关键词:甘蓝型油菜; 菌核病; 相关性分析; 数量性状位点; 木质素; 单体G/S Correlation Analysis of SclerotiniaResistance with Lignin Content and Monomer G/S and Its QTL Mapping in Brassica napus L. CHEN Xue-Ping*, JING Ling-Yun*, WANG Jia, JIAN Hong-Ju, MEI Jia-Qin, XU Xin-Fu, LI Jia-Na, LIU Lie-Zhao* College of Agronomy and Biotechnology, Southwest University / Chongqing Engineering Research Center for Rapeseed, Chongqing 400715, China Fund:This study was supported by the National Natural Science Foundation of China (31371655) and the Science and Technology Committee of Chongqing (cstc2016shmszx80083) Abstract Sclerotinia sclerotiorumis a fungal pathogen causing disease in a wide range of plants, resulting in serious damage in crop production. The detached stem inoculation assay of RIL and F2 populations at final flowering stage was conducted, near infrared (NIR) spectroscopy was used to measure lignin content and monomer G/S in the stem, and correlation analysis and QTL mapping for these traits were performed. The lesion size of the RIL had a significantly negative correlation with lignin content, with a correlation coefficient at -0.348 and -0.286 in 2013 and 2014, respectively. The monomer G/S was significantly correlated with lesion size in the RIL population, and the correlation coefficient at 0.198 and 0.167 in 2013 and 2014, respectively. The lesion size of F2 in 2014 was significantly and negatively correlated with lignin content in the stem, with a correlation coefficient at -0.306. The cross sections of resistant and susceptible plants from F2:3family were stained with phloroglucinol-HCl, showing that the content of lignin was significantly lower in the less resistant materials than in more resistant plants. According to the high density SNP genetic maps and composite interval mapping, a total of 18 QTLs were identified, which were located on A04, A05, A06, A08, C01, C03, C04, C06, and C07 chromosomes, with the explained phenotypic variation by individual QTL ranging from 2.38% to 12.05% for nine QTLs of lesion size, from 2.03% to 13.75% for three QTLs of lignin content, and from 2.06% to 8.66% for six QTLs of monomer G/S. The research results provide some new insights for the Sclerotiniaresistance breeding in B. napus.
Keyword: Brassica napus; Sclerotinia sclerotiorum; Correlation; QTL; Lignin; Monomer G/S Show Figures Show Figures
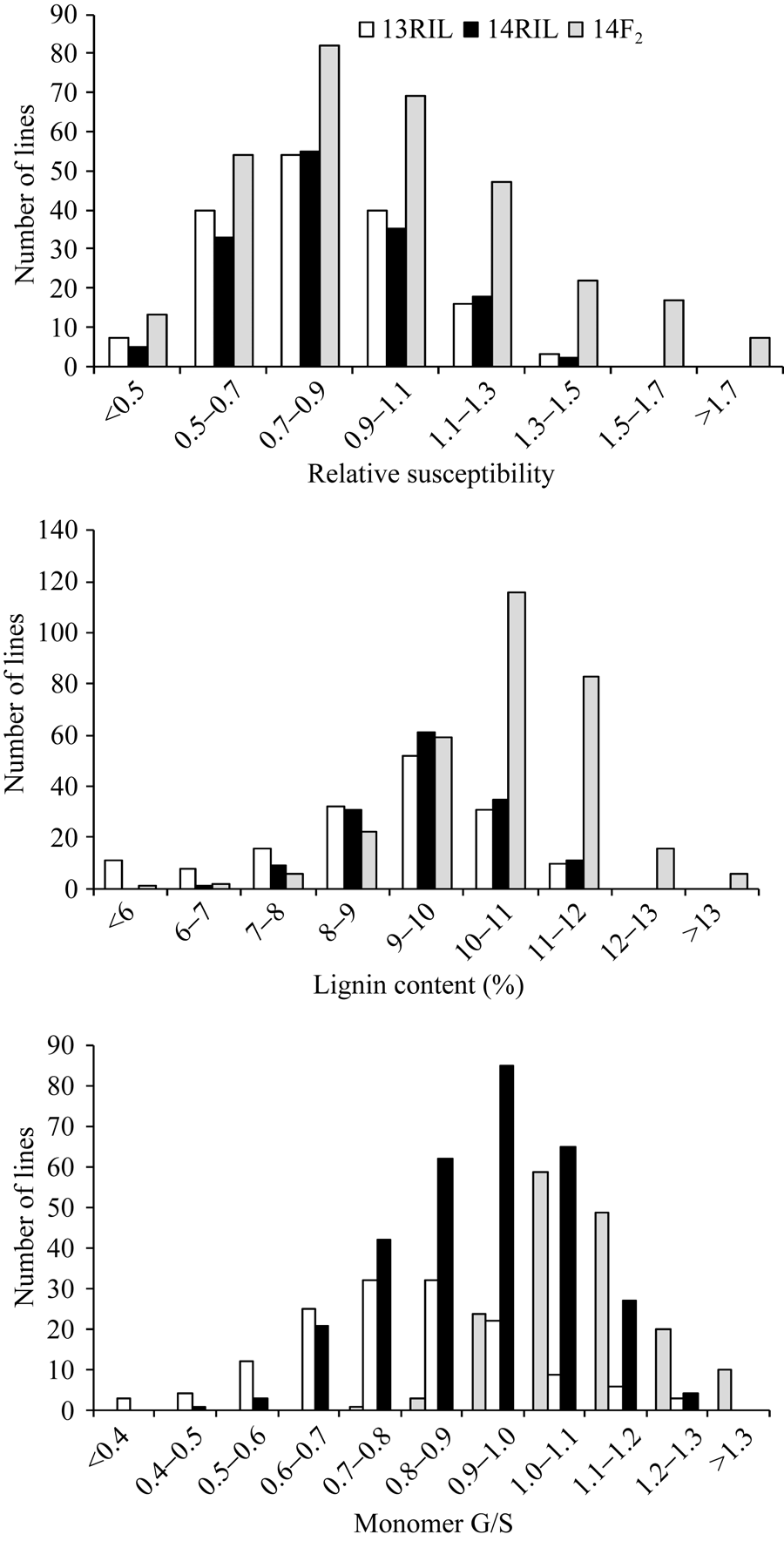
图1 RIL和F2群体茎秆相对感病度、木质素含量和单体G/S在2013年和2014年的频率分布Fig. 1 Frequency distribution of stem relative susceptibility, lignin content and monomer G/S fromB. napusRIL in 2013 and 2014 and F2in 2014
表1 Table 1 表1(Table 1)
表1 RIL群体茎秆相对感病度、木质素含量及单体G/S的表型分布特征 Table 1 Phenotype distribution feature of stem relative susceptibility, lignin content and monomer G/S for RIL populations
表1 RIL群体茎秆相对感病度、木质素含量及单体G/S的表型分布特征 Table 1 Phenotype distribution feature of stem relative susceptibility, lignin content and monomer G/S for RIL populations
表2 Table 2 表2(Table 2)
表2 F2群体茎秆相对感病度、木质素含量及单体G/S的表型分布特征 Table 2 Phenotype distribution feature of stem relative susceptibility, lignin content and monomer G/S for F2 populations
表2 F2群体茎秆相对感病度、木质素含量及单体G/S的表型分布特征 Table 2 Phenotype distribution feature of stem relative susceptibility, lignin content and monomer G/S for F2 populations
表3 2013年和2014年RIL群体菌斑大小、木质素含量及单体G/S的相关性分析 Table 3 Correlation coefficients among lesion size , lignin content and monomer G/S for RIL population in 2013 and 2014
性状 Trait
13LS
14LS
13ADL
14ADL
13G/S
2014LS
0.865* *
2013ADL
-0.348* *
-0.280* *
2014ADL
-0.236* *
-0.286* *
0.219* *
2013G/S
0.198*
0.178*
0.064
-0.152
2014G/S
0
0.167*
-0.073
0.088
0.206* *
LS: lesion size; ADL: lignin content; G/S: monomer G/S. * and * * denote significant correlation at the 0.05 and 0.01 probability levels, respectively. LS: 菌斑大小; ADL: 木质素含量; G/S: 单体G/S。* 和* * 分别代表在0.05和0.01显著水平。
表3 2013年和2014年RIL群体菌斑大小、木质素含量及单体G/S的相关性分析 Table 3 Correlation coefficients among lesion size , lignin content and monomer G/S for RIL population in 2013 and 2014
表4 Table 4 表4(Table 4)
表4 2014年F2群体菌斑大小、木质素含量及单体G/S的相关性分析 Table 4 Correlation coefficients among lesion size, lignin content and monomer G/S for F2 population in 2014
性状 Trait
14LS
14ADL
2014ADL
-0.306* *
2014G/S
0.142
0.079
LS: lesion size; ADL: lignin content; G/S: monomer G/S. * and * * denote significant correlation at the 0.05 and 0.01 probability levels, respectively. LS: 菌斑大小; ADL: 木质素含量; G/S: 单体G/S。* 和* * 分别代表在0.05和0.01显著水平。
表4 2014年F2群体菌斑大小、木质素含量及单体G/S的相关性分析 Table 4 Correlation coefficients among lesion size, lignin content and monomer G/S for F2 population in 2014
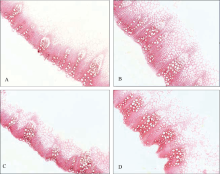
图2 抗病较弱材料(A, C)和抗病较强材料(B, D)在终花期的茎部横切面间苯三酚-盐酸染色观察以示木质素含量差异(10× 4)Fig. 2 Stem cross-section stained with phloroglucinol-HCl to show the difference of lignin content in the less resistant (A, C) and more resistant (B, D) lines by the end of flowering time (10× 4)
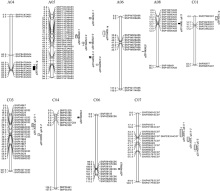
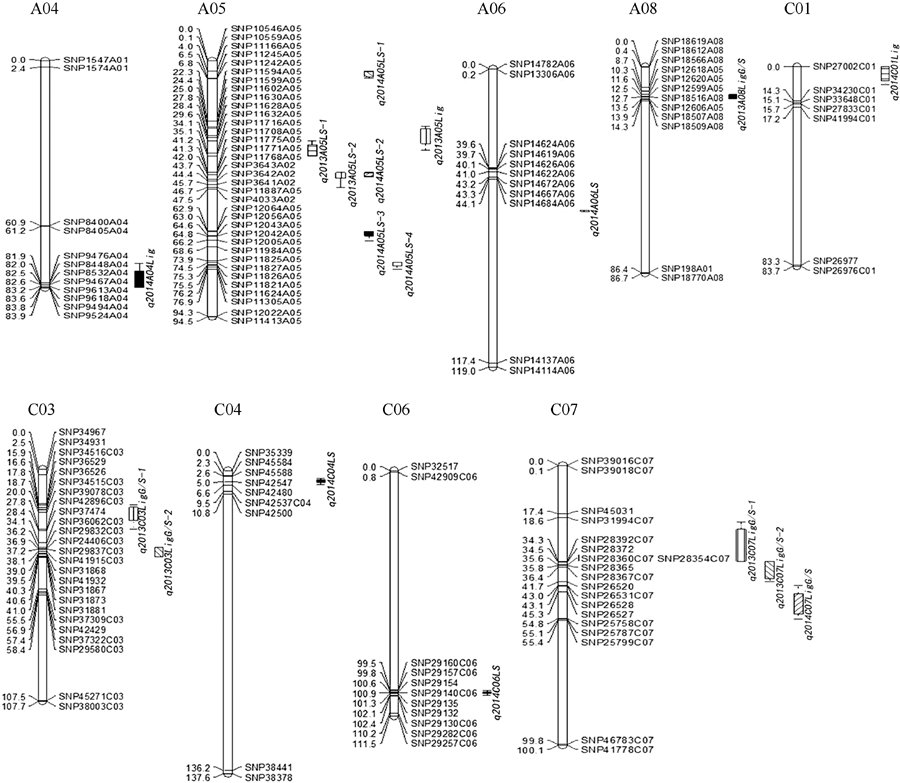
图3 甘蓝型油菜茎秆菌斑大小、木质素含量及单体G/S QTL在SNP连锁群上分布图QTL的染色体上仅展示了QTL区段的标记及染色体两端各2个标记。Fig. 3 Putative QTL locations of lesion size, lignin content, and monomer G/S for stem on the SNP genetic mapShowing the markers in the QTL confidence intervals, along with the terminal two markers at each end of chromosomes containing the QTL.
Carr RA, McDonald B E. Rapeseed in a changing world: Processing and utilization. GCIRC Eighth International Rapeseed Congress. Saskatoon, 1991. pp 39-56[本文引用:1]
[2]
李加纳, 谌利, 张学昆. 甘蓝型黄籽油菜的研究与思考. 北京: 中国农业科技出版社, 2004. pp 29-39Li JN, ChenL, Zhang XK. Research and thinking of yellow- seeded rapeseed (Brassica napus L. ). Beijing: China Science and Technology Press, 2004. pp 29-39(in Chinese)[本文引用:1]
[3]
ZhaoJ, MengJ. Genetic analysis of loci associated with partial resistance to Sclerotina sclerotiorum in rapeseed (Brassica napus L. ). Theor Appl Genet, 2003, 106: 759-764[本文引用:3]
[4]
GargH, SivasithamparamK, BangaS. Cotyledon assay as a rapid and reliable method of screening for resistance against Sclerotinia sclerotiorum in Brassica napus genotypes. Australas Plant Path, 2008, 37: 106-111[本文引用:1]
[5]
BradleyC, HameyH. Canola disease situation in North Dakota, USA, 1993-2004. 14th Australian Research Assembly on Brassicas. Port Lincoln, 2005. pp 33-34[本文引用:1]
[6]
Turkington TK, Morrall R A A. Use of petal infestation to forecast Sclerotinia stem rot of canola: the influence of inoculum variation over the flowering period and canopy density. Phytopathology, 1993, 83: 682-689[本文引用:1]
[7]
del Río LE, Bradley CA, Henson RA, Endres GJ, Hanson BK, McKayK, HalvorsonM, Porter PM, Le Gare DG, Lamey HA. Impact of Sclerotinia stem rot on yield of canola. Plant Dis, 2007, 91: 191-194[本文引用:1]
[8]
王汉中, 刘贵华, 郑元本, 王新发, 杨庆. 抗菌核病双低油菜新品种中双9号选育及其重要防御酶活性变化规律的研究. 中国农业科学, 2004, 37: 23-28Wang HZ, Liu GH, Zheng YB, Wang XF, YangQ. Breeding of the Brassica napus cultivar zhongshuang 9 with high-resistance to Sclerotinia sclerotiorum and dynamics of its important defense enzyme activity. Sci Agric Sin, 2004, 37: 23-28 (in Chinese with English abstract)[本文引用:1]
[9]
ZhaoJ, Udall JA, Quijada PA, Grau CR, MengJ, Osborn TC. Quantitative trait loci for resistance to Sclerotinia sclerotiorum and its association with a homeologous non-reciprocal transposition in Brassica napus L. Theor Appl Genet, 2006, 112: 509-516. [本文引用:3]
[10]
WuJ, CaiG, TuJ, LiL, LiuS, LuoX, ZhouL, FanC, ZhouY. Identification of QTLs for resistance to Sclerotinia stem rot and BnaC. IGMT5. a as a cand idate gene of the major resistant QTL SRC6 in Brassica napus. PloS One, 2013, 8: e67740[本文引用:2]
梅家琴. 甘蓝与甘蓝型油菜C亚基因组遗传关系调查及甘蓝抗菌核病QTL定位. 西南大学博士学位论文, 重庆, 2011. pp 62-73Mei JQ. Genetic Investigation of Relationships between Brassica oleracea and C Subgenome of B. napus and Mapping QTL for Sclerotinia sclerotiorum Resistance in B. oleracea. PhD Dissertation of Southwest University, Chongqing, China, 2011. pp 62-73 (in Chinese with English abstract)[本文引用:3]
[13]
WeiL, JianH, LuK, FilardoF, YinN, LiuL, QuC, WeiL, DuH, LiJ. Genome-wide association analysis and differential expression analysis of resistance to Sclerotinia stem rot in Brassica napus. Plant Biotechnol J, 2015, 14: 1368-1380[本文引用:2]
[14]
WuJ, ZhaoQ, YangQ, LiuH, LiQ, YiX, ChengY, GuoL, FanC, ZhouY. Comparative transcriptomic analysis uncovers the complex genetic network for resistance to Sclerotinia sclerotiorum in Brassica napus. Sci Rep, 2016, 6: 19007[本文引用:1]
[15]
Hoffman DD, Diers BW, Hartman GL, Nickell CD, Nelson RL, Pedersen WL, Cober ER, Graef GL, Steadman JR, Grau CR, Nelson B D, del Rio L E, Helms T, Anderson T, Poysa V, Rajcan I, Stienstra W C. Selected soybean plant introductions with partial resistance to Sclerotinia sclerotiorum. Plant Dis, 2002, 86: 971-980[本文引用:1]
[16]
Kim HS, Diers BW. Inheritance of partial resistance to Sclerotinia stem rot in soybean. Crop Sci, 2000, 40: 55-61[本文引用:1]
Boudet AM, LapierreC, Grima-PettenatiJ. Biochemistry and molecular biology of lignification. New Phytologist, 1995, 129: 203-236[本文引用:1]
[19]
Nicholson RL, HammerschmidtR. Phenolic compounds and their role in disease resistance. Annu Rev Phytopathol, 1992, 30: 369-389[本文引用:1]
[20]
Vance CP, Kirk TK, Sherwood RT. Lignification as a mechanism of disease resistance. Annu Rev Phytopathol, 1980, 18: 259-288[本文引用:1]
[21]
Dushnicky LG, Ballance GM, Sumner MJ, MacGregor A W. The role of lignification as a resistance mechanism in wheat to a toxin-producing isolate of Pyrenophora tritici-repentis. Can J Plant Pathol, 1998, 20: 35-47[本文引用:1]
[22]
HammerschmidtR, KućJ. Lignification as a mechanism for induced systemic resistance in cucumber. Physiol Plant Pathol, 1982, 20: 61-71[本文引用:1]
[23]
Southerton SG, Deverall BJ. Histochemical and chemical evidence for lignin accumulation during the expression of resistance to leaf rust fungi in wheat. Physiol Mol Plant Pathol, 1990, 36: 483-494[本文引用:1]
GayosoC, PomarF, Novo-UzalE, Merino F, de Ilárduya Ó M. The Ve-mediated resistance of the tomato to Verticillium dahliae involves H2O2, peroxidase and lignins and drives PAL gene expression. BMC Plant Biol, 2010, 10: 1[本文引用:2]
[26]
HammerschmidtR, Bonnen AM, Bergstrom GC, Baker KK. Association of epidermal lignification with nonhost resistance ofcucurbits to fungi. Can J Bot, 1985, 63: 2393-2398[本文引用:2]
[27]
PomarF, NovoM, Bernal MA, MerinoF, Barceló AR. Changes in stem lignins (monomer composition and crosslinking) and peroxidase are related with the maintenance of leaf photosynthetic integrity during Verticillium wilt in Capsicum annuum. New Phytol, 2004, 163: 111-123[本文引用:2]
[28]
Barceló AR. Lignification in plant cell walls. Int Rev Cytol, 1997, 176: 87-132[本文引用:2]
[29]
Snowdon RJ, WittkopB, RezaidadA, HasanM, LipsaF, SteinA, FriedtW. Regional association analysis delineates a sequenced chromosome region influencing antinutritive seed meal compounds in oilseed rape. Genome, 2010, 53: 917-928[本文引用:2]
[30]
LiuL, SteinA, WittkopB, SarvariP, LiJ, YanX, DreyerF, FrauenM, FriedtW, Snowdon RJ. A kockout mutation in the lignin biosynthesis gene CCR1 explains a major QTL for acid detergent lignin content in Brassica napus seeds. Theor Appl Genet, 2012, 124: 1573-1586[本文引用:2]
[31]
曲存民, 付福友, 卢坤, 谢景梅, 刘晓兰, 黄杰恒, 李波, 王瑞, 谌利, 唐章林, 李加纳. 不同环境中甘蓝型油菜种皮木质素含量的QTL定位. 作物学报, 2011, 37: 1398-1405Qu CM, Fu FY, LuK, Xie JM, Liu XL, Huang JH, LiB, WangR, ChenL, Tang ZL, Li JN. Identification of QTLs for lignin content of seed coat in Brassica napus L. in different environments. Acta Agron Sin, 2011, 37: 1398-1405 (in Chinese with English abstract)[本文引用:1]
[32]
Liu LZ, Qu CM, WittkopB, YiB, XiaoY, He HY, SnowdonR, LiJ. A high-density SNP map for accurate mapping of seed fibre QTL in Brassica napus L. PLoS One, 2013, 8: e83052[本文引用:3]
EynckC, Séguin-SwartzG, Clarke WE, Parkin IA. Monolignol biosynthesis is associated with resistance to Sclerotinia sclerotiorum in Camelina sativa. Mol Plant Pathol, 2012, 13: 887-899[本文引用:3]
[36]
McCouch SR, Cho YG, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14[本文引用:1]
[37]
LiJ, ZhaoZ, HaywardA, ChengH, FuD. Integration analysis of quantitative trait loci for resistance to Sclerotinia sclerotiorum in Brassica napus. Euphytica, 2015, 205: 483-489[本文引用:1]
[38]
Behla RS, Fernand o W G D, Li G. Identification of quantitative trait loci for resistance against Sclerotinia stem rot in Brassica napus. Can J Plant Pathol, 2009, 31: 477-478[本文引用:1]
[39]
Yin XR, YiB, ChenW, Zhang WJ, Tu JX, Fernand o W G D, Fu T D. Mapping of QTLs detected in a Brassica napus DH population for resistance to Sclerotinia sclerotiorum in multiple environments. Euphytica, 2010, 173: 25-35[本文引用:1]
[40]
WeiD, MeiJ, FuY, Disi JO, LiJ, QianW. Quantitative trait loci analyses for resistance to Sclerotinia sclerotiorum and flowering time in Brassica napus. Mol Breed, 2014, 34: 1797-1804[本文引用:1]
[41]
TuberosaR, SalviS, Sanguineti MC, Land iP, MaecaferriM, ContiS. Mapping QTLs regulating morpho-physiological traits and yield: case studies, shortcomings and perspectives in drought-stressed maize. Ann Bot, 2002, 89: 941-963[本文引用:1]
 , 荆凌云
, 荆凌云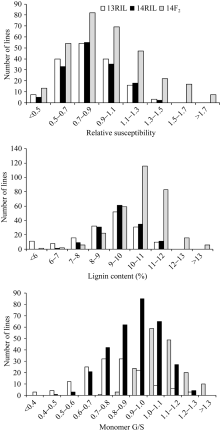


{kind=link}
{kind=link}
{kind=link}