 ,1,*, 李文旭3, 陈新宏2
,1,*, 李文旭3, 陈新宏2Molecular Cloning and Functional Analysis of 1-FFT in Wheat Relatives
WANG Jian-Wei1, HE Xiao-Lan,1,*, LI Wen-Xu3, CHEN Xin-Hong2通讯作者:
第一联系人:
收稿日期:2017-08-26接受日期:2018-03-26网络出版日期:2018-06-12
| 基金资助: |
Received:2017-08-26Accepted:2018-03-26Online:2018-06-12
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (11458KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王建伟, 贺晓岚, 李文旭, 陈新宏. 小麦近缘属植物1-FFT基因的克隆及功能分析[J]. 作物学报, 2018, 44(6): 814-823. doi:10.3724/SP.J.1006.2018.00814
WANG Jian-Wei, HE Xiao-Lan, LI Wen-Xu, CHEN Xin-Hong.
干旱、低温、高盐等非生物胁迫是影响植物生长发育的重要环境胁迫因子, 严重影响着作物的产量和品质[1]。果聚糖既是温带和寒带地区植物组织中重要的能量贮藏物质, 又是提高植物抗逆性(抗旱、抗寒、抗盐等)的重要成分[2,3]。基于糖苷键的连锁类型植物中的果聚糖可划分为5个类型, 在高等植物中, 主要有4种不同的果聚糖基转移酶参与果聚糖的合成[4]。其中1-FFT的主要功能是促进果聚糖碳链的进一步延伸, 产生高聚合度果聚糖[5]。除1-FFT之外, 果聚糖:果聚糖6G-果糖基转移酶(fructan: fructansyltrans 6G-fructosyltcallsferase 6G-FFT)和蔗糖:果聚糖6-果糖基转移酶(sucrose:fructan-6- fructosyltransferase 6-SFT)也参与糖苷键进一步延伸的生化反应, 但是不同酶催化可能形成不同的糖苷键类型[6]。对小麦族果聚糖合成酶的克隆和功能研究, 可为定向调控植物果聚糖的含量和组分, 提高植物抗逆性提供遗传依据。
早在1966年就有关于菊芋1-FFT的报道[7], 但直到20世纪80年代才从菊芋中分离纯化得到1-FFT[8,9], 随后在蓝刺头[5]、维氏菊[10]、圆锥小麦[11]、普通小麦[11]、乌拉尔图小麦[11]等植物中克隆得到1-FFT, 并且对其结构和功能进行了深入研究。小麦族1-FFT基因在进化上相对较保守, 除节节麦外, 乌拉尔图小麦、西尔斯山羊草、圆锥小麦和普通小麦1-FFT基因的开放阅读框(ORF)长度相等, 均为1947 bp, 并且不同物种1-FFT基因所编码氨基酸均含有3个高度保守的活性结构域, 即NDPNG、RDP和ECID [2]。1-FFT对增强植物的抗旱和抗寒性起重要作用[2], 但是1-FFT是如何提高植物抗逆性的机制还不明确。转入冰草1-FFT基因的黑麦草, 其果聚糖积累量和可溶性总糖含量显著增加[12]。Knipp和Honermeier[13]报道果聚糖合成酶基因的表达可能对脯氨酸的积累具有多向性效应。Li等[14]报道果聚糖合成酶基因能增强转基因植株氧化胁迫抗性, 降低植株电解质渗漏率和丙二醛含量。在逆境胁迫条件下, 可溶性糖和脯氨酸等抗逆重要调节物质的积累有利于增强植物对旱胁迫和冷胁迫等的抗性[15,16]。因此, 在非果聚糖积累植物中表达果聚糖合成相关基因对改良植物抗旱及抗寒性具有重要意义。
不同物种1-FFT基因的功能不尽相同, 且1-FFT的聚合度具有物种特异性[17]。华山新麦草(Psathyrostachys huashanica, 2n = 2x = 14, NsNs)是多年生异花授粉植物, 属禾本科小麦亚家族, 具有早熟、耐旱、抗寒、抗盐、抗病等优异性状[18]; 簇毛麦(Dasypyrum villosum, 2n=2x=14, VV)是一年生异花授粉植物[19], 属禾本科小麦亚家族[20], 具有抗旱、抗寒、抗盐、抗多种病害、分蘖力强、小穗数多、蛋白质含量高等优良性状[20,21]; 大赖草(Leymus racemosus, 2n=4x=28, NsNsXmXm)属禾本科赖草属多年生根茎型草本植物, 具有大穗、多花、抗旱、抗寒、抗多种病害及高耐盐碱性[22]。目前, 关于果聚糖酶基因的研究主要集中于挖掘来源于细菌[23]、真菌[24]、植物[25]等的果聚糖合成相关基因, 并转化非果聚糖植物, 研究转基因植株的果聚糖积累效应、相关生理生化指标及非生物胁迫抗性等。本研究从华山新麦草、簇毛麦和大赖草中分别克隆1-FFT基因, 比较序列间结构差异, 并在转基因烟草中初步验证该基因的抗逆功能, 以期为深入研究不同来源1-FFT基因在干旱和寒冷逆境中的作用, 以及利用转基因技术创制抗逆作物新种质奠定基础。
1 材料与方法
1.1 cDNA全长克隆
1.1.1 1-FFT保守区的克隆 将华山新麦草、簇毛麦和大赖草材料种植于西北农林科技大学试验地(陕西杨凌), 用总RNA提取试剂盒(RNeasy Plant Mini kit, Qiagen, Germany)从叶片中提取总RNA。使用SuperScript III Reverse Transcriptase cDNA第1链合成试剂盒(Invitrogen)合成cDNA第1链。参照乌拉尔图小麦、普通小麦、粗山羊草、大麦和圆锥小麦5种植物的1-FFT蛋白氨基酸保守区序列设计1对简并引物(Dp1-FFTsen: 5′-AYGAYCCNAAYGGN CCNGT-3′, DP1-FFTantisen: 5′-NSWRTCCATRTCR TGNAC-3′; Y=CT, N=ACGT, R=AG), 并用LA Taq (TaKaRa)扩增小麦近缘属植物Ph/Dv/Lr1-FFT基因的cDNA中间保守区。反应程序为94°C预变性3 min; 94°C变性30 s, 55°C退火30 s, 72°C延伸2 min, 35个循环; 72°C延伸10 min。扩增产物经琼脂糖凝胶电泳后回收纯化目的片段, 与pMD19-T载体(TaKaRa)连接, 转化至大肠杆菌E. coli DH5α感受态细胞, 将菌落PCR和质粒单双酶切鉴定正确的5个重组质粒送生工生物工程(上海)有限公司测序。1.1.2 3°端和5°端序列的克隆 按照3¢-Full RACE Core Set Ver.2.0试剂盒和5°-Full RACE Kit试剂盒(TaKaRa)说明书分别克隆小麦近缘属植物Ph/Dv/Lr1-FFT基因cDNA的3¢和5′端序列, 特异引物序列为GSOP3¢-PDL: 5′-TCCAGTCGATTCCGAG AACG-3′, GSIP3¢-PDL: 5′-TACCCTCCGCATCAAC ACC-3′; GSOP5°-PDL: 5′-GGTCATCCTGCTCTACA CG-3′, GSIP5°-PDL: 5′-ATTGGTACGACATCGAG GGC-3′。小麦近缘属植物不同组织总RNA提取、PCR产物的克隆和测序方法如前所述。
1.2 cDNA编码区的获得及生物信息学分析
根据5°和3¢-RACE的结果, 并参考小麦1-FFT基因cDNA序列设计克隆Ph/Dv/Lr-1-FFT基因的引物(PDL-1-FFT-F: 5°-CATGGAGTCGTCACGCGGC ATC-3¢; PDL-1-FFT-R: 5°-CCGACCGAGCAAACAG TCCATAAC-3¢), 通过PCR法扩增PDL-1-FFT基因编码区序列, 并测序验证。采用National Center for Biotechnology Information (NCBI)网站(http://www.ncbi.nlm.nih.gov/)的在线工具DNAMAN 4.0、MegAlign软件和多序列比对程序ClustalX2进行cDNA序列BLAST分析, 用WebLogo (http://weblogo.threeplusone.com/create.cgi)分析残基保守性。蛋白质1-FFT结构域及核定位信号预测软件分别为2ZIP (http://www.expasy.org/)和PSORT II (http://www.genscript.com/cgi-bin/tools/- psort2.pl)。采用ExPASy proteomics server (http:// www.expasy.ch/tools/dna.html)氨基酸的翻译工具; 应用Conserved Domains数据库分析氨基酸序列的保守区域; 使用NCBI的ORF Finder (http://www. ncbi.nlm.nih.gov/gorf/gorf.html) 进行基因ORF分析, 借助ClustalX2和MEGA5.0软件构建系统进化树。
1.3 叶盘法转化烟草及转基因植株的验证
分别将Ph-1-FFT、Dv-1-FFT和Lr-1-FFT的完整编码区序列插入植物表达载体p1300-35SN的Hind III-EcoR I位点, 构建表达载体p1300-35SN- Ph-1-FFT/Dv-1-FFT/Lr-1-FFT (图1), 通过测序确保目的基因在载体中的插入方向及完整性。采用冻融法[27]将重组质粒转入农杆菌GV3101 (本实验室保存), 再利用叶盘法[28]转化烟草W38, 最后将经过10 mg L-1潮霉素筛选的抗性植株移入温室培养。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1植物表达载体p1300-35SN-Ph-1-FFT/Dv-1-FFT/Lr-1-FFT示意图
LB和RB: T-DNA的左、右边界; 35S°: CaMV 35S poly A; 35S2: 含双增强子的CaMV 35S启动子; p35S1: CaMV 35S启动子; Tnos: 终止子。
Fig. 1Structure of p1300-35SN-Ph-1-FFT/Dv-1-FFT/Lr-1-FFT transformation vectors used in the study
LB and RB: left and right border of T-DNA; 35S°: CaMV 35S poly A; p35S2: CaMV 35S promoter with double enhancer sequences; p35S1: CaMV 35S promoter; Tnos: terminator.
采用CTAB法提取烟草叶片基因组DNA, 利用特异引物1-FFT (F: 5°-CATGGAGTCGTCACGC GGCATC-3¢; R: 5°-CCGACCGAGCAAACAGTCCA TAAC-3¢)进行PCR扩增, 检测目的基因。反应条件为94°C 5 min; 94°C 30 s, 60°C 30 s, 72°C 2 min, 35个循环; 72°C 10 min。扩增产物经1.0 %琼脂糖凝胶电泳、染色并照相。
为了分析不同来源1-FFT基因在RNA水平的表达情况, 对PCR阳性转基因植株进行转录分析。按前述方法提取烟草总RNA, 并合成cDNA第1链。以测试烟草叶片cDNA为模板, 首先以烟草actin基因特异引物(ActF: 5°-TGGCATCATACCTTTTACA A-3¢; ActR: 5°-TCCGGGCATCTGAACCTCT-3¢)对各样本进行扩增, 根据扩增结果调整模板浓度, 以使模板cDNA浓度均一化。反应程序为95°C预变性3 min; 94°C 30 s, 55°C 30 s, 72°C 30 s, 35个循环; 72°C延伸10 min。然后用PDL-1-FFT基因的特异引物(Rp1-FFT-F: 5°-GCATCATCATCGGCTCCAA-3¢; Rp1- FFT-R: 5°-TCGTCAAGCTCCACCGTTCT-3¢)进行半定量RT-PCR分析, 反应条件为95°C预变性3 min; 94°C 45 s, 55°C 35 s, 72°C 30 s, 35个循环; 72°C延伸10 min。重复3次。
1.4 转基因烟草的抗旱及抗寒性分析
按He等[26]、贺晓岚等[27]描述的方法鉴定转基因植株的抗旱和抗寒性。选择基因表达量相对较高的T0代植株, 采集逆境胁迫前及干旱胁迫(20 d)或冷胁迫(10°C 10 d或-20°C 5 min)后T0植株和对照植株的叶片, 用蒽酮比色法测定可溶性糖含量[28], 用茚三酮法测定脯氨酸含量[29], 用植物丙二醛测定试剂盒(南京建成)测定丙二醛含量, 并用Plant Fructan Colorimetric Assay Kit (GenMed Scientifics Inc., USA)测定果聚糖含量, 重复测定4次。用SAS V8.1软件统计分析数据, 用t检验法比较均数(P<0.05), 用OriginPro 2015软件作图。
2 结果与分析
2.1 基因克隆
以华山新麦草叶片、簇毛麦根、大赖草叶片的cDNA为模板进行PCR扩增, 3个基因的保守区扩增产物均约1300 bp (图2)。电泳结果显示3′端和5′端片段长约450 bp和500 bp, 而测序结果显示3′端的片段长度均为448 bp, 华山新麦草和簇毛麦该基因5′端片段长514 bp, 簇毛麦该基因5′端片段长475 bp (图2)。通过PCR扩增获得3条约2000 bp的全长cDNA序列(图2)。回收目的片段, 测序结果证明3个片段为Ph-1-FFT、Dv-1-FFT和Lr-1-FFT的编码区。图2
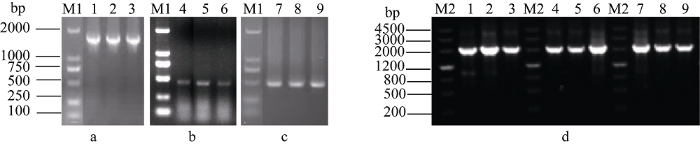 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2华山新麦草、簇毛麦和大赖草1-FFT基因PCR扩增产物电泳分析
M1: DL2000; M2: marker Ш. a: 基因保守区段扩增产物; b: 3¢ RACE扩增产物; c: 5¢ RACE 扩增产物; d: Ph-1-FFT (1~3泳道)、Dv-1-FFT (4~6泳道)和Lr-1-FFT (7~9泳道)的cDNA。
Fig. 2Electrophoresi s analysis of PCR products of 1-FFT gene cloned from P. huashanica, D. villosum, and L. racemosus
M1: DL2000; M2: marker Ш. a: amplification of gene conserved fragment; b: PCR product of 3¢ RACE; c: PCR product of 5¢ RACE; d: cDNA of Ph-1-FFT (lanes 1-3), Dv-1-FFT (lanes 4-6), and Lr-1-FFT (lanes 7-9).
2.2 基因序列分析
cDNA序列分析表明, Ph-1-FFT和Lr-1-FFT的ORF长度相等, 均为1989 bp; Dv-1-FFT的ORF为1950 bp, 等电点为4.87~5.00, 分子量预测为71.3~ 72.2 kD (表1)。经Blast比对, 发现Ph-1-FFT、Lr-1-FFT和Dv-1-FFT基因与普通小麦1-FFT基因序列相似度分别为86%、85%和91%。与已知1-FFT序列比对发现, Ph-1-FFT、Lr-1-FFT和Dv-1-FFT均存在3个高度保守的结构域; 与其他植物不同的是, Ph-1-FFT和Lr-1-FFT基因的蔗糖结合框由“NDPNG”变为“NDPCG”, 但是仍然包含一个活跃的天冬氨酸(D)残基, 在YECID保守区包含一个谷氨酸(E)残基, 在YRDP框包含一个天门冬氨酸(D)残基(图3)。Ph-1-FFT和Lr-1-FFT序列相似性最高, 在进化树上聚为一类; Dv-1-FFT与小麦族其他物种的1-FFT序列相似性最高, 在进化树上聚为一类(图4)。Ph-1-FFT、Lr-1-FFT和Dv-1-FFT的序列相似度高达96%, Dv-1-FFT除缺失长度为9 nt (5°-CCTCGGCCG-3°)和30 nt (5°-ACGGAGTTCCC GAGGAGCAGGGGCAAGGAC-3°)的2个序列片段外, 还存在207个单碱基变异, 但这些单碱基变异大多属同义突变, 未改变编码蛋白, 即在氨基酸水平上, Dv-1-FFT比Ph-1-FFT和Lr-1-FFT在N端缺失两处共13个氨基酸。Dv-1-FFT编码的氨基酸序列与小麦族其他物种1-FFT编码的氨基酸序列相似度较高。Ph-1-FFT和Lr-1-FFT与小麦族其他物种1-FFT编码的氨基酸序列在N-端差异较大, 但功能区域较保守。图4
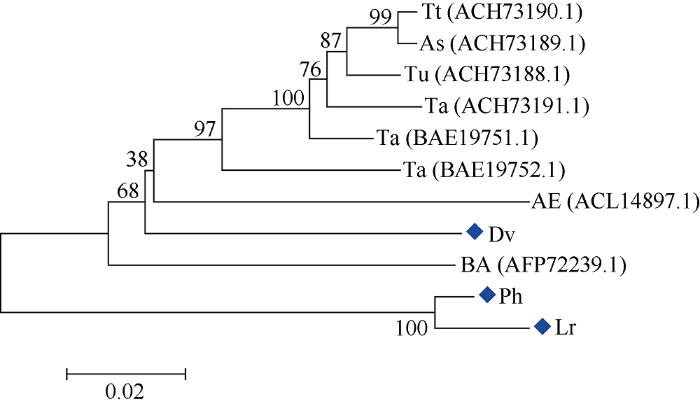 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4华山新麦草、大赖草和簇毛麦与其他物种1-FFT序列系统进化树分析
各节点处数值表示bootstrap值(迭代1000次)。比对物种为华山新麦草(Ph)、大赖草(Lr)、簇毛麦(Dv)、圆锥小麦(Tt)、西尔斯山羊草(As)、乌拉尔图小麦(Tu)、普通小麦(Ta)、粗山羊草(AE)和大麦(BA)。
Fig. 4Phylogenic tree of 1-FFTs from Psathyrostachys huashanica, Leymus racemosus, Dasypyrum villosum and other plant species
The bootstrap percentages (>50%) are shown next to the branches (based on 1000 replicates). The 1-FFT sequences were obtained from Psathyrostachys huashanica (Ph), Leymus racemosus (Lr), Dasypyrum villosum (Dv), Triticum turgidum subsp. durum (Tt) , Aegilops searsii (As), T. urartu (Tu), T. aestivum (Ta), Ae. tauschii (AE), and Hordeum vulgare subsp. vulgare (BA).
Table 1
表1
表1不同来源1-FFT基因的cDNA及其编码产物的长度、分子量和等电点比较
Table 1
| 基因 Gene | cDNA长度 cDNA length (bp) | 编码产物长度 Length of coding product (aa) | 分子量 Mw (kD) | 等电点 pI |
|---|---|---|---|---|
| Ph-1-FFT | 1989 | 662 | 72.2 | 4.91 |
| Dv-1-FFT | 1950 | 649 | 71.3 | 5.00 |
| Lr-1-FFT | 1989 | 662 | 72.0 | 4.87 |
新窗口打开|下载CSV
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3华山新麦草、簇毛麦、大赖草及其他物种1-FFT序列分析
方框显示序列保守区; 星号(*)表示相同残基; 冒号(:)表示保守替换; 点号(.)表示半保守替换。比对物种有普通小麦(Ta)、圆锥小麦(Tt)、西尔斯山羊草(As)、乌拉尔图小麦(Tu)、簇毛麦(Dv)、华山新麦草(Ph)、粗山羊草(AE)、大麦(BA)和大赖草(Lr)。
Fig. 3Sequence alignment of 1-FFTs from Psathyrostachys huashanica, Dasypyrum villosum, Leymus racemosus, and other plant species
Conserved regions are boxed. Asterisks (*), colons (:), and periods (.) show the identical residues, conserved substitutions, and semiconserved substitutions, respectively. The 1-FFT sequences were obtained from Triticum aestivum (Ta), T. turgidum subsp. durum (Tt), Aegilops searsii (As), T. urartu (Tu), Dasypyrum villosum (Dv), Psathyrostachys huashanica (Ph), Ae. tauschii (AE), Hordeum vulgare subsp. vulgare (BA), and Leymus racemosus (Lr).
2.3 转基因植株的获得与鉴定
通过叶盘转化法获得T0代转基因烟草, 经抗生素筛选后, 首先利用1-FFT-F/1-FFTR引物进行PCR验证, 共获得195株阳性植株(图5-A), 再用Rp1- FFT-F/Rp1-FFT-R引物分析验证基因表达, 鉴定获得172株阳性植株。RT-PCR分析结果显示, Ph-1-FFT/Dv-1-FFT/Lr-1-FFT在转基因烟草中均能正常表达, 而在对照植株中未检测到转录信号(图5-B)。图5
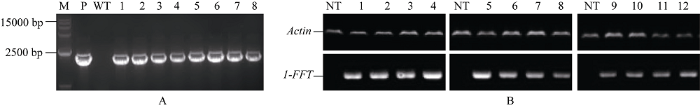 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5T0代转基因植株的鉴定
A: 转基因植株的PCR鉴定; B: 转基因植株1-FFT的表达鉴定。M: D15000 marker; P: 阳性对照(质粒); WT: 阴性对照; NT: 转空载体的阴性对照; 1~12: 转基因植株。
Fig. 5Validation of transgenic T0 plants
A: PCR analysis of transgenic plants; B: expression patterns of 1-FFT gene; M: D15000 marker; P: positive control (plasmid); WT: negative control plants; NT: negative control (non-transgenic plants); 1-12: T0-tobacco transgenic plants.
2.4 转基因烟草逆境胁迫前后表型分析
在正常生长条件下, 转Ph-1-FFT、Dv-1-FFT和Lr-1-FFT基因的烟草植株之间及转基因植株与对照植株的生长无明显差异。干旱胁迫35 d后, 对照植株全部干枯, 而转基因植株的心叶仍然保持绿色, 转不同来源1-FFT基因的植株间无明显差异(图6-a); 复水后, 对照植株未能复活, 而转基因植株在14 d后有少许新叶长出, 转不同来源的1-FFT基因的植株间生长状态无明显差异。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6转华山新麦草(Ph)、簇毛麦(Dv)和大赖草(Lr) 1-FFT基因烟草干旱和冷胁迫后的表型
a: 干旱胁迫处理35 d后; b: 冷胁迫(10°C 10 d, -20°C 35 min)处理后转移至温室培养7 d。NT为转空质粒对照。
Fig. 6Phenotypes of tobacco plants transformed with 1-FFT from Psathyrostachys huashanica (Ph), Dasypyrum villosum (Dv), and Leymus racemosus (Lr)
a: after thirty-five days of drought stress; b: recovery growth at room temperature for seven days after cold stress (10°C for 10 days and -20°C for 35 min). NT indicates the non-transgenic tobacco plant.
低温胁迫处理后, 对照全部枯萎, 而转基因植株心叶仍然保持正常生长势, 转1-FFT基因的植株间生长状态无明显差异(图6-b); 转移至室温继续培养7 d, 对照植株全部死亡, 转基因植株能够恢复生长, 只是生长发育迟缓, 且转不同1-FFT基因的植株间生长状态无明显差异。
2.5 转基因植株逆境胁迫前后生理指标分析
正常条件下, 转基因T0植株的果聚糖含量显著高于对照植株, 转华山新麦草、簇毛麦和大赖草1-FFT基因植株的果聚糖含量分别为5.37~5.52、5.86~6.40和5.40~5.80 mg g-1, 差异不显著, 而在对照植株中没有检测到果聚糖; 转基因植株可溶性糖、脯氨酸的含量均略高于对照植株, 丙二醛的含量略低于对照植株, 未达显著差异(图7)。图7
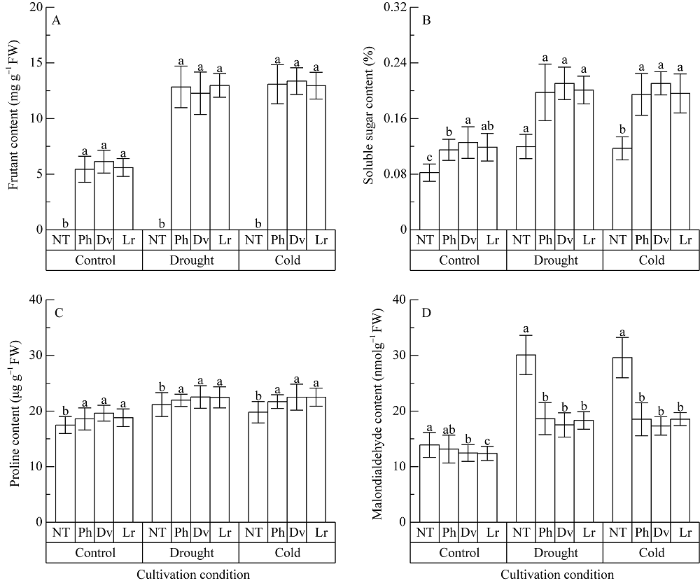 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7转华山新麦草(Ph)、簇毛麦(Dv)和大赖草(Lr) 1-FFT基因烟草干旱和冷胁迫后的生理指标变化
NT为转空质粒对照。在相同条件下, 误差线上不同字母表示4类转基因植株间差异显著(P < 0.05)。
Fig. 7Changes of physiological indices after drought and freezing stressed in tobacco plants transformed with 1-FFT from Psathyrostachys huashanica (Ph), Dasypyrum villosum (Dv), and Leymus racemosus (Lr)
NT indicates the non-transgenic tobacco plant. Different letters above the error bars indicate significant difference among the four types of transgenic plants (P < 0.05).
干旱胁迫20 d后, 转基因植株的果聚糖含量显著增加, 转华山新麦草、簇毛麦和大赖草1-FFT基因植株的果聚糖含量分别为12.84、12.27和12.97 mg g-1, 是胁迫前的2.0~2.2倍, 而在对照植株中没有检测到果聚糖; 转基因植株的可溶性糖为0.20%~ 0.21%, 脯氨酸含量为21.94~22.52 μg g-1, 均显著高于对照植株; 转基因植株的丙二醛含量仅为17.50~ 18.64 nmol g-1, 显著低于对照植株的28.95~29.64 nmol g-1 (图7)。
与干旱胁迫相似, 冷胁迫后转华山新麦草、簇毛麦和大赖草1-FFT基因植株的果聚糖含量分别为13.08、13.36和12.96 mg g-1, 是胁迫前的2.0~2.4倍, 而在对照植株中没有检测到果聚糖; 转基因植株的可溶性糖和脯氨酸含量也均显著高于对照植株, 而丙二醛含量(17.31~18.53 nmol g-1)却显著低于对照植株(图7)。
总之, 干旱和冷胁迫后, 转不同来源1-FFT基因植株中丙二醛的含量略有提高, 而对照植株中丙二醛的含量显著增加。在胁迫条件下, 转不同来源1-FFT基因植株中的碳水化合物和脯氨酸含量都显著高于对照植株, 无论在正常条件还是在胁迫条件下, 转不同来源1-FFT基因植株之间果聚糖、可溶性糖、脯氨酸和丙二醛含量无显著差异。
3 讨论
3.1 Ph-1-FFT/Dv-1-FFT/Lr-1-FFT基因的cDNA及其编码氨基酸序列的特征分析
植物中果聚糖合成相关基因广泛参与植物抗性逆境胁迫[30]。本研究克隆获得的3个小麦近缘属1-FFT基因与其他物种的1-FFT相似性在85%以上, 蛋白质结构分析表明这3个基因编码产物属于GH32家族。簇毛麦1-FFT的氨基酸序列包含NDPNG、RDP和ECID保守结构域; 与其他植物不同, 华山新麦草和大赖草1-FFT基因的蔗糖结合框由NDPNG变为NDPCG, 表明华山新麦草和大赖草1-FFT可能属于GH32基因家族F型的FT类[31]。在1-FFT蛋白系统进化树上, 小麦近缘属植物1-FFT序列与小麦族其他物种的同源序列处于不同分支, 华山新麦草与大赖草1-FFT序列处于同一分支(图4)。簇毛麦1-FFT序列与小麦族其他物种1-FFT序列相似度较高, 华山新麦草和大赖草1-FFT序列虽然与小麦族其他物种在N-端差异较大, 但功能区域相对较保守。因此推测, 1-FFT可能在果聚糖合成路径中起重要作用, 在进化过程中保持着较高的遗传稳定性。华山新麦草和大赖草1-FFT基因的cDNA序列长度相等, 但其推导氨基酸存在较大序列差异, 暗示华山新麦草和大赖草1-FFT基因的cDNA序列差异较大, 因此推测大赖草的1-FFT可能来自Xm基因组。另外, 本研究克隆的小麦近缘属植物1-FFT基因与Gao等[11]从小麦族中分离到的多个1-FFT基因cDNA序列长度不等, 说明本研究克隆的小麦近缘属植物1-FFT基因可能是该家族的新基因。
3.2 Ph-1-FFT、Dv-1-FFT和Lr-1-FFT表达对转基因烟草果聚糖含量和抗逆性的影响
Suárez-González等[30]报道, 1-FFT或1-SST基因在龙舌兰中表达可以提高其对逆境胁迫的抗性, 说明1-FFT基因对提高作物抗逆性起一定作用。本研究中, 在干旱和寒冷胁迫后, 转不同材料1-FFT烟草植株表现出明显高于对照植株的抗逆性, 转基因植株叶片的平均果聚糖含量为5.40~5.86 mg g-1, 逆境胁迫后, 转不同来源1-FFT基因植株的果聚糖含量显著增加, 提高植株的抗旱、抗寒性, 这些结果与前人报道[2, 32]相似。可溶性糖含量增加对提高植物活性氧清除能力有重要作用[33], 脯氨酸含量也与抗逆性密切相关。无论在逆境胁迫前或逆境胁迫后, 转不同来源1-FFT的阳性植株, 叶片中可溶性糖和脯氨酸含量均高于对照植株, 说明不同来源的1-FFT基因均能提高植株的抗旱、抗寒性。碳水化合物代谢的改变可能对脯氨酸的积累起诱导作用[13], 但二者之间的关系很复杂, 其机制有待深入研究。
通过对转基因烟草丙二醛含量的测定, 发现正常条件下转基因植株丙二醛含量较对照植株略低; 干旱和冷胁迫后, 对照的丙二醛含量显著增加, 转基因植株略有增加, 显著低于对照。这表明本研究克隆的3个1-FFT基因, 在转基因烟草中合成的果聚糖降低了逆境胁迫下的膜脂质过氧化程度, 减轻了细胞膜受损。因为小麦属果聚糖合成植物, 将外源果聚糖合成相关基因导入小麦是否可以提高其抗旱和抗寒性尚不清楚, 但是小麦果聚糖合成相关基因在水稻中表达可以使转基因水稻叶片中积累大量的果聚糖, 并且明显提高转基因水稻苗期的抗寒性[34]。
无论是否存在胁迫条件, 3个不同来源的1-FFT基因在转入烟草后, 所观测的生理指标均没有显著差异, 因此还需设计新的实验来比较这3个1-FFT基因的功能, 如合成果糖成分、参与非生物胁迫过程及生物学功能等。
4 结论
通过RT-PCR结合RACE技术, 克隆获得小麦3个近缘属植物的果聚糖合成相关基因, 生物信息学分析表明它们都属于糖基水解酶第32家族, 在转基因烟草中初步验证了3个基因能够显著提高抗旱和抗寒性。3个不同来源的1-FFT基因的编码产物相似度较高, 转入烟草后生理指标差异不显著。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.1007/s00299-012-1332-yURLPMID:22911265 [本文引用: 4]

Seven kinds of transgenic tobacco plants transformed with combinations of three FBE genes were obtained. The transgenic plants transformed with Ta1-SST + Ta6-SFT genes appeared to have the highest fructan or soluble sugar content and the strongest salt tolerance. Fructan is thought to be one of the important regulators involved in plant tolerance to various abiotic stresses. In this study, wheat-derived genes, Ta1-SST, Ta6-SFT, and Ta1-FFT, encoding fructan biosynthesis enzymes (FBE) were isolated and cloned into vectors modified pBI121 or pZP211. Seven different combinations of the three target genes were transformed into tobacco plants through an Agrobacterium-mediated approach, and transgenic tobacco plants were identified by PCR, ELISA, and Southern blotting. Compared with tobacco plants transformed with other six combinations of the three target genes and with wild-type plants, the transgenic plants transformed with Ta1-SST + Ta6-SFT genes contained the highest fructan and soluble sugar content. All seven types of transgenic tobacco plants displayed a much higher level of tolerance to drought, low temperature, and high salinity compared with the wild type. Differences of drought and low temperature tolerance between the transgenic plants containing a single FBE gene and those harboring two or three FBE genes were not significant, but the salt tolerance level of the transgenic plants with different FBE gene combinations from high to low was: Ta1-SST + Ta6-SFT > Ta1-SST + Ta6-SFT + Ta1-FFT > Ta1-SST + Ta1-FFT > Ta1-SFT + Ta1-FFT > single FBE gene. These results indicated that the tolerances of the transgenic tobacco plants to various abiotic stresses were associated with the transformed target gene combinations and the contents of fructan and soluble sugar contained in the transgenic plants.
DOI:10.1007/s10725-014-9908-xURL [本文引用: 1]
DOI:10.1046/j.1469-8137.2003.00858.xURL [本文引用: 1]
DOI:10.1093/jxb/erj065URLPMID:16449376 [本文引用: 2]
Inulin-type fructans are the simplest and most studied fructans and have become increasingly popular as prebiotic health-improving compounds. A natural variation in the degree of polymerization (DP) of inulins is observed within the family of the Asteraceae. Globe thistle (Echinops ritro), artichoke (Cynara scolymus), and Viguiera discolor biosynthesize fructans with a considerably higher DP than Cichorium intybus (chicory), Helianthus tuberosus (Jerusalem artichoke), and Dahlia variabilis. The higher DP in some species can be explained by the presence of special fructan: fructan 1-fructosyl transferases (high DP 1-FFTs), different from the classical low DP 1-FFTs. Here, the RT-PCR-based cloning of a high DP 1-FFT cDNA from Echinops ritro is described, starting from peptide sequence information derived from the purified native high DP 1-FFT enzyme. The cDNA was successfully expressed in Pichia pastoris. A comparison is made between the mass fingerprints of the native, heterodimeric enzyme and its recombinant, monomeric counterpart (mass fingerprints and kinetical analysis) showing that they have very similar properties. The recombinant enzyme is a functional 1-FFT lacking invertase and 1-SST activities, but shows a small intrinsic 1-FEH activity. The enzyme is capable of producing a high DP inulin pattern in vitro, similar to the one observed in vivo. Depending on conditions, the enzyme is able to produce fructo-oligosaccharides (FOS) as well. Therefore, the enzyme might be suitable for both FOS and high DP inulin production in bioreactors. Alternatively, introduction of the high DP 1-FFT gene in chicory, a crop widely used for inulin extraction, could lead to an increase in DP which is useful for a number of specific industrial applications. 1-FFT expression analysis correlates well with high DP fructan accumulation in vivo, suggesting that the enzyme is responsible for high DP fructan formation in planta.
.
URL [本文引用: 1]
果聚糖是以β(2→1)和/或β(2→6)糖苷键连接的线性或分支型果糖聚合物,通常在末端连接一个葡萄糖残基,是水溶性的非还原性多糖。果聚糖广泛存在于细菌、真菌以及超过40000种高等植物中。研究较多的是菊糖型果聚糖,是一个蔗糖以β(2→1)糖苷键与多个果糖相连的线性分子。 果聚糖作为一种功能性食品,不仅可以增强人体钙的吸收,还被用作低热量值的甜味剂、食品添加剂中的水溶性膳食纤维等,更重要的是,它不能被人体代谢吸收,可以作为一种益生素,因此成为近年研究的热点。 菊糖型果聚糖代谢中主要涉及两个关键酶:蔗糖-1-果糖基转移酶(1-SST,EC2.4.1.99)和果聚糖-1-果糖基转移酶(1-FFT,EC2.4.1.100)。1-SST催化蔗糖转变为蔗果三糖(DP=3),1-FFT负责糖链的延长,将蔗果三糖转变为高聚合度的果聚糖。 在前期研究中已将由莴苣(Lactuca sativa)中克隆得到的1-SST、1-FFT基因分别与玉米醇溶蛋白基因启动子结合,构建目标基因串联的植物表达载体,使用农杆菌介导的芽尖转化法转化玉米自交系,获得初步鉴定的转基因植株。在此基础上,本研究以T_2代转基因植株为材料,通过除草剂初步筛选和PCR检测鉴定转基因植株,进而通过RT-PCR筛选出目标基因表达较高的株系,通过连续自交获得转基因纯合系。Southern分析表明,在入选材料中编码这两个酶的基因在玉米基因组中是以单拷贝的形式存在。对转基因植株的生长发育进行系统观察,与未转基因对照植株相比,转基因植株未出现明显不同的变化。 对转基因表达强度较高的株系,使用TLC(薄板层析)分离和鉴定玉米籽粒可溶性糖提取液中的各种糖分,同时采用高效液相色谱结合示差折光检测器测定技术,对玉米籽粒糖提取液中的可溶性糖进行定量分析。结果表明,在玉米籽粒中合成了一定量的果聚糖,合成量在0.05~2.0mg/g范围。蔗糖、葡萄糖、果糖其他可溶性糖的含量与对照植株差异不明显。 对玉米籽粒的蛋白提取液进行酶活性测定,确定了最适底物及浓度为100mmol/L蔗糖,最适反应时间为3h。然后从3个转基因株系籽粒中分别提取酶蛋白,进行酶活性测定。结果显示,该提取液具有果糖基转移酶活性,能以蔗糖为底物,生成果聚糖。这表明向不合成果聚糖的植物导入1-SST和1-FFT基因能够合成高分子量果聚糖。 本研究通过对转基因植株糖组分的分析和酶活性的测定,证明转入玉米中的果糖基转移酶1-SST和1-FFT发挥了作用,得到了能在玉米胚乳中积累果聚糖的新种质。该工作对于研究植物糖代谢网络及其调控机制也有启示作用。
URL [本文引用: 1]
果聚糖是以β(2→1)和/或β(2→6)糖苷键连接的线性或分支型果糖聚合物,通常在末端连接一个葡萄糖残基,是水溶性的非还原性多糖。果聚糖广泛存在于细菌、真菌以及超过40000种高等植物中。研究较多的是菊糖型果聚糖,是一个蔗糖以β(2→1)糖苷键与多个果糖相连的线性分子。 果聚糖作为一种功能性食品,不仅可以增强人体钙的吸收,还被用作低热量值的甜味剂、食品添加剂中的水溶性膳食纤维等,更重要的是,它不能被人体代谢吸收,可以作为一种益生素,因此成为近年研究的热点。 菊糖型果聚糖代谢中主要涉及两个关键酶:蔗糖-1-果糖基转移酶(1-SST,EC2.4.1.99)和果聚糖-1-果糖基转移酶(1-FFT,EC2.4.1.100)。1-SST催化蔗糖转变为蔗果三糖(DP=3),1-FFT负责糖链的延长,将蔗果三糖转变为高聚合度的果聚糖。 在前期研究中已将由莴苣(Lactuca sativa)中克隆得到的1-SST、1-FFT基因分别与玉米醇溶蛋白基因启动子结合,构建目标基因串联的植物表达载体,使用农杆菌介导的芽尖转化法转化玉米自交系,获得初步鉴定的转基因植株。在此基础上,本研究以T_2代转基因植株为材料,通过除草剂初步筛选和PCR检测鉴定转基因植株,进而通过RT-PCR筛选出目标基因表达较高的株系,通过连续自交获得转基因纯合系。Southern分析表明,在入选材料中编码这两个酶的基因在玉米基因组中是以单拷贝的形式存在。对转基因植株的生长发育进行系统观察,与未转基因对照植株相比,转基因植株未出现明显不同的变化。 对转基因表达强度较高的株系,使用TLC(薄板层析)分离和鉴定玉米籽粒可溶性糖提取液中的各种糖分,同时采用高效液相色谱结合示差折光检测器测定技术,对玉米籽粒糖提取液中的可溶性糖进行定量分析。结果表明,在玉米籽粒中合成了一定量的果聚糖,合成量在0.05~2.0mg/g范围。蔗糖、葡萄糖、果糖其他可溶性糖的含量与对照植株差异不明显。 对玉米籽粒的蛋白提取液进行酶活性测定,确定了最适底物及浓度为100mmol/L蔗糖,最适反应时间为3h。然后从3个转基因株系籽粒中分别提取酶蛋白,进行酶活性测定。结果显示,该提取液具有果糖基转移酶活性,能以蔗糖为底物,生成果聚糖。这表明向不合成果聚糖的植物导入1-SST和1-FFT基因能够合成高分子量果聚糖。 本研究通过对转基因植株糖组分的分析和酶活性的测定,证明转入玉米中的果糖基转移酶1-SST和1-FFT发挥了作用,得到了能在玉米胚乳中积累果聚糖的新种质。该工作对于研究植物糖代谢网络及其调控机制也有启示作用。
[本文引用: 1]
URL [本文引用: 1]
DOI:10.1109/38.79453URL [本文引用: 1]
Fructan: fructan fructosyl transferase (FFT, EC 2.4.1.100) was purified from chicory (Cichorium intybus L. var. foliosum cv. Flash) roots by a combination of ammonium sulfate precipitation, concanavalin A affinity chromatography, and anion- and cation-exchange chromatography. This protocol produced a 60-fold purification and a specific activity of 14.5 mu mol .(mg protein)(-1) . min(-1). The mass of the enzyme was 69 kDa as estimated by gel filtration. On sodium dodecyl sulfate-polyacrylamide gel electrophoresis and mass spectrometry, 52-kDa and 17-kDa fragments were found, suggesting that the enzyme was a heterodimer. Optimal activity was found between pH 5.5 and 6.5. The enzyme used 1-kestose, 1,1-nystose, oligofructan and commercial chicory root inulin (degree of polymerization greater than or equal to 10) as donors and accepters. Sucrose was the best acceptor but could not be used as a donor. However, at higher concentrations sucrose acted as a competitive inhibitor for donors of FFT. 1-Kestose was the most efficient and 1,1-nystose the least efficient donor. The purified enzyme exhibited beta-fructosidase activity, specially at higher temperatures and lower substrate concentrations. The synthesis of fructans from 1-kestose decreased at higher temperatures (5-50 degrees C). Therefore enzyme assays were performed at 0 degrees C. The same fructan oligosaccharides, with a distribution similar to that observed in vivo, were obtained upon incubation of the enzyme with sucrose and commercial chicory root inulin. [References: 26]
DOI:10.1111/j.1399-3054.2005.00579.xURL [本文引用: 1]
Inulin- type fructans are stored in the tuberous roots of the Brazilian cerrado plant Viguiera discolor Baker ( Asteraceae). In Cynara scolymus ( artichoke) and Echinops ritro ( globe thistle), the fructans have a considerably higher degree of polymerization ( DP) than in Cichorium intybus ( chicory) and Helianthus tuberosus ( Jerusalem artichoke). It was shown before that the higher DP in some species can be attributed to the properties of their fructan: fructan 1- fructosyl transferases ( 1- FFTs; EC 2.4.1.100), enzymes responsible for chain elongation. Here, we describe the cloning of a high DP ( hDP) 1- FFT cDNA from V. discolor and its heterologous expression in Pichia pastoris. Starting from 1- kestose and Neosugar P ( a mixture of oligo- inulins from microbial origin) as substrates, the recombinant enzyme produces a typical hDP inulin profile in vitro, closely resembling the one observed in vivo. The enzyme shows no invertase activity and sucrose: sucrose 1- fructosyl transferase ( 1- SST; EC 2.4.1.99) activity in vitro. Pattern evolution during incubation suggests that inulins with DP >= 6 are much better substrates than sucrose or lower DP oligo- fructans. Because hDP inulin- type fructans show superior properties for specific food and non- food applications, the hDP 1- FFT gene from V. discolor has potential for the production of hDP inulin in vitro or in transgenic crops.
DOI:10.1016/S1671-2927(09)60099-7URL [本文引用: 4]
DOI:10.11686/cyxb20110116URLMagsci [本文引用: 1]
以多年生黑麦草(品种卡特)胚性愈伤组织为转化受体,利用农杆菌介导的遗传转化方法将冰草果聚糖:果聚糖-1-果糖基转移酶基因(<em>Ac</em>1-<em>FFT</em>)导入黑麦草中,对再生植株喷洒basta溶液和PCR 法检测,共获得18个阳性株系,RT-PCR 结果表明,该基因在转基因黑麦草中正常表达。转基因黑麦草株系中的可溶性总糖含量和果聚糖含量明显高于对照植株,耐旱性提高,干旱胁迫6d时其相对含水量和叶绿素含量明显高于对照植株,且下降速度慢,但其电解质渗漏率和丙二醛含量显著低于对照植株,复水后很快复原,而对照植株无法恢复,说明转基因植株中由于干旱处理发生的损伤是可逆的,而对照植株中的损伤是非可逆的。以上结果表明,转基因黑麦草中犃<em>Ac</em>1-<em>FFT</em>的表达及果聚糖合成可能是其耐旱性提高的最重要原因。
DOI:10.11686/cyxb20110116URLMagsci [本文引用: 1]
以多年生黑麦草(品种卡特)胚性愈伤组织为转化受体,利用农杆菌介导的遗传转化方法将冰草果聚糖:果聚糖-1-果糖基转移酶基因(<em>Ac</em>1-<em>FFT</em>)导入黑麦草中,对再生植株喷洒basta溶液和PCR 法检测,共获得18个阳性株系,RT-PCR 结果表明,该基因在转基因黑麦草中正常表达。转基因黑麦草株系中的可溶性总糖含量和果聚糖含量明显高于对照植株,耐旱性提高,干旱胁迫6d时其相对含水量和叶绿素含量明显高于对照植株,且下降速度慢,但其电解质渗漏率和丙二醛含量显著低于对照植株,复水后很快复原,而对照植株无法恢复,说明转基因植株中由于干旱处理发生的损伤是可逆的,而对照植株中的损伤是非可逆的。以上结果表明,转基因黑麦草中犃<em>Ac</em>1-<em>FFT</em>的表达及果聚糖合成可能是其耐旱性提高的最重要原因。
DOI:10.1016/j.jplph.2005.03.014URLPMID:16455352 [本文引用: 2]
This study aimed to investigate the effect of water stress on the accumulation of proline in the leaves of transgenic potato lines generating fructans. The results of the series of bifactorial experiments in 2002 and 2003 indicated an increase of the proline level in leaves of all potato lines examined under water deficit. In addition, an increase in proline concentration during plant development was observed. The proline content was related to leaf water potential and relative water content (RWC), which indicates that proline could be involved in osmoregulation of potato plants under the experimental conditions. Surprisingly, under water deficit, the proline level was lowest in most of the transgenic SST/FFT-lines, which generate fructan molecules with a high degree of polymerization. Therefore, a pleiotropic effect can not be excluded as the reason for the divergence in behavior of these transgenic lines. The present results suggest that the modification of carbohydrate metabolism, especially the high content of soluble carbohydrates, may affect water stress-induced proline accumulation.
[本文引用: 1]
In: Jacquot J, Delseny M, eds.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
簇毛麦属(Dasypyrum原作Haynaldia)包括二倍体一年生簇毛麦(D. villosum,染色体组为VV),二倍体多年生簇毛麦(D. breviaristatum,染色体组为VbVb)和四倍体多年生簇毛麦(D. breviaristatum,染色体组为VbVbVbVb)。一年生簇毛麦主要分布在地中海沿岸,而多年生簇毛麦主要分布在西北非的山区,希腊和摩洛哥。在植物分类上属禾本科(Gramineae)小麦族(Triticeae)的小麦亚族(Triticinae),是小麦的野生近缘种,具有抗白粉病、抗叶锈病和秆锈、抗全蚀病、分蘖力强、小穗数多、耐旱耐寒、耐盐和蛋白质含量高等许多栽培小麦所需要的优良性状,是小麦高产、抗病、优质育种的重要遗传资源。 迄今为止,对簇毛麦属物种研究仍有如下不足:(1)四倍体多年生簇毛麦在该属中的遗传进化关系一直存在争议。特别是对四倍体多年生簇毛麦与二倍体多年生簇毛麦的染色体组关系研究上,缺乏分子和细胞学证据。(2)可在育种中利用的簇毛麦染色体组(臂)特异分子标记不能满足小麦-簇毛麦新种质鉴定的需求,需要进一步挖掘和开发。(3)我们在开展将四倍体多年生簇毛麦抗白粉和抗条锈基因导入小麦的研究工作中,育成了一批小麦-四倍体多年生簇毛麦抗病渐渗系,但是这些抗病渐渗系中外源染色质的同源群关系尚未明确。(4)涉及二倍体一年生簇毛麦1V、4V和6V染色体的抗病(或高产)易位系已经育成,但是涉及2V、3V、5V和7V染色体的易位系还未见报道。 本文着重解决上述4个问题。本文对四倍体多年生簇毛麦在簇毛麦属的遗传进化关系进行了深入研究;创制并用建立的多个EST-STS标记和PLUG标记鉴定了小麦-四倍体簇毛麦附加系;诱导和鉴定了小麦-二倍体一年生簇毛麦2V、3V、5V和7V补偿性罗伯逊易位系。研究结果如下: 1.簇毛麦属物种的遗传进化关系:(1)利用RAPD技术对二倍体一年生簇毛麦、四倍体多年生簇毛麦和小麦族黑麦、中间偃麦草、拟鹅观草和黑麦等物种进行了聚类分析,结果表明二倍体一年生簇毛麦和四倍体多年生簇毛麦不能聚在一起,表明四倍体多年生簇毛麦不是由二倍体一年生簇毛麦加倍而来。(2)吉姆萨C分带结果显示,四倍体多年生簇毛麦与二倍体多年生簇毛麦具有较多的中间带,这些带纹多属弱带型,并且二者的带型还有所差异;而二倍体一年生簇毛麦具有较多的端带,这些带纹多属强带型。因此,C分带的证据只能粗略证明四倍体多年生簇毛麦与二倍体多年生簇毛麦遗传关系较近,而与二倍体一年生簇毛麦相对较远。(3)克隆了簇毛麦属三物种的叶绿体infA基因和线粒体trnfM及rrn18基因,并以此对这些基因序列分别进行了bootstrap分析。分析结果支持四倍体多年生簇毛麦是由二倍体多年生簇毛麦加倍而来的观点。(4)基因组原位杂交和利用核糖体基因pTa71为探针的原位杂交清楚的表明四倍体多年生簇毛麦是由二倍体多年生簇毛麦加倍产生的。(5)以黑麦基因组重复序列pSc74为探针的原位杂交表明,从二倍体多年生簇毛麦加倍到四倍体多年生簇毛麦的过程中,不仅发生了染色体的重排,还发生了基因组DNA水平上的重组现象。 2.簇毛麦基因组或单染色体分子标记:(1)分离得到簇毛麦基因组1个新的重复序列pDb12H(或称为OPH12),该重复序列和大麦基因组Sabrina反转录重复序列具有95%的同源性。根据pDb12H设计引物,对含簇毛麦染色质的材料和大量对照材料进行扩增,建立了簇毛麦基因组SCAR标记,该标记被用来筛选小麦-四倍体多年生簇毛麦抗病渐渗系和小麦-二倍体一年生簇毛麦易位系。以pDb12H为探针对小麦族大量物种进行原位杂交,结果表明,pDb12H仅分布在小麦族二倍体一年生簇毛麦V基因组、多年生簇毛麦Vb基因组、大麦H基因组和中间偃麦草Js基因组上,即pDb12H高拷贝存在于V基因组,Vb基因组,H基因组和Js基因组上,而寡拷贝存在于小麦族其他基因组上,Southern blot也证实了上述结果。根据FISH和Southern blot结果推测pDb12H在小麦族各基因组(V、Vb、H和JS基因组除外)分化前期可能就失去了转座活性。Kishii等曾用原位杂交和分子标记手段证实百萨偃麦草J基因组和黑麦R基因组参与了中间偃麦草Js基因组的形成,认为中间偃麦草基因组应为StJs(V-J-R)s。本文发现大麦H基因组也参与了中间偃麦草基因组进化,因此我们建议将中间偃麦草基因组暂时补充修订为StJs(V-J-R-H)s。(2)利用小麦第1到第7同源群的943对EST-STS引物对四倍体多年生簇毛麦和中国春进行筛选,PCR产物利用HaeIII、MspI、RsaI和AluI四种酶进行酶切,获得了474个STS标记。同时还筛选了小麦第1到第7同源群的56对PLUG引物,利用TaqI和HaeIII两种酶对PCR产物进行酶切,筛选到6个PLUG标记。 3.小麦-四倍体多年生簇毛麦附加系的鉴定:利用建立的SCAR分子标记对小麦-四倍体多年生簇毛麦抗病渐渗系进行初步筛选,然后利用C分带对筛选出的渐渗系进行普查,结果显示这批抗病渐渗系中只有2种不同类型的附加系Y93-1-6-1和Y93-1-A6-4。利用上述筛选到的STS标记和PLUG标记对Y93-1-6-1和Y93-1-A6-4进行鉴定。首次发现Y93-1-6-1和Y93-1-A6-4中四倍体簇毛麦染色体均发生了重组。附加系Y93-1-6-1中四倍体簇毛麦染色体长臂为6VbL,其短臂发生了6VbS和7VbS染色体片段的重组;而附加系Y93-1-A6-4中四倍体簇毛麦染色体发生了更为复杂的染色体重组,其短臂为6VbS和7VbS的重组,长臂为1VbL、2VbL和7VbL的重组。 4.小麦-二倍体一年生簇毛麦易位系的创制与鉴定:利用中国春-二倍体一年生簇毛麦附加系与相应的中国春单体进行杂交获得双单体,然后双单体进行自交,对所获得的自交种子进行筛选获得相关易位系。(1)筛选来自双单体40+2V+2D的自交后代660个单株,获得4个杂合T2VL.2DS易位系,3个杂合T2VS.2DL易位系。(2)筛选来自双单体40+3V+3D的自交后代496个单株,获得5个T3DS.3VL易位系,3个杂合T3DL.3VS易位系。(3)筛选来自双单体40+5V+5D的自交后代342个单株,获得12个杂合的T5DL.5VS易位系,未发现T5DS.5VL易位系。(4)筛选来自双单体40+7V+7D的自交后代384个单株,获得6个杂合T7DS.7VL易位系,2个杂合T7DL.7VS易位系。对上述杂合易位系的自交种子进行筛选,目前都已获得了纯合的易位系,这些优异的种质资源可用于小麦育种。综上,本论文对簇毛麦属种间的进化关系进行了系统研究,对簇毛麦特异分子标记进行了进一步挖掘和开发并利用开发的分子标记对新创制的种质进行了鉴定。
URL [本文引用: 2]
簇毛麦属(Dasypyrum原作Haynaldia)包括二倍体一年生簇毛麦(D. villosum,染色体组为VV),二倍体多年生簇毛麦(D. breviaristatum,染色体组为VbVb)和四倍体多年生簇毛麦(D. breviaristatum,染色体组为VbVbVbVb)。一年生簇毛麦主要分布在地中海沿岸,而多年生簇毛麦主要分布在西北非的山区,希腊和摩洛哥。在植物分类上属禾本科(Gramineae)小麦族(Triticeae)的小麦亚族(Triticinae),是小麦的野生近缘种,具有抗白粉病、抗叶锈病和秆锈、抗全蚀病、分蘖力强、小穗数多、耐旱耐寒、耐盐和蛋白质含量高等许多栽培小麦所需要的优良性状,是小麦高产、抗病、优质育种的重要遗传资源。 迄今为止,对簇毛麦属物种研究仍有如下不足:(1)四倍体多年生簇毛麦在该属中的遗传进化关系一直存在争议。特别是对四倍体多年生簇毛麦与二倍体多年生簇毛麦的染色体组关系研究上,缺乏分子和细胞学证据。(2)可在育种中利用的簇毛麦染色体组(臂)特异分子标记不能满足小麦-簇毛麦新种质鉴定的需求,需要进一步挖掘和开发。(3)我们在开展将四倍体多年生簇毛麦抗白粉和抗条锈基因导入小麦的研究工作中,育成了一批小麦-四倍体多年生簇毛麦抗病渐渗系,但是这些抗病渐渗系中外源染色质的同源群关系尚未明确。(4)涉及二倍体一年生簇毛麦1V、4V和6V染色体的抗病(或高产)易位系已经育成,但是涉及2V、3V、5V和7V染色体的易位系还未见报道。 本文着重解决上述4个问题。本文对四倍体多年生簇毛麦在簇毛麦属的遗传进化关系进行了深入研究;创制并用建立的多个EST-STS标记和PLUG标记鉴定了小麦-四倍体簇毛麦附加系;诱导和鉴定了小麦-二倍体一年生簇毛麦2V、3V、5V和7V补偿性罗伯逊易位系。研究结果如下: 1.簇毛麦属物种的遗传进化关系:(1)利用RAPD技术对二倍体一年生簇毛麦、四倍体多年生簇毛麦和小麦族黑麦、中间偃麦草、拟鹅观草和黑麦等物种进行了聚类分析,结果表明二倍体一年生簇毛麦和四倍体多年生簇毛麦不能聚在一起,表明四倍体多年生簇毛麦不是由二倍体一年生簇毛麦加倍而来。(2)吉姆萨C分带结果显示,四倍体多年生簇毛麦与二倍体多年生簇毛麦具有较多的中间带,这些带纹多属弱带型,并且二者的带型还有所差异;而二倍体一年生簇毛麦具有较多的端带,这些带纹多属强带型。因此,C分带的证据只能粗略证明四倍体多年生簇毛麦与二倍体多年生簇毛麦遗传关系较近,而与二倍体一年生簇毛麦相对较远。(3)克隆了簇毛麦属三物种的叶绿体infA基因和线粒体trnfM及rrn18基因,并以此对这些基因序列分别进行了bootstrap分析。分析结果支持四倍体多年生簇毛麦是由二倍体多年生簇毛麦加倍而来的观点。(4)基因组原位杂交和利用核糖体基因pTa71为探针的原位杂交清楚的表明四倍体多年生簇毛麦是由二倍体多年生簇毛麦加倍产生的。(5)以黑麦基因组重复序列pSc74为探针的原位杂交表明,从二倍体多年生簇毛麦加倍到四倍体多年生簇毛麦的过程中,不仅发生了染色体的重排,还发生了基因组DNA水平上的重组现象。 2.簇毛麦基因组或单染色体分子标记:(1)分离得到簇毛麦基因组1个新的重复序列pDb12H(或称为OPH12),该重复序列和大麦基因组Sabrina反转录重复序列具有95%的同源性。根据pDb12H设计引物,对含簇毛麦染色质的材料和大量对照材料进行扩增,建立了簇毛麦基因组SCAR标记,该标记被用来筛选小麦-四倍体多年生簇毛麦抗病渐渗系和小麦-二倍体一年生簇毛麦易位系。以pDb12H为探针对小麦族大量物种进行原位杂交,结果表明,pDb12H仅分布在小麦族二倍体一年生簇毛麦V基因组、多年生簇毛麦Vb基因组、大麦H基因组和中间偃麦草Js基因组上,即pDb12H高拷贝存在于V基因组,Vb基因组,H基因组和Js基因组上,而寡拷贝存在于小麦族其他基因组上,Southern blot也证实了上述结果。根据FISH和Southern blot结果推测pDb12H在小麦族各基因组(V、Vb、H和JS基因组除外)分化前期可能就失去了转座活性。Kishii等曾用原位杂交和分子标记手段证实百萨偃麦草J基因组和黑麦R基因组参与了中间偃麦草Js基因组的形成,认为中间偃麦草基因组应为StJs(V-J-R)s。本文发现大麦H基因组也参与了中间偃麦草基因组进化,因此我们建议将中间偃麦草基因组暂时补充修订为StJs(V-J-R-H)s。(2)利用小麦第1到第7同源群的943对EST-STS引物对四倍体多年生簇毛麦和中国春进行筛选,PCR产物利用HaeIII、MspI、RsaI和AluI四种酶进行酶切,获得了474个STS标记。同时还筛选了小麦第1到第7同源群的56对PLUG引物,利用TaqI和HaeIII两种酶对PCR产物进行酶切,筛选到6个PLUG标记。 3.小麦-四倍体多年生簇毛麦附加系的鉴定:利用建立的SCAR分子标记对小麦-四倍体多年生簇毛麦抗病渐渗系进行初步筛选,然后利用C分带对筛选出的渐渗系进行普查,结果显示这批抗病渐渗系中只有2种不同类型的附加系Y93-1-6-1和Y93-1-A6-4。利用上述筛选到的STS标记和PLUG标记对Y93-1-6-1和Y93-1-A6-4进行鉴定。首次发现Y93-1-6-1和Y93-1-A6-4中四倍体簇毛麦染色体均发生了重组。附加系Y93-1-6-1中四倍体簇毛麦染色体长臂为6VbL,其短臂发生了6VbS和7VbS染色体片段的重组;而附加系Y93-1-A6-4中四倍体簇毛麦染色体发生了更为复杂的染色体重组,其短臂为6VbS和7VbS的重组,长臂为1VbL、2VbL和7VbL的重组。 4.小麦-二倍体一年生簇毛麦易位系的创制与鉴定:利用中国春-二倍体一年生簇毛麦附加系与相应的中国春单体进行杂交获得双单体,然后双单体进行自交,对所获得的自交种子进行筛选获得相关易位系。(1)筛选来自双单体40+2V+2D的自交后代660个单株,获得4个杂合T2VL.2DS易位系,3个杂合T2VS.2DL易位系。(2)筛选来自双单体40+3V+3D的自交后代496个单株,获得5个T3DS.3VL易位系,3个杂合T3DL.3VS易位系。(3)筛选来自双单体40+5V+5D的自交后代342个单株,获得12个杂合的T5DL.5VS易位系,未发现T5DS.5VL易位系。(4)筛选来自双单体40+7V+7D的自交后代384个单株,获得6个杂合T7DS.7VL易位系,2个杂合T7DL.7VS易位系。对上述杂合易位系的自交种子进行筛选,目前都已获得了纯合的易位系,这些优异的种质资源可用于小麦育种。综上,本论文对簇毛麦属种间的进化关系进行了系统研究,对簇毛麦特异分子标记进行了进一步挖掘和开发并利用开发的分子标记对新创制的种质进行了鉴定。
DOI:10.1023/A:1004095705460URL [本文引用: 1]
Dasypyrum villosum (L.) Candargy (DV) is adiploid (2n = 14, VV genomes), allogamous grass of theMediterranean region. It may be hybridized with wheatand is thus a gene resource for wheat improvement. Westudied grain protein concentration andSDS-sedimentation (SED) as indicators of end-usequality. The latter is a good predictor of glutenstrength. A-PAGE and SDS-PAGE were used to identifymonomeric and polymeric seed storage proteins,respectively, to relate proteins of DV to those foundin Chinese Spring (CS), Triticum aestivum L.,wheat. Two full-sib lines of DV had high grain protein(19.3 and 20.3%), but one had very low mean SED (69mm) and one had very high (118 mm) based on onegreenhouse and one field test. CS had very low grainprotein (12.0%) and weak gluten (33 mm). Single-DVchromosome addition and substitution lines and twoDV-wheat recombinant lines all had higher grainprotein than CS (range 13.9 to 16.7%). SED valuesshowed a different pattern. CS=4V and CS=6V hadlow SED, 63 and 44 mm, similar to CS, whereas CS=1Vand full sib DV 200 had very strong gluten, 118 mm, asdid substitution lines CS 1V (1A) and CS 1V (1B), 125and 131 mm, respectively. One hybrid-derived line withDV-wheat 1V recombinant chromosome had SED of 99 mmand one line with a 6V added chromosome had SED of 64mm. The large positive effects of quality in the wheathaving DV chromosome 1V are believed to be due to DValleles at the Glu-V1 and Gli-V1/Glu-V3 loci. DV chromosomes 4V and 6V did not contribute toimproved quality probably due to Gli-V2 and Gli-V3 which, as the orthologous loci in wheat, donot enhance wheat quality. Based on the positiveeffects of alleles on DV chromosome 1V in a breadwheat background, we conclude that D. villosum is a source of allelic diversity that can beconsidered for improving end-use quality in breadwheat.
DOI:10.2135/cropsci1981.0011183X002100050018xURL [本文引用: 1]
To determine the potential of various species of wheatgrasses (Elytrigia Desv.) as gene sources for the improvement of salt tolerance in wheat (Triticum aestivum L. em. Thell.), six wheat accessions, previously identified as salt-tolerant, and 36 wheatgrass accessions, representing 13 species, were grown from the seedling stage. Plants were grown in gradually increasing concentrations of NaCl in hydroculture with one-half strength Hoagland's solution. Tolerance to the NaCl was determined for the wheat plants by scoring survival in 250 mM NaCl and for the wheatgrass plants by scoring survival in 500 mM NaCl and in 750 mM NaCl. while no wheat plants survived the stress with 250 nM NaCl, several wheatgrass accessions had large percentages of surviving individuals in 750 MM NaCl. The four most tolerant species were: Elytrigia scirpea (Presl) Holub, E. pontica (Podp.) Holub, E. junceiformis Love et Love, and E. diae (Runemark) nom. nud. Since these species are also among those most easily hybridizable with wheat, they are the best candidates for sources of salt tolerance.
DOI:10.1104/pp.107.1.33URLPMID:12228347 [本文引用: 1]
Abstract Fructans are polyfructose molecules produced by approximately 15% of the flowering plant species. It is possible that, in addition to being a storage carbohydrate, fructans have other physiological roles. Owing to their solubility they may help plants survive periods of osmotic stress induced by drought or cold. To investigate the possible functional significance of fructans, use was made of transgenic tobacco (Nicotiana tabacum) plants that accumulate bacterial fructans and hence possess an extra sink for carbohydrate. Biomass production was analyzed during drought stress with the use of lines differing only in the presence of fructans. Fructan-producing tobacco plants performed significantly better under polyethylene-glycol-mediated drought stress than wild-type tobacco. The growth rate of the transgenic plants was significantly higher (+55%), as were fresh weight (+33%) and dry weight (+59%) yields. The difference in weight was observed in all organs and was particularly pronounced in roots. Under unstressed control conditions the presence of fructans had no significant effect on growth rate and yield. Under all conditions the total nonstructural carbohydrate content was higher in the transgenic plants. We conclude that the introduction of fructans in this non-fructan-producing species mediates enhanced resistance to drought stress.
DOI:10.1128/AEM.67.1.363-370.2001URLPMID:11133467 [本文引用: 1]
We have purified a fructosyltransferase from conidia of the inulin-producing fungus Aspergillus sydowi IAM 2544 and obtained peptide sequences from proteolytic fragments of the protein. With degenerated primers, we amplified a PCR fragment that was used to screen a cDNA library. The fructosyltransferase gene from Aspergillus sydowi (EMBL accession no. AJ289046) is expressed in conidia, while no expression could be detected in mycelia by Northern blot analysis of mycelial RNA. The gene encodes a protein with a calculated molecular mass of 75 kDa that is different from all fructosyltransferases in the databases. The only homology that could be detected was to the invertase of Aspergillus niger (EMBL accession no. L06844). The gene was functionally expressed in Escherichia coli, yeast, and potato plants. With protein extracts from transgenic bacteria and yeast, fructooligosaccharides could be produced in vitro. In transgenic potato plants, inulin molecules of up to 40 hexose units were synthesized in vivo. While in vitro experiments with protein extracts from conidia of Aspergillus sydowi yielded the same pattern of oligosaccharides as extracts from transformed bacteria and yeast, in vivo inulin synthesis with fungal conidia leads to the production of a high-molecular-weight polymer.
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1515/bchm2.1951.286.1-6.270URLPMID:14907695 [本文引用: 2]
FALES FW.
[本文引用: 1]
.
[本文引用: 2]
DOI:10.1104/pp.108.125559URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11248-013-9776-6URLPMID:24398893 [本文引用: 1]
The cultivation of rice ( Oryza sativa L.), a major food crop, requires ample water (30 % of the fresh water available worldwide), and its productivity is greatly affected by drought, the most significant environmental factor. Much research has focussed on identifying quantitative trait loci, stress-regulated genes and transcription factors that will contribute towards the development of climate-resilient/tolerant crop plants in general and rice in particular. The transcription factor DREB1A , identified from the model plant Arabidopsis thaliana , has been reported to enhance stress tolerance against drought stress. We developed transgenic rice plants with AtDREB1A in the background of indica rice cultivar Samba Mahsuri through Agrobacterium -mediated transformation. The AtDREB1A gene was stably inherited and expressed in T 1 and T 2 plants and in subsequent generations, as indicated by the results of PCR, Southern blot and RT-PCR analyses. Expression of AtDREB1A was induced by drought stress in transgenic rice lines, which were highly tolerant to severe water deficit stress in both the vegetative and reproductive stages without affecting their morphological or agronomic traits. The physiological studies revealed that the expression of AtDREB1A was associated with an increased accumulation of the osmotic substance proline, maintenance of chlorophyll, increased relative water content and decreased ion leakage under drought stress. Most of the homozygous lines were highly tolerant to drought stress and showed significantly a higher grain yield and spikelet fertility relative to the nontransgenic control plants under both stressed and unstressed conditions. The improvement in drought stress tolerance in combination with agronomic traits is very essential in high premium indica rice cultivars, such as Samba Mahsuri, so that farmers can benefit in times of seasonal droughts and water scarcity.
DOI:10.1093/jxb/erm367URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}