关键词:β; 淀粉酶; 茶树; 基因克隆; 低温胁迫; 表达调控 Cloning of β-amylase Gene ( CsBAM3) and Its Expression Model Response to Cold Stress in Tea Plant HAO Xin-Yuan1,**, YUE Chuan1,2,**, TANG Hu1, QIAN Wen-Jun1, WANG Yu-Chun1, WANG Lu1, WANG Xin-Chao1,*, YANG Ya-Jun1,* 1 Tea Research Institute, Chinese Academy of Agricultural Sciences / National Center for Tea Improvement / Key Laboratory of Tea Plant Biology and Resources Utilization, Ministry of Agriculture, Hangzhou 310008, China
2 College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002
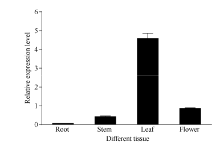
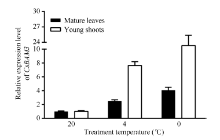
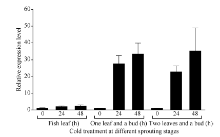
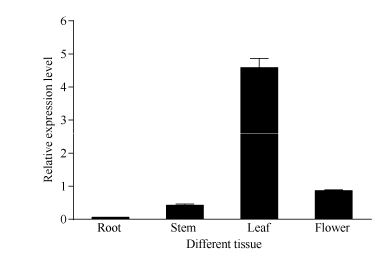
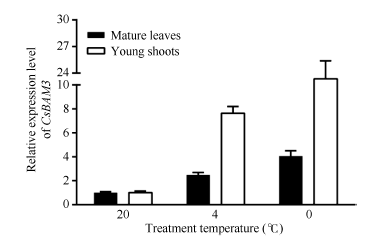
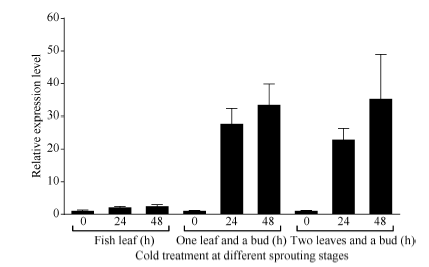
Fund:This study was supported by the Natural Science Foundation of Zhejiang Province (LY16C160001), the China Agriculture Research System (CARS-19), and the Chinese Academy of Agricultural Sciences through an Innovation Project for Agricultural Sciences and Technology (CAAS-ASTIP-2017-TRICAAS). AbstractA major enzyme of starch breakdown in plant leaf, β-amylase (BAM), plays important roles in response to abiotic stresses. In tea plant, a differentially expressed gene involved in starch metabolism was identified from previous RNA-Seq analysis of cold acclimation. Its full-length cDNA was cloned and sequenced, showing that it is a homolog of BAM3 in Arabidopsis (named CsBAM3). CsBAM3 contains 548 amino acid residues and is grouped into subfamily II together with BAM1 and BAM3 from Arabidopsis. According to protein sequence and 3D structure analyses, CsBAM3 was considered as a chloroplast-targeted protein with β-amylase activity. Promoter cloning and sequence analysis showed a couple of responsive elements related to circadian, light, cold, and phytohormone, indicating its complex regulating mechanism. CsBAM3 showed the expression higher in leaf, relative lower in stem and flower and extremely low in root. The expression of CsBAM3 in mature leaf was significantly up-regulated during the early time courses of winter cold acclimation and maintained at relative high level during the following period. With the treatments of 4°C and 0°C, the transcript levels of CsBAM3 in both mature leaves and young shoots were dramatically increased, interestingly, much higher level was detected in young shoots. The cold environment similar to spring cold spell was created using climate chamber. And the expression changes of CsBAM3 in shoots were determined then after the tea plants were moved into climatic chamber at different sprouting phrases. CsBAM3 was quickly up-regulated by cold once the first leaf unfolded. Our study illuminated that CsBAM3 is a key β-amylase coding gene involved in starch metabolism of tea plant, which can be stimulated in both mature leaves and young shoots by cold stress.
Keyword:β; -amylase; Tea plant; Gene clone; Cold stress; Expression regulation Show Figures Show Figures
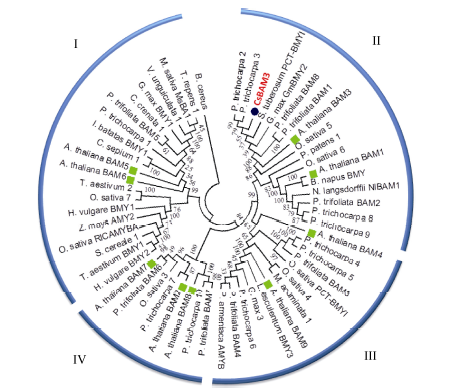
图2 CsBAM3与其他物种中BAMs的系统发育树分析 BAMs被聚为I、II、III、IV亚家族。CsBAM3用红色字体及蓝色实心圆标出, 拟南芥中BAMs用绿色实心方块儿标出。Fig. 2 Phylogeny of CsBAM3 and BAMs from other species Plant BAM proteins fall into four subfamilies (I, II, III, IV). Red and blue filled circle mean CaBAM3; green solid squares for BAMs from Arabidopsis.
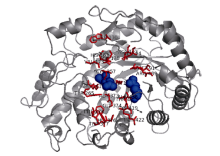
图3 CsBAM3的3D结构预测 与底物结合的作用位点用红色表示, 蓝色圆球为具有催化功能的Glu186和Glu380残基。Fig. 3 3D structure prediction of CsBAM3 Red sticks indicate substrate binding residues. Blue dots indicate the catalytic residues of Glu186 and Glu380.
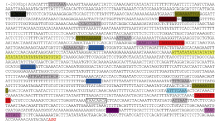
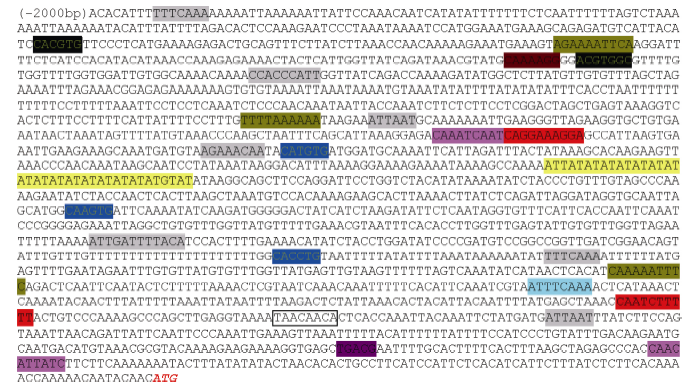
图4CsBAM3启动子序列及包含的顺式作用元件Fig. 4 Sequence andcis-acting element information of CsBAM3 promoter
表2 Table 2 表2(Table 2)
表2CsBAM3启动子区顺式作用元件功能注释 Table 2 Functional annotation of the cis-acting elements in CsBAM3 promoter region
序列 Sequence
位点名称 Site name
功能 Function
CAANNNNATC
Circadian
cis-acting regulatory element involved in circadian control
TGACG
TGACG-motif
cis-acting regulatory element involved in the MeJA-responsiveness
CAGAAAAGGA, CCATCTTTTT
TCA-element
cis-acting element involved in salicylic acid responsiveness
TATATATATATATATATATATA
TA-rich region
Enhancer
AAAAAATTTC, AGAAAATTCG
HSE
cis-acting element involved in heat stress responsiveness
ATTTCAAA
ERE
Ethylene-responsive element
TTTCAAA
Box I
Light responsive element
ATTAATTTTACA
AT1-motif
Part of a light responsive module
ATTAAT
Box 4
Part of a conserved DNA module involved in light responsiveness
AGAAACAA
AE-box
Part of a module for light response
CC(G/A)CCC
Sp1
Light responsive element
CCTTTTG
P-box
Gibberellin-responsive element
CACGTG, ACGTGGC
ABRE
cis-acting element involved in the abscisic acid responsiveness
TAACACA
Amylase box
Conserved sequence found in 5'-upstream region of alpha-amylase gene of rice, wheat, and barley
CANNTG
MYCCONSENSUSAT
MYC recognition site found in the promoters of the dehydration- responsive gene rd22 and many other genes in Arabidopsis; MYC recognition sequence in CBF3 promoter; Binding site of ICE1 (inducer of CBF expression 1) that regulates the transcription of CBF/DREB1 genes in the cold in Arabidopsis
表2CsBAM3启动子区顺式作用元件功能注释 Table 2 Functional annotation of the cis-acting elements in CsBAM3 promoter region
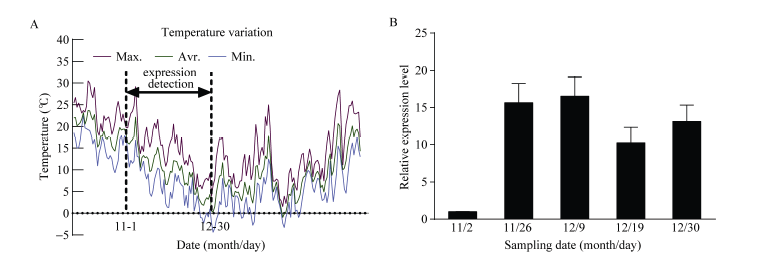
图6 温度变化及CsBAM3在冷驯化阶段的表达模式分析 A: 采样期间10月至第2年3月份每日温度变化, 其中虚线间为11月和12月份冷驯化阶段温度变化。 B: 冷驯化阶段CsBAM3基因的表达变化。Fig. 6 Temperature information and expression level of CsBAM3 during cold acclimation periods A: daily temperature from October to the next March, in which the temperature during cold acclimation (November and December) was marked between dotted lines. B: expression changes of CsBAM3 during cold acclimation.
蒋跃林, 李倬. 我国茶树栽培界限的气候划分. 生态农业研究, 2000, 8(1): 89-92Jiang YL, LiZ. Climate divison of tea tree cultivated limit in China. Eco-Agric Res, 2000, 8(1): 89-92 (in Chinese with English abstract)[本文引用:1]
[2]
郭湘, 唐茜, 许燕, 陈玖琳, 王自琴. 早春霜冻对不同茶树品种芽叶的生化成分及制茶品质的影响. 云南大学学报, 2015, 37: 930-938GuoX, TangQ, XuY, Chen JL, Wang ZQ. Effect of frost in early spring on biochemical composition and sensory qulity of new shoots in different tea cultivars. J Yunnan Univ, 2015, 37: 930-938 (in Chinese with English abstract)[本文引用:2]
[3]
Wang XC, Zhao QY, Ma CL, Zhang ZH, Cao HL, Kong YM, YueC, Hao XY, ChenL, Ma JQ, Jin JQ, LiX, Yang YJ. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom, 2013, 14: 415[本文引用:4]
[4]
BeninaM, ObataT, MehterovN, IvanovI, PetrovV, TonevaV, Fernie AR, Gechev TS. Comparative metabolic profiling of Haberlea rhodopensis, Thellungiella halophyla, and Arabidopsis thalianaexposed to low temperature. Front Plant Sci, 2013, 4: 499[本文引用:1]
[5]
Lee JH, Yu DJ, Kim SJ, ChoiD, Lee HJ. Intraspecies differences in cold hardiness, carbohydrate content and beta-amylase gene expression of Vaccinium corymbosum during cold acclimation and deacclimation. Tree Physiol, 2012, 32: 1533-1540[本文引用:1]
[6]
JanskaA, AprileA, ZamecnikJ, CattivelliL, OvesnaJ. Transcriptional responses of winter barley to cold indicate nucleosome remodelling as a specific feature of crown tissues. Funct Integr Genom, 2011, 11: 307-325[本文引用:1]
[7]
Gupta AK, KaurN. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J Biosci, 2005, 30: 761-776[本文引用:2]
[8]
Baena-GonzalezE, SheenJ. Convergent energy and stress signaling. Trends Plant Sci, 2008, 13: 474-482[本文引用:1]
[9]
SmeekensS, MaJ, HansonJ, Rolland F. Sugar signals and molecular networks controlling plant growth. Curr Opin Plant Biol, 2010, 13: 274-279[本文引用:1]
[10]
StittM, Zeeman SC. Starch turnover: pathways, regulation and role in growth. Curr Opin Plant Biol, 2012, 15: 282-292[本文引用:2]
[11]
Yu TS, Zeeman SC, ThorneycroftD, Fulton DC, DunstanH, Lue WL, HegemannB, Tung SY, UmemotoT, ChappleA, Tsai DL, Wang SM, Smith AM, ChenJ, Smith SM. Alpha-amylase is not required for breakdown of transitory starch in Arabidopsis leaves. J Biol Chem, 2005, 280: 9773-9779[本文引用:1]
[12]
YueC, Cao HL, WangL, Zhou YH, Huang YT, Hao XY, Wang YC, WangB, Yang Y J. Wang X C. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season. Plant Mol Biol, 2015, 88: 591-608[本文引用:1]
[13]
Fulton DC, StettlerM, MettlerT, Vaughan CK, LiJ, FranciscoP, GilM, ReinholdH, EickeS, MesserliG, DorkenG, HallidayK, Smith AM, Smith SM, Zeeman SC. Beta-Amylase 4, a noncatalytic protein required for starch breakdown, acts upstream of three active beta-amylases in Arabidopsischloroplasts. Plant Cell, 2008, 20: 1040-1058[本文引用:5]
[14]
LiJ, FranciscoP, ZhouW, EdnerC, SteupM, RitteG, Bond CS, Smith SM. Catalytically-inactive beta-amylase BAM4 required for starch breakdown in Arabidopsisleaves is a starch-binding- protein. Arch Biochem Biophys, 2009, 489: 92-98[本文引用:1]
[15]
Monroe JD, Storm AR, Badley EM, Lehman MD, Platt SM, Saunders LK, Schmitz JM, Torres CE. Beta-amylase 1 and beta-amylase 3 are plastidic starch hydrolases in Arabidopsis that seem to be adapted for different thermal, pH, and stress conditions. Plant Physiol, 2014, 166: 1748-1763[本文引用:2]
[16]
PengT, ZhuX, DuanN, Liu JH. PtrBAM1, a beta-amylase- coding gene of Poncirus trifoliata, is a CBF regulon member with function in cold tolerance by modulating soluble sugar levels. Plant Cell Environ, 2014, 37: 2754-2767[本文引用:5]
[17]
KaplanF, Guy CL. Beta-amylase induction and the protective role of maltose during temperature shock. Plant Physiol, 2004, 135: 1674-1684[本文引用:2]
[18]
FowlerS, Thomashow MF. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell, 2002. 14: 1675-1690[本文引用:2]
[19]
ReinholdH, SoykS, SimkovaK, HostettlerC, MarafinoJ, MainieroS, Vaughan CK, Monroe JD, Zeeman SC. Beta- amylase-like proteins function as transcription factors in Arabidopsis, controlling shoot growth and development. Plant Cell, 2011, 23: 1391-1403[本文引用:1]
[20]
丁菲. 低温胁迫下与茶树糖代谢相关基因的克隆与表达. 安徽农业大学硕士学位论文, 安徽合肥, 2012DingF. Cloning and Expression of Genes Related to Glyconmetabolism in Tea Plant ( Camellia sinensis (L. ) O. Kuntze) Under Low Temperature. MS Thesis of Anhui Agricultural University, Hefei, China, 2012 (in Chinese with English abstract)[本文引用:1]
[21]
岳川. 茶树糖类相关基因的挖掘及其在茶树冷驯化中的表达研究. 中国农业科学院博士学位论文, 北京, 2015YueC. Cloning and Expression Analysis of Sugar-Related Genes during Cold Acclimation in Tea Plant. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2015 (in Chinese with English abstract)[本文引用:1]
[22]
ChangS, PuryearJ, CairneyJ. A simple and efficient method for isolating RNA from pine trees. Plant Mol Biol Rep, 1993, 11: 113-116[本文引用:1]
[23]
HaoX, Horvath DP, Chao WS, YangY, WangX, XiaoB. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plant ( Camellia sinensis(L. ) O. Kuntze). Int J Mol Sci, 2014, 15: 22155-22172[本文引用:1]
[24]
彭婷. 枳叶片抑制差减杂交cDNA文库构建及 PtrBAM1基因抗寒功能鉴定. 华中农业大学博士学位论文, 湖北武汉, 2013PengT. Suppression Subtractive Hybridization cDNA Library Construction for Low Temperature-treated Poncirus trifoliata and Characterization of PtrBAM1. PhD Dissertation of Huazhong Agurcultural University, Wuhan, China, 2013 (in Chinese with English abstract)[本文引用:2]
[25]
SparlaF, CostaA, Lo SchiavoF, PupilloP, TrostP. Redox regulation of a novel plastid-targeted beta-amylase of Arabidopsis. Plant Physiol, 2006, 141: 840-850[本文引用:1]
[26]
KaplanF, Guy CL. RNA interference of Arabidopsisbeta-amylase 8 prevents maltose accumulation upon cold shock and increases sensitivity of PSII photochemical efficiency to freezing stress. Plant J, 2005, 44: 730-743[本文引用:3]
Davies P J. Plant Hormones. Netherland s: Springer-Verlag, XI, 2010, p 833[本文引用:1]
[29]
TakedaS, KowyamaY, TakeuchiY, MatsuokaK, NishimuraM, NakamuraK. Spatial patterns of sucrose-inducible and polygalacturonic acid-inducible expression of genes that encode sporamin and β-amylase in sweet potato: gene structure and expression. Plant Cell Physiol, 1995, 36: 321-333[本文引用:1]
[30]
SamojednyD, OrzechowskiS. New look at starch degradation in Arabidopsis thaliana L. chloroplasts. Postepy Biochem, 2007, 53: 74-83[本文引用:1]
[31]
ShinH, OhY, KimD. Differences in cold hardiness, carbohydrates, dehydrins and related gene expressions under an experimental deacclimation and reacclimation in Prunus persica. Physiol Plant, 2015, 154: 485-499[本文引用:1]
 , 岳川
, 岳川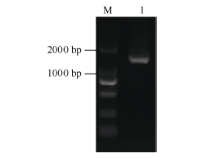




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}