关键词:甘蓝型油菜; 脯氨酸合成; 多倍化; 偏向性表达; 亚功能化 Evolutionary Fate and Expression Pattern of Genes Related to Proline Biosynthesis in Brassica napus WANG Cui-Ping1, HUA Xue-Jun2,*, LIN Bin3, LIU Ai-Hua4 1 State Key Laboratory of Seedling Bioengineering, Ningxia Forestry Institute, Yinchuan 750004, China
2 Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
3 University of Chinese Academy of Sciences, Beijing 100049, China
4 Department of Plant Science, University of Manitoba, Winnipeg, R3T2N2, MB, Canada
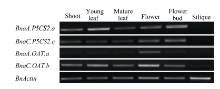
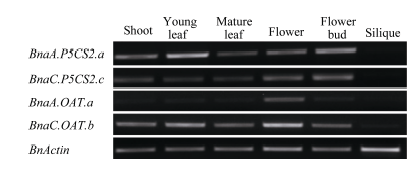
Fund:The study was supported by the Ningxia Natural Science Foundation of China (NZ16215). AbstractProline accumulation is a widespread metabolic adaptation in many organisms in response to various environmental stresses, and it was proved to play protective roles for plants under adverse conditions. Polyploidization is a prominent driving force during plant evolution and many important crops have experienced this process during their evolutionary history. Brassica napus (AACC) is believed to be a newly formed allotetraploid, evolving from the inter-specific hybridization of two diploids, B. rapa (AA) and B. oleracea (CC) followed by chromosome doubling. In this research, we studied the evolutionary fate of genes related to proline synthesis in allotetraploid ( B. napus), and its diploids ancestors ( B. rapa and B. oleracea), to explore the effect of polyploidition on homologous gene evolution. First, we obtained the genes for proline biosynthesis ( P5CSs, OAT) by database searching, and studied the similarities and the expression regulation pattern of homologous genes in allotetraploid ( B. napus), in comparison with its diploids progenitors ( B. rapa and B. oleracea), in different organs and in response to salt stress. Sequence analysis and phylogenetic analysis revealed that BnaA.P5CS2.a, BnaA.P5CS2.b, and BnaC.P5CS2.c originated from BraA.P5CS2.a, BraA.P5CS2.b, and BolC.P5CS2.a, respectively; BnaA.OAT.a, BnaC.OAT.b originated from BraA.OAT.a and BolC.OAT.a, respectively. And, one copy of gene loss from B. oleracea occurred for BnP5CS2but not for BnOAT. Expression patterns of these homologous genes in response to salt stress in different organs were also characterized by semi-quantitive RT-PCR. In B. napus, two homologous gene pairs with different origins, BnaA.P5CS2.a and BnaC.P5CS2.c, BnaA.OAT.a and BnaC.OAT.b exhibited biased expression in different organs, implying possible sub-functionalization of P5CS2 and OAT. The genes BnaA.P5CS1.aand BnaC.P5CS1.dwith different diploid ancestors were induced by NaCl treatment, and the expression of BnaC.P5CS1.d was higher than that of BnaA.P5CS1.a, showing a biased expression. RT-PCR manifested that preservation of expression pattern of original genes in diploid was found for P5CS1( BnaA.P5CS1.aand BnaC.P5CS1.d), P5CS2( BnaA.P5CS2.a and BnaC.P5CS2.c). These results suggest that the gene sequence and expression pattern existing in allotetraploid ( B. napus) were conserved, which is benefit to proline accumulation for plant adaptation to environmental stresses.
Keyword:Polyploidization; Proline biosynthesis; Biased expression; Salt stress; Sub-functionalization Show Figures Show Figures
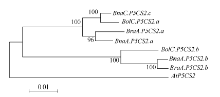
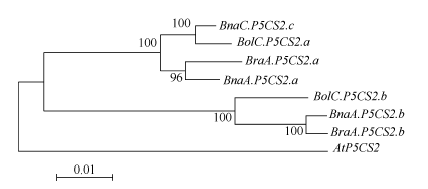
图2 芸薹属植物P5CS2基因进化关系分析 通过软件MEGA4的邻位相连法构建进化树, 所用数据包括P5CS2基因的基因组DNA序列(从起始密码子到终止密码子)。Fig. 2 Phylogenetic analysis of Brassica P5CS2 genes The phylogenetic tree was constructed using neighbor-joining method of MEGA4. The dataset covered partial genomic sequences of P5CS2 genes. The tree was drawn to scale, and the lengths of tree branches had the same units as evolutionary distances. Percentages refer to significant bootstrap value of 1000 calculated trees (from start codon to stop codon).
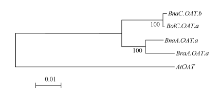
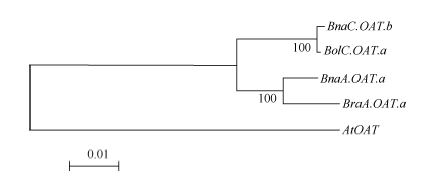
图3 芸薹属植物OAT基因进化关系分析 通过软件MEGA4的邻位相连法构建进化树, 所用数据包括OAT基因基因组DNA序列(从起始密码子到终止密码子)。Fig. 3 Phylogenetic analysis of Brassica OATgenes The phylogenetic tree was constructed using neighbor-joining method of MEGA4. The dataset covered partial genomic sequences of OAT genes. The tree was drawn to scale, and the lengths of tree branches had the same units as evolutionary distances. Percentages refer to significant bootstrap value of 1000 calculated trees (from start codon to stop codon).
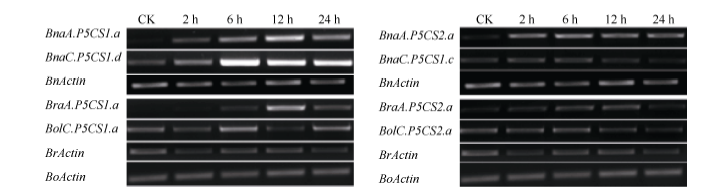
图5 250 mmol L-1 NaCl胁迫下甘蓝型油菜、白菜和甘蓝的P5CS1和P5CS2的表达模式Fig. 5 Expression pattern of P5CS1 and P5CS2 in B. napus, B. rapa, and B. oleracea under 250 mmol L-1 NaCl
4 结论甘蓝型油菜中脯氨酸代谢途径基因(P5CS和OAT)存在多个拷贝, 它们和其二倍体亲本白菜及甘蓝的对应基因序列高度相似; 和亲本相比, 甘蓝型油菜的P5CS2基因来自二倍体祖先甘蓝的一个拷贝丢失, 而OAT基因没有发生丢失; 盐胁迫下, 甘蓝型油菜P5CS1和P5CS2基因的诱导模式都和亲本中的祖先基因诱导模式一致, 说明甘蓝型油菜进化中该基因盐诱导模式的保守性。 致谢: 感谢Calgary大学的Maurice MOLONEY博士, Ed YEUNG博士和唐明娟博士提供甘蓝型油菜(B. napus L. cv. Westar)种子, 感谢中国农业科学院蔬菜花卉研究所王晓武研究员提供白菜和甘蓝的P5CS基因和OAT基因的相关序列信息。 The authors have declared that no competing interests exist.
Kishor P BK, Hong ZL, Miao GH, Hu C AA, Verma D PS . Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants . Plant Physiol, 1995, 108: 1387-1394[本文引用:1]
[2]
SchatH, Sharma SS, Vooijs R . Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol Plant, 1997, 101: 477-482[本文引用:1]
[3]
Yang SL, Lan SS, GongM. Hydrogen peroxide-induced proline and metabolic pathway of its accumulation in maize seedlings . J Plant Physiol, 2009, 166: 1694-1699[本文引用:1]
[4]
TavakoliM, PoustiniK, Alizadeh H . Proline accumulation and related genes in wheat leaves under salinity stress . J Agric Sci Tech, 2016, 18: 707-716[本文引用:3]
[5]
Dar MI, Naikoo MI, RehmanF, NaushinF, Khan FA. Proline accumulation in plants: roles in stress tolerance and plant development. In: Iqbal N, Nazar R, Khan N A, eds. Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies. New Delhi: Springer India Press, 2016. pp 155-166[本文引用:1]
[6]
Chaves MM, FlexasJ, Pinheiro C . Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell . Ann Bot, 2009, 103: 551-560[本文引用:1]
[7]
Wu HH, Zou YN, Rahman MM, Ni QD, Wu Q S . Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress . Sci Rep, 2017, 7: 42389[本文引用:1]
[8]
MattioliR, MarcheseD, D’Angeli S, Altamura M M, Costantino P, Trovato M . Modulation of intracellular proline levels affects flowering time and inflorescence architecture in Arabidopsis. Plant Mol Biol, 2008, 66: 277-288[本文引用:1]
[9]
MattioliR, CostantinoP, Trovato M . Proline accumulation in plants: not only stress . Plant Signal Behav, 2009, 4: 1016-1018[本文引用:1]
[10]
MattioliR, FalascaG, SabatiniS, Altamura MM, CostantinoP, Trovato M . The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development . Physiol Plant, 2009, 137: 72-85[本文引用:1]
[11]
FunckD, WinterG, BaumgartenL, Forlani G . Requirement of proline synthesis during Arabidopsis reproductive development . BMC Plant Biol, 2012, 12: 191[本文引用:1]
[12]
Delauney AJ, Hu C A A, Kishor P B K, Verma D P S . Cloning of ornithine delta-aminotransferase cDNA from Vigna-Aconitifolia by transcomplementation in Escherichia coliand regulation of proline biosynthesis . J Biol Chem, 1993, 268: 18673-18678[本文引用:1]
[13]
Kishor P BK, SangamS, Amrutha RN, Laxmi PS, Naidu KR, Rao K R S S, RaoS, Reddy KJ, TheriappanP, SreenivasuluN . Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance . Curr Sci India, 2005, 88: 424-438[本文引用:1]
[14]
WangL, GuoZ, ZhangY, WangY, YangG, YangL, WangR, Xie Z . Characterization of LhSorP5CS, a gene catalyzing proline synthesis in Oriental hybrid lily Sorbonne: molecular modelling and expression analysis . Bot Stud, 2017, 58(1): 10[本文引用:1]
[15]
YoshibaY, KiyosueT, KatagiriT, UedaH, MizoguchiT, YamaguchishinozakiK, WadaK, HaradaY, Shinozaki K . Correlation between the induction of a gene for Δ1-pyrroline-5-carboxylate synthetase and the accumulation of proline in Arabidopsis thaliana under osmotic-stress . Plant J, 1995, 7: 751-760[本文引用:1]
[16]
FabroG, KovacsI, PavetV, SzabadosL, Alvarez M E . Proline accumulation and AtP5CS2 gene activation are induced by plant-pathogen incompatible interactions in Arabidopsis. Mol Plant Microbe In, 2004, 17: 343-350[本文引用:1]
[17]
Soltis PS, Soltis D E . The role of genetic and genomic attributes in the success of polyploids . Proc Nati Acad Sci USA, 2000, 97: 7051-7057[本文引用:1]
[18]
Buggs RJ, Doust AN, Tate JA, KohJ, SoltisK, Feltus FA, Paterson AH, Soltis PS, Soltis D E . Gene loss and silencing in Tragopogon miscellus(Asteraceae): comparison of natural and synthetic allotetraploids . Heredity, 2009, 103: 73-81[本文引用:1]
[19]
WangJ, TianL, Lee HS, Chen Z J . Nonadditive regulation of FRIand FLC loci mediates flowering-time variation in Arabidopsisallopolyploids . Genetics, 2006, 173: 965-974[本文引用:2]
[20]
Zhao JW, BuchwaldtL, Rimmer SR, BrkicM, BekkaouiD, Hegedus D . Differential expression of duplicated peroxidase genes in the allotetraploid Brassica napus. Plant Physiol Bioch, 2009, 47: 653-656[本文引用:3]
[21]
NagaharaU . Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilisation . J Jpn Bon, 1935, 7: 389-452[本文引用:1]
[22]
BlancG, Wolfe K H . Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution . Plant Cell, 2004, 16: 1679-1691[本文引用:2]
[23]
Yang YW, Lai KN, Tai PY, Li W H . Rates of nucleotide substitution in angiosperm mitochondrial DNA sequences and dates of divergence between Brassica and other angiosperm lineages . J Mol Evol, 1999, 48: 597-604[本文引用:1]
[24]
Park JY, Koo DH, Hong CP, Lee SJ, Jeon JW, Lee SH, Yun PY, Park BS, Kim HR, Bang JW, PlahaP, BancroftI, Lim Y P . Physical mapping and microsynteny of Brassica rapa ssp. pekinensis genome corresponding to a 222 kbp gene-rich region of Arabidopsis chromosome 4 and partially duplicated on chromosome 5 . Mol Genet Genom, 2005, 274: 579-588[本文引用:1]
Udall JA, Wendel J F . Polyploidy and crop improvement . Crop Sci, 2006, 46: S3-S14[本文引用:2]
[27]
Wang CP, LinB, Zhang YQ, Lin YH, Liu AH, Hua X J . The evolutionary fate of Δ1-pyrroline-5-carboxylate synthetase 1( P5CS1) genes in allotetraploid Brassica napus. J Syst Evol, 2014, 52: 566-579[本文引用:2]
Hua XJ, van de CotteB, Van MontaguM, VerbruggenN . The 5 ' untranslated region of the At-P5R gene is involved in both transcriptional and post-transcriptional regulation . Plant J, 2001, 26: 157-169[本文引用:1]
[30]
ØstergaardL, King G J . Stand ardized gene nomenclature for the Brassicagenus . Plant Methods, 2008, 4: 10[本文引用:1]
[31]
HuaS, Shamsi IH, GuoY, PakH, ChenM, ShiC, MengH, Jiang L . Sequence, expression divergence, and complementation of homologous ALCATRAZ loci in Brassica napus. Planta, 2009, 230: 493-503[本文引用:1]
[32]
Cardenas PD, Gajardo HA, HuebertT, Parkin IA, Iniguez-Luy F L, Federico M L . Retention of triplicated phytoene synthase( PSY) genes in Brassica napus L. and its diploid progenitors during the evolution of the Brassiceae. Theor Appl Genet, 2012, 124: 1215-1228[本文引用:2]
[33]
DengW, ZhouL, Zhou YT, Wang YJ, Wang ML, Zhao Y . Isolation and characterization of three duplicated PISTILLATAgenes in Brassica napus. Mol Biol Rep, 2011, 38: 3113-3120[本文引用:1]
[34]
ForceA, LynchM, Pickett FB, AmoresA, Yan YL, Postlethwait J . Preservation of duplicate genes by complementary, degenerative mutations . Genetics, 1999, 151: 1531-1545[本文引用:1]
[35]
Adams KL, Liu Z L . Expression partitioning between genes duplicated by polyploidy under abiotic stress and during organ development . Curr Biol, 2007, 17: 1669-1674[本文引用:2]
 , 华学军
, 华学军



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}