关键词:水稻; 染色体片段代换系; 紫鞘 PSH1; 基因定位 Identification of Rice Chromosome Segment Substitution Line Z519 with Purple Sheath and Candidate Gene Analysis of PSH1 ZHOU Ke1,**, LI Yan1,2,**, WANG Shi-Ming1, CUI Guo-Qing1, YANG Zheng-Lin1, HE Guang-Hua1, LING Ying-Hua1, ZHAO Fang-Ming1,* 1 Rice Research Institute, Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400716, China
2 Tea Research Institute of Guizhou Province, Guiyang 550006, China
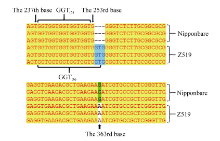
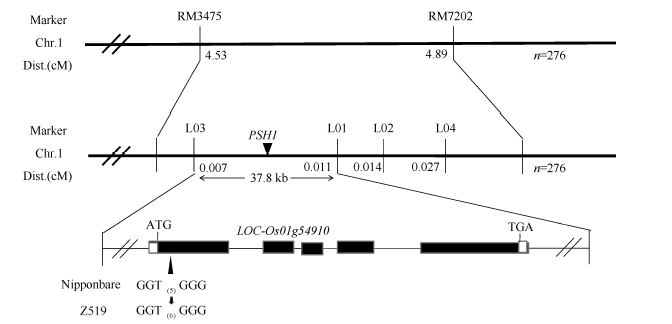
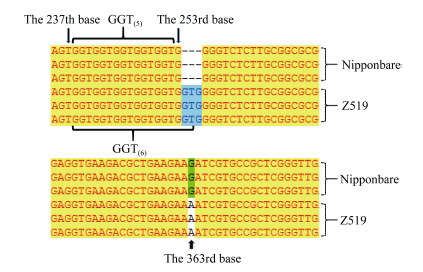
Fund:This study was supported by the subject “New Technology of Heterosis Utilization in Rice” (2016YFD0101107) in the National Key Research and Development Program (SQ2016ZY03001818), and Chongqing Key Laboratory Capacity Improvement Project (cstc2014pt-sy80001). AbstractAnthocyanins as plant pigments are widely liked by people and play a very important role in food processing and hybrid purity identification. Here, a rice chromosome segment substitution line (CSSL) Z519 with purple sheath was identified deriving from recipient Nipponbare and donor R225. Z519 contained 16 substitution segments with 6.85 Mb of average length, which were distributed on 11 chromosomes of rice except the 10th chromosome. The bud sheath of Z519 began to appear the purple color stripes when it was about 3 mm long. Then the purple stripes displayed on sheaths, leaf margins, vascular bundles of stem and stigmas. While all parts of Nipponbare were green. Anthocyanin content in leaf sheath of Z519 was significantly higher than that of Nipponbare, whereas no significant difference was in flag leaf. Compared with Nipponbare, plant height of Z519 was significantly decreased, spikelets number and grain number of main panicle, and 1000-grain weight of Z519 were significantly increased. There was no significant difference between Z519 and Nipponbare in the other traits such as panicle number, main panicle length and seed-setting rate. Then, F2 population from the cross of Nipponbare and Z519 were used for genetic analysis and gene mapping of the purple sheath. The purple sheath in Z519 was controlled by a single dominant gene, named as PSH1, which was mapped on the chromosome 1 between InDel marker L03 and SSR marker L01 with the physical distance of 37.8 kb. By sequencing and gene-predicting in the region, Z519 had three bases (GTG) insertion in the GTG repeat area of the 238th-252th base in the first exon compared with Nipponbare, which resulted in increasing a Gly amino acid. Furthermore, the expression of LOC_Os01g45910was obviously decreased in Z519 by qRT-PCR analysis. Thus, LOC_Os01g45910was preliminary identified as the candidate gene of PSH1. The results lay a good foundation for studying molecular mechanisms of regulating anthocyanin by PSH1.
Keyword:Rice ( Oryza sativa L.); Chromosome segment substitution line (CSSL); Purple sheath PSH1; Gene mapping Show Figures Show Figures
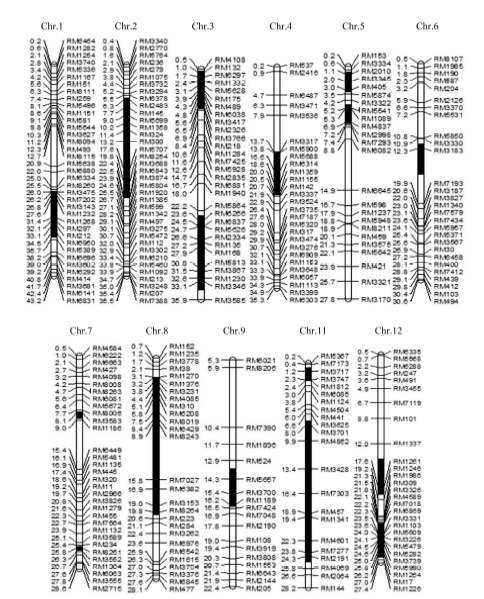
图1 Z519的染色体代换片段(图中黑色区段为代换片段所在的位置, 染色体左端数字为物理图距(Mb), 右侧为分子标记。)Fig. 1 Chromosome substitution segments of Z519(Black intervals indicate substitution segments. The digits on the left of chromosomes represent physical distance (Mb) while the markers are on the right.)
图2 日本晴和Z519的表型(A: 苗期日本晴和Z519的表型; B: 分蘖期日本晴、Z519和供体R225的鞘; C: 出芽期日本晴和Z519的鞘; D: 日本晴和Z519的雌蕊柱头; E: 抽穗期日本晴和Z519的表型。)Fig. 2 Phenotype of Nipponbare and Z519(A: plant morphology of Nipponbare and Z519 at the seeding stage; B: sheath of Nipponbare, Z519 and donor R225 at the tillering stage; C: sheath of Nipponbare and Z519 at the sprouting stage; D: pistil stigma of Nipponbare and Z519; E: plant morphology of Nipponbare and Z519 at the heading stage.)
表1 Table 1 表1(Table 1)
表1 日本晴和Z519的农艺性状 Table 1 Agronomic traits of Nipponbare and Z519
材料 Material
株高 Plant height (cm)
有效穗数 Panicles number per plant
主穗长 Main panicle length (cm)
主穗总粒数 Spikeletes of main panicle
主穗实粒数 Grains of main panicle
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
日本晴 Nipponbare
110.20± 2.85
11.80± 2.14
24.10± 1.29
192.60± 11.77
159.50± 10.57
83.04± 6.39
20.67± 1.27
Z519
104.30± 2.84* *
10.10± 1.97
23.60± 2.35
214.38± 8.34* *
172.44± 12.80*
79.64± 6.79
24.55± 2.17* *
* 和* * 分别表示在0.05和0.01水平差异显著。* and * * indicate difference at P< 0.05 and P< 0.01, respectively.
表1 日本晴和Z519的农艺性状 Table 1 Agronomic traits of Nipponbare and Z519
图3 日本晴和Z519的叶绿素及花青素含量(A: 抽穗期剑叶的叶绿素含量; B: 抽穗期鞘的叶绿素含量; C: 抽穗期剑叶和鞘中的花青素含量。)Fig. 3 Contents of chlorophyll and anthocyanidin of Nipponbare and Z519(A: chlorophyll content of the flag leaves at the heading stage; B: chlorophyll content of the sheath at the heading stage; C: anthocyanin content in the sheath and flag leaves of Nipponbare and Z519 at the heading stage.)
唐忠厚, 周丽. 花青素对人类健康影响的研究进展及其前景. 食品研究与开发, 2009, (7): 159-162Tang ZH, ZhouL. Study on anthocyanins influencing on human health and its prospect. Food R&D, 2009, (7): 159-162 (in Chinese with English abstract)[本文引用:1]
[2]
Holton TA, Cornish EC. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell, 1995, 7: 1071-1083[本文引用:3]
[3]
Austin MB, Noe L J P. The chalcone synthase superfamily of type III polyketide synthases. Nat Prod Rep, 2003, 20: 79-110[本文引用:1]
[4]
YamazakiM, NakajimaJ, YamanashiM, SugiyamaM, MakitaY, SpringobK, AwazuharaM, SaitoK. Metabolomics and differential gene expression in anthocyanin chemo-varietal forms of Perilla frutescens. Phytochemistry, 2003, 62: 987-995[本文引用:1]
[5]
BogsJ, EbadiA, McDavid D, Robinson P. Identification of the flavonold hydroxylases from grapevine and their regulation during fruit development. Plant Physiol, 2006, 140: 279-291[本文引用:1]
[6]
Jeong ST, Goto-YamamotoN, HashizumeK, EsakaM. Expression of the flavonoid 3'-hydroxylase and flavonoid 3', 5' -hydroxylase genes and flavonoid composition in grape( Vitis vinifera). Plant Sci, 2006, 170: 61-69[本文引用:1]
[7]
Castellarin SD, Di GasperoG, MarconiR, NonisA, PeterlungerE, PaillardS, Adam-Blondon A F, Testolin R. Colour variation in red grapevines ( Vitis vinifera L. ): genomic organisation, expression of flavonoid 3'-hydroxylase, flavonoid 3', 5’-hydroxylase genes and related metabolite profiling of red cyanidin-/blue delphinidin- based anthocyanins in berry skin. BMC Genomics, 2006, 7: 12[本文引用:1]
[8]
Ramsay NA, Glover BJ. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci, 2005, 10: 63-70[本文引用:1]
[9]
Xie DY, Sharma SB, WrightE, Wang ZY, Dixon RA. Metabolic engineering of proanthocyanidins through co-expression of anthocyanidin reductase and the PAP1 MYB transcription factor. Plant J, 2006, 45: 895-907[本文引用:1]
[10]
GonzalezA, Zhao MZ, Leavitt JM, Lloyd AM. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsisseedings. Plant J, 2007, 53: 814-827[本文引用:1]
[11]
GrotewoldE. The challenges of moving chemicals within and out of cells: insights into the transport of plant natural products. Planta, 2004, 219: 906-909[本文引用:1]
[12]
PourcelL, Irani NG, LuY, RiedlK, SchwartzS, GrotewoldE. The formation of anthocyanic vacuolar inclusions in Arabidopsis thalianaand implications for the sequestration of anthocyanin pigments. Mol Plant, 2010, 3: 78-90[本文引用:1]
[13]
Marrs KA, Alfenito MR, Lloyd AM, WalbotV. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature, 1995, 395: 397-400[本文引用:1]
[14]
KitamuraS, ShikazonoN, TanakaA. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant J, 2004, 37: 104-114[本文引用:1]
[15]
张毅. 水稻两个芽鞘紫线相关基因的图位克隆与应用研究. 西南大学博士学位论文, 重庆, 2009ZhangY. Map-based Cloning and Application Research of Two Coleoptile Purple Line Related Genes in Rice ( Oryza sativaL. ). PhD Dissertation of Southwest University, Chongqing, China, 2009 (in Chinese with English abstract)[本文引用:4]
[16]
Wang WY, Ding HF, Li GX, Jiang MS, Li LF, LiuX, ZhangY, Yao FY. Delimitation of the PSH1(t)for rice purple leaf sheath to a 23. 5 kb DNA fragment. Genome, 2009, 52: 268-274[本文引用:4]
[17]
孙旭, 柳絮, 李润芳, 王文英, 姚方印, 刘开启. 水稻紫色叶鞘基因的遗传分析. 山东农业科学, 2009, (10): 22-24SunX, LiuX, Li RF, Wang WY, Yao FY, Liu KQ. Genetic analysis of gene for purple leaf sheath in rice. Shand ong Agric Sci, 2009, (10): 22-24 (in Chinese with English abstract)[本文引用:1]
[18]
SakamotoW, OhmoriT, KageyamaK, MiyazakiC, SaitoA, MurataM, NodaK, MaekawaM. The purple leaf (pl)locus of rice: plwallele has a complex organization and includes two genes encoding basic helix-loop-helix proteins involved in anthocyanin biosynthesis. Plant Cell Physiol, 2001, 42: 982-991[本文引用:1]
[19]
赵芳明, 郭超, 魏霞, 杨正林, 凌英华, 桑贤春, 王楠, 张长伟, 李云峰, 何光华. 日本晴与5个优良恢复系的多态性标记筛选及遗传差异分析. 西南大学学报(自然科学版), 2016, 38(11): 1-7Zhao FM, GuoC, WeiX, Yang ZL, Ling YH, Sang XC, WangN, Zhang CW, Li YF, He GH. Polymorphic SSR markers screening and genetic difference analysis beween Nipponbare and five excellent restorer lines. J Southwest Univ(Nat Sci Edn), 2016, 38(11): 1-7 (in Chinese with English abstract)[本文引用:1]
[20]
Paterson AH, DamonS, Hewitt JD, ZamirD, Rabinowitch HD, Lincoln SD, Land er ES, Tanksley SD. Mendelian factors underlying quantitative traits in tomato: comparison across species, generations, and environments. Genetics, 1991, 127: 181-197[本文引用:1]
[21]
邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2000. pp 27-29ZouQ. Experimental Guide of Plant Physiology. Beijing: China Agriculture Press, 2000. pp 27-29(in Chinese)[本文引用:1]
[22]
于晓南. 植物叶片中花青素的分析与研究. 现代仪器, 2000, (4): 37-38Yu XN. Analysis and study on anthocyanin in plant leaves. Modern Instruments, 2000, (4): 27-29 (in Chinese)[本文引用:1]
[23]
向佳, 李燕, 樊亚伟, 许军红, 郑丽媛, 何光华, 杨正林, 王楠, 赵芳明. 一个具有主效晚抽穗基因的水稻染色体片段代换系的鉴定、形态分析及 Ehd4-2定位. 作物学报, 2015, 41: 683-691XiangJ, LiY, Fan YW, Xu JH, Zheng LY, He GH, Yang ZL, WangN, Zhao FM. Identification and morphological analysis of a rice chromosome segment substitution line carrying a major effect gene for late heading date and mapping of Ehd4-2. Acta Agron Sin, 2015, 41: 683-691 (in Chinese with English abstract)[本文引用:2]
[24]
Rogers SO, Bendich AJ. Extraction of DNA from plant tissues. Plant Mol Biol, 1988, A6: 1-10[本文引用:1]
[25]
EbitaniT, TakeuchiY, NonoueY, YamamotoT, TakeuchiK, YanoM. Construction and evaluation of chromosome segment substitution lines carrying over lapping chromosome segments of indica rice cultivar “ Kasalath” in a genetic background of japonica elite cultivar Koshihikari. Breed Sci, 2005, 55: 65-73[本文引用:1]
[26]
TakaiT, NonoueY, YamamotoS, YamanouchiU, MatsubaraK, Liang ZW, Lin HX, OnoN, UgaY, YanoM. Development of chromosome segment substitution lines derived from backcross between indica donor rice cultivar “ Nona Bokra” and japonica recipient cultivar “ Koshihikari”. Breed Sci, 2007, 57: 257-261[本文引用:1]
[27]
熊振民. 中国水稻. 北京: 中国农业科技出版社, 1992. pp 55-57Xiong ZM. Rice in China. Beijing: China Agricultural Science and Technolegy Press, 1992. pp 55-77(in Chinese)[本文引用:1]
[28]
黄碧光, 卢泳全. 杂交水稻V优77 色素与育性的遗传关系研究. 福建农业学报, 2002, 17(4): 215-216Huang BG, Lu YQ. Study on the heredity of color and fertility in rice. Fujian J Agric Sci, 2002, 17(4): 215-216 (in Chinese with English abstract)[本文引用:1]
邓国富, 李杨瑞, 梁世荣, 陈仁天, 周萌, 高利军, 周维永, 戴高兴. 紫叶水稻IR1552紫叶性状的遗传及其育种利用. 杂交水稻, 2007, 22(5): 71-75Deng GF, Li YR, Liang SR, Chen RT, ZhouM, Gao LJ, Zhou WY, Dai GX. Genetics and breeding of purple leaf character of purple leaf rice IR1552. Hybrid Rice, 2007, 22(5): 71-75 (in Chinese with English abstract)[本文引用:1]
[31]
钱前, 唐绍清, 林建荣, 吕子同. 一种具有遗传特异性的紫叶稻及其在两系法杂交稻中的应用. 浙江农业学报, 1995, 7: 259-262QianQ, Tang SQ, Lin JR, Lyu ZT. A purple leaf rice CN-plwith genetic specificity and its utilization on two-line hybrid rice. Acta Agric Zhejiangensis, 1995, 7: 259-262 (in Chinese with English abstract)[本文引用:1]
[32]
谢国生, 蔡得田, 马平福, 祝虹. 水稻显性紫色基因的遗传分析. 湖北农业科学, 1997, 10(3): 3-5Xie GS, Cai DT, Ma PF, ZhuH. Genetic analysis of dominant purple gene in Rice. Hubei Agric Sci, 1997, 10(3): 3-5 (in Chinese with English abstract)[本文引用:1]
[33]
牟同敏, 李春海, 杨国才, 卢兴贵. 紫叶水稻苗期叶色的遗传研究. 中国水稻科学, 1995, 9: 45-48Mou TM, Li CH, Yang GC, Lu XG. Genetical studies on seedling leaf color of purple rice. Chin J Rice Sci, 1995, 9: 45-48 (in Chinese with English abstract)[本文引用:1]
[34]
Hu JP, AndersonB, Wessler SR. Isolation and characterization of rice R genes: evidence for distinct evolutionary paths in rice and maize. Genetics, 1996, 142: 1021-1031[本文引用:1]
[35]
谢小东, 程廷才, 王根洪, 段军, 夏庆友. 植物ABC和MATE转运蛋白与次生代谢物跨膜转运. 植物生理学报, 2011, 47: 752-758Xie XD, Cheng TC, Wang GH, DuanJ, Xia QY. ABC and MATEtransporters of plant and their roles in membrane transport of secondary metabolites. Plant Physiol Commun, 2011, 47: 752-758 (in Chinese with English abstract)[本文引用:1]
 , 李燕
, 李燕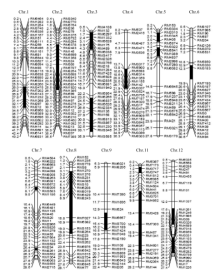








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}