关键词:木薯; MeHDZ14; 结构特性; 表达分析 Cloning and Analysis of the Transcription Factor Gene MeHDZ14 in Cassava YU Xiao-Ling, RUAN Meng-Bin, WANG Bin, YANG Yi-Ling, WANG Shu-Chang*, PENG Ming* Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Science / Key Laboratory of Biology and Genetic Resources of Tropical Crops, Ministry of Agriculture, Haikou 571101, China Fund:This study was supported by the International Science & Technology Cooperation Program of China (31561143012, 2013DFA32020) and the National Natural Science Foundation of China (31501378) AbstractHD-Zip family genes play important roles in plant growth and stress response. To reveal the role of MeHDZ14gene in abiotic stresses (e.g. drought) in cassava, we cloned MeHDZ14 gene by using RT-PCR from cassava cultivar SC124, which was relatively more resistant to drought stress. Bioinformatics methods were used to analyze its structural characteristics, and semi-quantitative RT-PCR/qRT-PCR was used to explore its expression patterns in response to abiotic stresses in different plant tissues and varieties. MeHDZ14 has a 726 bp open reading frame, encoding 241 amino acids, and contains the typical HD and ZIP domain. Blastp analysis showed that MeHDZ14 has close genetic relationship with ATHB-7, which is a member of the family I HD-Zip gene. Yeast and subcellular localization test showed that the MeHDZ14 gene is a transcription factor and specifically expresses in the nucleus. Genetic structural variation analysis revealed a total of four mis-sense mutations in eight tested varieties. However, amino acid mutations were not found between wild and cultivated cassavas. This indicates the MeHDZ14 proteins are highly conserved. Semi-quantitative RT PCR analysis revealed that MeHDZ14was specifically expressed in petioles, and induced by drought stress in root and leaf, suggesting that MeHDZ14 plays an important role in the early drought stage. Analysis by qRT-PCR showed that MeHDZ14 gene had different expression levels in different cassava varieties, but the same mode under drought stress and ABA treatment. These data indicate that MeHDZ14 is a member of the ABA pathway responding to drought. Our results showed that MeHDZ14 plays an important role in the molecular pathways of cassava drought resistance, underlining its potential in genetic improvement of cassava drought tolerance.
Keyword:Cassava; MeHDZ14; Structure characteristics; Expression analysis Show Figures Show Figures
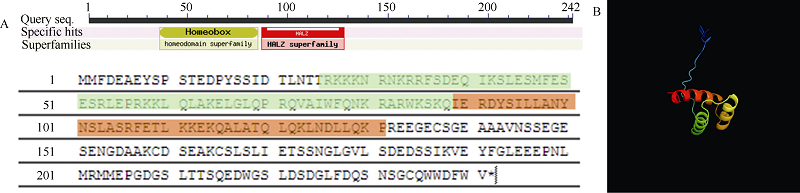
图1MeHDZ14基因结构分析 A: 基因结构及序列示意图; B: 蛋白质三维空间结构示意图。Fig. 1 Bioinformatics analysis results of MeHDZ14 A: sequence and structural analysis of MeHDZ14; B: the three-dimensional space structure of protein.
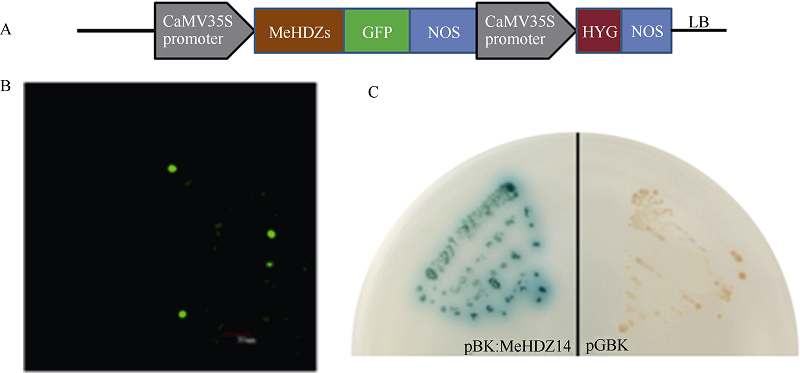
图2 转录因子验证结果 A: 植物表达载体载体骨架; B: 亚细胞定位结果(烟草叶表皮细胞); C: 目标基因酵母自激活试验。Fig. 2 Validation experiments of transcription factors A: schematic representation of the T-DNA region of MeHDZ14; B: subcellular localization of MeHDZ14 as revealed by GFP fusion protein in tobacco leaf-cells; C: α -X-gal developing of MeHDZ14.
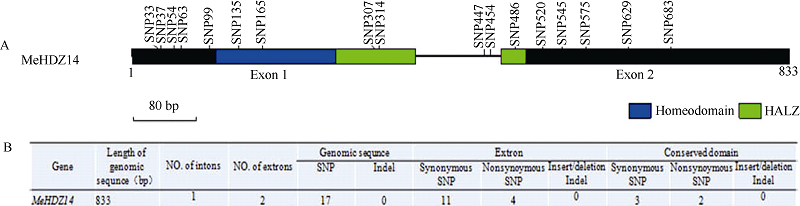
图5MeHDZ14基因在不同木薯品种中的核酸差异分析 A: 基因结构及SNP和Indel多态性位点分析; B: 基因自然变异分析。Fig. 5 DNA sequence of MeHDZ14 gene in different cassava varieties A: gene structure and their SNP and Indel polymorphism sites; B: the natural variation analysis of MeHDZ14.
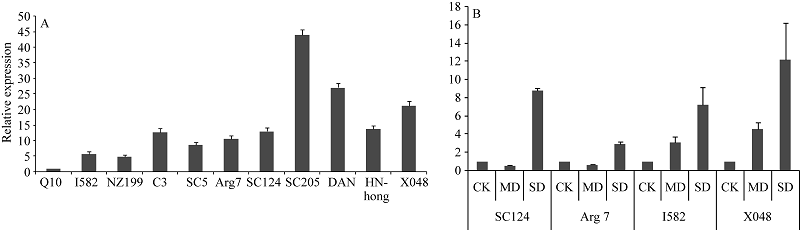
图6 不同品种木薯叶片中MeHDZ14基因的表达 A: 不同木薯品种中的相对表达; B: 干旱处理下, 4个木薯品种叶片中MeHDZ14基因的表达; MD: 中度干旱; SD: 重度干旱。Fig. 6 Expression profile of MeHDZ14 gene in different varieties of cassava A: relative expression level in different cassava varieties; B: under drought stress, the expression of MeHDZ14 in four different varieties of cassava. MD: moderate drought; SD: severe drought.
MizoiJ, ShinozakiK, Yamaguchi-ShinozakiK. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim Biophys Acta, 2012, 1819: 86-96[本文引用:1]
[2]
ShinozakiK, Yamaguchi-ShinozakiK. Gene networks involved in drought stress response and tolerance. J Exp Bot, 2007, 58: 221-227[本文引用:1]
[3]
康宗利, 杨玉红, 张立军. 植物响应干旱胁迫的分子机制. 玉米科学, 2006, 14(2): 96-100Kang ZL, Yang YH, Zhang LJ. Molecular mechanism of responsing to drought stress in plants. Maize Sci, 2006, 14(2): 96-100 (in Chinese with English abstract)[本文引用:1]
[4]
AsoK, KatoM, Banks JA, HasebeM. Characterization of homeodomain-leucine zipper genes in the fern Ceratopteris richardii and the evolution of the homeodomain-leucine zipper gene family in vascular plants. Mol Biol Evol, 1999, 16: 544-552[本文引用:1]
[5]
SakakibaraK, NishiyamaT, KatoM, HasebeM. Isolation of homeodomain-leucine zipper genes from the moss Physcomitrella patens and the evolution of homeodomain-leucine zipper genes in land plants. Mol Biol Evol, 2001, 18: 491-502[本文引用:1]
[6]
SchenaM, Davis RW. HD-Zip proteins: members of an Arabidopsis homeodomain protein superfamily. Proc Natl Acad Sci USA, 1992, 89: 3894-3898[本文引用:1]
[7]
Meijer AH, Scarpella E, van Dijk E L, Qin L, Taal A J, Rueb S, Harrington S E, McCouch S R, Schilperoort R A, Hoge J H. Transcriptional repression by Oshox1, a novel homeodomain leucine zipper protein from rice. Plant J, 1997, 11: 263-276[本文引用:1]
[8]
ZhaoY, ZhouY, JiangH, LiX, GanD, PengX, ZhuS, ChengB. Systematic analysis of sequences and expression patterns of drought-responsive members of the HD-Zip gene family in maize. PLoS One, 2011, 6: e28488[本文引用:1]
[9]
Re DA, Dezar CA, Chan RL, Baldwin IT, BonaventureG. Nicotiana attenuata NaHD20 plays a role in leaf ABA accumulation during water stress, benzylacetone emission from flowers, and the timing of bolting and flower transitions. J Exp Bot, 2011, 62: 155-166[本文引用:1]
[10]
ZhangZ, ChenX, GuanX, LiuY, ChenH, WangT, MouekoubaLD, LiJ, WangA. A genome-wide survey of homeodomain- leucine zipper genes and analysis of cold-responsive HD-Zip I members’ expression in tomato. Biosci Biotechnol Biochem, 2014, 78: 1337-1349[本文引用:1]
[11]
MatsumotoT, MorishigeH, TanakaT, KanamoriH, KomatsudaT, SatoK, ItohT, WuJ, NakamuraS. Transcriptome analysis of barley identifies heat shock and HD-Zip I transcription factors up-regulated in response to multiple abiotic stresses. Mol Breed, 2014, 34: 761-768[本文引用:1]
[12]
ChenX, ChenZ, ZhaoH, ZhaoY, ChengB, XiangY. Genome-wide analysis of soybean HD-Zip gene family and expression profiling under salinity and drought treatments. PLoS One, 2014, 9: e87156[本文引用:1]
[13]
Rueda EC, Dezar CA, Gonzalez DH, Chan RL. Hahb-10a sunflower homeobox-leucine zipper gene, is regulated by light quality and quantity, and promotes early flowering when expressed in Arabidopsis. Plant Cell Physiol, 2005, 46: 1954-1963[本文引用:2]
[14]
SongA, LiP, XinJ, ChenS, ZhaoK, WuD, FanQ, GaoT, ChenF, GuanZ. Transcriptome-wide survey and expression profile analysis of putative chrysanthemum HD-Zip I and II genes. Genes, 2016, 7: 19[本文引用:1]
[15]
Ariel FD, Manavella PA, Dezar CA, Chan RL, Chan RL. The true story of the HD-Zip family. Trends Plant Sci, 2007, 12: 419-426[本文引用:2]
[16]
Chan RL, Gago GM, Palena CM, Gonzalez DH. Homeoboxes in plant development. Biochim Biophys Acta, 1998, 1442: 1-19[本文引用:2]
[17]
Olsson AS, EngstromP, SodermanE. The homeobox genes ATHB12 and ATHB7 encode potential regulators of growth in response to water deficit in Arabidopsis. Plant Mol Biol, 2004, 55: 663-677[本文引用:1]
[18]
Gago GM, Jordano C A J, Gonzalez D H, Chan R L. Hahb-4, a homeobox-leucine zipper gene potentially involved in abscisic acid-dependent responses to water stress in sunflower. Plant Cell Environ, 2002, 25: 633-640[本文引用:1]
[19]
Cabello JV, Arce AL, Chan RL. The homologous HD-Zip I transcription factors HaHB1 and AtHB13 confer cold tolerance via the induction of pathogenesis-related and glucanase proteins. Plant J, 2012, 69: 141-153[本文引用:2]
[20]
AgalouA, PurwantomoS, OvernasE, JohannessonH, ZhuX, Estiati A, de Kam R J, Engstrom P, Slamet-Loedin I H, Zhu Z Wang M, Xiong L, Meijer A, Ouwerkerk P. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol Biol, 2008, 66: 87-103[本文引用:2]
[21]
HansonJ, JohannessonH, EngstromP. Sugar-dependent alterations in cotyledon and leaf development in transgenic plants expressing the HD-Zip gene ATHB13. Plant Mol Biol, 2001, 45: 247-262[本文引用:1]
[22]
HenrikssonE, Olsson AS, JohannessonH, JohanssonH, HansonJ, EngstromP, SodermanE. Homeodomain leucine zipper class I genes in Arabidopsis. expression patterns and phylogenetic relationships. Plant Physiol, 2005, 139: 509-518[本文引用:1]
[23]
WangY, HenrikssonE, SodermanE, Henriksson KN, SundbergE, EngstromP. The Arabidopsis homeobox gene, ATHB16, regulates leaf development and the sensitivity to photoperiod in Arabidopsis. Dev Biol, 2003, 264: 228-239[本文引用:1]
[24]
朱莹莹, 于亮亮, 汪杏芬, 李来庚. HD-ZIP III转录因子家族与植物细胞分化. 植物学报, 2013, 48: 199-209Zhu YY, Yu LL, Wang XF, Li LG. HD-Zip III transcription factor and cell differentiation in plants. Chin Bull Bot, 2013, 48: 199-209 (in Chinese with English abstract)[本文引用:1]
[25]
LechnerE, LeonhardtN, EislerH, ParmentierY, AliouaM, JacquetH, LeungJ, GenschikP. MATH/BTB CRL3 receptors target the homeodomain-leucine zipper ATHB6 to modulate abscisic acid signaling. Dev Cell, 2011, 21: 1116-1128[本文引用:1]
[26]
ParkJ, Lee HJ, Cheon CI, Kim SH, Hur YS, Auh CK, Im KH, Yun DJ, LeeS, Davis KR. The Arabidopsis thaliana homeobox gene ATHB12 is involved in symptom development caused by geminivirus infection. PLoS One, 2011, 6: e20054[本文引用:1]
[27]
SonO, Hur YS, Kim YK, Lee HJ, KimS, Kim MR, Nam KH, Lee MS, Kim BY, ParkJ. ATHB12, an ABA-inducible homeodomain-leucine zipper (HD-Zip) protein of Arabidopsis, negatively regulates the growth of the inflorescence stem by decreasing the expression of a gibberellin 20-oxidase gene. Plant Cell Physiol, 2010, 51: 1537-1547[本文引用:1]
[28]
ElhitiM, StasollaC. Structure and function of homodomain- leucine zipper (HD-Zip) proteins. Plant Signal Behav, 2009, 4: 86-88[本文引用:1]
[29]
HimmelbachA, HoffmannT, LeubeM, HohenerB, GrillE. Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis. EMBO J, 2002, 21: 3029-3038[本文引用:1]
[30]
MaydaE, TorneroP, ConejeroV, VeraP. A tomato homeobox gene (HD-Zip) is involved in limiting the spread of programmed cell death. Plant J, 1999, 20: 591-600[本文引用:1]
[31]
ZhaoY, MaQ, JinX, PengX, LiuJ, DengL, YanH, ShengL, JiangH, ChengB. A novel Maize homeodomain-leucine zipper (HD-Zip) I gene, Zmhdz10positively regulates drought and salt tolerance in both rice and Arabidopsis. Plant Cell Physiol, 2014, 55: 1142-1156[本文引用:1]
[32]
Manavella PA, Arce AL, Dezar CA, BittonF, Renou JP, CrespiM, Chan RL. Cross-talk between ethylene and drought signalling pathways is mediated by the sunflower Hahb-4 transcription factor. Plant J, 2006, 48: 125-137[本文引用:1]
[33]
ZhaoP, LiuP, ShaoJ, LiC, WangB, GuoX, YanB, XiaY, PengM. Analysis of different strategies adapted by two cassava cultivars in response to drought stress: ensuring survival or continuing growth. J Exp Bot, 2015, 66: 1477-1488[本文引用:2]
[34]
于晓玲, 王淦, 阮孟斌, 刘恩世, 彭明. 水分胁迫对不同木薯品种叶片生理生化的影响. 中国农学通报, 2012, 28(33): 60-64Yu XL, WangG, Ruan MB, Liu ES, PengM. Physiological and biochemical changes of leaves in different cassava varieties under water stress. Chin Agric Sci Bull, 2012, 28(33): 60-64 (in Chinese with English abstract)[本文引用:1]
[35]
Hebsgaard SM, Korning PG, TolstrupN, EngelbrechtJ, RouzeP, BrunakS. Splice site prediction in Arabidopsis thaliana pre-mRNA by combining local and global sequence information. Nucl Acids Res, 1996, 24: 3439-3452[本文引用:1]
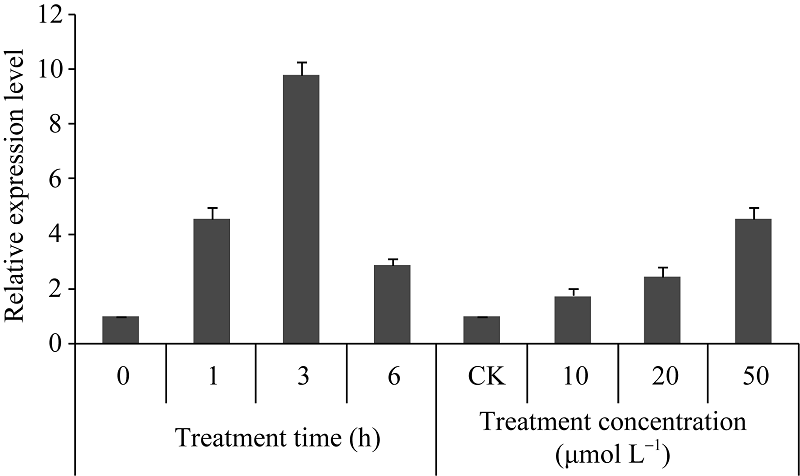
SodermanE, MattssonJ, EngstromP. The Arabidopsis homeobox gene ATHB-7 is induced by water deficit and by abscisic acid. Plant J, 1996, 10: 375-381[本文引用:2]
[38]
Van HoveJ, StefanowiczK, De SchutterK, EggermontL, LannooN, Al AtalahB, Van Damme E J. Transcriptional profiling of the lectin ArathEULS3 from Arabidopsis thaliana toward abiotic stresses. J Plant Physiol, 2014, 171: 1763-1773[本文引用:1]
[39]
Raghavendra AS, Gonugunta VK, ChristmannA, GrillE. ABA perception and signaling. Trends Plant Sci, 2010, 15: 395-401[本文引用:1]
[40]
ChristmannA, Weiler EW, SteudleE, GrillE. A hydraulic signal in root-to-shoot signaling of water shortage. Plant J, 2007, 52: 167-174[本文引用:1]
[41]
ZellerG, Henz SR, Widmer CK, SachsenbergT, RatschG, WeigelD, LaubingerS. Stress-induced changes in the Arabidopsis thaliana transcriptome analyzed using whole-genome tiling arrays. Plant J, 2009, 58: 1068-1082[本文引用:1]
 , 阮孟斌, 王斌, 杨义伶, 王树昌
, 阮孟斌, 王斌, 杨义伶, 王树昌





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}