关键词:小麦; 假禾谷镰孢; 生长素; 细胞分裂素; 脱落酸; 基因差异表达 Differential Expression of Three Plant Hormone Related Genes in Wheat Infected by Fusarium pseudograminearum LI Yong-Hui, CHEN Lin-Lin, SUN Bing-Jian, WANG Li-Min, XING Xiao-Ping, YUAN Hong-Xia, DING Sheng-Li*, LI Hong-Lian* Henan Agricultural University / National Key Laboratory of Wheat and Maize Crop Science / Collaborative Innovation Center of Henan Grain Crops, Zhengzhou 450002, China Fund:This study was supported by China Special Fund for Agro-scientific Research in the Public Interest (201503112) Abstract Fusarium pseudograminearum is a pathogenic fungus in wheat newly found in China. The purpose of this study was to disclose the responses of genes involved in the hormone signals pathway after F. pseudograminearum infection. Wheat cultivar Zhoumai 24 was infected by the wild-type strain WZ2-8A, and the seedling samples at five and fifteen days after inoculation (DAI) were subject to transcriptome analysis by RNA-seq method. The selected differentially expressed genes were subsequently validated by qRT-PCR assay. The infection of F. pseudograminearum had a negative impact on wheat growth with significantly decreased root length, seedling height, and fresh weights of root and shoot. A total of 29 genes expressed differentially in the signal transduction pathways of auxin, cytokinin (CTK) and abscisic acid (ABA). At 5 DAI, 11 genes were expressed differentially compared with the control group (without WZ2-8A inoculation), in which two and nine genes were up- and down-regulated, respectively. At 15 DAI, 25 genes were expressed differentially, including eight up-regulated and seventeen down-regulated genes. In the auxin signaling pathway, auxin input transporter AUX1 was differentially expressed, influencing polar transport of auxin and disturbing cell elongation of wheat roots. In the CTK signaling pathway, the up-regulating role of B-ARR suggests that it could facilitate CTK signal transduction to inhibit cellular division, resulting in weaker wheat growth along with the interaction of the auxin signaling pathway. In the ABA signaling pathway, ABA receptor PYR/PYL was down-regulated, while the negative regulator PP2C was up-regulated. ABA plays a negative role in regulating host resistance to fungi and bacteria and its transduction has the antagonistic action against the jasmine acid (JA) / ethylene (ET) pathway, i.e., the repression of ABA signaling pathway might result in the opening of JA/ET pathway. These results of transcriptome analysis were generally validated by the qRT-PCR evidence, indicating that inhibition of ABA signal transduction after F. pseudograminearum infection might be the physiological basis of moderate resistance to the fungus in Zhoumai 24.
Keyword:Wheat; Fusarium pseudograminearum; Auxin cytokinin; Abscisic acid; Gene differential expression Show Figures Show Figures
图1 小麦受假禾谷镰孢侵染后的症状及对植株生长的影响 A1和A2分别为接种后5 d和15 d的小麦。* 和* * 分别表示处理组(ZF)与对照组(ZC)之间有显著(P < 0.05)和极显著差异(P< 0.01)。Fig. 1 Symptom and effect on growth of wheat plant infected by F. pseudograminearum A1 and A2 show wheat growth at 5 days and 15 days after inoculation, respectively. * and * * indicate significant difference between the treatment group (ZF) and the control group (ZC) atP< 0.05 and P< 0.01, respectively.
表2 假禾谷镰孢侵染诱导后植物激素差异表达基因数 Table 2 Number of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum
植物激素 Plant hormone
转录因子 Transcription factor
ZF-5 vs. ZC
ZF-15 vs. ZC
ZF-15 vs. ZF-5
上调 Up-regulated
下调 Down-regulated
上调 Up-regulated
下调 Down-regulated
上调 Up-regulated
下调 Down-regulated
IAA
AUX1
1
1
0
0
0
0
SAUR
0
6
2
8
0
0
CTK
B-ARR
1
1
2
0
0
0
ABA
SnRK2
0
1
2
1
0
0
PYR/PYL
0
0
0
8
0
0
PP2C
0
0
2
0
0
0
总计 Total
2
9
8
17
0
0
ZF-5 and ZF-15 refer to the treatment group at 5 and 15 days after inoculation, respectively, and ZC refers to the control group. ZF-5和ZF-15分别表示处理组接种后5 d和15 d, ZC表示对照组。
表2 假禾谷镰孢侵染诱导后植物激素差异表达基因数 Table 2 Number of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum
细胞分裂素途径中的一个B-ARR基因Traes_7AS_503B57D77和脱落酸途径中的一个PP2C基因Traes_2AS_048E13951在接种后15 d差异表达倍数最大, 其log2FC可达到10以上; 而下调表达最明显的是1个SAUR基因Traes_5BL_80B9C8E10, 其log2FC值为-5.9, 其次是1个SAUR基因Traes_5DL_677150034和1个PYR/PYL基因Traes_1AL_B82F1573F, 它们的log2FC值均为-4.6 (表3)。 将各个差异表达基因表达量的FPKM (fragments per kilobase of transcript per million mapped reads)值取以10为底的对数(lg FPKM)制作热图能够直观看出这些基因的差异表达情况。图2显示, SAUR和PYR/PTL差异表达基因最多, 大部分为在接种5 d后下调表达的, 而在接种5 d和15 d后基因表达量变化不大。而B-ARR和PP2C的基因大部分上调表达。其中, B-ARR在接种后5 d至接种后15 d逐渐上调表达, PP2C的基因在接种后5 d不差异表达, 而在接种后15 d明显上调表达。 表3 Table 3 表3(Table 3)
表3 假禾谷镰孢侵染诱导后植物激素相关基因差异表达倍数 Table 3 Fold change (FC) of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum
植物激素 Plant hormone
转录因子 Transcription factor
基因代号 Gene code
log2 FC
ZF-5 vs. ZC
ZF-15 vs. ZC
ZF-15 vs. ZF-5
IAA
AUX1
Traes_5DS_0CBB9E7E6
4.7
—
—
Traes_1DL_F70395059
-3.2
—
—
SAUR
Traes_5BL_80B9C8E10
-3.8
-5.9
—
Traes_5DL_677150034
-3.5
-4.6
—
Traes_5DL_D5D75CCDF
-3.2
-3.6
—
Traes_5DL_32B83E57E
-3.2
-4.1
—
Traes_5DL_E0171FA9B
-3.1
-4.3
—
Traes_5BL_32E4F15BB
-2.8
-3.6
—
Traes_2DL_EC3448C9F
—
6.4
—
Traes_7AL_AEC178775
—
6.2
—
Traes_6AS_3470A0E8E
—
-4.5
—
Traes_5AL_2CDA282711
—
-4.4
—
CTK
B-ARR
Traes_7AS_503B57D77
8.8
10.0
—
Traes_3DL_D82CA7827
-1.8
—
—
Traes_7AS_DECB8080D
—
3.5
—
ABA
SnRK2
Traes_5DS_4ABE4DD6F
-1.5
—
—
Traes_2BL_616F08780
—
3.5
—
Traes_2AL_2FF604DA9
—
3.0
—
Traes_1BL_D20749C1D
—
-2.2
—
PYR/PYL
Traes_3DS_D7629B48F
—
-5.4
—
Traes_1AL_B82F1573F
—
-4.6
—
Traes_3AS_FF831326D
—
-4.2
—
Traes_3B_B2A7272D2
—
-3.9
—
Traes_4AS_72BEF89AC
—
-3.6
—
Traes_4DL_3A1814A74
—
-3.1
—
Traes_3B_6FEADCE61
—
-2.9
—
Traes_4BL_E43C1BB11
—
-2.7
—
PP2C
Traes_2AS_048E13951
—
10.1
—
Traes_2BS_C48CB635C
—
4.8
—
The log2 FC values represent changing folds of differentially expressed genes. The positive and negative log2 FC value indicates up- and down-regulation, respectively, and “ — ” indicates no significant changes (significant when log2 FC > ± 1 and P < 0.05). ZF-5 and ZF-15 refer to the treatment group at 5 and 15 days after inoculation, respectively, and ZC refers to the control group. log2 FC表示差异表达基因的差异倍数, 正、负值分别表示上调和下调, “ — ” 表示表达水平变化不显著(log2 FC > ± 1且P < 0.05为变化显著)。ZF-5和ZF-15分别表示处理组接种后5 d和15 d, ZC表示对照组。
表3 假禾谷镰孢侵染诱导后植物激素相关基因差异表达倍数 Table 3 Fold change (FC) of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum
图2 植物激素途径中差异表达基因表达量热图 不同颜色表示基因表达量(lg FPKM)。ZC:对照组; ZF-5:接种后5 d; ZF-15:接种后15 d。Fig. 2 Heatmap of differentially expressed genes involved in plant-hormone pathways Gene expression levels in lg FPKM represent in different colors. ZC:control group; ZF-5:5 days after inoculation; ZF-15:15 days after inoculation.
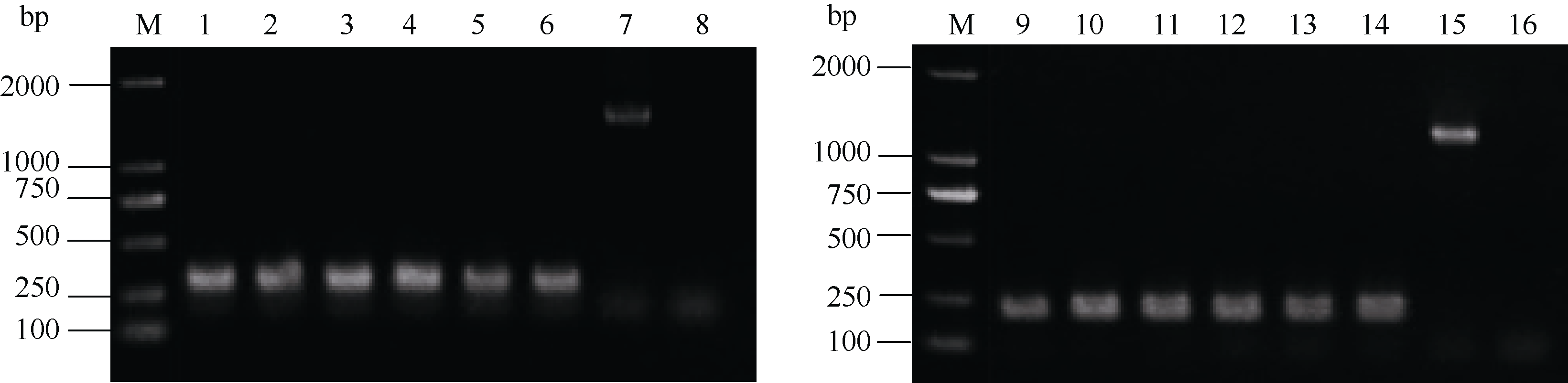
图3 cDNA中基因组DNA污染验证电泳图 M:DL2000 marker; 1~3:接种后5 d对照组; 4~6:接种后5 d处理组; 7和15:gDNA; 8和16:ddH2O; 9~11:接种后15 d对照组; 12~14:接种后15 d处理组。Fig. 3 Electrophoretic pattern of verification of genomic DNA contamination in cDNA pool M:DL2000 marker; 1-3:control group at 5 days after inoculation (DAI); 4-6:treatment group at 5 DAI; 7 and 15:gDNA; 8 and 16:ddH2O; 9-11:control group at 15 DAI; 12-14:treatment group at 15 DAI.
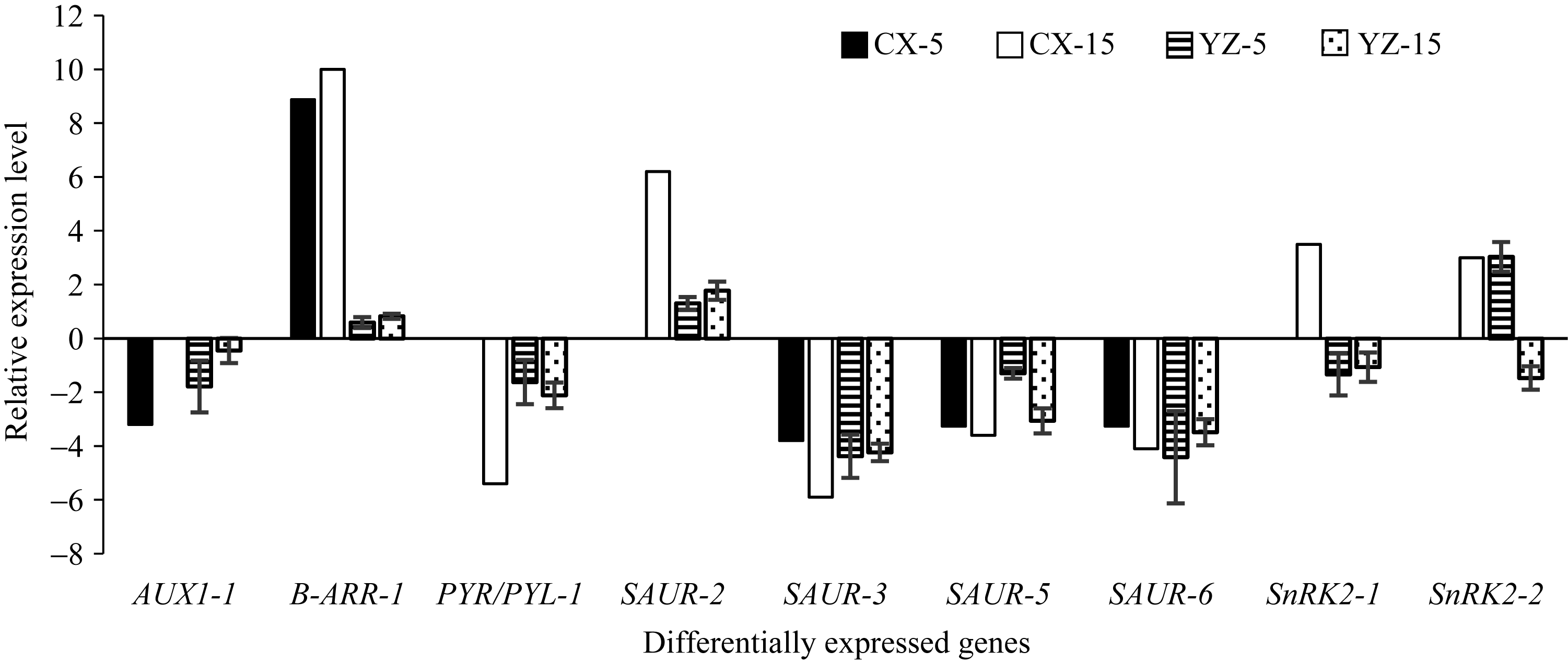
图4 差异表达基因qRT-PCR验证结果 CX-5和CX-15代表5 d和15 d测序样品; YZ-5和YZ-15代表5 d和15 d验证样品。Fig. 4 qRT-PCR validation for the differentially expressed genes CX-5 and CX-15 represent sequencing samples at 5 days after inoculation (DAI) and 15 DAI, respectively; YZ-5 and YZ-15 represent validation samples at 5 DAI and 15 DAI, respectively.
BackhouseD, AbubakarA, BurgessL, DenniscJ, HollawayG, WildermuthG, WallworkH, HenryF. Survey of Fusarium species associated with crown rot of wheat and barley in eastern Australia. Aust Plant Pathol, 2004, 33:255-261[本文引用:1]
[2]
贺小伦, 周海峰, 袁虹霞, 施艳, 孙炳剑, 李洪连. 河南和河北冬小麦区假禾谷镰孢的遗传多样性. 中国农业科学, 2016, 49:272-281He XL, Zhou HF, Yuan HX, ShiY, Sun BJ, Li HL. Genetic diversity of Fusarium pseudograminearum collected from Henan and Hebei winter wheat regions. Sci Agric Sin, 2016, 49:272-281 (in Chinese with English abstract)[本文引用:1]
[3]
张向向, 孙海燕, 李伟, 邓渊钰, 张爱香, 于汉寿, 陈怀谷. 我国冬小麦主产省小麦茎基腐镰孢菌的组成及其致病力. 麦类作物学报, 2014, 34:272-278Zhang XX, Sun HY, LiW, Deng YY, Zhang AX, Yu HS, Chen HG. Composition and pathogenicity of Fusarium crown rot pathogens of wheat in major winter wheat. J Triticeae Crops, 2014, 34:272-278 (in Chinese with English abstract)[本文引用:1]
[4]
周海峰, 杨云, 牛亚娟, 袁虹霞, 李洪连. 小麦茎基腐病的发生动态与防治技术. 河南农业科学, 2014, 43(5):114-117Zhou HF, YangY, Niu YJ, Yuan HX, Li HL. Occurrence and control methods of crown rot of wheat. J Henan Agric Sci, 2014, 43(5):114-117 (in Chinese with English abstract)[本文引用:1]
[5]
杨云, 贺小伦, 胡艳峰, 侯莹, 牛亚娟, 代君丽, 袁虹霞, 李洪连. 黄淮麦区主推小麦品种对假禾谷镰刀菌所致茎基腐病的抗性. 麦类作物学报, 2015, 35:339-345YangY, He XL, Hu YF, HouY, Niu YJ, Dai JL, Yuan HX, Li HL. Resistance of wheat cultivars in Huang-Huai region of China to crown rot caused by Fusarium pseudograminearum. J Triticeae Crops, 2015, 35:339-345 (in Chinese with English abstract)[本文引用:1]
[6]
Smiley RW, Gourlie JA, Easley SA, Patterson LM, Whittaker RG. Crop damage estimates for crown rot of wheat and barley in the Pacific Northwest. Plant Dis, 2005, 89:595-604[本文引用:1]
[7]
贺小伦, 周海峰, 侯莹, 胡艳峰, 袁虹霞, 李洪连. 黄淮麦区小麦茎基腐病病原菌的分离鉴定. 见:郭泽建, 吴元华主编. 中国植物病理学会2014年学术年会论文集, 北京:中国农业科学技术出版社, 2014. pp 40-43He XL, Zhou HF, HouY, Hu YF, Yuan HX, Li HL. Isolation and identification of Huang-Huai wheat root rot pathogen. In:Guo Z J, Wu Y H eds. Proceedings of the Annual Meeting of the Chinese Society for Plant Pathology (2014), Beijing:China Agricultural Science and Technology Press, 2014. pp 40-43(in Chinese)[本文引用:2]
[8]
陈厚德, 王彰明, 袁树忠, 陈占荣, 柳玉标, 梅爱中. 小麦茎基褐腐病的发生特点及其对产量的影响. 扬州大学学报(自然科学版), 2001, 4(4):43-46Chen DH, Wang ZM, Yuan SZ, Chen ZR, Liu YB, Mei AZ. Occurrence characteristics of brown rot and its effect on yield of Wheat. J Yangzhou Univ (Nat Sci Edn), 2001, 4(4):43-46 (in Chinese)[本文引用:1]
[9]
樊少华, 李敏权. 定西地区小麦根部镰刀菌及其致病性研究. 甘肃农业大学学报, 2007, 42(4):71-74Fan SH, Li MQ. Pathogenicity of species in Fuarium associated with root of in Dingxi. J Gansu Agric Univ, 2007, 42(4):71-74 (in Chinese with English abstract)[本文引用:1]
[10]
Li HL, Yuan HX, FuB, XingX, SunB, TangW. First report of Fusarium pseudograminearum causing crown rot of wheat in Henan, China. Plant Dis, 2012, 96:1065-1065[本文引用:1]
[11]
付博, 杨云, 周海峰, 袁虹霞, 孙炳剑, 邢小萍, 李洪连. 河南省小麦茎基腐病原鉴定及其致病力研究. 见:郭泽建, 李宝笃主编. 中国植物病理学会2012年学术年会论文集. 北京:中国农业科学技术出版社, 2012. pp 160-163FuB, YangY, Zhou HF, Yuan HX, Sun BJ, Xing XP, Li HL. Pathogen identification and pathogenicity of wheat root rot in Henan. In:Guo Z J, Li B D eds. Proceedings of the Annual Meeting of the Chinese Society for Plant Pathology (2012), Beijing:China Agricultural Science and Technology Press, 2012. pp 160-163(in Chinese)[本文引用:1]
[12]
王志卫, 贝学军, 朱世平, 马岩岩, 杨润婷. 植物激素在植物抗病过程中的作用研究进展. 安徽农业科学, 2011, 39:9035-9038Wang ZW, Bei XJ, Zhu SP, Ma YY, Yang RT. Recent advances in phytohormone regulated plant resisitance to pathogens. J Anhui Agric Sci, 2011, 39:9035-9038 (in Chinese with English abstract)[本文引用:1]
[13]
WangD, Pajerowska-MukhtarK, Culler AH, DongX. Salicylic acid inhibits pathogen growth in plants through repression of the auxin signaling pathway. Curr Biol, 2007, 17:1784-1790[本文引用:1]
[14]
Gonzalez-LamotheR, El OirdiM, BrissonN, BouarabK. The conjugated auxin indole-3-acetic acid-aspartic acid promotes plant disease development. Plant Cell, 2012, 24:762-777[本文引用:1]
[15]
张碧玉, 苏良辰, 孙叙卓. 拟南芥生长素输入载体AUX1研究. 亚热带植物科学, 2010, 39:88-91Zhang BY, Su LC, Sun XZ. Research on auxin influx transporter AUXl of Arabidopsis. Subtrop Plant Sci, 2010, 39:88-91 (in Chinese with English abstract)[本文引用:1]
[16]
SwarupR, FrimlJ, MarchantA, LjungK, Sand bergG, PalmeK, BennettM. Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev, 2001, 15:2648-2653[本文引用:1]
[17]
MullerA, GuanC, GalweilerL, TanzlerP, HuijserP, MarchantA, ParryG, BennettM, WismanE, PalmeK. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J, 1998, 17:6903-6911[本文引用:1]
[18]
朱宇斌, 孔莹莹, 王君晖. 植物生长素响应基因SAUR的研究进展. 生命科学, 2014, 26:407-413Zhu BY, Kong YY, Wang JH. Research advances in auxin- responsive SAUR genes. Chin Bull Life Sci, 2014, 26:407-413 (in Chinese with English abstract)[本文引用:1]
[19]
McClure BA, GuilfoyleT. Characterization of a class of small auxin-inducible soybean polyadenylated RNAs. Plant Mol Biol, 1987, 9:611-623[本文引用:1]
[20]
KnaussS, RohrmeierT, LehleL. The auxin-induced maize gene ZmSAUR2 encodes a short-lived nuclear protein expressed in elongating tissues. J Biol Chem, 2003, 278:23936-23943[本文引用:2]
[21]
张红梅, 王俊丽, 廖祥儒. 细胞分裂素的生物合成、代谢和受体. 植物生理学通讯, 2003, 39:267-272Zhang HM, Wang JL, Liao XR. Biosynthesis metabolize and receptor of cytomin. Plant Physiol Commun, 2003, 39:267-272 (in Chinese)[本文引用:1]
[22]
郭瑾, 薛永来, 杜道林. 植物激素调控拟南芥根系发育的研究进展. 江苏农业科学, 2014, 42(5):7-10GuoJ, Xue YL, Du DL. Research progress of plant hormones regulating root development in Arabidopsis. Jiangsu Agric Sci, 2014, 42(5):7-10 (in Chinese with English abstract)[本文引用:1]
[23]
HwangI, SheenJ. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature, 2001, 413:383-389[本文引用:1]
[24]
ZhangH, HanW, De SmetI, TalboysP, LoyaR, HassanA, RongH, Jürgens GJ, Knox JP, Wang MH. ABA promotes quiescence of the quiescent centre and suppresses stem cell differentiation in the Arabidopsis primary root meristem. Plant J, 2010, 64:764-774[本文引用:1]
[25]
韩淑鸿, 罗锋, 付嵘. ABA在植物与病原菌互作中的作用. 山西农业科学, 2014, 42:1236-1238Han SH, LuoF, FuR. ABA in Interaction between plant and pathogen. J Shanxi Agric Sci, 2014, 42:1236-1238 (in Chinese with English abstract)[本文引用:1]
阮海华. 植物PP2C蛋白磷酸酶负调控ABA信号转导途径研究进展. 安徽农业科学, 2007, 35:652-653Ruan HH. Research progress in the negative regulation of PP2C-type protein phosphatase in ABA signaling of plant. J Anhui Agric Sci, 2007, 35:652-653 (in Chinese with English abstract)[本文引用:1]
[28]
胡帅, 王芳展, 刘振宁, 刘亚培, 余小林. PYR/PYL/RCAR蛋白介导植物ABA的信号转导. 遗传, 2012, 34:560-572HuS, Wang FZ, Liu ZN, Yu XL. ABA signaling mediated by PYR/PYL/RCAR in plants. Hereditas (Beijing), 2012, 34:560-572 (in Chinese with English abstract)[本文引用:1]
[29]
曹婧, 兰海燕. 植物激素脱落酸受体及其信号转导途径研究进展. 生物技术通报, 2014, (6):22-28CaoJ, Lan HY. Research progress on abscisic acid receptor and signal transdction pathway. Biotechnol Bull, 2014, (6):22-28 (in Chinese with English abstract)[本文引用:1]
[30]
DiX, Takken F L W, Tintor N. How phytohormones shape interactions between plants and the soil-borne fungus Fusarium oxysporum. Front Plant Sci, 2015, 7:170-170[本文引用:1]
[31]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25:402-408[本文引用:1]
[32]
VermaV, RavindranP, Kumar PP. Plant hormone-mediated regulation of stress responses. BMC Plant Biol, 2016, 16:1-10[本文引用:1]
[33]
SpaepenS, Vand erleydenJ. Auxin and plant-microbe interactions. Cold Spring Harb Perspect Biol, 2011, 3:704-704[本文引用:1]
[34]
康书静, 钱前, 朱丽. 生长素对水稻根系生长发育调控的研究进展. 中国稻米, 2014, 20(4):1-8Kang SJ, QianQ, ZhuL. Research progress of auxin on the growth and development of rice root system. China Rice, 2014, 20(4):1-8 (in Chinese)[本文引用:1]
[35]
Gee MA, HagenG, Guilfoyle TJ. Tissue-specific and organ- specific expression of soybean auxin-responsive transcripts GH3 and SAURs. Plant Cell, 1991, 3:419-430[本文引用:1]
[36]
任怡怡, 戴绍军, 刘炜. 生长素的运输及其在信号转导及植物发育中的作用. 生物技术通报, 2012, (3):9-16Ren YY, Dai SJ, LiuW. Auxin transport and its roles in signal transduction and plant development. Biotechnol Bull, 2012, (3):9-16 (in Chinese with English abstract)[本文引用:1]
[37]
NavarroL, DunoyerP, JayF, ArnoldB, DharmasiriN, EstelleM, VoinnetO, Jones JD. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science, 2006, 312:436-439[本文引用:1]
[38]
AloniR, AloniE, LanghansM, Ullrich CI. Role of cytokinin and auxin in shaping root architecture:regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann Bot, 2006, 97:883-893[本文引用:1]
[39]
KuderováA, HejátkoJ. Spatiotemporal aspect of cytokinin-auxin interaction in hormonal regulation of the root meristem. Plant Signal Behav, 2009, 4:156-157[本文引用:1]
唐莹莹. 细胞分裂素B类ARRs通过负调节生长素合成酶基因YUCCAs的表达参与拟南芥茎端分生组织的形成. 山东农业大学硕士学位论文, 山东泰安, 2012Tang YY. Cytokinin Type-B ARRs Negatively Regulate Auxin Biosynthetic Genes YUCCAs Expression Invovled in Formation of Shoot Apical Merstem Arabidopsis. MS Thesis of Shand ong Agricultural University, Tai’an, China, 2012 (in Chinese with English abstract)[本文引用:1]
[42]
丁新伦, 谢荔岩, 林奇英, 吴祖建, 谢联辉. 水稻条纹病毒胁迫下抗、感病水稻品种胼胝质的沉积. 植物保护学报, 2008, 35:19-22Ding XL, Xie LY, Lin QY, Wu ZJ, Xie LH. Callose deposition in resistant and susceptible rice varieties under Rice stripe virus stress. Acta Phytophyl Sin, 2008, 35:19-22 (in Chinese with English abstract)[本文引用:1]
[43]
Morrison EN, Emery RJ, Saville BJ. Phytohormone Involvement in the Ustilago maydis-Zea mays pathosystem:relationships between abscisic acid and cytokinin levels and strain virulence in infected cob tissue. PLoS One, 2015, 10:e0130945[本文引用:1]
[44]
KaliffM, StaalJ, MyrenåsM, DixeliusC. ABA is required for Leptosphaeria maculans resistance via ABI1- and ABI4- dependent signaling. Mol Plant Microbe Interact, 2007, 20 :335-345[本文引用:1]
[45]
Mauch-ManiB, MauchF. The role of abscisic acid in plant-pathogen interactions. Curr Opin Plant Biol, 2005, 8:409-414[本文引用:1]
 , 陈琳琳, 孙炳剑, 王利民, 邢小萍, 袁虹霞, 丁胜利
, 陈琳琳, 孙炳剑, 王利民, 邢小萍, 袁虹霞, 丁胜利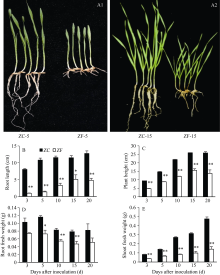






{kind=link}
{kind=link}
{kind=link}
{kind=link}