关键词:RNA干扰; 寄主诱导的基因沉默(HIGS); 真菌病害; 转基因植物 Progress in Enhancement of Plant Resistance against Fungal Diseases through Host-Induced Gene Silencing CHENG Wei1,2, LI He-Ping2, HE Shui-Lin1, LIAO Yu-Cai2,* 1 Key Laboratory of Plant Genetic Improvement and Comprehensive Utilization, Ministry of Education / College of Crop Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2 Molecular Biotechnology Laboratory of Triticeae Crops / College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China
Fund:This research was supported by the National Major Project for Developing New GM Crops (2016ZX08002001-003), the National Natural Science Foundation of China (31601761), and the Research Project for Young Teachers of Fujian Province (JAT160180) AbstractHost-induced gene silencing (HIGS) uses the key genes that are involved in growth and development, sporulation and propagation, and pathogenesis of pathogens as targets, and expresses the RNAi constructs in host plants targeting to the target genes; during infection of plants, pathogens take up dsRNA or siRNA molecules, specifically bind to nucleotide sequences after recognition, and interfere the expression of the target genes, thereby inhibiting infection and spreading of invaded pathogens and thus conferring resistance phenotypes in plants. This technology serves as the solid foundation of plant resistance based on pathogen-specific sequences and has great potential for application in crop improvement against fungal diseases. In this review, recent advances in methodologies, technological routes and application of the HIGS strategy in plants against invading fungal pathogens are summarized; prospects for the future in HIGS-based plant resistance to fungal pathogens were also discussed.
Keyword:RNA interfering; Host-induced gene silencing (HIGS); Fungal disease; Transgenic plants Show Figures Show Figures
Fire AZ. Gene silencing by double-strand ed RNA. Cell Death Differ, 2007, 14: 1998-2012[本文引用:1]
[2]
Ketting RF. The many faces of RNAi. Dev Cell, 2011, 20: 148-161[本文引用:1]
[3]
Nunes CC, Dean AR. Host-induced gene silencing: A tool for understand ing fungal host interaction and for developing novel disease control strategies. Mol Plant Pathol, 2012, 13: 519-529[本文引用:2]
[4]
KochA, Kogel KH. New wind in the sails: improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol J, 2014, 12: 821-831[本文引用:1]
[5]
PanwarV, McCallum B, Bakkeren G. A functional genomics method for assaying gene function in phytopathogenic fungi through host-induced gene silencing mediated by agroinfiltration. Methods Mol Biol, 2015, 1287: 179-189[本文引用:1]
Tinoco ML, Dias BB, Dall’Astta R C, Pamphile J A, Aragao F J. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-strand ed RNA. BMC Biol, 2010, 8: 27[本文引用:1]
[8]
Melnyk CW, MolnarA, Baulcombe DC. Intercellular and systemic movement of RNA silencing signals. EMBO J, 2011, 30: 3553-3563[本文引用:1]
[9]
NishkuraK. A short primer on RNAi: RNA-directed RNA polymerase acts as a key catalyst. Cell, 2001, 107: 415-418[本文引用:1]
[10]
Vaistij FE, JonesL, Baulcombe DC. Spreading of RNA targeting and DNA methylation in RNA silencing requires transcription of the target gene and a putative RNA dependent RNA polymerase. Plant Cell, 2002, 14: 857-867[本文引用:1]
[11]
WasseneggerM, KrczalG. Nomenclature and functions of RNA-directed RNA polymerases. Trends Plant Sci, 2006, 11: 142-151[本文引用:1]
[12]
PliegoC, NowaraD, BoncianiG, Gheorghe DM, XuR, SuranaP, WhighamE, NettletonD, Bogdanove AJ, Wise RP, SchweizerP, Bindschedler LV, Spanu PD. Host-induced gene silencing in barley powdery mildew reveals a class of ribonuclease-like effectors. Mol Plant Microbe Interact, 2013, 26: 633-642[本文引用:1]
[13]
ZhangH, GuoJ, Voegele RT, ZhangJ, DuanY, LuoH, KangZ. Functional characterization of calcineurin homologs PsCNA1/ PsCNB1 in Puccinia striiformis f. sp. tritici using a host-induced RNAi system. PLoS One, 2012, 7: e49262[本文引用:1]
[14]
PanwarV, McCallum B, Bakkeren G. Endogenous silencing of Puccinia triticina pathogenicity genes through in planta- expressed sequences leads to suppression of rust diseases on wheat. Plant J, 2013, 73: 521-532[本文引用:1]
[15]
PanwarV, McCallum B, Bakkeren G. Host-induced gene silencing of wheat leaf rust fungus Puccinia triticina pathogenicity genes mediated by the Barley stripe mosaic virus. Plant Mol Biol, 2013, 81: 595-608[本文引用:1]
[16]
KochA, KumarN, WeberL, KellerH, ImaniJ, Kogel KH. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc Natl Acad Sci USA, 2013, 110: 19324-19329[本文引用:1]
[17]
Ghag SB, Shekhawat UK, Ganapathi TR. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium wilt in banana. Plant Biotechnol J, 2014, 12: 541-553[本文引用:1]
[18]
ChengW, Song XS, Li HP, Cao LH, SunK, Qiu XL, Xu YB, YangP, HuangT, Zhang JB, QuB, Liao YC. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol J, 2015, 13: 1335-1345[本文引用:1]
WangM, WeibergA, Lin FM, Thomma BP, Huang HD, JinH. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat Plants, 2016, 2: 16151[本文引用:3]
[21]
Zhang MX, Wang QH, XuK, Meng YL, Quan JL, Shan WX. Production of dsRNA sequences in the host plant is not sufficient to initiate gene silencing in the colonizing oomycete pathogen Phytophthora parasitica. PLoS One, 2011, 6: e28114[本文引用:1]
YinC, Jurgenson JE, Hulbert SH. Development of a host- induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f sp. tritici. Mol Plant Microbe Interact, 2011, 24: 554-561[本文引用:2]
[24]
Franco-Zorrilla JM, ValliA, TodescoM, MateosI, Puga MI, Rubio-SomozaI, LeyvaA, WeigelD, García JA, Paz-AresJ. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat Genet, 2007, 39: 1033-1037[本文引用:1]
[25]
TodescoM, Rubio-SomozaI, Paz-AresJ, WeigelD. A collection of target mimics for comprehensive analysis of microRNA function in Arabidopsis thaliana. PLoS Genet, 2010, 6: e1001031[本文引用:1]
[26]
MengY, ShaoC, WangH, JinY. Target mimics: an embedded layer of microRNA-involved gene regulatory networks in plants. BMC Genomics, 2012, 13: 197[本文引用:1]
[27]
WeibergA, WangM, Lin FM, ZhaoH, ZhangZ, KaloshianI, Huang HD, JinH. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science, 2013, 342: 118-123[本文引用:2]
[28]
KochA, BiedenkopfD, FurchA, WeberL, RossbachO, AbdellatefE, LinicusL, JohannsmeierJ, JelonekL, GoesmannA, CardozaV, McMillan J, Mentzel T, Kogel K H. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog, 2016, 12: e1005901[本文引用:1]
 , 李和平
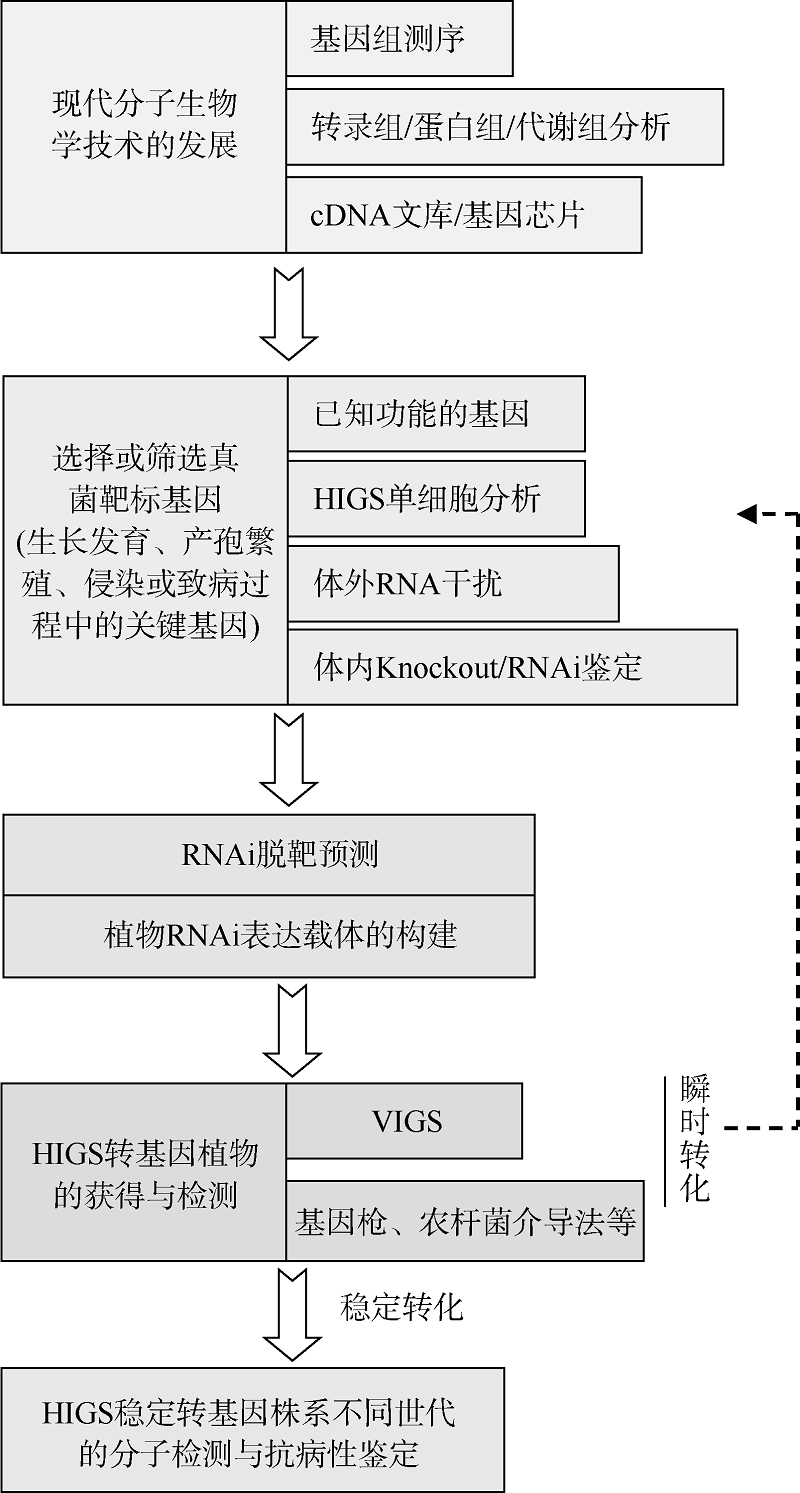
, 李和平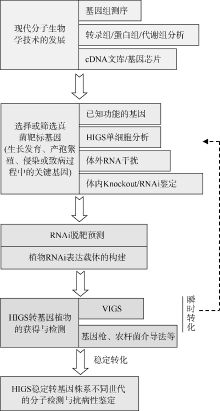


{kind=link}