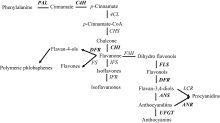
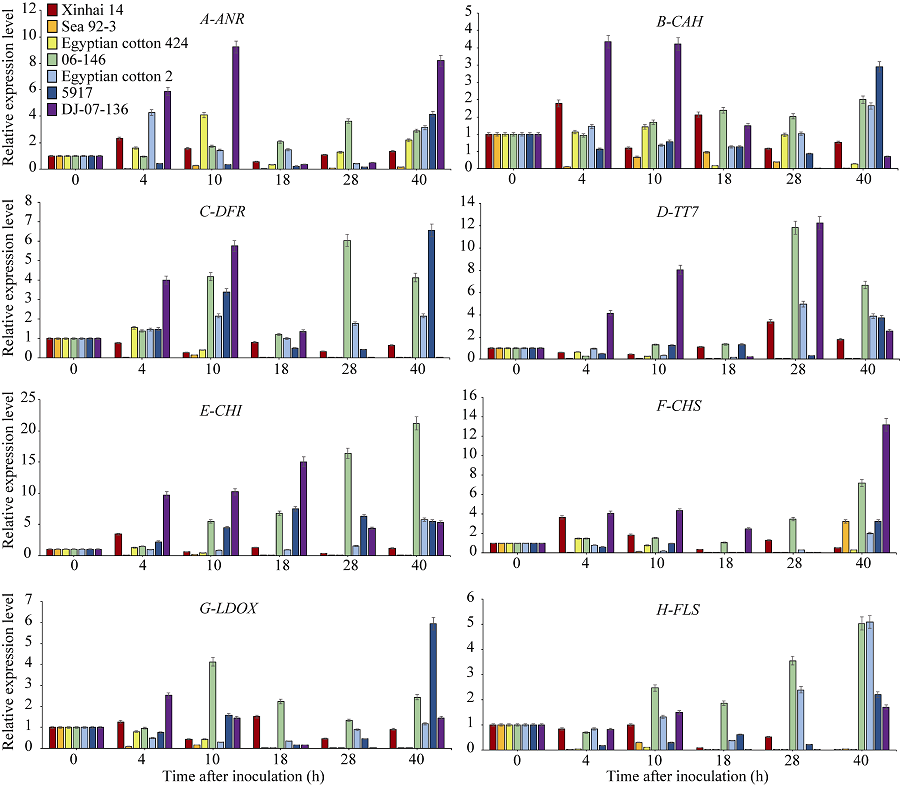
关键词:海岛棉; 枯萎病; 类黄酮; 表达分析; 转录组测序 Correlation between Fusarium wilt Resistance and Expression Levels of Genes Involved in Flavonoid Metabolism Pathway in Gossypium barbadense L HUANG Qi-Xiu, QU Yan-Ying, YAO Zheng-Pei, LI Meng-Yu, CHEN Quan-Jia* College of Agronomy / Key Laboratory of Agriculture Biological Technology, Xinjiang Agricultural University, Urumqi 830052, China Fund:This study was supported by the National Natural Science Foundation of China (31560409) AbstractFusarium wilt is one of the important factors threatening the production of island cotton ( Gossypium barbadense L.). Understand the molecular mechanism of Fusarium wilt resistance will facilitate cotton breeding against Fusarium wilt and therefore alleviate the problem in cotton production. On the basis of RNA-seq results in previous studies, differentially expressed genes (DEG) were further analyzed in this study. After inoculating the pathogenic fungus for 0-40 hours, the DEGs expression levels of seven varieties in different wilt resistance were studied using qRT-PCR method. The correlation between gene expression level and disease index was analyzed. The flavonoid biosynthetic pathway genes were found to be related to Fusarium wilt resistance. The qRT-PCR result indicated the significantly higher expression levels of flavonoid metabolic pathway genes in resistant genotypes than those susceptible genotypes. The expression levels of TT7, CHI, and DFR, the key genes in flavonoid metabolic pathways, were significantly higher in resistant genotypes than in susceptible genotypes ( P < 0.05 or P < 0.01) at several time points after inoculation. Particularly, the expression levels of CHIand DFRwere negatively correlated with disease index. Therefore, we conclude that the flavonoid metabolic pathway related genes might be involved in the resistance to Fusarium wilt in cotton, among which CHI, TT7, and DFRare the key genes.
Keyword: Gossypium barbadense; Fusarium wilt; Flavonoids; Expression analysis; RNA-seq Show Figures Show Figures
表4 转录组测序中类黄酮代谢相关基因的表达 Table 4 Expression of related genes in flavonoids by RNA-seq
基因 Gene
品种 Variety
表达趋势 Expression trend
差异倍数 Fold change
基因 Gene
品种 Variety
表达趋势 Expression trend
差异倍数 Fold change
CHS
06-146
Up
15.81
LDOX/ANS
06-146
Up
13.30
RIL 10893
Up
23.90
RIL 10893
Up
11.36
RIL 10895
Up
4.88
RIL 10895
Up
2.80
Xinhai 14
Up
2.34
Xinhai 14
Up
2.00
RIL 10796
Up
2.18
RIL 10796
Up
1.53
RIL 10897
Up
2.15
RIL 10897
Up
1.18
CHI-1
06-146
Up
11.54
DFR-1
06-146
Up
8.27
RIL 10893
Up
9.22
RIL 10893
Up
8.35
RIL 10895
Up
2.52
RIL 10895
Up
1.38
Xinhai 14
Up
1.03
Xinhai 14
Up
1.02
RIL 10796
Up
1.83
RIL 10796
Down
0.46
RIL 10897
Up
1.06
RIL 10897
Up
1.18
CHI-2
06-146
Up
12.99
DFR-2
06-146
Up
8.17
RIL 10893
Up
10.97
RIL 10893
Up
9.12
RIL 10895
Up
2.63
RIL 10895
Up
3.23
Xinhai 14
Up
1.45
Xinhai 14
Up
1.68
RIL 10796
Up
2.01
RIL 10796
Up
1.25
RIL 10897
Up
1.84
RIL 10897
Up
1.74
TT7-1
06-146
Up
11.54
ANR
06-146
Up
6.37
RIL 10893
Up
10.61
RIL 10893
Up
4.97
RIL 10895
Up
6.10
RIL 10895
Up
2.04
Xinhai 14
Up
5.90
Xinhai 14
Up
1.28
RIL 10796
Up
4.50
RIL 10796
Down
0.94
RIL 10897
Up
4.04
RIL 10897
Up
1.29
TT7-2
06-146
Up
5.72
RIL 10893
Up
4.50
RIL 10895
Up
2.62
Xinhai 14
Up
1.89
RIL 10796
Down
0.90
RIL 10897
Up
1.72
The same gene has two forms, for example, CHI gene is displayed as CHI-1 and CHI-2, in fact, CHI-1and CHI-2 are similar to the same sequence of TAIR, this phenomenon is caused by different parts of a sequence alignment. But its expression trends are similar. So in the following gene analysis process unified the display for CHS. 在表中出现同一基因的2个表达, 例如: CHI基因为CHI-1和CHI-2, 实际上CHI-1和CHI-2序列均与拟南芥库中的同一个基因相似, 这种现象是由比对到序列部位不同而导致, 但其表达趋势均类似。因此后续基因分析过程中统一用CHI代表。
表4 转录组测序中类黄酮代谢相关基因的表达 Table 4 Expression of related genes in flavonoids by RNA-seq
表6 抗、感材料间基因表达量方差分析 Table 6 Variance analysis of gene expression between resistant and susceptible varieties
基因 Gene
接菌后时间 Times after inoculation
4 h
10 h
18 h
28 h
40 h
CHI
2.283
16.527* *
15.362* *
11.335* *
9.414* *
DFR
6.991*
32.341* *
19.942* *
5.232*
12.719* *
TT7
3.094
4.641*
2.529
11.492* *
37.741* *
ANR
3.268
0.773
6.473*
0.421
13.133* *
C4H
1.596
4.316
1.921
1.323
16.759* *
CHS
0.011
1.910
4.446*
0.882
9.133* *
LDOX
2.225
6.885*
0.317
6.990*
13.033* *
FLS
1.348
5.079*
3.690
6.276*
30.590* *
* and * * indicate significant difference between resistant and susceptible genotypes at P< 0.05 and P< 0.01, respectively. * 和* * 分别表示在P< 0.05和P< 0.01水平抗、感材料间差异显著。
表6 抗、感材料间基因表达量方差分析 Table 6 Variance analysis of gene expression between resistant and susceptible varieties
孔庆平. 我国海岛棉生产概况及比较优势分析. 中国棉花, 2002, 12: 19-23Kong QP. Production situation and comparative advantage analysis of Gossypium barbadense in China. China Cotton, 2002, 12: 19-23 (in Chinese with English abstract)[本文引用:1]
[2]
徐秋华, 张献龙, 聂以春, 冯纯大. 我国棉花抗枯萎病品种的遗传多样性分析. 中国农业科学, 2002, 35: 272-276Xu QH, Zhang XL, Nie YC, Feng CD. Genetic diversity evaluation of cultivars (G. hirsumtum L. ) resistant to fusarium wilt by RAPD markers. Sci Agric Sin, 2002, 35: 272-276 (in Chinese with English abstract)[本文引用:1]
[3]
彭姜龙, 曲延英, 王莉萍, 王聪, 郝维维, 汪铖亮. 不同温度条件下接种棉花枯萎病菌海岛棉和陆地棉抗病性研究. 新疆农业科学, 2013, 50: 89-93Peng JL, Qu YY, Wang LP, WangC, Hao WW, Wang CL. A study on properties of resistance to Fusarium wilt in leaves of sea island cotton (Gossipium barbadense) and upland cotton (Gossipium hirsutum L. ) under different temperature conditions. Xinjiang Agric Sci, 2013, 50: 89-93 (in Chinese with English abstract)[本文引用:1]
[4]
校百才. 陆地棉抗枯、黄萎病性状配合力、遗传力的初步研究. 作物学报, 1985, 11: 267-273Xiao BC. Preliminary study on cotton Fusarium and Verticillium wilt resistance traits, combining ability and heritability. Acta Agron Sin, 1985, 11: 267-273 (in Chinese with English abstract)[本文引用:1]
[5]
Smith AL, Dick JB. Inheritance of resistance to Fusarium wilt in upland and sea island cottons as complicated by nematodes under field conditions. Phytopathology, 1960, 50: 44-48[本文引用:1]
[6]
YangJ, MaQ, ZhangY, Wang XF, Zhang GY, Ma ZY. Molecular cloning and functional analysis of GbRVd, a gene in Gossypium barbadense that plays an important role in conferring resistance to Verticillium wilt. Gene, 2016, 575: 687-694[本文引用:1]
[7]
Gao XQ, WheelerT, Li ZH, Charles MK, HeP, Shan LB. Silencing GhNDR1 and GhMKK2 compromises cotton resistance to Verticillium wilt. Plant J, 2011, 66: 293-375[本文引用:1]
[8]
SnyderM. RNA-seq: a revolutionary tool for transcriptomics. Nat Rev Genet, 2009, 10: 57-63[本文引用:1]
[9]
XuL, Zhu LF, Tu LL, Liu LL, Yuan DJ, JinL, LongL, Zhang XL. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-Seq-dependent transcriptional analysis and histochemistry. J Exp Bot, 2011, 62: 5607-5621[本文引用:4]
HichriI, BarrieuF, BogsJ, KappelC, DelrotS, LauvergeatV. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J Exp Bot, 2011, 62: 2465-2483[本文引用:2]
[12]
MalhotraB, Onyilagha JC, Bohm BA, Towers G H N, James D, Harborne J B, French C J. Inhibition of Tomato ringspot virus by flavonoids. Phytochemistry, 1996, 43: 1271-1276[本文引用:1]
Anna KP, EdwardF. S, Jaroslav P. Effect of flavonoids on mycelial growth of Verticillium albo-atrum. Biochem Syst & Ecol, 1995, 23: 683-693[本文引用:1]
[15]
左涛, 赵树堂, 卢孟柱, 孙爱东, 王延伟, 贺伟. 杨树二氢黄酮醇-4-还原酶基因(DFR)的克隆及反义表达对儿茶素合成的影响. 东北林业大学学报, 2016, 10: 49-55ZuoT, Zhao ST, Lu MZ, Sun AD, Wang YW, HeW. Cloning dihydroflavonol-4-reductase gene (DFR) of poplar and its antisense expression effects on catechin synthesis. J Northeast For Univ, 2016, 10: 49-55 (in Chinese with English abstract)[本文引用:1]
[16]
马银平, 王付欣, 杨淳淋, 沈法富, 夏桂先. 海岛棉几丁质酶基因GbCHI的克隆与功能分析. 遗传, 2012, 34: 240-247Ma YP, Wang FX, Yang CL, Shen FF, Xia GX. Cloning and functional analysis of chitinase gene GbCHI from sea-island cotton (Gossypium barbadense). Hereditas (Beijing), 2012, 34: 240-247 (in Chinese with English abstract)[本文引用:1]
[17]
Leslie AW, Li GQ, DoreenW, Imre ES, Keith RD. The phenylalanine ammonialyase gene family in Arabidopsis thaliana. Plant Mol Biol, 1995, 27: 327-338[本文引用:1]
[18]
WajadN, Abdul LT, SaghirA, KhalidM, AbidM, Zhou BL. Evaluation of cotton leaf curl virus resistance in BC1, BC2, and BC3 progenies from an interspecific cross between Gossypium arboreum and Gossypium hirsutum. PLoS One, 2014, 9: e111861[本文引用:1]
[19]
Emilie FF, Bart P H J T. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol Plant Pathol, 2006, 7: 71-86[本文引用:1]
Kunkel BN, Brooks DM. Cross talk between signaling pathways in pathogen defense. Curr Opin Plant Biol, 2002, 5: 325-331[本文引用:1]
[22]
Patil MA, Pierce ML, Phillips AL, Venters BJ, EssenbergM. Identification of genes up-regulated in bacterial-blight-resistant upland cotton in response to inoculation with Xanthomonas campestris pv. malvacearum. Physiol Mol Plant Pathol, 2005, 67: 319-335[本文引用:1]
[23]
AtsushiN, ChikaraM, MineoS, HideyukiM, AtsushiK, Hong JS, KeisukeK, JunA, AkiraK. Functional analysis of soybean genes involved in flavonoid biosynthesis by virus-induced gene silencing. Plant Biotechnol J, 2007, 5: 778-790[本文引用:2]
[24]
Boots AW, Haenen G R M M, Bast A. Health effects of quercetin: from antioxidant to nutraceutical. Eur J Pharmacol, 2008, 585: 325-337[本文引用:2]
[25]
包改红, 毕阳, 李永才, 王毅, 王婷, 唐瑛, 马朝玲, 白小东. 硫色镰刀菌接种对抗病/易感品种马铃薯块茎苯丙烷代谢的影响比较. 食品科学, 2015, 36: 251-256Bao GH, BiY, Li YC, WangY, WangT, TangY, Ma ZL, Bai XD. Comparison of phenylpropanoid pathway metabolism in slices of susceptible and resistant potato cultivars inoculated with Fusarium sulphureum. Food Sci, 2015, 36: 251-256 (in Chinese with English abstract)[本文引用:1]
Tan BA, Daim L D J, Ithnin N, Ooi T E K, Md-Noh N, Mohamed M, Mohd-Yusof H, Appleton D R, Kulaveerasingam H. Expression of phenylpropanoid and flavonoid pathway genes in oil palm roots during infection by Ganoderma boninense. Plant Gene, 2016, 7: 11-20[本文引用:1]
[28]
Sun WG, XuY. Study on the composition of the pigments in the nature colored cotton. J Xi’an Polytechnic Univ, 2009, 23: 119-124[本文引用:1]
[29]
SunQ, Jiang HZ, Zhu XY, Wang WN, He XH, Shi YZ, Yuan YL, Du XM, Cai YF. Analysis of sea-island cotton and upland cotton in response to Verticillium dahliae infection by RNA sequencing. BMC Genomics, 2013, 14: 852[本文引用:1]
[30]
FedericoP, MartaN, María AB, FuencislaM, Barceló AR. Changes in stem lignins (monomer composition and crosslinking) and peroxidase are related with the maintenance of leaf photosynthetic integrity during Verticillium wilt in Capsicum annuum. New Phyt, 2004, 163: 111-123[本文引用:1]
[31]
GayosoC, PomarF, Novo-UzalE, MerinoF, Martínez D I O. The Ve-mediated resistance response of the tomato to Verticillium dahliae involves H2O2, peroxidase and lignins and drives PAL gene expression. J Med Hum, 2010, 10: 1-19[本文引用:1]
[32]
Irina OV, Robert TR, Jane E G. David J R, Kent AM, NicholiV. Characterization of flavonols in Vaccinium macrocarpon powder. J Agric Food Chem, 2003, 52: 188-195[本文引用:1]
[33]
LiF, JinZ, QuW, ZhaoD, MaF. Cloning of a cDNA encoding the Saussurea medusa chalcone isomerase and its expression in transgenic tobacco. Plant Physiol Biochem, 2006, 44: 455-461[本文引用:1]
[34]
FaweA, Abou ZM, Menzies JG, Belanger RR. Silicon-mediated accumulation of flavonoid phytoalexins in cucumber. Phytopathology, 1998, 88: 396-401[本文引用:1]
[35]
马春雷, 乔小燕, 陈亮. 茶树无色花色素还原酶基因克隆及表达分析. 茶叶科学, 2010, 30: 27-36Ma CL, Qiao XY, ChenL. Cloning and expression analysis of Leucoanthocyantin reducase gene of tea plant (Camellia sinensis). J Tea Sci, 2010, 30: 27-36 (in Chinese with English abstract)[本文引用:1]
[36]
宋凤鸣, 郑重, 童贤明, 葛秀春. 儿茶素对棉枯萎病菌胞壁降解酶的抑制及在棉花抗病性中的作用. 真菌学报, 1996, 4: 297-303Song FM, ZhengZ, Tong XM, Ge XC. Inhibition of tea catechins on cell wall degrading enzymes of cotton Fusarium wilt and its role in disease resistance of cotton. Mycosystema, 1996, 4: 297-303 (in Chinese with English abstract)[本文引用:1]
[37]
Shih CH, Chu IK, Yip WK, LoC. Differential expression of two flavonoid 3’-hydroxylase cDNAs involved in biosynthesis of anthocyanin pigments and 3’-deoxyanthocyanidin phytoalexins in sorghum. Plant Cell Physiol, 2006, 47: 1412-1419[本文引用:1]
[38]
张松焕, 李春奇, 郭惠明, 裴熙祥, 程红梅. 过量表达紫茎泽兰类黄酮3’-羟化酶基因对转基因烟草POD、PAL活性的影响. 中国农业科技导报, 2009, 11(3): 98-101Zhang SH, Li CQ, Guo HM, Pei XX, Cheng HM. Effects of eupatorium adenophorum flavonoid 3'-hydroxylase overexpression on pod and pal activity in transgenic tobacco. J Agric Sci Technol, 2009, 11(3): 98-101 (in Chinese with English abstract)[本文引用:1]
[39]
KohliD, JoshiG, Deokar AA, Bhardwaj AR, AgarwalM, Katiyar-AgarwalS, SrinivasanR, Jain PK. Identification and characterization of wilt and salt stress-responsive microRNAs in chickpea through high-throughput sequencing. PLoS One, 2014; 9: e108851[本文引用:1]
 , 曲延英, 姚正培, 李梦雨, 陈全家
, 曲延英, 姚正培, 李梦雨, 陈全家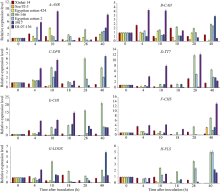


{kind=link}
{kind=link}