关键词:玉米; SNAC基因; 遗传变异; 耐旱性 Genetic Variations and Drought Tolerance of SNAC Genes in Common Maize Inbred Lines of China LI Guo-Jun1,3,**, MA Yi-Wen2,3,**, XU Dan-Yang3, WU Yong-Bo3, SONG Jie3, WANG Nan3, HAO Zhuan-Fang3,*, ZHAO Juan1,* 1 College of Agronomy, Shanxi Agricultural University, Taigu 030801, China
2 Tonghua Academy of Agricultural Sciences, Tonghua 135007, China
3 Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
Fund:This study was supported by the National Natural Science Foundation of China (31661143010, 31271735) AbstractThe coding regions and their upstream 800 bp promoter regions of SNAC genes ( Sress-responsive NAM, ATAF1/2, CUC2) were sequenced in 16 maize inbred lines commonly used in China. Among 12 SNAC genes, genetic variations in promoter region were only identified in four SNAC genes, and more than 30 variations were identified in four SNAC genes, showing higher polymorphism in the four genes than other in SNAC genes. Although most of the SNAC genes were mainly SNP (Single nucleotide polymorphism) mutations, more insertion/deletion mutations were detected in ZmNAC031467 gene, reaching 63.3% of the total genetic variations. The PLACE software was used to predict three kinds of stress-tolerant binding elements in SNAC gene, but little effect was found to be related with the variations. Additionally, high nucleotide polymorphisms were identified in seven SNAC genes, especially with the highest π value of 0.00962 in ZmNAC030308, which suggested that they were greatly influenced by natural selection in the genetic drift. With the t-test, two mutations of ZmNAC070395 and ZmNAC080398 genes were associated with drought-tolerant traits, which provides references for further analysing the relationship between nucleotide variation in SNAC and drought tolerance traits.
Keyword: Zea mays L.; SNAC genes; Genetic variations; Drought tolerance Show Figures Show Figures
表1 研究用玉米自交系的基本情况 Table 1 Basic information of tested maize inbred lines
自交系 Inbred line
类群a Heterotic group
系谱 Pedigree
耐旱性(SI)b Drought tolerance
齐318 Qi 318
PBc
源自78599杂交系 Derived from hybrid 78599
-2.05
齐319 Qi 319
PB
源自78599杂交系 Derived from hybrid 78599
1.13
X178
PB
源自78599杂交系 Derived from hybrid 78599
1.48
豫12 Yu 12
LAN (Lancaster)
Mo17改良系 Improved from Mo17
1.52
吉412 Ji 412
LAN (Lancaster)
源自Mo17 Derived from Mo17
-0.01
Mo17
LAN (Lancaster)
C103× 187-2
0.97
CN962
唐四平头 SPT
黄早四改良系 Improved from Huangzaosi
-1.51
黄早四 Huangzaosi
唐四平头 SPT
源自唐四平头 Derived from Tangsipingtou
0.35
昌7-2 Chang 7-2
唐四平头 SPT
(黄早四× Wei95)× S901 (Huangzaosi× Wei 95)× S901
0.18
铁7922 Tie 7922
Reid
源自3382杂交系 Derived from hybrid 3382
2.26
掖478 Ye 478
Reid
U8112× 沈5003 U8112× Shen 5003
0.25
郑58 Zheng 58
Reid
掖478改良系 Improved from Ye 478
-0.52
B73
Reid
BSSSC5
0.96
CA339
旅大红骨 LRC
Pool 33 优质蛋白玉米 Pool 33 QPM
-0.90
丹340 Dan 340
旅大红骨 LRC
Baigulv 9× Lemma corn Co60
-0.26
吉81162 Ji 81162
旅大红骨 LRC
525× 掖107 525× Ye 107
-1.82
aThe division of heterotic breeding groups for maize inbred lines is shown by Liu et al.[15] in detail in 2015; bThe assessment of drought tolerance is based on the maize selection index (SI) by Hao et al.[16] in 2011; cPB is described by Zhang S H[17] in 2014. SPT: Tangsipingtou; LRC: Lyuda Red Cob. a玉米自交系杂种优势群的划分详见Liu等[15], 2015; b耐旱性的评估以玉米选择指数(SI)判断为准, 详见Hao等[16], 2011; c详见张世煌[17], 2014。
表1 研究用玉米自交系的基本情况 Table 1 Basic information of tested maize inbred lines
表2 扩增12个SNAC基因序列所用引物信息 Table 2 Basic information of primers for 12 SNAC genes
基因名称a SNAC genea
引物名称b Primerb
引物序列 Primer sequence (5° -3° )
长度 Primer length (bp)
退火温度 Annealing tempreture (℃)
ZmNAC010312
GRMZM2G347043-1F
AGCACGATAATGAACCCTC
19
53
GRMZM2G347043-1R
TGTACGCTACACCGACACT
19
53
ZmNAC010373
GRMZM2G011598-1F
GACTCGTTGGTGGAGGTGG
19
56
GRMZM2G011598-1R
AAGTACAAATCCGTCAATCCC
21
56
ZmNAC020303
GRMZM2G162739-1F
TGTCGGCTGCGGTTGAGA
18
58
GRMZM2G162739-1R
GGCAGAGCACCCAGTCAT
18
58
GRMZM2G162739-2F
GGTCAAGACGGAGTGGAT
18
55
GRMZM2G162739-2R
TTGCGATGACTAAACGAAG
19
55
ZmNAC030295
GRMZM2G014653-1F
AGTTACGGGTGGGTTCAT
18
54
GRMZM2G014653-1R
ATTGGTTCACGGCTGCTA
18
54
GRMZM2G014653-2F
ACCTCTACAAGTTCGACCCA
20
56
GRMZM2G014653-2R
AGTCAACAATGGCGGCTC
18
56
ZmNAC031467
GRMZM2G312201-1F
AGCGAGCAAACCATCTAA
18
53
GRMZM2G312201-1R
TAAAGGGAACATAGCAACAC
20
53
GRMZM2G312201-2F
TGGTCGCAAGGAGTGGTA
18
56
GRMZM2G312201-2R
TGGTGGGTGACGGAGTGAGG
20
56
GRMZM2G312201-3F
GCCCACTTGTACGATTGA
18
54
GRMZM2G312201-3R
ACGCTAACCCTTGCTCTA
18
54
GRMZM2G312201-4F
CAGTTGAGGCACTTGAGC
18
55
GRMZM2G312201-4R
TCTTCAGCCTGCATTAGC
18
55
ZmNAC040359
GRMZM2G123667-1F
AAATTGACGCTTGGGAAGA
19
54
GRMZM2G123667-1R
GGAAGAGGCTTGGAGATGA
19
54
GRMZM2G123667-2F
CATCTCCAAGCCTCTTCC
18
56
GRMZM2G123667-2R
CGAGCACTCCTGGTTTCT
18
56
ZmNAC050302
GRMZM2G336533-1F
TGGCTGATGCGGTGTTTG
18
55
GRMZM2G336533-1R
GGTCCCTGGGCGTGAAGA
18
55
GRMZM2G336533-2F
CACCGCTTCTAGTTCTCG
18
55
GRMZM2G336533-2R
CTTCAAACATTGCCATTCG
19
55
ZmNAC070395
GRMZM2G430849-1F
ACGATTGTTGATTGCTCCAT
20
56
GRMZM2G430849-1R
CGCACGACCCATCTTTATT
19
56
ZmNAC080308
GRMZM2G068973-1F
GTGATCGCATTGATACCG
18
54
GRMZM2G068973-1R
GTCTGCCACATCTTCGTC
18
54
ZmNAC080398
GRMZM2G109627-1F
CCTTCGCTTGAATCTCCC
18
55
GRMZM2G109627-1R
GCAGCGGTATTCTTTCCT
18
55
ZmNAC090241
GRMZM2G042494-1F
GCTAGTTATTGTTGTCCTCCTG
22
54
GRMZM2G042494-1R
AAGTACCACTCCTTGTCCC
19
54
GRMZM2G042494-2F
GACTCAGCTCGCATCACA
18
56
GRMZM2G042494-2R
ACCTCCTCGAACAAGTGC
18
56
ZmNAC100475
GRMZM2G127379-1F
ATGATTCTAAGCCTGGTATGTC
22
58
GRMZM2G127379-1R
GACGCCGACGACGATGTT
18
58
GRMZM2G127379-2F
AGCCAAGGCATTGTTCGG
18
56
GRMZM2G127379-2F
CGTGCATGGGTCGTCATC
18
56
aThe name of maize SNAC transcription factors was described in detail by Li et al.[19]in 2015, with the name of chromosome and the amino acid fragment size each; bThe name of primers is followed the gene name in the NCBI database. a玉米SNAC基因的名称编号详见Li等19], 2015, 以ZmNAC加基因所在染色体再加氨基酸片段大小命名。b玉米SNAC基因引物以基因所在NCBI数据库中的名称为主进行命名。
表2 扩增12个SNAC基因序列所用引物信息 Table 2 Basic information of primers for 12 SNAC genes
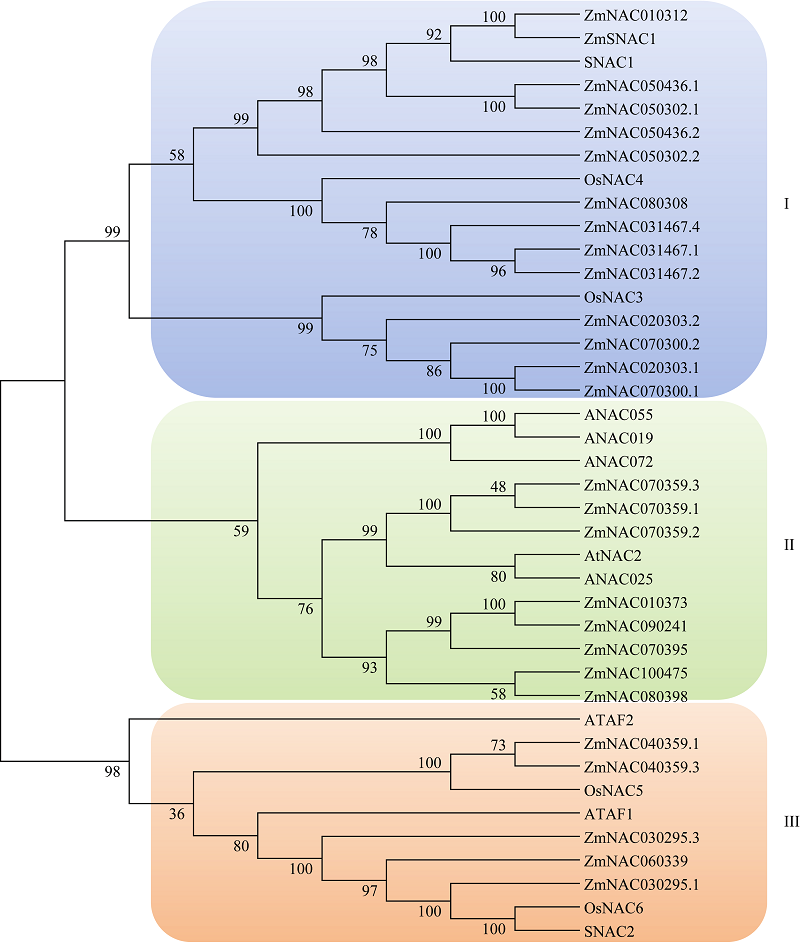
图1 玉米SNAC家族和已知的拟南芥、水稻SNAC蛋白的聚类分析 图中SNAC转录因子及拟南芥和水稻SNAC蛋白序列见Li等, 2015[19]。Fig. 1 Phylogenetic tree among the SNAC proteins of maize and NAC proteins of Arabidopsisand rice Protein sequences of SNAC TF from maize, Arabidopsisand rice were described by Li et al. in 2015[19].
表3 核苷酸多样性与中性检测 Table 3 Nucleotide diversity and neutrality test
基因名称 Gene
Tajima’ s D
核苷酸多样性(Pi/π ) Nucleotide diversity
ZmNAC010312
-1.08744 NS
0.00217
ZmNAC010373
-1.78041 NS
0.00062
ZmNAC020303
-2.21101* *
0.00051
ZmNAC031467
-2.29433* *
0.00068
ZmNAC030295
-0.78442 NS
0.00269
ZmNAC040359
-0.37821 NS
0.00085
ZmNAC050302
2.14811*
0.00206
ZmNAC070395
0.70217 NS
0.00189
ZmNAC080398
0.93093 NS
0.00360
ZmNAC080308
1.54676 NS
0.00962
ZmNAC090241
0.69718 NS
0.00230
ZmNAC100475
-0.90066 NS
0.00201
* * The gene is subjected to negative selection pressure; * The gene is subjected to positive selection pressure; NS means nonsignificance, the gene is not subjected to selection pressure. * * 为基因受负向选择压力; * 为基因受正向选择压力; NS表示不显著, 为基因不受选择压力。
表3 核苷酸多样性与中性检测 Table 3 Nucleotide diversity and neutrality test
Tajima’ s D检验是鉴定目标DNA序列在进化过程中是否遵循中性进化模型。如果是负的显著结果, 那么这个结果可能是负选择造成的, 反之则受到平衡选择[22]。本研究利用有限的群体针对自然选择是否影响SNAC亚族基因初步分析, 通过使用Tajima’ s D值来检测群体是否符合中性进化模型(表3), 其中大多数SNAC基因差异不显著, 说明这9个基因在群体中未偏离中性进化, 基因突变受自然选择影响较小; ZmNAC020303、ZmNAC031467基因的Tajima’ s D值分别为-2.21101、-2.29433, 达到极显著水平, 说明这2个基因受负向选择压力较大, 突变受自然选择影响较大; 同时ZmNAC050302基因Tajima’ s D为2.14811, 达极显著水平, 说明这个基因受正向选择压力较大, 基因突变受自然选择影响较大。 2.5 SNAC基因的等位变异与耐旱选择指数的t检验结果 t检验适用于总体标准差未知且样本容量小于30的小群体范围。本实验利用t检验, 对12个SNAC基因的393个多态性位点与耐旱选择指数(表4)进行差异显著性分析。结果在P-value < 0.01水平上检测到2个突变位点差异显著, 分别是位于第7染色体上的ZmNAC070395基因的553 bp位点和第8染色体上的ZmNAC080398基因的226 bp位点。初步推测这2个基因变异位点可能对耐旱性有着一定的影响, 需要进一步的验证(表4)。 表4 Table 4 表4(Table 4)
表4 SNAC基因多态性位点与选择指数差异显著性分析 Table 4 Significant polymorphic variations in SNAC gene associated with drought-tolerance selection index
基因名称 Gene
位点 Site
内含子/外显子 Intron/exon
显著性 P-value
ZmNAC070395
553
外显子2 Exon 2
0.0051* *
ZmNAC080398
226
内含子1 Intron 1
0.0079* *
* * Significant diffence at the 0.01 probability level.
表4 SNAC基因多态性位点与选择指数差异显著性分析 Table 4 Significant polymorphic variations in SNAC gene associated with drought-tolerance selection index
4 结论12个玉米SNAC基因与拟南芥及水稻中已知的SNAC基因具有相似的功能基序, 推测参与类似的耐逆功能代谢途径。4个玉米SNAC基因在上游800 bp检测到遗传变异, 序列含有3种逆境相关的顺式作用元件。其中有7个SNAC基因核苷酸多态性较高, ZmNAC080308基因的多态性达到0.00962, 说明这些基因在遗传漂移过程中受自然选择影响较大。ZmNAC070395的553 bp位点和ZmNAC080398的226 bp位点与耐旱选择指数极显著相关, 推测这两个核苷酸多态性位点对玉米耐旱性有着一定影响, 可以被考虑作为耐旱标记辅助选择。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
李月. 棉花逆境胁迫应答Trihelix转录因子的鉴定及功能分析. 石河子大学博士学位论文, 新疆石河子, 2013LiY. The Identification of Trihelix Transcription Factor and Functional Analysis in Response to Stress in Cotton. PhD Dissertation of Shihezi University, Shihezi, China, 2013 (in Chinese with English abstract)[本文引用:1]
[2]
陈儒钢, 巩振辉, 逯明辉, 李大伟, 黄炜. 植物抗逆反应中的转录因子网络研究进展. 农业生物技术学报, 2010, 18(1): 126-134Chen RG, Gong ZH, Lu MH, Li DW, HuangW. Research advanced of the transcription factors networks related to plant adverse environmental stress. J Agric Biotechnol, 2010, 18(1): 126-134 (in Chinese with English abstract)[本文引用:1]
[3]
康桂娟, 曾日中, 聂智毅, 黎瑜, 代龙军, 段翠芳. 植物NAC 转录因子的研究进展. 生物技术通报, 2012, (11): 21-26Kang GJ, Zeng RZ, Nie ZY, LiY, Dai LJ, Duan CF. Research progress of plant NAC transcription factors. Biotechnol Bull, 2012, 11: 21-26 (in Chinese with English abstract)[本文引用:1]
[4]
SouerE, Houwelingen VA, KloosD, MolJ, KoesR. The no apical meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordial boundaries. Cell, 1996, 85: 159-170[本文引用:1]
[5]
AidaM, IshidaT, FukakiH, FujisawaH, TasakaM. Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell, 1997, 9: 841-857[本文引用:1]
[6]
Tran LS, NakashimaK, SakumaY, Simpson SD, FujitaY, MaruyamaK, FujitaM, SekiM, ShinozakiK, Yamaguchi-ShinozakiK. Isolation and functional analysis of Arabidopsis stress- inducible NAC transcription factors that bind to a drought- responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell, 2004, 16: 2481-2498[本文引用:1]
[7]
MitsudaN, HisaboriT, TakeyasuK, Sato MH. VOZ; isolation and characterization of novel vascular plant transcription factors with a one-zinc finger from Arabidopsis thaliana. Plant Cell Physiol, 2004, 45: 845-854[本文引用:1]
[8]
Fang YJ, YouJ, Xie KB, Xie WB, Xiong LZ. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol Genet Genomics, 2008, 280: 535-546[本文引用:1]
[9]
ShenH, Yin YB, ChenF, XuY, Dixon RA. A bioinformatic analysis of NAC genes for plant cell wall development in relation to lignocellulosic bioenergy production. BioEnergy Res, 2009, 2: 217-232[本文引用:1]
[10]
NuruzzamanM, ManimekalaiR, Sharoni AM, SatohK, KondohH, OokaH, KikuchiS. Genome-wide analysis of NAC transcription factor family in rice. Gene, 2010, 465: 30-44[本文引用:1]
LuM, YingS, Zhang DF, Shi YS, Song YC, Wang TY, LiY. A maize stress-responsive NAC transcription factor, ZmSNAC1, confers enhanced tolerance to dehydration in transgenic Arabidopsis. Plant Cell Rep, 2012, 31: 1701-1711[本文引用:2]
[14]
Mao HD, Wang HW, Liu SX, Li ZG, Yang XH, Yan JB, Li JS, Phan Tran L S, Qin F. A transposable element in a NAC gene is associated with drought tolerance in maize seedlings. Nat Commun, 2015, 6: 8326[本文引用:3]
[15]
Liu SS, Hao ZF, Weng JF, Li MS, Zhang DG, Pan TG, Zhang SH, Li XH. Identification of two functional markers associated with drought resistance in maize. Mol Breed, 2015, 35: 1-10[本文引用:1]
[16]
Hao ZF, Li XH, Su ZJ, Xie CX, Li MS, Liang XL, Weng JF, Zhang DG, LiL, Zhang SH. A proposed selection criterion for drought resistance across multiple environments in maize. Breed Sci, 2011, 61: 101-108[本文引用:1]
[17]
张世煌. 商业育种只需要两个杂种优势群. 种子科技, 2014, (7): 7-8Zhang SH. Only need two heterotic groups in commercial breeding. Seed Sci Technol, 2014, (7): 7-8 (in Chinese)[本文引用:1]
[18]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4326[本文引用:1]
[19]
LiL, Ma YW, Zhang SH, Hao ZF, Li XH. Zea mays NAC transcription factor family members: their genomic characteristics and relationship with drought stress. Res J Biotechnol, 2015, 1: 63-77[本文引用:2]
[20]
ZhaoY, Zhou YQ, Jiang HY, Li XY, Gan DF, Peng XJ, Zhu SW, Cheng BJ. Systematic analysis of sequences and expression patterns of drought-responsive members of the HD-Zip gene family in maize. PLoS One, 2011, 6: e28488[本文引用:1]
[21]
NeiM, Li WH. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci USA, 1979, 10: 5269-5273[本文引用:1]
[22]
周琦, 王文. DNA水平自然选择作用的检测. 动物学研究, 2004, 25(1): 73-80ZhouQ, WangW. Detecting natural selection at the DNA level. Zool Res, 2004, 25(1): 73-80 (in Chinese with English abstract)[本文引用:1]
[23]
李伟, 韩蕾, 钱永强, 孙振元. 植物NAC转录因子的种类、特征及功能. 应用与环境生物学报, 2011, 17: 596-606LiW, HanL, Qian YQ, Sun ZY. Characteristics and functions of NAC transcription factors in plants. Chin J Appl Environ Biol, 2011, 17: 596-606 (in Chinese with English abstract)[本文引用:1]
[24]
Hu HH, Dai MQ, Yao JL, Xiao BZ, Li XH, Zhang QF, Xiong LZ. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc Natl Acad Sci USA, 2006, 103: 12987-12992[本文引用:1]
[25]
KanedaT, TagaY, TakaiR, IwanoM, MatsuiH, TakayamaS, IsogaiA, CheF. The transcription factor OsNAC4 is a key positive regulator of plant hypersensitive cell death. EMBO J, 2009, 28: 926-936[本文引用:1]
[26]
NakayamaA, FukushimaS, GotoS, MatsushitaA, ShimonoM, SuganoS, Jiang CJ, AkagiA, YamazakiM, InoueM, TakatsujiH. Genome-wide identification of WRKY45-regulated genes that mediate benzothiadiazole-induced defense responses in rice. BMC Plant Biol, 2013, 13: 1-11[本文引用:1]
[27]
Song SY, ChenY, ChenJ, Dai XY, Zhang WH. Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta, 2011, 234: 331-345[本文引用:1]
[28]
OhnishiT, SugaharaS, YamadaT, KikuchiK, YoshibaY, Hirano HY, TsutsumiN. OsNAC6, a member of the NAC gene family, is induced by various stresses in rice. Genes Genet Syst, 2005, 80: 135-139[本文引用:1]
[29]
Hu HH, YouJ, Fang YJ, Zhu XY, Qi ZY, Xiong LZ. Characterization of transcriotion factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol Biol, 2008, 67: 169-18[本文引用:1]
[30]
FujitaM, FujitaY, MaruyamaK, SekiM, HiratsuK, Ohme- TakagiM, Tran LS, Yamaguchi-ShinozakiK, ShinozakiK. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J, 2004, 39, 863-876[本文引用:1]
[31]
Lu PL, Chen NZ, AnR, SuZ, Qi BS, RenF, ChenJ, Wang XC. A novel drought-inducible gene, ATAF1, encodes a NAC family protein that negatively regulates the expression of stress-responsive genes in Arabidopsis. Plant Mol Biol, 2007, 63: 289-305[本文引用:2]
[32]
WuJ, Wang LF, Wang SM. Comprehensive analysis and discovery of drought-related NAC transcription factors in common bean. BMC Plant Biol, 2016, 16: 193[本文引用:1]
[33]
SekiM, NarusakaM, AbeH, KasugaM, Yamaguchi-ShinozakiK, CarninciP, HayashizakiY, ShinozakiK. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell, 2001, 13: 61-72[本文引用:1]
[34]
FowlerS, Thomashow MF. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell, 2002, 14: 1675-1690[本文引用:1]
[35]
MaruyamaK, SakumaY, KasugaM, ItoY, SekiM, GodaH, ShimadaY, YoshidaS, ShinozakiK, Yamaguchi-ShinozakiK. Identification of cold-inducible downstream genes of the Arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. Plant J, 2004, 38: 982-993[本文引用:1]
[36]
ShinozakiK, Yamaguchi-ShinozakiK. Gene networks involved in drought stress response and tolerance. J Exp Bot, 2007, 58: 221-227[本文引用:1]
[37]
ChoiH, HongJ, HaJ, KangJ, Kim SY. ABFs, a family of ABA-responsive element binding factors. J Biol Chem, 2000, 275: 1723-1730[本文引用:1]
[38]
UnoY, FurihataT, AbeH, YoshidaR, ShinozakiK, Yamaguchi-ShinozakiK. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc Natl Acad Sci USA, 2000, 97: 11632-11637[本文引用:1]
[39]
NakashimaK, Tran LS, Van NguyenD, FujitaM, MaruyamaK, TodakaD, ItoY, HayashiN, ShinozakiK, Yamaguchi-ShinozakiK. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J, 2007, 51: 617-630[本文引用:1]
[40]
SakumaY, LiuQ, Dubouzet JG, AbeH, ShinozakiK, Yamaguchi-ShinozakiK. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem Biophys Res Commun, 2002, 290: 998-1009[本文引用:1]
[41]
赵洪阳. 水稻抗旱基因分子进化研究. 华中农业大学硕士学位论文, 湖北武汉, 2012Zhao HY. Molecular Evolution of Drought-Resistant Genes in Rice. MS Thesis of Huazhong Agricultural University, Wuhan, China, 2012 (in Chinese with English abstract)[本文引用:1]
[42]
HickmanR, HillC, Penfold CA, BreezeE, BowdenL, Moore JD, ZhangP, JacksonA, CookeE, Bewicke-CopleyF, MeadA, BeynonJ, Wild DL, Denby KJ, OttS, Buchanan-WollastonV. A local regulatory network around three NAC transcription factors in stress responses and senescence in Arabidopsis leaves. Plant J, 2013, 75: 26-39[本文引用:1]
[43]
BhattramakkiD, DolanM, HanafeyM, Wineland R, VaskeD, Register J C 3rd, Tingey S V, Rafalski A. Insertion deletion polymorphisms in 3' regions of maize genes occur frequently and can be used as highly informative genetic markers. Plant Mol Biol, 2002, 48: 539-547[本文引用:1]
[44]
YangD, YangX, LiuJ, Wang BH, Liu BL, Wang YZ. Pod shattering resistance associated with domestication is mediated by an NAC gene in soybean. Nat Commun, 2014, 5: 3352[本文引用:1]
 , 马艺文
, 马艺文





{kind=link}
{kind=link}
{kind=link}