关键词:水稻; 合成启动子; GUS报告基因; 顺式调控元件 Designing, Construction and Functional Characterization of Tissue-specific Synthetic Promoter in Rice WANG Rui1,2, ZHU Meng-Lin1, GAO Fang-Yuan2, REN Juan-Sheng2, LU Xian-Jun2, REN Guang-Jun2, LIN Yong-Jun1,* 1 National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University, Wuhan 430070, China
2Crop Research Institute, Sichuan Academy of Agricultural Sciences, Chengdu 610066, China
Fund:This study was supported by the National Major Project for Developing New GM Crops (2014ZX08001001) and the National High Technology Research and Development Program of China (863 Program). AbstractAs an important part of synthetic biology, synthetic promoter has gradually become a hotspot in current biology. Rice is one of the most important food crops in the world and a model plant for functional genomic research in cereals. The purpose of the present study was to design and construct tissue-specific synthetic promoters in rice. We selected several tissue-specific cis-elements based on previous reports and assembled them in different patterns. The combinations were respectively fused to Mini 35S promoter. GUS assays of the transgenic plants indicated that our work successfully created tissue-specific synthetic promoter with the above approach and also revealed that the assembly patterns of cis-elements are critical for the expression patterns and expression efficiencies of synthetic promoters. Our study provides useful information and a practical basis for the design ideas and construction approaches of synthetic promoters in plants.
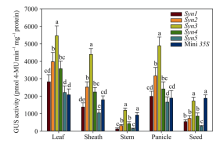
图3 不同合成启动子驱动GUS报告基因的转基因水稻各个组织的GUS活性定量检测Fig. 3 Quantitative analysis of GUS activity in various tissues of the transgenic plants containing different synthetic promoters/GUS fusions 柱上标以不同字母表示不同转基因植株之间以及转基因植株与对照之间存在显著差异(P< 0.05)。 Bars denoted by different letters are significantly different between different transgenic plants and also between the transgenic plants and the control at P< 0.05.
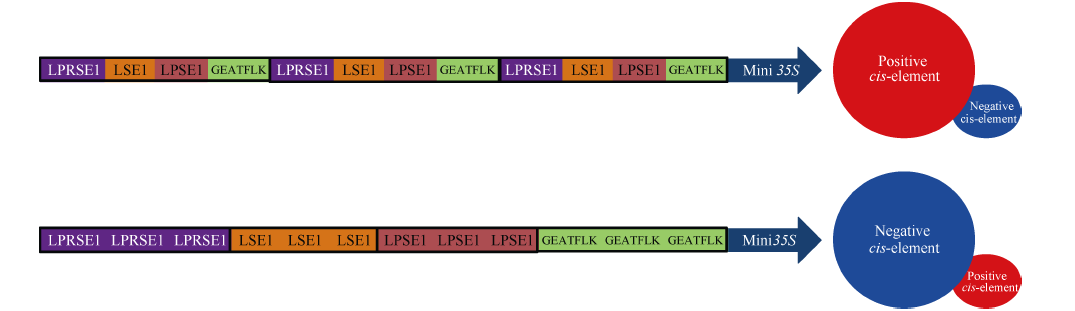
图4 多种顺式元件的不同组合方式对合成启动子表达活性影响的推测Fig. 4 Speculation of the influence of different assembly patterns of multiple cis-elements on the expression efficiencies of synthetic promoters
4 结论利用顺式调控元件连接核心启动子的方式构建出种子不表达的组织特异型合成启动子, 并推测出合成启动子中顺式调控元件的组合规律: 单拷贝顺式元件串联后以组合的形式重复多次的设计方式有利于正调控顺式元件发挥功能; 而顺式元件以多拷贝形式组合的设计方式则有利于负调控顺式元件发挥功能。该结果表明, 控制启动子的表达活性和表达模式的因素不仅来自于顺式调控元件自身的功能, 更重要的是顺式元件之间的位置关系和相互作用。 The authors have declared that no competing interests exist.
CaiM, WeiJ, LiX, XuC, WangS. A rice promoter containing both novel positive and negative cis-elements for regulation of green tissue-specific gene expression in transgenic plants. Plant Biotechnol J, 2007, 5: 664-674[本文引用:3]
[2]
扆珩, 李昂, 刘惠民, 景蕊莲. 小麦蛋白磷酸酶2A基因 TaPP2AbB’’-α启动子的克隆及表达分析. 作物学报, 2016, 42: 1282-1290YiH, LiA, Liu HM, Jing RL. Cloning and expression analysis of protein phosphatase 2A gene TaPP2AbB’’-α promoter in wheat. Acta Agron Sin, 2016, 42: 1282-1290 (in Chinese with English abstract)[本文引用:1]
[3]
习雨琳, 周朋, 宋梅芳, 李志勇, 孟凡华, 杨建平. 拟南芥 RBCS-1A基因受光调节表达模式及其启动子遗传转化应用评价. 作物学报, 2012, 38: 1561-1569Xi YL, ZhouP, Song MF, Li ZY, Meng FH, Yang JP. Expression patterns of Arabidopsis RBCS-1Agene in response to light treatments and application evaluation of its promoter in transgenic engineering. Acta Agron Sin, 2012, 38: 1561-1569 (in Chinese with English abstract)[本文引用:1]
[4]
Yim SS, An SJ, KangM, LeeJ, Jeong KJ. Isolation of fully synthetic promoters for high-level gene expression in Corynebacterium glutamicum. Biotechnol Bioeng, 2013, 110: 2959-2969[本文引用:1]
Sohoni SV, FazioA, Workman CT, MijakovicI, Lantz AE. Synthetic Promoter Library for modulation of actinorhodin production in Streptomyces coelicolor A3 (2). PLoS One, 2014, 9: e99701[本文引用:1]
[7]
DaiJ, ZhangQ, LiuS, ZhangM, RenX, HaoL, JiangQ, ZhangY. The combination of a synthetic promoter and a CMV promoter improves foreign gene expression efficiency in myocytes. J Biotechnol, 2012, 158: 91-96[本文引用:1]
[8]
KangW, KwonJ, KimH, JeongH, KangH, JeongM, ChoJ, ParkJ, KimY, AhnY. A macrophage-specific synthetic promoter for therapeutic application of adiponectin. Gene Ther, 2014, 21: 353-362[本文引用:1]
[9]
Rushton PJ, ReinstadlerA, LipkaV, LippokB, Somssich IE. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling. Plant Cell, 2002, 14: 749-762[本文引用:1]
[10]
RoychoudhuryA, SenguptaD. The promoter-elements of some abiotic stress-inducible genes from cereals interact with a nuclear protein from tobacco. Biol Plant, 2009, 53: 583-587[本文引用:1]
[11]
KoschmannJ, MachensF, BeckerM, NiemeyerJ, SchulzeJ, BülowL, Stahl DJ, HehlR. Integration of bioinformatics and synthetic promoters leads to the discovery of novel elicitor- responsive cis-regulatory sequences in Arabidopsis. Plant Physiol, 2012, 160: 178-191[本文引用:1]
[12]
LiuW, MazareiM, Rudis MR, Fethe MH, PengY, Millwood RJ, SchoeneG, Burris JN, Jr Stewart C N. Bacterial pathogen phytosensing in transgenic tobacco and Arabidopsis plants. Plant Biotechnol J, 2013, 11: 43-52[本文引用:1]
[13]
LiuW, MazareiM, PengY, Fethe MH, Rudis MR, LinJ, Millwood RJ, Arelli PR, Jr Stewart C N. Computational discovery of soybean promoter cis-regulatory elements for the construction of soybean cyst nematode-inducible synthetic promoters. Plant Biotechnol J, 2014, 12: 1015-1026[本文引用:1]
[14]
Goff SA, RickeD, Lan TH, PrestingG, WangR, DunnM, GlazebrookJ, SessionsA, OellerP, VarmaH. A draft sequence of the rice genome ( Oryza sativa L. ssp. japonica). Science, 2002, 296: 92-100[本文引用:1]
[15]
PanY, DengY, LinH, Kudrna DA, Wing RA, LiL, ZhangQ, LuoM. Comparative BAC-based physical mapping of Oryza sativassp. indica var. 93-11 and evaluation of the two rice reference sequence assemblies. Plant J, 2014, 77: 795-805[本文引用:1]
[16]
YuJ, HuS, WangJ, Wong GK, LiS, LiuB, DengY, DaiL, ZhouY, ZhangX. A draft sequence of the rice genome ( Oryza sativa L. ssp. indica). Science, 2002, 296: 79-92[本文引用:1]
[17]
WangL, XieW, ChenY, TangW, YangJ, YeR, LiuL, LinY, XuC, XiaoJ, ZhangQ. A dynamic gene expression atlas covering the entire life cycle of rice. Plant J, 2010, 61: 752-766[本文引用:1]
[18]
YeR, ZhouF, LinY. Two novel positive cis-regulatory elements involved in green tissue-specific promoter activity in rice ( Oryza sativa L. ssp. ). Plant Cell Rep, 2012, 31: 1159-1172[本文引用:2]
[19]
WangR, ZhuM, YeR, LiuZ, ZhouF, ChenH, LinY. Novel green tissue-specific synthetic promoters and cis-regulatory elements in rice. Sci Rep, 2015, 5: 18256[本文引用:2]
[20]
HieiY, OhtaS, KomariT, KumashiroT. Efficient transformation of rice ( Oryza sativaL. ) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J, 1994, 6: 271-282[本文引用:1]
[21]
WuC, LiX, YuanW, ChenG, KilianA, LiJ, XuC, LiX, Zhou DX, WangS, ZhangQ. Development of enhancer trap lines for functional analysis of the rice genome. Plant J, 2003, 35: 418-427[本文引用:1]
[22]
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976, 72: 248-254[本文引用:1]
[23]
XuL, YeR, ZhengY, WangZ, ZhouP, LinY, LiD. Isolation of the endosperm-specific LPAAT gene promoter from coconut( Cocos nuciferaL. ) and its functional analysis in transgenic rice plants. Plant Cell Rep, 2010, 29: 1061-1068[本文引用:1]
[24]
LiC, WangY, LiuL, HuY, ZhangF, MergenS, WangG, Schläppi MR, ChuC. A rice plastidial nucleotide sugar epimerase is involved in galactolipid biosynthesis and improves photosynthetic efficiency. PLoS Genet, 2011, 7: e1002196[本文引用:1]
[25]
ZhouY, HuangW, LiuL, ChenT, ZhouF, LinY. Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence. BMC Plant Biol, 2013, 13: 132[本文引用:1]
[26]
ZhuL, HuJ, ZhuK, FangY, GaoZ, HeY, ZhangG, GuoL, ZengD, DongG, YanM, LiuJ, QianQ. Identification and characterization of SHORTENED UPPERMOST INTERNODE 1, a gene negatively regulating uppermost internode elongation in rice. Plant Mol Biol, 2011, 77: 475-487[本文引用:1]
[27]
HuaL, WuJ, ChenC, WuW, HeX, LinF, WangL, AshikawaI, MatsumotoT, WangL, PanQ. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Theor Appl Genet, 2012, 125: 1047-1055[本文引用:1]
[28]
YangZ, ChenH, TangW, HuaH, LinY. Development and characterisation of transgenic rice expressing two Bacillus thuringiensisgenes. Pest Manag Sci, 2011, 67: 414-422[本文引用:1]
 , 朱梦琳
, 朱梦琳





{kind=link}
{kind=link}
{kind=link}
{kind=link}