关键词:水稻; 温度敏感型; 遗传分析; 叶色; 基因定位 Phenotypes and Gene Mapping of a Thermo-sensitive Yellow Leaf Mutant of Rice ZHANG Tian-Yu1, ZHOU Chun-Lei1, LIU Xi1, SUN Ai-Ling1, CAO Peng-Hui1, Thanhliem NGUYEN1, TIAN Yun-Lu1, ZHAI Hu-Qu1,2, JIANG Ling1,* 1State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology, Genetics and Breeding of Japonica Rice in Mid-lower Yangtze River, Ministry of Agriculture / Research Center of Jiangsu Plant Gene Engineering, Nanjing Agricultural University, Nanjing 210095, China
2Chinese Academy of Agricultural Sciences, Beijing 100081, China
Fund:This study was supported by the National Key Research and Development Program of China (2016YFD0100101-08), the Science and Technology Support Project of Jiangsu Province (BE2015363), the Independent Innovation Project for Agricultural Science and Technology of Jiangsu Province [CX(16)1029], and the Science and Technology Major Projects of Anhui Province (16030701068). AbstractPhenotypic analysis and gene mapping of rice leaf color could lay a foundation for map-based cloning of related genes and the function research of rice photosynthetic system. Leaf yellow mutant dy1 was obtained from rice cultivar “Nanjing 11” (abbreviated as NJ11) mutated by ethyl methanesulfonate (EMS). The mutant dy1 showed leaf yellowing at seedling stage and maturity stage in natural environment, with an abnormal structure of thylakoids under TEM, and significant differences in plant height, tiller number, seed setting rate and so on. The mutant dy1 showed albinism in 20°C, etiolation in 25°C and virescence in 30°C. The rice leaf yellow mutant dy1 was controlled by a single recessive gene. F2 population was constructed by crossing the mutant with “02428”, and the mutant phenotypes of extreme individuals were selected to map gene dy1. The gene was located in the 115 kb region of the long arm of chromosome 1, and contained 16 ORFs. Sequencing analysis showed that LOC_Os01g73450 controlling a uracil nucleotide kinase, with a single base substitution in the junction of the fourth intron and the fifth exon in dy1, might be a candidate gene. And the expression of genes related to chloroplast synthesis was significantly decreased, indicating dy1 may involve in the chloroplast synthesis
Keyword:Rice; Thermo-sensitive; Genetic analysis; Leaf color; Gene mapping Show Figures Show Figures
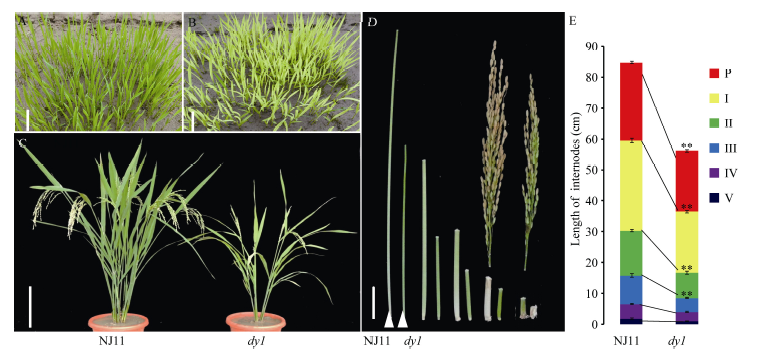
图1 自然条件下NJ11和dy1在苗期和灌浆期的表型 A, B: 在南京土桥自然条件下播种2周NJ11 (A)和dy1 (B)的幼苗表现。C: NJ11和dy1在灌浆期植株表现, 比例尺为15 cm。D, E: 在南京土桥自然条件下至成熟期NJ11 (A)和dy1 (B)主穗各节间长度, 比例尺为3 cm。* * 表示在P=0.01水平上差异极显著。Fig. 1 Phenotypes of NJ11 and dy1at seedling and mature stages under natural condition A, B: Phenotypes of NJ11 (A) and dy1(B) in the field at two weeks after seeding. C: phenotypes of NJ11 and dy1 at tilling period. Bar = 15 cm. D, E: the internode length of NJ11 and dy1. Bar = 3 cm. * * significantly different at P=0.01.
表2 Table 2 表2(Table 2)
表2 野生型NJ11和突变体dy1的农艺性状 Table 2 Agronomic traits of the wild type NJ11 and dy1 mutant
性状Trait
NJ11
dy1
P-value
株高Plant height (cm)
87.4± 3.72
53.9± 2.63* *
1.28× 10-11
分蘖数Number of tillers
8.92± 2.15
4.15± 1.07* *
1.91× 10-8
穗长Panicle length (cm)
25.5± 1.7
21.4± 2.2* *
1.38× 10-3
每穗粒数Grains per panicle
147.6± 13.0
118.5± 17.9* *
1.78× 10-2
结实率 Seed setting mate (%)
88.2± 2.4
67.4± 3.2* *
1.91× 10-8
千粒重 1000-grain weight (g)
23.6± 1.01
23.4± 0.68
0.17
粒长 Grain length (mm)
7.58± 0.40
7.40± 0.38
0.13
粒宽 Grain width (mm)
2.78± 0.19
2.92± 0.22
0.15
Values are means± SD (n=20, except for 1000-grain weight n=3). Significance analyses were conducted by t-test. * * Significantly different at P=0.01. 除千粒重n=3, 其余各项n=20, 取平均值; 显著性分析采用t测验。* * 表示在P=0.01水平上差异极显著。
表2 野生型NJ11和突变体dy1的农艺性状 Table 2 Agronomic traits of the wild type NJ11 and dy1 mutant
图2 3种温度下野生型NJ11和突变体dy1表型Fig. 2 Phenotypes of NJ11 and dy1 under three temperature conditions
表3 Table 3 表3(Table 3)
表3 3种温度下野生型NJ11和dy1三叶期幼苗叶片中叶绿素含量 Table 3 Chlorophyll contents in leaves of dy1 and NJ11 seedlings at 3-leaf stage under the three temperature conditions
叶绿素含量 Chlorophyll content
30° C
25° C
20° C
NJ11
dy1
NJ11
dy1
NJ11
dy1
叶绿素a含量 Content of chlorophyll a (mg g-1)
0.87± 0.034
0.69± 0.03* *
0.82± 0.01
0.44± 0.01* *
0.78± 0.03
0.24± 0.03* *
叶绿素b含量 Content of chlorophyll b (mg g-1)
0.37± 0.03
0.28± 0.02* *
0.34± 0.04
0.19± 0.04* *
0.32± 0.06
0.10± 0.01* *
* * 表示在P=0.01水平上差异极显著。* * Significantly different at P=0.01. WT: wild type.
表3 3种温度下野生型NJ11和dy1三叶期幼苗叶片中叶绿素含量 Table 3 Chlorophyll contents in leaves of dy1 and NJ11 seedlings at 3-leaf stage under the three temperature conditions
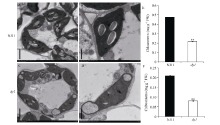
图3 NJ11和dy1剑叶叶绿体超微结构和叶绿素含量 A, B: NJ11叶绿体超微结构; C, D: dy1叶绿体超微结构, 比例尺(A, C) =2 μ m, 比例尺(B, D) =1 μ m; E, F: NJ11和dy1叶绿素a和叶绿素b含量。* * 表示在P=0.01水平上差异极显著。Fig. 3 TEM images of chloroplast structure and the chlorophyll contents of the flag leaf in NJ11 and dy1 A, B, C, D: TEM images of chloroplast structure in NJ11 (A, B) and dy1 (C, D), Bar (A, C) =2 μ m, Bar (B, D) = 1 μ m; E, F: the chlorophyll a and chlorophyll b contents of NJ11 and dy1. * * Significantly different at P=0.01.
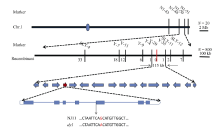
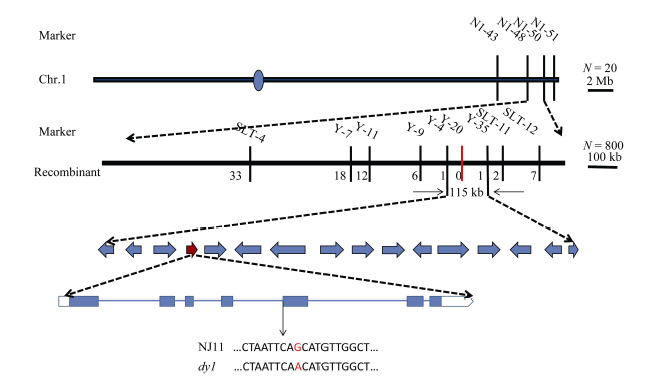
图4 水稻叶色基因在第1染色体的精细定位及候选基因 蓝色圆圈表示着丝粒; 红色竖线表示共分离标记; 红色箭头表示候选基因; 红色字母表示测序差异。Fig. 4 Fine mapping of the putative gene controlling leaf color on chromosome 1 in rice The blue circle means the centromere; the red vertical line means a co-segregate marker; the red arrow means the candidate gene; the red letter means sequencing difference.
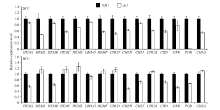
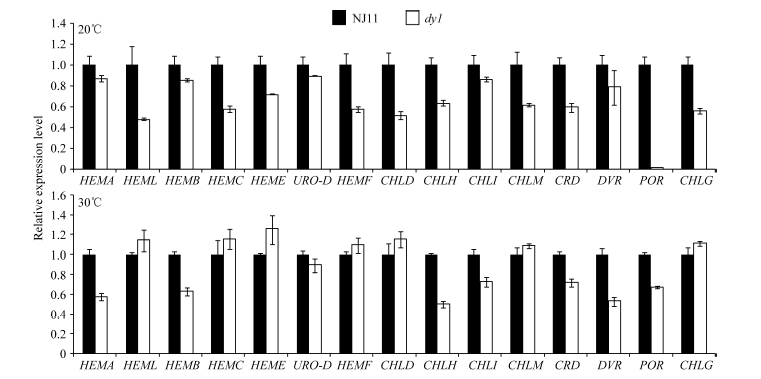
图5 两种温度下NJ11和dy1幼苗中第2叶叶绿素合成相关基因的相对表达水平Fig. 5 Relative expression levels of genes involved in chlorophyll biosynthesis in the second leaf of NJ11 and dy1 seedlings under two temperature conditions
LeisterD. Chloroplast research in the genomic age. Trends Genet, 2003, 19: 47-56[本文引用:1]
[2]
Afsar AwanM, Konzak CF, Rutger JN, Nilan RA. Mutagenic effects of sodium azide in rice. Crop Sci, 1980, 76: 663-668[本文引用:1]
[3]
胡忠, 彭丽萍, 蔡永华. 一个黄绿色的水稻细胞核突变体. 遗传学报, 1981, 8: 256-261HuZ, Peng LP, Cai YH. A yellow rice mutants of nuclei. Acta Genet Sin, 1981, 8: 256-261 (in Chinese)[本文引用:1]
[4]
陈青, 卢芙萍, 徐雪莲. 水稻叶色突变体研究进展. 热带生物学报, 2010, 1: 269-281ChenQ, Lu FP, Xu XL. Progress in research of rice leaf coloration mutant. J Tropical Agric, 2010, 1: 269-281 (in Chinese)[本文引用:1]
[5]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
[6]
Yang YL, XuJ, Huang LC, Leng YJ, Rao YC, ChenL, Wang QQ, Tu ZJ, HuJ, Ren DY, Zhang GH, ZhuL, Guo LB, QianQ, Zeng DL. PGL, encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice. J Exp Bot, 2007, 67: 297-310[本文引用:1]
[7]
Zhang HT, Li JJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Peak NC. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1]
[8]
Zhao CF, Xu JM, ChenY, Mao CZ, Zhang SL, Bai YH, JiangD, WuP. Molecular cloning and characterization of OsCHR4, a rice chromatin-remodeling factor required for early chloroplast development in adaxial mesophyll. Planta, 2012, 236: 1165-1176[本文引用:1]
[9]
Yoo SC, Cho SH, SugimotoH, Li JJ, KusumiK, Koh HJ, IbaK, Peak NC. Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol, 2009, 150: 388-401[本文引用:1]
[10]
Park SY, Yu JW, Park JS, Li JJ, Yoo SC, Lee SK, Lee NY, Jeong SW, Jeong SW, Seo HS, Koh HJ, Jeon JS, Park YL, Peak NC. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1]
[11]
HartmutK, Lichtenthaler. Biosynthesis, accumulation and emission of carotenoids, α-tocopherol, plastoquinone, and isoprene in leaves under high photosynthetic irradiance. Photosynth Res, 2007, 92: 163-179[本文引用:1]
[12]
刘喜, 周春雷, 任雅琨, 杨春艳, 何旎清, 柳周, 江玲, 万建民. 水稻叶色白化转绿突变体 WGL的遗传分析与基因定位. 南京农业大学学报, 2015, 38: 712-719LiuX, Zhou CL, Ren YK, Yang CY, He NQ, LiuZ, JiangL, Wan JM. Genetic analysis and gene mapping of virescent albino leaf mutant WGL in rice. J Nanjing Agric Univ, 2015, 38: 712-719 (in Chinese with English abstract)[本文引用:2]
[13]
何旎清, 柳周, 张龙, 白苏阳, 田云录, 江玲, 万建民. 一个新的水稻黄绿叶突变体的遗传分析及突变基因的精细定位. 作物学报, 2015, 41: 1155-1163He NQ, LiuZ, ZhangL, Bai SY, Tian YL, JiangL, Wan JM. Genetic analysis of a new yellow-green leaf mutant and fine-mapping of mutant gene in rice. Acta Agron Sin, 2015, 41: 1155-1163 (in Chinese with English abstract)[本文引用:1]
[14]
孔飞, 蔡跃, 汪鹏, 尤小满, 张杰, 江玲, 张文伟, 万建民. 水稻温度敏感型黄叶突变体 yl2(t)的表型分析和基因定位. 南京农业大学学报, 2016, 39: 703-710KongF, CaiY, WangP, You XM, ZhangJ, JiangL, Zhang WW, Wan JM. Characterization and gene mapping of a thermo- sensitive yellow leaf mutant yl2(t)in rice. J Nanjing Agric Univ, 2016, 39: 703-710 (in Chinese with English abstract)[本文引用:2]
[15]
刘艳霞, 林冬枝, 董彦君. 水稻温敏感叶色突变体研究进展. 中国水稻科学, 2015, 29: 439-446Liu YX, Lin DZ, Dong YJ. Research advances in thermo- sensitive leaf coloration mutants in rice. Chin J Rice Sci, 2015, 29: 439-446 (in Chinese with English abstract)[本文引用:1]
[16]
KensukeK, ShokoH, HiroshiS, YokoC, OsanuM, KohI. Contribution of chloroplast biogenesis to carbon-nitrogen balance during early leaf development in rice. J Plant Res, 2010, 123: 617-622[本文引用:1]
[17]
HirokiS, KensukeK, KoN, MasahiroY, AtsushiY, KohI. The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria. Plant J, 2007, 52: 512-527[本文引用:2]
[18]
Gong XD, Su QQ, Lin DZ, JiangQ, Xu JL, Zhang JH, TengS, Dong YJ. The rice OsV4encoding a novel pentatricopeptide repeat protein is required for chloroplast development during the early leaf stage under cold stress. J Integr Plant Biol, 2014, 56: 400-410[本文引用:2]
[19]
JiangQ, MeiJ, Gong XD, Xu JL, Zhang JH, TengS, Lin DZ, Dong Y J. Importance of the rice TCD9 encoding subunit of chaperonin protein 60 (Cpn60) for the chloroplast development during the early leaf stage. Plant Sci#/magtechI#, 2014, 215/216: 172-179[本文引用:1]
[20]
SongJ, Wei XJ, Shao GN, Sheng ZH, Chen DB, Liu CL, Jiao GA, Xie LL, Tang SQ, Hu PS. The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions. Plant Mol Biol, 2014, 84: 301-314[本文引用:2]
[21]
KensukeK, ChikakoS, TakahiroN, ShinjiK, AtsushiY, KohI. A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions. Plant J, 2011, 68: 1039-1050[本文引用:1]
[22]
TodaT, FujiS, NoguchiK, TomohikoK, KinyaT. Rice MPR25 encodes a pentatricopeptide repeat protein and is essential for RNA editing of nad5 transcripts in mitochondria. Plant J, 2012, 72: 450-460[本文引用:1]
[23]
Peng LW, FukaoY, MyougaF, MotohashiR, ShinozakiK, ShikanaiT. A chaperonin subunit with unique structures is essential for folding of a specific substrate. PLoS Biol, 2011, 9: 1-13[本文引用:1]
[24]
ZhouL, LacrouteF, ThornburgR. Cloning, expression in Escherichia coli, and characterization of Arabidopsis thaliana UMP/CMP kinase. Plant Physiol, 1998, 117: 245-254[本文引用:1]
[25]
HeinP, StockelJ, BennewitzS, OelmullerR. A protein related to prokaryotic UMP kinases is involved in psaA/B transcript accumulation in Arabidopsis. Plant Mol Biol, 2009, 69: 517-528[本文引用:1]
 , 周春雷
, 周春雷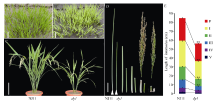




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}