 ,1,2,*1
,1,2,*1 2
Alleviation Effects of Exogenous Growth Regulators on Seed Germination of Sweet Sorghum under Salt Stress and Its Physiological Basis
ZHU Guang-Long1,**, SONG Cheng-Yu2,**, YU Lin-Lin2, CHEN Xu-Bing2, ZHI Wen-Fang2, LIU Jia-Wei2, JIAO Xiu-Rong1, ZHOU Gui-Sheng,1,2,*12
ͨѶ����:
��һ��ϵ��:
�ո�����:2017-11-21��������:2018-08-20�����������:2018-09-04
| ��������: |
Received:2017-11-21Accepted:2018-08-20Online:2018-09-04
| Fund supported: |

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1387KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
�����, �γ���, ������, ������, ���ķ�, ������, ������, �ܹ���. ��Դ�����������ʶ�����������ȷ��������η�в�ȵĻ���ЧӦ������������[J]. ����ѧ��, 2018, 44(11): 1713-1724. doi:10.3724/SP.J.1006.2018.01713
ZHU Guang-Long, SONG Cheng-Yu, YU Lin-Lin, CHEN Xu-Bing, ZHI Wen-Fang, LIU Jia-Wei, JIAO Xiu-Rong, ZHOU Gui-Sheng.
Ŀǰ�����μ����Ӿ�, �ѳ�Ϊȫ�����̬����Դ����[1,2]���ҹ��μ�ش�1��108 hm2 [3], �������������[4], ����Լ4/5δ�ܿ�������, ����μ�صĸ������ۺϿ��������ѳ�Ϊũҵ����������������[5]����ֲ���������ǿ��������μ�ص���Ч;��֮һ�������(Sorghum bicolor [L.] Moench)��һ�����˵����ϡ����Ϻ���Դ����[6,7], ��ԭ�����������ȸ������ƶ�ķ����ȴ���½, ���п��������ԡ����¡����μ����������Ч�ʸߵ�����[8]��Ȼ��, �������ηָ���0.3% ʱ, ��������ӵ��ȷ��������ܵ�Ӱ��, ��������[9]�����, �η�в���½������������ȷ����������ѵȻ���������, ���ƹ���ֲ������Ϳ��������μ����Դ��ǰ�ᡣ
Ŀǰ, ������������η�в����Ӧ������о���Ҫ����������ˮƽ�ļ���������������Դ��ɸѡ�������ȷ����������������е����������仯, ��ʩ���ػ����ֶ����������в����������������Ӱ��ȷ��档�߽����[10]��66�ݸ���������Դ�������Լ�������, �յ�����ĸ���Ʒ�������Խ�ǿ, ������ϵ��ĸ������϶��ηֽ�Ϊ���С��ڲ�ͬŨ���η�в����, ������������ȷ����ܵ�����, �������½�, ��������[11]����������, �η�в�ȵ���������������˺���в��, �ƻ�ϸ��Ĥ�ṹ����������ʡ��ƻ�����ƽ���[12,13]����в�����������ȷ���������������Ҫ����������в����ɵ������ɺ�[14]����ʩ����[15]��ֲ���������ڼ�[16,17]�ȿ����������ηֵ�Σ�������ӵ��ȷ��ͳ���������Ը������Ӫ��������, �Ե����ס��طʵ����ս���[18,19,20,21,22], ��ʩ����Դ�����������ʺ����������ʿ���Ч�����������η�в�ȵ��µ�ˮ�ƽ��͡����Ӷ�����Ӱ��[23]�������о���������ʩ��һ��Դ���ʶ���в�ȵ�Ӱ��, �����ٹ��ڲ�ͬ������Դ�����������ʶ��η�в�ȵĻ���ЧӦ����ػ��ơ�
������ǰ���о��Ļ�����, ��ʩ��-��������(GABA)����ù��(GA3)��������(KT)��ˮ����(SA) 4��������������, �����η�в���²�ͬ�����������ʶ���������ӵ��ȷ����������������Եĵ���ЧӦ, Ϊ��ʾֲ����κ����ƺ�̽����Դ�������ʻ����η�в�ȵ����û����ṩ�ο���
1 �����뷽��
1.1 �������
������2016�������ݴ�ѧ����ʡ�������������ص�ʵ���Һͽ�����ũҵ��ũ��Ʒ��ȫ���ʺ�������ʵ���ҽ��С�ǰ��������2�����ε������Ʒ�ֹ���2011����106��Ϊ������Ͻ��������Լ������顣Ȼ���������������Խ����еĹ���106Ϊ�Բ�, ������Դ�����������ʶ������Ե���ЧӦ���о���������ñ������ֻ��ص����ջ������(��ѿ��>85%)������Һ��NaCl (������)������ˮ���ö���, �����Լ�����������5���η��ݶ�Ũ��, ��0��50��100��150��200 mmoL L-1; �����������ʵ����������� 4���η�Ũ���ݶ�, 0��50��100��150 mmoL L-1��
ѡȡ��������Сһ�µ�����, ��1%�Ĵ���������Һ����10 min, ����ˮ������ϴ�ɾ�, ��ɱ��á�ѡ�ھ�15 cm�Ĵ��Dz���������, ����ֱ��Ϊ15 cm����ֽ(˫Ȧ��, �����ֻ���ֽ����˾, �й�), ÿ����ȷ�������100��, ע��Hoagland[10]Ӫ��Һ50 mL, ÿ������3���ظ����������зֱ������Ӧ��NaCl����Һ10 mL, Ȼ������������������Ϊ12 h/12 h (��/ҹ)���¶�Ϊ25���������, ���趨��ʱ�俪�Ǽ�¼������ˮ���ȷ���������������ÿ��24 h��һ�δ���Һ�Ա�֤�������η�Ũ�ȵ�һ���ԡ�
ѡȡ500 ��mol L-1 ��-��������(GABA)��300 ��mol L-1��ù��(GA3)��232.3 ��mol L-1 ������(KT)��362 ��mol L-1 ˮ����(SA) 4����Դ�����������ʽ���������������Щ����Ũ����Ԥ����ɸѡ�ó���25�������¸������������ʽ�������12 h, ���ֺ������̳�, ������ˮ��ϴ3��, ������ˮֽ����, ������¼ԭʼ������Ȼ��ͬ������Դ���������������������ӷ��벻ͬŨ���ݶȵ�NaCl��Һ��, ��������ͬ���������Լ������顣
1.2 �ⶨ����
���鿪ʼ��4~48 h�ڼ�, ��ʱ�ⶨ���ӵ���ˮ��, ��ÿ���������е�����ȡ��, ����ˮֽ��ȥ���ӱ����ˮ�ֺ����, ������ˮ�ʺ���ˮ���ʡ�ÿ��12 h����������ķ�ѿ������ѿ�ơ���ѿ�ʡ����ӷ�ѿָ��������κ��ʺ����ӷ�ѿ����κ��ʵ�ָ�ꡣͬʱ, ���α꿨�߲����߸�����ѿ���ȡ����ȷ���5��, ѡ��������(����ѿ)�ⶨ���������ʺ����Ϳ�����ø���ԡ�1.2.1 �����ȷ�ָ�� ������ˮ��Yt (%) = [(Xt-X0)/X0]��100; ������ˮ����(%h-1) = (Xt-Xt-1)/ [t-(t-1)], ʽ��, X0Ϊ100����������ӵ�ԭʼ����(g); XtΪ������е�tСʱ100�����ӵ�����(g)��
���ӷ�ѿ��(%) = ��7�췢ѿ������/������������100; ���ӷ�ѿ��(%) = ��3�����ӷ�ѿ��/������������100; ���ӷ�ѿָ��(GI) = ��Gt /Dt , ʽ��, GtΪtʱ��ķ�ѿ��, DtΪ��Ӧ�ķ�ѿ������
���ӷ�ѿ����κ���(%) = (�������ӷ�ѿ��-�������ӷ�ѿ��) /�������ӷ�ѿ����
1.2.2 ���������ʵIJⶨ ��������ͱ�����(TBA)��Һ���ⶨ����ȩ(MDA)����[13,16]��MDA = [6.45 �� (D532-D600) -0.56D450] / 100 �� ��Ʒ���ء�������ͪ��ɫ���ⶨ�������Ǻ���[16]���������Ǻ���(mg g-1) = (�ӱ���������������ǵ�mg����ϡ�ͱ���)/��Ʒ��(g)�����ÿ���˹����G-250Ⱦɫ���ⶨ�����Ե���[17]�������ʺ���(mg) = (�ӵ����ʱ���������õ����ʦ�g����ϡ�ͱ���)/1000��
1.2.3 ������ø���Բⶨ �����������շ����вⶨ��������ø(CAT)����[8]����������ľ��Ⱦɫ���ⶨ��������ø(POD)����[18]�����õ�������(NBT)��ɫ���ⶨ���������绯ø(SOD)����[7]��
1.2.4 ���Ӻ����IJⶨ ������ɫ��Ӧ���вⶨK+��Na+; ����ԭ�����չ����ⶨCa2+��Mg2+��
1.3 ���ݷ���
����Microsoft Excel 2007¼�����ݡ��������ͼ, ����SPSS17.0ͳ�����������ݽ��е����ط������(One-way ANOVA)�Ͷ����ط������(Two-way ANOVA), ���رȽϲ�����С�������취(LSD), ������ˮƽ�趨Ϊ0.05��2 ��������
2.1 ��ͬŨ���η�в�ȶ�����������ȷ���Ӱ��
2.1.1 ��������ˮ���ʵ�Ӱ�� ��ͼ1��֪, ���ӵ���ˮ�������η�Ũ�ȵ��������С�����ӵĿ�����ˮʱ����0~8 h���ڿ�ʼ��8 h������12 h, ����2011����ˮ�����������ڹ���106, ���м�ʱ����Ʒ�ֵ���ˮ��������С����0��50��100 mmol L-1�η�������, ����4 hʱ������ˮ�������, 8 h��������ˮ���ʿ����½�; ����150 mmol L-1��200 mmol L-1 �η�������, ����8 h���ӵ���ˮ�������ͼ1
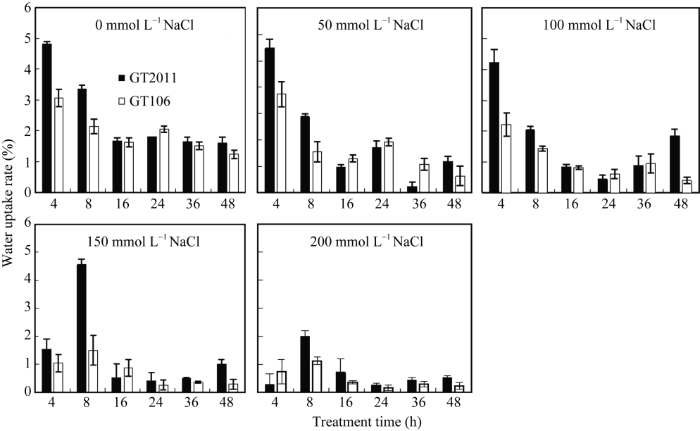 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��ͬ�η�Ũ�ȴ����������������ˮ���ʵ�Ӱ��
GT2011: ����2011; GT106: ����106��
Fig. 1Effect of different salt concentrations on water uptake rate of sweet sorghum seeds
GT2011: Guotian 2011; GT106: Guotian 106.
2.1.2 �����ӷ�ѿ�ʡ���ѿ�Ƽ���ѿָ����Ӱ��
���η�Ũ������, 2��Ʒ�ֵķ�ѿ�ơ���ѿ�ʺͷ�ѿָ��������(��1)����������, 50 mmol L-1 �η�Ũ�ȴ�����, ����2011�ķ�ѿ�ơ���ѿ�ʺͷ�ѿָ���½�������, ������106��������, ���������106���ηֽ�Ϊ���С����η�Ũ�ȴ���100 mmol L-1 ʱ, 2��Ʒ�ֵķ�ѿ�ơ���ѿ�ʺͷ�ѿָ�����������͡����η�Ũ�ȴ�����, ����2011�ķ�ѿ�ʡ���ѿ�ƺͷ�ѿָ�����������ڹ���106��
Table 1
��1
��1�����Ʒ���ڲ�ͬ�ηִ����µ����ӷ�ѿ�ơ���ѿ�ʺͷ�ѿָ��
Table 1
| �η�Ũ�� Salt concentration (mmol L-1) | ����2011 Guotian 2011 | ����106 Guotian 106 | ||||||
|---|---|---|---|---|---|---|---|---|
| ��ѿ��Germination energy (%) | ��ѿ�� Germination rate (%) | ��ѿָ�� Germination index | ��ѿ�� Germination energy (%) | ��ѿ�� Germination rate (%) | ��ѿָ�� Germination index | |||
| 0 | 80 a | 88 a | 53 a | 78 a | 85 a | 51 bc | ||
| 50 | 79 a | 84 a | 53 ab | 74 b | 80 b | 49 c | ||
| 100 | 48 b | 60 b | 35 d | 39 c | 45 c | 29 e | ||
| 150 | 25 c | 32 c | 19 f | 17 d | 21 d | 16 g | ||
| 200 | 15 d | 21 d | 11 g | 9 e | 13 e | 6 h | ||
| ƽ��ֵMean | 49 | 57 | 34 | 43 | 49 | 30 | ||
�´��ڴ�|����CSV
2.1.3 ������κ��ʵ�Ӱ�� ��������ӵ�����κ������η�Ũ�ȵ����Ӷ�����(��2)��200 mmol L-1 �η�Ũ�ȴ�����, 2��Ʒ�ֵ�����κ��ʾ��ﵽ����ȷ�ʱ��, ��ͬ�η�Ũ���¹���106������κ��ʵ��������ڹ���2011, �ҹ���106������κ����������ڹ���2011����Ҳ��������106���η�в�Ƚ�Ϊ���С�
Table 2
��2
��2��ͬ�η�Ũ��в������������ӵ�����κ���
Table 2
| Ʒ�� Genotype | �η�Ũ�� Salt concentration (mmol L-1) | ����κ��� Relative salt stress rate (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 12 h | 24 h | 36 h | 48 h | 60 h | 72 h | |||
| ����2011 | 50 | 26 d | 5 d | 2 d | 0 d | 4 d | 2 d | |
| Guotian 2011 | 100 | 59 c | 35 c | 34 c | 28 c | 36 c | 40 c | |
| 150 | 78 b | 60 b | 65 b | 69 b | 68 b | 69 b | ||
| 200 | 81 a | 78 a | 84 a | 83 a | 83 a | 82 a | ||
| ����106 | 50 | 28 d | 10 d | 8 d | 3 d | 5 d | 6 d | |
| Guotian 106 | 100 | 61 c | 67 c | 48 c | 46 c | 48 c | 51 c | |
| 150 | 89 b | 82 b | 80 b | 78 b | 77 b | 79 b | ||
| 200 | 94 a | 89 a | 87 a | 89 a | 89 a | 89 a | ||
�´��ڴ�|����CSV
2.1.4 ��ͬ�η�Ũ��в�ȶ�����MDA������Ӱ��
���η�Ũ������, ������������ӵ�MDA������������(��3)����������, ��50��100��150��200 mmol L-1 NaCl������, ����2011��MDA�����ֱ�������1.6��1.7��2.3��2.5��; ������106��ֱ������� 1.4��1.6��2.1��2.2�����η�в����, ����106��MDA�����������ڹ���2011�����ηִ���Ũ����, ����106��MDA�����ȹ���2011�ֱ��69.6%��50.0%��55.0%��56.6%��50.9%��
Table 3
��3
��3��ͬ�η�в���������������MDA����
Table 3
| �ηִ��� Salt treatment (mmol L-1) | MDA���� MDA content (��mol g-1 FW) | |
|---|---|---|
| ����2011 Guotian 2011 | ����106 Guotian 106 | |
| 0 | 2.3 e | 3.9 e |
| 50 | 3.6 d | 5.4 d |
| 100 | 4.0 c | 6.2 c |
| 150 | 5.3 b | 8.3 b |
| 200 | 5.7 a | 8.6 a |
| ƽ��ֵMean | 4.2 | 6.5 |
�´��ڴ�|����CSV
2.2 �η�в������Դ�����������ʶ�����������ȷ���Ӱ��
2.2.1 ������ˮ�� ��4���������֪, �η�в�ȶ�������ˮ�ȷ���Ӱ����Ҫ�ڴ���8 h��ﵽ����ˮƽ, 0~8 h������Ӱ�졣��Դ�����������ʶ�������ˮ�ȷ���Ӱ���ڸ�ʱ�����ﵽ������ˮƽ���η�Ũ�Ⱥ���Դ�����������ʵĽ������ö�������ˮ�ȷ���Ӱ����ڴ���24~36 h�ﵽ������ˮƽ, ���ദ��ʱ��ξ���������Table 4
��4
��4��ͬ��Դ�����������ʶԲ�ͬ�η�Ũ���������������ˮ��Ӱ��ķ������
Table 4
| ������Դ Source of variation | Fֵ F-value | |||||
|---|---|---|---|---|---|---|
| 0-4 h | 4-8 h | 8-16 h | 16-24 h | 24-36 h | 36-48 h | |
| �η�Ũ�� Salt concentration (SC) | ns | ns | 3.68* | 6.07** | 23.30** | 19.315** |
| ������������ Exogenous growth regulators (EGR) | 107.71** | 121.47** | 105.49** | 180.52** | 2544.96** | 1361.21** |
| �����������ʡ��η�Ũ�� SC��EGR | ns | ns | ns | ns | 3.52** | ns |
�´��ڴ�|����CSV
����5��֪, ���ȷ�ʱ����ӳ�, ��������ӵ���ˮ��������, �������ˮ�ھ������ڴ������16~24 h��������ˮ����50 mmol L-1�ηִ�������������, ���ηִ���Ũ�ȴ���100 mmol L-1ʱ, ������ˮ�ʼ��罵�͡��ڸ��η�Ũ�ȴ�����, ��Դ�����������ʿ���Ч�����η�в�ȶ�������ˮ�ʵ�Ӱ�졣��50��100��150 mmol L-1�η�������, GA3���������ӵ���ˮ�����, �����GABA������Ũ�ȷִ�����, ��ͬ������Դ�����������ʶ�������ˮ�ʵĵ���ЧӦ���ȷ�ǰ�ڲ�����(0~16 h), �ȷ�����Ч������(16~48 h)��
Table 5
��5
��5��ͬ��Դ�����������ʶԲ�ͬ�η�Ũ���������������ˮ�ʵ�Ӱ��
Table 5
| �η�Ũ�� Salt stress (mmol L-1 ) | ������������ Exogenous growth regulator | ��ˮ��Water uptake rate (%) | |||||
|---|---|---|---|---|---|---|---|
| 0-4 h | 4-8 h | 8-16 h | 16-24 h | 24-36 h | 36-48 h | ||
| 0 | 0 | 12.64 b | 21.91 b | 28.70 c | 52.08 b | 67.7 c | 78.20 b |
| KT | 12.80 b | 25.60 ab | 29.44 bc | 54.87 b | 71.2 ab | 83.75 a | |
| GA3 | 13.84 a | 28.79 ab | 41.05 a | 66.26 a | 74.1 a | 84.89 a | |
| SA | 12.85 b | 27.43 ab | 37.50 ab | 56.09 b | 72.0 b | 84.21 a | |
| GABA | 13.44 ab | 27.98 a | 39.11 a | 58.14 ab | 73.4 ab | 84.73 a | |
| ƽ��ֵMean | 13.03 | 26.34 | 35.16 | 57.49 | 71.7 | 83.16 | |
| 50 | 0 | 13.33 a | 23.50 a | 30.83 b | 52.95 b | 71.5 b | 81.83 b |
| KT | 13.67 a | 26.78 a | 30.79 b | 55.04 b | 72.5 ab | 84.58 a | |
| GA3 | 13.85 a | 29.25 a | 42.06 a | 67.60 a | 75.3 a | 86.03 a | |
| SA | 13.73 a | 28.90 a | 37.57 a | 56.37 b | 72.8 ab | 85.51 a | |
| GABA | 13.79 a | 28.97 a | 40.77 a | 58.97 b | 73.8 ab | 85.74 a | |
| ƽ��ֵMean | 13.68 | 27.48 | 36.41 | 58.19 | 73.2 | 84.74 | |
| 100 | 0 | 8.41 a | 14.35 b | 17.11 a | 28.64 a | 41.9 a | 43.73 c |
| KT | 10.34 a | 16.54 a | 17.60 a | 30.21 a | 42.6 a | 44.70 bc | |
| GA3 | 11.46 a | 17.16 a | 18.30 a | 32.98 a | 44.4 a | 48.60 a | |
| SA | 10.61 a | 16.62 a | 17.67 a | 30.24 a | 43.0 a | 46.43 abc | |
| GABA | 11.41 a | 17.06 a | 17.97 a | 32.68 a | 43.6 a | 47.03 ab | |
| ƽ��ֵMean | 10.45 | 16.34 | 17.73 | 30.95 | 43.1 | 46.10 | |
| 150 | 0 | 4.08 a | 6.87 b | 9.48 a | 22.61 c | 25.2 d | 27.15 a |
| KT | 5.13 a | 6.99 b | 9.77 a | 23.59 c | 32.3 c | 35.10 a | |
| GA3 | 5.85 a | 8.83 a | 10.21 a | 28.89 a | 35.6 a | 38.19 a | |
| SA | 5.49 a | 7.15 b | 9.78 a | 27.20 b | 33.8 b | 36.52 a | |
| GABA | 5.62 a | 8.24 ab | 9.92 a | 27.87 ab | 34.8 ab | 37.45 a | |
| ƽ��ֵMean | 5.23 | 7.62 | 9.83 | 26.03 | 32.3 | 34.88 | |
�´��ڴ�|����CSV
2.2.2 ���ӷ�ѿ�� �������ȷ�������, �η�Ũ�Ⱥ���Դ�����������ʶ���������ӷ�ѿ�ʵ�Ӱ����ﵽ������ˮƽ, �����ߵĽ������ö������ȷ���Ӱ������ȷ�����(12 h)���ȷ�ĩ��(72 h)�ﵽ����ˮƽ(��6)�������η�Ũ������, ���ӷ�ѿ������; ��Դ�����������ʿ������������ӵķ�ѿ��, �ڸ��η�Ũ����, �����������ʴ��������ӵķ�ѿ�ʾ����ڶ���; ����GABA�����µķ�ѿ�����, �����SA��GA3 (ͼ2)��
Table 6
��6
��6��ͬ�����������ʶԲ�ͬ�η�Ũ�����������ѿ��Ӱ��ķ������
Table 6
| ������Դ Source of variation | Fֵ F-value | |||||
|---|---|---|---|---|---|---|
| 12 h | 24 h | 36 h | 48 h | 60 h | 72 h | |
| �η�Ũ�� Salt concentration (SC) | 72.45** | 20.61** | 35.79** | 55.47** | 78.56** | 30.29** |
| ������������ Exogenous growth regulators (EGR) | 303.91** | 1586.50** | 3397.20** | 6028.48** | 7744.33** | 6326.75** |
| �����������ʡ��η�Ũ�� SC��EGR | 7.37** | ns | ns | ns | ns | 2.57* |
�´��ڴ�|����CSV
ͼ2
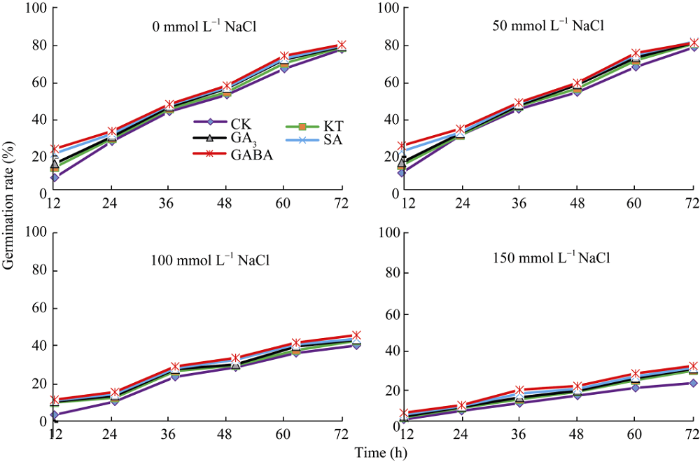 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2��Դ�����������ʶԲ�ͬ�η�Ũ������������ӷ�ѿ�ʵ�Ӱ��
KT: ������; GA3: ��ù��; SA: ˮ����; GABA: ��-�������ᡣ
Fig. 2Effect of exogenous growth regulators on germination rate of sweet sorghum under salt stress
KT: kinetin; GA3: gibberellin; SA: salicylic acid; GABA: ��-aminobutyric acid.
2.2.3 �����������������ʵ�Ӱ�� �����������, �η�в�ȡ���Դ�����������ʼ����ߵĽ������ö�����������ڿ����Ե��Ϳ������Ǻ�����Ӱ����ﵽ������ˮƽ(P < 0.01), ���Ա���ȩ������Ӱ������η�в���´ﵽ������ˮƽ(P < 0.01), ��Դ�����������ʼ����ηֵĻ�����MDA����������Ӱ��(��7)�������η�Ũ�ȵ�����, ��������ӵ�MDA����������, ��������, 50��100��150 mmol L-1 NaCl������MDA�����ֱ�ƽ������12.4%��11.4%��21.7%������Դ�����������ʵ�����MDA�ĺ�����������, ��0��50��150 mmol L-1 NaCl������, GABA�����µ�MDA�������; ��100 mmol L-1 NaCl��Һ������GA3�����µ�MDA�������(��8)��
Table 7
��7
��7��ͬ��Դ�����������ʶ��η�в�������������������������Ӱ��ķ������
Table 7
| ������Դ Source of variation | Fֵ F-value | ||
|---|---|---|---|
| MDA ���� MDA content (��mol g-1 FW) | �����Ե����� Content of soluble protein (mg g-1 FW) | �������Ǻ��� Content of soluble sugar (mg g-1 FW) | |
| �η�Ũ�� Salt concentration (SC) | 2.33** | 92.30** | 110.61** |
| ������������ Exogenous growth regulators (EGR) | ns | 310.16** | 128.91** |
| �����������ʡ��η�Ũ�� SC��EGR | ns | 13.29** | 2.56* |
�´��ڴ�|����CSV
Table 8
��8
��8��ͬ��Դ�����������ʶ��η�в����������������������ʺ�����Ӱ��
Table 8
| �η�Ũ�� Salt concentration (mmol L-1 ) | ������������ Exogenous growth regulator | ���������ʺ��� Content of osmotic adjustment substance | ||
|---|---|---|---|---|
| MDA (��mol g-1 FW) | �����Ե��� Soluble protein (mg g-1 FW) | �������� Soluble sugar (mg g-1 FW) | ||
| 0 | 0 | 29.0 a | 3.53 d | 12.72 c |
| KT | 26.2 b | 6.60 c | 12.74 c | |
| GA3 | 21.1 c | 7.23 c | 17.09 b | |
| SA | 12.4 d | 8.75 b | 25.26 a | |
| GABA | 7.6 e | 14.09 a | 25.42 a | |
| ƽ��ֵMean | 19.3 | 8.04 | 18.65 | |
| 50 | 0 | 32.6 a | 4.21 c | 14.66 e |
| KT | 30.7 b | 6.97 b | 16.79 d | |
| GA3 | 29.2 b | 7.78 b | 20.80 c | |
| SA | 14.8 c | 14.57 a | 28.60 b | |
| GABA | 11.4 d | 14.43 a | 30.21 a | |
| ƽ��ֵMean | 23.7 | 9.59 | 22.21 | |
| 100 | 0 | 32.3 a | 4.63 d | 19.21 e |
| KT | 15.5 b | 8.55 c | 21.10 d | |
| GA3 | 13.6 b | 8.61 c | 25.61 c | |
| SA | 32.7 a | 21.38 a | 40.51 a | |
| GABA | 31.5 a | 15.23 b | 35.40 b | |
| Mean | 25.1 | 11.68 | 28.37 | |
| 150 | 0 | 35.3 a | 8.44 b | 23.78 d |
| KT | 17.7 b | 23.69 a | 42.95 b | |
| GA3 | 33.4 a | 8.55 b | 48.27 a | |
| SA | 32.6 a | 9.30 b | 27.33 c | |
| GABA | 17.2 b | 21.66 a | 28.52 c | |
| ƽ��ֵMean | 27.2 | 14.33 | 34.17 | |
�´��ڴ�|����CSV
���Ƶ�, �����Ե��Ϳ������Ǻ������ηִ���Ũ�ȵ����Ӷ����ӡ���������, ��������, �ڸ��ηִ�����, �����Ե��ֱ�������19.3%��45.3%��74.9%, �������Ƿֱ�������19.1%��52.1%��83.2%���ڸ��η�Ũ�ȴ�����, ��Դ��������������������������ڿ����Ե��Ϳ������ǵĺ���, �����������ӵķ��ȴﵽ1�����ϡ���0��50��100 mmol L-1 NaCl������, SA��GABA�����¿����Ե��Ϳ������ǵĺ������, ����150 mmol L-1 NaCl��Һ������, KT��GABA�����µ����ӵĿ����Ե��������, ���������ǵĺ�����GA3�����µĺ�����ߡ�
2.2.4 �Կ�����ø���Ե�Ӱ�� ����9��֪, ���η�Ũ�ȵ�����, 3�ֿ�����ø�Ļ��������ߺ�, ��50 mmol L-1 �ηִ�����SOD��POD��CAT�Ļ��Ծ��ﵽ���ֵ���ڸ��ηִ�����, ��Դ�����������ʿ���������3�ֱ���ø�Ļ��ԡ���0��50 mmol L-1�ηִ�����, SA��GABA��3��ø���Ե���Ч���, SA������ø���Էֱ�ȶ���������97.8%��131.6% (SOD)��107.3%��109.4% (POD)��74.2%��75.2% (CAT)(0 mmol L-1 NaCl), GABA������3��ø���Էֱ�ȶ���������86.6%��96.3% (SOD)��70.7%��60.0% (POD)��43.7%��45.9% (CAT)(50 mmol L-1 NaCl)��100 mmol L-1 �ηִ�����SA�����µ�SOD��POD�������, �ȶ��շֱ�������304.4%��142.9%, ��GABA������CAT�Ļ������, �ȶ��������80.9%������150 mmol L-1�ηִ�����, SOD��KT�����µĻ������, �ȶ���������365.0%, ��POD��CAT��SA�����µĻ������, �ֱ�������147.1%��50.5%��
Table 9
��9
��9�η�в���²�ͬ�����������ʶ���������ӿ�����ø���Ե�Ӱ��
Table 9
| �η�Ũ�� Salt concentration (mmol L-1) | ������������ Exogenous growth regulator | ø���� Enzyme activity (U g-1 FW) | ||
|---|---|---|---|---|
| SOD | POD | CAT | ||
| 0 | 0 | 31.35 e | 257.98 d | 251.04 c |
| KT | 40.94 d | 287.47 c | 267.88 b | |
| GA3 | 51.35 c | 361.24 b | 271.99 b | |
| SA | 62.02 b | 534.74 a | 437.42 a | |
| GABA | 72.60 a | 540.22 a | 439.85 a | |
| ƽ��ֵMean | 51.65 | 396.33 | 333.64 | |
| 50 | 0 | 41.56 c | 343.57 e | 323.38 c |
| KT | 49.98 c | 372.75 d | 323.83 c | |
| GA3 | 63.51 b | 455.42 c | 335.06 b | |
| SA | 77.53 a | 586.33 a | 464.59 a | |
| GABA | 81.56 a | 549.64 b | 471.70 a | |
| ƽ��ֵMean | 62.83 | 461.54 | 383.71 | |
| 100 | 0 | 15.38 e | 185.61 e | 199.89 e |
| KT | 27.55 d | 211.66 b | 213.30 d | |
| GA3 | 43.78 b | 318.92 c | 245.10 c | |
| SA | 62.20 a | 450.79 a | 298.31 b | |
| GABA | 31.13 c | 395.49 b | 361.51 a | |
| ƽ��ֵMean | 36.01 | 312.49 | 263.62 | |
| 150 | 0 | 10.80 c | 120.92 e | 153.75 e |
| KT | 50.22 a | 223.88 c | 193.73 c | |
| GA3 | 20.64 b | 271.57 b | 178.00 d | |
| SA | 24.15 b | 298.78 a | 231.31 a | |
| GABA | 28.99 b | 185.78 d | 212.83 b | |
| ƽ��ֵMean | 26.96 | 220.19 | 193.92 | |
�´��ڴ�|����CSV
2.2.5 �����Ӻ�����Ӱ�� ����10��֪, �����η�Ũ������, �����������Na+����������, ��K+��Ca2+��Mg2+�������͡�������������, 50��100��150 mmol L-1 NaCl������Na+�����ֱ�������29.8%��66.7%��104.6%, ��K+�����ֱ���4.1%��4.6%��8.2%, Ca2+�����ֱ���20.0%��26.7%��40.0%, Mg2+�����ֱ���6.0%��8.0%��14.0%����Դ�����������ʿ���Ч������в�ȵ�Σ��, ����������, ��Դ�����������ʿ���������Na+�������еĻ��ۡ�������Ӷ�K+��Ca2+��Mg2+�����ա���Na+����, ���ηִ�����GABA���������Ӷ�Na+������������, ���κ��Ļ���ЧӦ��ѡ�0��50 mmol L-1�ηִ�����K+��Ca2+��GABA�����º������, ��100��150 mmol L-1 �ηִ�����GA3���������Ӷ�K+��Ca2+����������ࡣ����, KT�ɴٽ�Mg2+������, �ڸ��η�в����KT�����µ�Mg2+��������ߡ�
Table 10
��10
��10 �η�в���²�ͬ�����������ʶ�������������Ӻ�����Ӱ��
Table 10
| �η�Ũ�� Salt concentration (mmol L-1) | ������������ Exogenous growth regulator | ���Ӻ��� Ions content (mg g-1) | |||
|---|---|---|---|---|---|
| Na+ | K+ | Ca2+ | Mg2+ | ||
| 0 | 0 | 2.85 a | 1.96 c | 0.15 d | 0.50 c |
| KT | 2.57 b | 1.99 c | 0.18 b | 0.62 a | |
| GA3 | 2.56 b | 2.03 b | 0.18 b | 0.53 b | |
| SA | 2.22 c | 2.08 b | 0.16 c | 0.53 b | |
| GABA | 2.09 d | 2.13 a | 0.20 a | 0.54 b | |
| ƽ��ֵMean | 2.55 | 2.04 | 0.17 | 0.54 | |
| 50 | 0 | 3.70 a | 1.88 c | 0.12 c | 0.47 d |
| KT | 3.47 b | 1.96 b | 0.14 b | 0.55 a | |
| GA3 | 3.36 c | 1.99 ab | 0.14 b | 0.49 c | |
| SA | 3.24 d | 2.04 a | 0.14 b | 0.51 bc | |
| GABA | 3.12 e | 2.03 a | 0.16 a | 0.52 b | |
| ƽ��ֵMean | 3.38 | 1.98 | 0.14 | 0.51 | |
| 100 | 0 | 4.75 a | 1.87 c | 0.11 c | 0.46 d |
| KT | 4.61 b | 1.97 ab | 0.12 b | 0.52 a | |
| GA3 | 4.50 c | 2.00 a | 0.13 a | 0.48 b | |
| SA | 4.34 d | 1.93 b | 0.12 b | 0.47 c | |
| GABA | 4.23 e | 1.96 ab | 0.12 b | 0.47 c | |
| ƽ��ֵMean | 4.49 | 1.95 | 0.12 | 0.48 | |
| 150 | 0 | 5.83 a | 1.80 d | 0.09 d | 0.43 c |
| KT | 5.47 c | 1.85 c | 0.10 c | 0.47 a | |
| GA3 | 5.21 d | 1.95 a | 0.12 a | 0.45 b | |
| SA | 5.65 b | 1.89 b | 0.10 c | 0.45 b | |
| GABA | 5.17 d | 1.93 ab | 0.11 b | 0.46 ab | |
| ƽ��ֵMean | 5.47 | 1.88 | 0.10 | 0.45 | |
�´��ڴ�|����CSV
3 ����
�ηֶ������Ӱ�����˫��ЧӦ, һ�����������ЧӦ, ��ֲ����������Σ��[24,25]; ��һ����ʹ���ӷ������߶�����κ�[26]���ڱ��о���, ���κ�������ЧӦΪ��, ���η�Ũ��в������������ӵ��ȷ�ָ�����������, ����ɳ������[24]���ܹ�����[25]���о����һ�¡�����Ҫԭ�����η�в��������в����ɵ������ɺ�, �η�в���γɵĸ�����������������ˮ����, �����ȷ������������ڵ��۵�ˮ�����ø�Ļȱ�ٱ�Ҫ��ˮ����, ����ȷ����ѡ���ѿ�ʽϵ�[14]�����η�Ũ����ɵ���в��, ��ʹϸ��Ĥ����, �ƻ�Ĥ��ͨ�Ժ����ӵ��ڹ���, ��������������κ���[27]������κ��ʵ����ӵ�������������ƽ���ƻ�, �������Na+�����պͻ���, �����ƶ��������ӵ�����[12]���ڱ��о���, ��������ӵ�����κ������η�Ũ�ȵ����Ӷ�����, Na+�������ڵĸ���, ���������Ӷ�K+��Ca2+��Mg2+������, ��������ƽ����ƻ����������ζ���[28], ���ƵĽ��������Farooq��[12]��Nimir��[27]���о��С�
����, ���о�����, ���η�в����, ֲ�����ڲ��������Ļ�����(ROS), ����Ĥ֬��������Ӧ(��������ȩ)��������, ֱ���ƻ�����͵����ʵȴ�л�[29]�����о��и��η�Ũ�ȴ�����, ����106��MDA���������ڹ���2011, ��������2011�������Ը��ڹ���106������ȩ�Ƿ�ӳϸ��Ĥ֬����������Ĥ���˳̶ȵ���Ҫָ��, �京������Ĥ���˳̶ȳ����ȡ���Ʒ�ֿ��Գɷ���[30]�����η�Ũ�ȴ�����, SOD��POD��CAT�Ļ��������ߺ�, ��50 mmol L-1 �ηִ����»������, �������Ũ�ȵ��η�������������ˮ�ȷ�����ߴ�лˮƽ, ����Ũ�ȵ��η�в�������������ȷ���л, �������ӵ���ˮ�ʡ���ˮ���ʡ���ѿ�ʡ���ѿָ���Ȳ����ϱ���һ��, ���ƵĽ��Ҳ�������ܹ�����[25]��Nimir��[27]���о���, ����������ۡ�
��Դ�����������ʿ���Ч�����κ�, �ٽ�����������ȷ�, ����ܵ����û��ƿɹ���Ϊ����4��: (1)��Դ�����������ʿ��յ����ӿ��ٴ�������, ������ӻ����������ȷ�״̬, �ӿ��˶�ˮ�ֵ����ն��ٽ��ȷ�[27]��(2)��Դ������������ͨ����߿�����ø�Ļ���, ������������Ļ����������ɻ�, �Ӷ������κ���SOD�ɽ�(O2-)�绯��H2O2��O2, H2O2��ͨ��CAT��POD�����ñ��ֽ�Ϊ������H2O��O2, �Ӷ�ʹֲ�������˺�[31]��(3)ֲ���ͳ����øϵͳ�������κ�, ��ͨ���������кͻ��к����ʵ�����[32]������Դ������������������, ���������ʿ������ǺͿ����Ե�������������, ����������һ����Ϊ�����ȷ��ṩ���������ʻ���, ��һ�������ϻ��кͲ������к�����, �Ӷ����Ͷ����ӵ����ˡ�����ϸ���ṹ��, ���������ʵ����ÿɱ���Ĥ���ס������˵�DNA�����ӵ��ϳ�����, ��������ȷ�[33]��(4)��Դ��������������Ҫͨ��������������ƽ�������⡢�����κ����η�в����, Na+�Ĺ�����ۿ�����K+���Ӽ��������ӵ�����, �ƻ����������ӵ�ƽ�⡣һ��������Na+����ɵ��ζ�����ȱ��[34,35]; ��һ����Na+��ֱ�ӽ���ϸ��, �ƻ�Ĥ�Ľṹ��Ӱ��ϸ���ʵĴ�л�[36]����Դ�������������ܹ��ٽ�������ˮ�ȷ��������κ���Ҫ����Ϊ�併���˶�Na+������, ά�������ڵ�����ƽ��[37]����ͬ��Դ�����������ʶ��κ��Ļ���ЧӦ��ͬ, ��������, GA3��GABA��������ˮ�ȷ�ʱ�κ��Ļ���ЧӦ�Ϻ�, ��KT�ٽ����Ӷ�Mg2+������Ч���Ϻá�����������Դ�����������ʵĴ�л;����ͬ�й�, ����ػ����д���һ���о���
4 ����
��в��ͨ������ˮ�����ա��Ӿ��������˺��ƻ�����ƽ��Ӱ����������ӵ��ȷ����̡���Դ��������������Ҫͨ���������ߴٽ���ˮ����߿�����ø���ԡ��������������ʺ����͵�������ƽ���;�������κ�������, GA3��GABA������������κ��Ļ���ЧӦ�Ϻ�, KT�ٽ����Ӷ�Mg2+������Ч���Ϻá�������Դ������������(������GA3��GABA)�����ڸ�������������, �ٽ��μ���и������ƹ���ֲ���μ�ؿ������á��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.3969/j.issn.1674-3466.1999.03.002URL [��������: 1]

This paper summerized the studies on the halophytes in China which includes ( 1 ) the halophyte species; (2) the types of halophytes; (3) the vegetation types of halopytes; (4) halopy- tes failed to be recorded in Holoph of the world; (5) economic potential of halophytes. The further directions and key area of research on the Chinese halophytes were suggested at last.
DOI:10.3969/j.issn.1674-3466.1999.03.002URL [��������: 1]
This paper summerized the studies on the halophytes in China which includes ( 1 ) the halophyte species; (2) the types of halophytes; (3) the vegetation types of halopytes; (4) halopy- tes failed to be recorded in Holoph of the world; (5) economic potential of halophytes. The further directions and key area of research on the Chinese halophytes were suggested at last.
[��������: 1]
[��������: 1]
DOI:10.16420/j.issn.0513-353x.2015-0906URL [��������: 1]
Ϊ̽����Դˮ���ᣨSA�����ⷬ��������в���˺��Ĺ����������,�ԡ��طᱣ�ڡ���������Ϊ�Բ�,��ˮ��������,����Ҷ����ӫ���ѧ�����о�Ӫ��Һ�У�1/4 Hoagland��ʩ��SA��200��mol��L~��-1������100 mmol��L~��-1�� Na Clв���·�������PS��⻯ѧЧ�ʡ������ܷ��������ɫ�����չ������õ�Ӱ�졣�����ʾ��Na Clв��15 d��,SA����������ҶƬPS��DZ�ڹ⻯ѧ���ԣ�F_v/F_o�������⻯ѧЧ�ʣ�F_v/F_m��������ת��Ч�ʣ�F_v��/F_m�䣩��ʵ�ʹ⻯ѧЧ�ʣ���_��PS�����⻯ѧӫ�����ϵ����q_P�������չ������ڽ��й⻯ѧ��Ӧ�ķݶP����Ҷ����ӫ��˥���ʣ�R_��fd������ͬ�̶�����;P_��S�ǹ⻯ѧӫ�����ϵ����NPQ����������ѹ����1�Cqp���������Ⱥ�ɢ�ķݶD������Ϲ����������ֵ��L_��PFD������˫��ϵͳ�伤���ܷ��䲻ƽ��ƫ��ϵ������/���C1�����Խ���;PS��Ӧ���ķǹ⻯ѧ��ɢ�ķݶE_x���仯�����Թ��ɡ����Ͻ������,��ԴSA����ͨ�����PS��⻯ѧ������������в�ȵ��µĹ�����,�����ٽ��˷������������������
DOI:10.16420/j.issn.0513-353x.2015-0906URL [��������: 1]
Ϊ̽����Դˮ���ᣨSA�����ⷬ��������в���˺��Ĺ����������,�ԡ��طᱣ�ڡ���������Ϊ�Բ�,��ˮ��������,����Ҷ����ӫ���ѧ�����о�Ӫ��Һ�У�1/4 Hoagland��ʩ��SA��200��mol��L~��-1������100 mmol��L~��-1�� Na Clв���·�������PS��⻯ѧЧ�ʡ������ܷ��������ɫ�����չ������õ�Ӱ�졣�����ʾ��Na Clв��15 d��,SA����������ҶƬPS��DZ�ڹ⻯ѧ���ԣ�F_v/F_o�������⻯ѧЧ�ʣ�F_v/F_m��������ת��Ч�ʣ�F_v��/F_m�䣩��ʵ�ʹ⻯ѧЧ�ʣ���_��PS�����⻯ѧӫ�����ϵ����q_P�������չ������ڽ��й⻯ѧ��Ӧ�ķݶP����Ҷ����ӫ��˥���ʣ�R_��fd������ͬ�̶�����;P_��S�ǹ⻯ѧӫ�����ϵ����NPQ����������ѹ����1�Cqp���������Ⱥ�ɢ�ķݶD������Ϲ����������ֵ��L_��PFD������˫��ϵͳ�伤���ܷ��䲻ƽ��ƫ��ϵ������/���C1�����Խ���;PS��Ӧ���ķǹ⻯ѧ��ɢ�ķݶE_x���仯�����Թ��ɡ����Ͻ������,��ԴSA����ͨ�����PS��⻯ѧ������������в�ȵ��µĹ�����,�����ٽ��˷������������������
DOI:10.3969/j.issn.1000-7091.2012.06.018Magsci [��������: 1]
��6��ˮ��Ʒ��Ϊ�������,���������״̬�¹��NaCl���Ƶ�ˮ��Һ������в��,��������ȡ������Na+��K+��Ca<sup>2</sup>+�����չ��ɺͽ�Ҷ����̬����������,��������Ʒ�������ԵĹ�ϵ���������,6��Ʒ�ֵ�����������ϵ���ߵ�����Ϊ:����5>����10>�ɾ�12>��ũ425>ĵ����26>ĵ����30����в�Ⱥ�ˮ��ֲ��Na+��������,K+��������,����Ʒ�ֵ��ϲ�Na+����������,���н϶�,����Ʒ�ֵ��ϲ�K+���۶�,���в��첻����;Ca<sup>2</sup>+��༯���ڸ���,в�Ⱥ�����Ʒ���ڸ����仯������,����Ʒ���½��϶ࡣ��в����,����Ʒ�ֵĽ�Ҷ������Ҷ���ͽ�Ҷ�����������յIJ����������������ˮƽ,����Ʒ�ֽ�Ҷ�������;����Ʒ�ֵĽ�Ҷ��в�Ⱥ�������,����Ʒ���ɾ�����Ϊƽչ,����Ʒ�ֵľ�����ҪС��������Ʒ�֡�Ҷ���غ������ֵ��Ʒ�������ԵĽ�������,���������������ˮƽ����в�Ⱥ�,����Ʒ�ֽ�������Ʒ�ֵ��������ʺ������ȸ�,���ڲ��¶�����Ʒ�������ԵĽ��Ͷ����ߡ������ڵ��ϲ�K+�������ֵ����Ҷ�����ֵ���ڲ��¶����ֵ����Na+�������ֵ����Ca<sup>2</sup>+�������ֵ����Ҷ������ֵ�������������ֵ����Ϊˮ��������ɸѡ������ָ�ꡣ
DOI:10.3969/j.issn.1000-7091.2012.06.018Magsci [��������: 1]
��6��ˮ��Ʒ��Ϊ�������,���������״̬�¹��NaCl���Ƶ�ˮ��Һ������в��,��������ȡ������Na+��K+��Ca<sup>2</sup>+�����չ��ɺͽ�Ҷ����̬����������,��������Ʒ�������ԵĹ�ϵ���������,6��Ʒ�ֵ�����������ϵ���ߵ�����Ϊ:����5>����10>�ɾ�12>��ũ425>ĵ����26>ĵ����30����в�Ⱥ�ˮ��ֲ��Na+��������,K+��������,����Ʒ�ֵ��ϲ�Na+����������,���н϶�,����Ʒ�ֵ��ϲ�K+���۶�,���в��첻����;Ca<sup>2</sup>+��༯���ڸ���,в�Ⱥ�����Ʒ���ڸ����仯������,����Ʒ���½��϶ࡣ��в����,����Ʒ�ֵĽ�Ҷ������Ҷ���ͽ�Ҷ�����������յIJ����������������ˮƽ,����Ʒ�ֽ�Ҷ�������;����Ʒ�ֵĽ�Ҷ��в�Ⱥ�������,����Ʒ���ɾ�����Ϊƽչ,����Ʒ�ֵľ�����ҪС��������Ʒ�֡�Ҷ���غ������ֵ��Ʒ�������ԵĽ�������,���������������ˮƽ����в�Ⱥ�,����Ʒ�ֽ�������Ʒ�ֵ��������ʺ������ȸ�,���ڲ��¶�����Ʒ�������ԵĽ��Ͷ����ߡ������ڵ��ϲ�K+�������ֵ����Ҷ�����ֵ���ڲ��¶����ֵ����Na+�������ֵ����Ca<sup>2</sup>+�������ֵ����Ҷ������ֵ�������������ֵ����Ϊˮ��������ɸѡ������ָ�ꡣ
DOI:10.11838/sfsc.20060302URL [��������: 1]
����ֲ������ض��Ŀ����ԣ������Ρ�ϡ�Ρ����Ρ����εȶ��ֶ�����������������Ӧ��ʽ��ȫ������ֲ��Լ��1560���֣��й���502�֡���������ֲ������Ҫ�ľ��ü�ֵ�����Թ㷺����ʳƷ����֯�����졢������ҽҩ�������ȷ��档����Ҫ������Ը�����������������ά����̬ƽ��Ļ���Ч�ѡ���ֲ����ֲ�����ӵ��渲�ǣ������������������մ��������е��η֡��Ӷ����ٸ������η��ۻ���ͬʱ���������л��ʣ���������������״���ٽ���������Ч���������ã�����Խ������������������������²�������������ٵر���ʴ��ʮ�ȷ�Σ����ͨ��������ѱ����ɸѡ�������ձ䡢������ֵ��ֶΣ��ҹ��Ѿ�ѡ�������ü�ֵ�ߡ���Ӧ�ü�ֵ������ֲ��150�֡����ļ�Ҫ��������Щ����ֲ��������ĸ���Ч���;������ü�ֵ��
DOI:10.11838/sfsc.20060302URL [��������: 1]
����ֲ������ض��Ŀ����ԣ������Ρ�ϡ�Ρ����Ρ����εȶ��ֶ�����������������Ӧ��ʽ��ȫ������ֲ��Լ��1560���֣��й���502�֡���������ֲ������Ҫ�ľ��ü�ֵ�����Թ㷺����ʳƷ����֯�����졢������ҽҩ�������ȷ��档����Ҫ������Ը�����������������ά����̬ƽ��Ļ���Ч�ѡ���ֲ����ֲ�����ӵ��渲�ǣ������������������մ��������е��η֡��Ӷ����ٸ������η��ۻ���ͬʱ���������л��ʣ���������������״���ٽ���������Ч���������ã�����Խ������������������������²�������������ٵر���ʴ��ʮ�ȷ�Σ����ͨ��������ѱ����ɸѡ�������ձ䡢������ֵ��ֶΣ��ҹ��Ѿ�ѡ�������ü�ֵ�ߡ���Ӧ�ü�ֵ������ֲ��150�֡����ļ�Ҫ��������Щ����ֲ��������ĸ���Ч���;������ü�ֵ��
DOI:10.3969/j.issn.1088-1631.2011.03.038URL [��������: 1]
������������Դ�о��ص㡢ũҵ��Դ������״��������������Դ�о���չ���ص�����˺ӱ�ʡֲ��ת�������Ľ��е�����ũ����ո�����������ȡ�����������ʺϳ��͵ļ����о�����������Դ���Ի�����Դѹ�����⡣Ϊ�ˣ���������Դ�о�����������˽�һ������������δ����������Դ���о�����̽�ֺ�չ����
DOI:10.3969/j.issn.1088-1631.2011.03.038URL [��������: 1]
������������Դ�о��ص㡢ũҵ��Դ������״��������������Դ�о���չ���ص�����˺ӱ�ʡֲ��ת�������Ľ��е�����ũ����ո�����������ȡ�����������ʺϳ��͵ļ����о�����������Դ���Ի�����Դѹ�����⡣Ϊ�ˣ���������Դ�о�����������˽�һ������������δ����������Դ���о�����̽�ֺ�չ����
DOI:10.1016/j.fcr.2010.08.011URL [��������: 2]
Sweet sorghum ( Sorghum bicolor (L.) Moench.) is a drought-tolerant crop with high resistance to saline-alkaline soils, and sweet sorghum may serve as an alternative summer crop for biofuel production in areas where irrigation water is limited. A two-year study was conducted in Northern Greece to assess the productivity (biomass, juice, total sugar and theoretical ethanol yields) of four sweet sorghum cultivars (Sugar graze, M-81E, Urja and Topper-76-6), one grain sorghum cultivar (KN-300) and one grass sorghum cultivar (Susu) grown in intermediate (3.2 dS m 611) or in high (6.9 dS m 611) soil salinity with either low (120 mm) or intermediate (210 mm) irrigation water supply (supplemented with 142�C261 mm of rainfall during growth). The soil salinity and irrigation water supply effects on the sorghum chlorophyll content index, photosystem II quantum yield, stomatal conductance and leaf K/Na ratio were also determined. The sorghum emergence averaged 75,083 plants ha 611 and 59,917 plants ha 611 in a soil salinity of 3.2 dS m 611 and 6.9 dS m 611, respectively. The most affected cultivar, as averaged across the two soil salinity levels, was the Susu grass sorghum emerging at 53,250 plants ha 611, followed by the Topper-76-6 sweet sorghum emerging at 61,250 plants ha 611. The leaf K/Na ratio decreased with decreasing irrigation water supply, in most cases, but it was not significantly affected by soil salinity. The dry biomass, juice and total sugar yields of sorghum that received 210 mm of irrigation water was 49�C88% greater than the yields of sorghum that received the 120 mm of irrigation water. Sorghum plants grown in a soil salinity of 3.2 dS m 611 produced 42�C58% greater dry biomass, juice and total sugar yields than the yields of sorghum plants grown in a soil salinity of 6.9 dS m 611. The greatest theoretical ethanol yield was produced by sweet sorghum plants grown in a soil salinity of 3.2 dS m 611 with 210 mm of irrigation water (6130 L ha 611, as averaged across cultivar), and the Urja and Sugar graze cultivars produced the most ethanol (7620 L ha 611 and 6528 L ha 611, respectively). Conclusively, sweet sorghum provided sufficient juice, total sugar and ethanol yields in fields with a soil salinity of 3.2 dS m 611, even if the plants received 50�C75% of the irrigation water typically applied to sorghum.
DOI:10.3969/j.issn.1671-5292.2005.04.013URL [��������: 2]
�������һ�־����������Ե���Դ����,���ҹ��������������ȼ�Ͼƾ����Ź����ķ�չǰ��.��������ϸ���������������ѧ���ԵĻ�����,���ҹ��������Ʒ�ֵ��о��Ϳ�����״�����˻��ܷ���,�Խ���ҹ���չ��Դ��������������ؿɹ�Ӧ�Ժ�����ȼ�Ͼƾ���DZ�������������о�.
DOI:10.3969/j.issn.1671-5292.2005.04.013URL [��������: 2]
�������һ�־����������Ե���Դ����,���ҹ��������������ȼ�Ͼƾ����Ź����ķ�չǰ��.��������ϸ���������������ѧ���ԵĻ�����,���ҹ��������Ʒ�ֵ��о��Ϳ�����״�����˻��ܷ���,�Խ���ҹ���չ��Դ��������������ؿɹ�Ӧ�Ժ�����ȼ�Ͼƾ���DZ�������������о�.
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
DOI:10.7666/d.d144050URL [��������: 1]
���о�ѡ�ò�ͬ����(�ӽ��ֺͳ�����)�������Ʒ��Ϊ�������,�о����ζȶԲ�ͬ���������Ʒ�ֵ��ȷ�������������������Ӱ�졣̽������в�ȶ�������ķ�ѿ�ơ���ѿ�ʡ���ѿָ��������ָ���������������ԡ����ز���������������Ӱ���Լ�Ӱ���������Ҫ���ӡ���Ҫ�������: 1.�ζȶԲ�ͬ���������Ʒ���ȷ���Ӱ�� ��0.5%��1.0%��1.5%��2.0%��2.5%��NaCl����Һв����,������ӽ��ֺͳ����ֵ��ȷ����ܵ�����,�����ڷ�ѿ�ơ���ѿ�ʡ���ѿָ��������ָ����ѿ����������ѿ�ء����ؾ����½�������ͬ��Ũ����,�ӽ��ֵ��½������������ڳ����֡���ͬ����Ʒ��֮��ķ�ѿ�ơ���ѿ�ʡ���ѿָ�������ز�������,������ָ����������ѿ����ѿ�ز��첻������ 2.�ζȶԲ�ͬ���������Ʒ������������Ӱ�� ������������0.3%��0.6%��0.8%����в��������,��ͬ���������Ʒ�ֵ����������ܵ�����Ӱ�졣������Ũ�ȵ�����,14�������Ʒ�ֵij������½�,�����������������������������ӳ�,ֲ�����ߡ��볤����֡����ֽ���,Ҷ�ء����ء����ء����ء�ǧ���ء������ء����������������ؾ������½�,�������Ǵ��������ߵ����ơ���ͬ���͵�Ʒ�ּ���ֻ���һ�¡��ӽ��ֺͳ�����֮����������������Ա���Ϊ,����������ǧ���ء������Ǵ���֮����첻����,������֮���������,����13����״(�����ʡ�����������������������ߡ��볤����֡����֡�Ҷ�ء����ء����ء����ء�����������������)֮�������������� 3.�ζȶԲ�ͬ���������Ʒ�����ز���������������Ӱ�� ��в����,��ͬ���������Ʒ�ֵ����ز�����������������ܴ�������Ũ�ȵ�����,���ز������������������½������ӽ����½��ķ����������ڳ����֡���ͬ����Ʒ�ֵIJ����������Ϊ,��������֮���������,���ز���֮����켫������Ӱ��������������Ҫ�����������ء����ء�ǧ���ء��볤�������������;Ӱ�����ز�������Ҫ�����е������ء����ء�Ҷ�ء����ء����֡���ߡ�
DOI:10.7666/d.d144050URL [��������: 1]
���о�ѡ�ò�ͬ����(�ӽ��ֺͳ�����)�������Ʒ��Ϊ�������,�о����ζȶԲ�ͬ���������Ʒ�ֵ��ȷ�������������������Ӱ�졣̽������в�ȶ�������ķ�ѿ�ơ���ѿ�ʡ���ѿָ��������ָ���������������ԡ����ز���������������Ӱ���Լ�Ӱ���������Ҫ���ӡ���Ҫ�������: 1.�ζȶԲ�ͬ���������Ʒ���ȷ���Ӱ�� ��0.5%��1.0%��1.5%��2.0%��2.5%��NaCl����Һв����,������ӽ��ֺͳ����ֵ��ȷ����ܵ�����,�����ڷ�ѿ�ơ���ѿ�ʡ���ѿָ��������ָ����ѿ����������ѿ�ء����ؾ����½�������ͬ��Ũ����,�ӽ��ֵ��½������������ڳ����֡���ͬ����Ʒ��֮��ķ�ѿ�ơ���ѿ�ʡ���ѿָ�������ز�������,������ָ����������ѿ����ѿ�ز��첻������ 2.�ζȶԲ�ͬ���������Ʒ������������Ӱ�� ������������0.3%��0.6%��0.8%����в��������,��ͬ���������Ʒ�ֵ����������ܵ�����Ӱ�졣������Ũ�ȵ�����,14�������Ʒ�ֵij������½�,�����������������������������ӳ�,ֲ�����ߡ��볤����֡����ֽ���,Ҷ�ء����ء����ء����ء�ǧ���ء������ء����������������ؾ������½�,�������Ǵ��������ߵ����ơ���ͬ���͵�Ʒ�ּ���ֻ���һ�¡��ӽ��ֺͳ�����֮����������������Ա���Ϊ,����������ǧ���ء������Ǵ���֮����첻����,������֮���������,����13����״(�����ʡ�����������������������ߡ��볤����֡����֡�Ҷ�ء����ء����ء����ء�����������������)֮�������������� 3.�ζȶԲ�ͬ���������Ʒ�����ز���������������Ӱ�� ��в����,��ͬ���������Ʒ�ֵ����ز�����������������ܴ�������Ũ�ȵ�����,���ز������������������½������ӽ����½��ķ����������ڳ����֡���ͬ����Ʒ�ֵIJ����������Ϊ,��������֮���������,���ز���֮����켫������Ӱ��������������Ҫ�����������ء����ء�ǧ���ء��볤�������������;Ӱ�����ز�������Ҫ�����е������ء����ء�Ҷ�ء����ء����֡���ߡ�
DOI:10.1016/j.plaphy.2017.06.020URLPMID:28648997 [��������: 3]
Salt stress is an ever-present threat to crop yields, especially in countries with irrigated agriculture. Efforts to improve salt tolerance in crop plants are vital for sustainable crop production on marginal lands to ensure future food supplies. Grain legumes are a fascinating group of plants due to their high grain protein contents and ability to fix biological nitrogen. However, the accumulation of excessive salts in soil and the use of saline groundwater are threatening legume production worldwide. Salt stress disturbs photosynthesis and hormonal regulation and causes nutritional imbalance, specific ion toxicity and osmotic effects in legumes to reduce grain yield and quality. Understanding the responses of grain legumes to salt stress and the associated tolerance mechanisms, as well as assessing management options, may help in the development of strategies to improve the performance of grain legumes under salt stress. In this manuscript, we discuss the effects, tolerance mechanisms and management of salt stress in grain legumes. The principal inferences of the review are: (i) salt stress reduces seed germination (by up to more than 50%) either by inhibiting water uptake and/or the toxic effect of ions in the embryo, (ii) salt stress reduces growth (by more than 70%), mineral uptake, and yield (by 12 100%) due to ion toxicity and reduced photosynthesis, (iii) apoplastic acidification is a good indicator of salt stress tolerance, (iv) tolerance to salt stress in grain legumes may develop through excretion and/or compartmentalization of toxic ions, increased antioxidant capacity, accumulation of compatible osmolytes, and/or hormonal regulation, (v) seed priming and nutrient management may improve salt tolerance in grain legumes, (vi) plant growth promoting rhizobacteria and arbuscular mycorrhizal fungi may help to improve salt tolerance due to better plant nutrient availability, and (vii) the integration of screening, innovative breeding, and the development of transgenics and crop management strategies may enhance salt tolerance and yield in grain legumes on salt-affected soils.
DOI:10.3321/j.issn:0496-3490.2007.08.009URL [��������: 2]
; while, 200 mmol L NaCl treatment induced an obvious decrease in chlorophyll content, relative water content and an increase in membrane lipid peroxidation. Net photosynthetic rate (P) and stomatal conductance (G) began to decline strongly at NaCl concentration higher than 50 mmol L, whereas, the intercellular COconcentration (C) increased strikingly. During this process, the stomatal limiting value (L) decreased with the increase of salt treatment. More important, the depression of carbon assimilation under salt stress could not be restored by the enhancement of COconcentration. The measurement of chlorophyll fluorescence showed that the initial fluorescence yield (F), the maximum fluorescence yield (F) and the maximal quantum yield of photosystem �� photochemistry (F/F) significantly went down in 200 mmol LNaCl treated seedlings. Moreover, chlorophyll fluorescence quenching kinetics parameters were not affected by 50 mmol L NaCl treatment. When NaCl concentration was above 50 mmol L, the efficiency of open centers of photosystem �� (F��/F��)��photochemical quenching (q) and actual photosystem �� efficiency (��) decreased sharply, while non-photochemical quenching (NPQ) increased gradually. Therefore, we suggest that the decreased net photosynthetic rate (Pn) in salt-stressed sweet sorghum seedlings is mainly due to non-stomatal limitation; the depression of carbon assimilation capacity significantly alters the excited energy distribution. Under moderate salt stress (100 mmol L NaCl), it is the enhancement of thermal dissipation that dissipate excess excited energy protecting sweet sorghum seedlings against photodamage; however, severely salt treatment (200 mmol L NaCl) causes the reduction of light absorption and improvement of thermal dissipation for keeping the balance in light capture and utilization.
DOI:10.3321/j.issn:0496-3490.2007.08.009URL [��������: 2]
; while, 200 mmol L NaCl treatment induced an obvious decrease in chlorophyll content, relative water content and an increase in membrane lipid peroxidation. Net photosynthetic rate (P) and stomatal conductance (G) began to decline strongly at NaCl concentration higher than 50 mmol L, whereas, the intercellular COconcentration (C) increased strikingly. During this process, the stomatal limiting value (L) decreased with the increase of salt treatment. More important, the depression of carbon assimilation under salt stress could not be restored by the enhancement of COconcentration. The measurement of chlorophyll fluorescence showed that the initial fluorescence yield (F), the maximum fluorescence yield (F) and the maximal quantum yield of photosystem �� photochemistry (F/F) significantly went down in 200 mmol LNaCl treated seedlings. Moreover, chlorophyll fluorescence quenching kinetics parameters were not affected by 50 mmol L NaCl treatment. When NaCl concentration was above 50 mmol L, the efficiency of open centers of photosystem �� (F��/F��)��photochemical quenching (q) and actual photosystem �� efficiency (��) decreased sharply, while non-photochemical quenching (NPQ) increased gradually. Therefore, we suggest that the decreased net photosynthetic rate (Pn) in salt-stressed sweet sorghum seedlings is mainly due to non-stomatal limitation; the depression of carbon assimilation capacity significantly alters the excited energy distribution. Under moderate salt stress (100 mmol L NaCl), it is the enhancement of thermal dissipation that dissipate excess excited energy protecting sweet sorghum seedlings against photodamage; however, severely salt treatment (200 mmol L NaCl) causes the reduction of light absorption and improvement of thermal dissipation for keeping the balance in light capture and utilization.
URL [��������: 2]
ѡ�á����ա���10��Ʒ�����������Ϊ����,�ֱ���Na Cl�͵����ĸ�¶�����д���,�Լ�����Ʒ�ֵĿ������뿹���ԡ��������:��ͬƷ�������������100 mmol��L-1 Na Cl�͵����ĸ�¶���������ȷ��������������ܵ��˲�ͬ�̶ȵ�����;�Ա��δ�����ɺ�������,�������Ʒ�ֵ���������ӵ��ȷ���������������ָ�궼û�������ԵIJ���,��˵����в�����������ȷ�����Ҫ����������в����ɵ������ɺ�;������������ֵ������������ȷ��ڵ�7����������ָ������ۺ�����,������ǿ��˳��Ϊ������1�š���������7�š���������10�š���������2�š���������11-6����MN-3867��������1�š������ա���MN-94����������12�š�,������ǿ��˳��Ϊ������1�š���������2�š���������11-6����������10�š���������7�š���MN-3867����MN-94��������1�š���������12�š������ա�,���Կ�����������ȷ��ڵĿ������뿹���Դ��ڽ�ǿ������ع�ϵ��
URL [��������: 2]
ѡ�á����ա���10��Ʒ�����������Ϊ����,�ֱ���Na Cl�͵����ĸ�¶�����д���,�Լ�����Ʒ�ֵĿ������뿹���ԡ��������:��ͬƷ�������������100 mmol��L-1 Na Cl�͵����ĸ�¶���������ȷ��������������ܵ��˲�ͬ�̶ȵ�����;�Ա��δ�����ɺ�������,�������Ʒ�ֵ���������ӵ��ȷ���������������ָ�궼û�������ԵIJ���,��˵����в�����������ȷ�����Ҫ����������в����ɵ������ɺ�;������������ֵ������������ȷ��ڵ�7����������ָ������ۺ�����,������ǿ��˳��Ϊ������1�š���������7�š���������10�š���������2�š���������11-6����MN-3867��������1�š������ա���MN-94����������12�š�,������ǿ��˳��Ϊ������1�š���������2�š���������11-6����������10�š���������7�š���MN-3867����MN-94��������1�š���������12�š������ա�,���Կ�����������ȷ��ڵĿ������뿹���Դ��ڽ�ǿ������ع�ϵ��
DOI:10.3724/SP.J.1011.2012.01303URL [��������: 1]
Ϊ���о�NaClв���µ��ʶ�����������ȷ���ѿ���������������Ե�Ӱ��,̽�������������������Ĵ�ʩ,�������ò�ͬ�η�Ũ�ȡ���ͬ��Դ��Ũ���������������ѿ���顣�������:NaClв�ȺͲ�ͬ��Դ���������ѿ��ѿ��������Ӱ����в�ͬ��NaClŨ�ȶ�����������ȷ�������Ӱ��,�������ѿ��������,ͨ����߱���ø���Ժ����������ʶ���ǿ�����˺����������ġ�100 mmol.L 1NaClв����,��ϵPOD�������,��ҶƬMDA���������������Ǻ�����POD�������,���κ��̶����û����в����������Ӳ�ͬ��Դ���������������Ҷ�������Ե�Ӱ���������,����Ũ����20 mmol.L 1ʱ,ϸ�����˺��̶����,������á���ͬ��̬��Դ���������ѿ������������Ӱ���������,NH4Cl�Ĵٽ�Ч������KNO3����100mmol.L 1��NaClв����,ʩ���̬������̬��Դ��������ǿ�����ѿ���ڵ�POD����,����MDA����,�Ӷ�������в�ȴ������˺����о�������ȡ�ʵ��ĵ��ʵ��ش�ʩ������������������������
DOI:10.3724/SP.J.1011.2012.01303URL [��������: 1]
Ϊ���о�NaClв���µ��ʶ�����������ȷ���ѿ���������������Ե�Ӱ��,̽�������������������Ĵ�ʩ,�������ò�ͬ�η�Ũ�ȡ���ͬ��Դ��Ũ���������������ѿ���顣�������:NaClв�ȺͲ�ͬ��Դ���������ѿ��ѿ��������Ӱ����в�ͬ��NaClŨ�ȶ�����������ȷ�������Ӱ��,�������ѿ��������,ͨ����߱���ø���Ժ����������ʶ���ǿ�����˺����������ġ�100 mmol.L 1NaClв����,��ϵPOD�������,��ҶƬMDA���������������Ǻ�����POD�������,���κ��̶����û����в����������Ӳ�ͬ��Դ���������������Ҷ�������Ե�Ӱ���������,����Ũ����20 mmol.L 1ʱ,ϸ�����˺��̶����,������á���ͬ��̬��Դ���������ѿ������������Ӱ���������,NH4Cl�Ĵٽ�Ч������KNO3����100mmol.L 1��NaClв����,ʩ���̬������̬��Դ��������ǿ�����ѿ���ڵ�POD����,����MDA����,�Ӷ�������в�ȴ������˺����о�������ȡ�ʵ��ĵ��ʵ��ش�ʩ������������������������
DOI:10.3969/j.issn.1001-7283.2011.05.017URL [��������: 3]
Ϊ���о���Ч�������������������������Ե�Ӱ��,���ò�ͬŨ�ȵĶ�Ч�������ںͳ����ڶ����������Ҷ����ʩ����.�������,����Ũ�ȵĶ�Ч����������Ч��������ҶƬ�Ŀ������ǡ�Ҷ���غͿ����Ե����ʵȵĺ���,������ǿ�������ҶƬ�����Ǻϳ�ø���ԡ��������ʺ;��������,��Ч���������������ߺ;���,���Ծ��ѵ������дٽ�����,�Ӷ������������Ŀ���������,����ʹ���Ѳ������.��Ч��Ƚ����˵�ʹ��Ũ��Ϊ0.75g/L.
DOI:10.3969/j.issn.1001-7283.2011.05.017URL [��������: 3]
Ϊ���о���Ч�������������������������Ե�Ӱ��,���ò�ͬŨ�ȵĶ�Ч�������ںͳ����ڶ����������Ҷ����ʩ����.�������,����Ũ�ȵĶ�Ч����������Ч��������ҶƬ�Ŀ������ǡ�Ҷ���غͿ����Ե����ʵȵĺ���,������ǿ�������ҶƬ�����Ǻϳ�ø���ԡ��������ʺ;��������,��Ч���������������ߺ;���,���Ծ��ѵ������дٽ�����,�Ӷ������������Ŀ���������,����ʹ���Ѳ������.��Ч��Ƚ����˵�ʹ��Ũ��Ϊ0.75g/L.
URL [��������: 2]
��ǰ����ʯ��Դ����Ļ��������������أ���չ����������������������Դ�ܵ��㷺��ע�����ҹ��μ����Դʮ�ַḻ�������μ�ط�չ��Դ������һ�������˫Ӯ����������һ���������Դ����֮һ����Ȼ�����һ�������������������ηֽϸߵ����������������ȷ��ѡ������ѡ������ʵ��Լ����������ܵ����Ƶ����⡣�������о����֣�ˮ������Ϊһ��ֲ�D�أ����л����η�в�ȶ�����������Ӱ�졣�������Ž�13�������Ʒ��Ϊ������ϣ��о����ڲ�ͬ�η�Ũ���벻ͬ��Դˮ����Ũ�������µĸ�Ʒ�����������ˮ�ȷ������������������ԡ����ڴ˻����ϣ�ѡ�����Ч����...
URL [��������: 2]
��ǰ����ʯ��Դ����Ļ��������������أ���չ����������������������Դ�ܵ��㷺��ע�����ҹ��μ����Դʮ�ַḻ�������μ�ط�չ��Դ������һ�������˫Ӯ����������һ���������Դ����֮һ����Ȼ�����һ�������������������ηֽϸߵ����������������ȷ��ѡ������ѡ������ʵ��Լ����������ܵ����Ƶ����⡣�������о����֣�ˮ������Ϊһ��ֲ�D�أ����л����η�в�ȶ�����������Ӱ�졣�������Ž�13�������Ʒ��Ϊ������ϣ��о����ڲ�ͬ�η�Ũ���벻ͬ��Դˮ����Ũ�������µĸ�Ʒ�����������ˮ�ȷ������������������ԡ����ڴ˻����ϣ�ѡ�����Ч����...
DOI:10.1017/S0014479700005640URL [��������: 2]
A field experiment was conducted to study the effect of irrigation with brackish water on the growth and composition of millet and corn, fertilized with various combinations of N and P. Millet irrigated with water of EC 3.30 mmho/cm. produced yields 79 to 93 per cent of the control (tap water of 0.61 mmho/cm.), depending on the N and P treatment applied. With water of EC 5.05 mmho/cm. yields equivalent to 52�C72 per cent were obtained. Corn yields were 78�C90 per cent of the control with irrigation water of 4.60 mmho/cm., and 55�C64 per cent when irrigated with water of 7.06 mmho/cm. The N and P contents were mostly higher in plants grown on plots irrigated with brackish water.
DOI:10.1007/BF02197756URL [��������: 1]
Tomato (Lycopersicon esculentum var. VF 145) plants were grown with Typic Xerofluvents soil in a greenhouse irrigated with recycled nutrient solutions having increasing levels of N and salinity. Positive response of plants to increasing levels of N was obtained at the lowest initial salinity level of 1 dS/m (dS/m = mmho/cm, referenced at 25 C). At the higher initial salinity levels of 5 and 9 dS/m, increasing N was ineffective in counteracting adverse effects on growth and yield caused by the presence of enhanced salt concentrations of the nutrient solution. Total N uptake was linearly correlated with the total water uptake and was severely suppressed by impaired growth associated with the two higher initial salinity levels, irrespective of N levels. The effect of salinity on leaf N concentrations changed over time. Leaf Cl and P concentrations indicated a possible suppressing effect of Cl on P uptake into plant tops.
DOI:10.2135/cropsci1990.0011183X003000010028xURL [��������: 1]
(...) Tomato plants (Lycopersicon esculentum Mill.) were grown in a greenhouse in a continuously flowing solution culture system to investigate whether P fertilization modified the detrimental effects of NaCl at low constant P concentrations similar to those in soil solutions. Increasing P fertilization enhanced the tolerance of tomato plants to NaCl. At 0.1, 1.0, and 10 mM P, the NaCl concentrations that reduced yields of fruit by 50% were 58,72, and 130 mM, respectively (...)
[��������: 1]
[��������: 1]
URL [��������: 1]
�η���Ӱ��ֲ��������һ����Ҫ�������ء��������ϵĸɺ��Ͱ�ɺ�����,������С,��������,�����Լ���������ˮ�����ö��ٽ����η�в�ȵļӾ硣��в��ͨ�����Ӷ�����Ӫ��ʧ�����в��,����ֲ��������������лʧ��,����Ӱ��������������������Ʒ�ʡ��ܽ����η�в�ȶ�ֲ����������Ӱ����о���չ,�������ɻ�������Ĥ֬�������������˺������˺����ж����ʻ��۵ȷ���ϵͳ��������в�ȶ�ֲ����˺�����,��������ֲ����η�в�ȵ���Ӧ����,�ܽ�����Ҫ�Ŀ�������ָ�ꡣ
URL [��������: 1]
�η���Ӱ��ֲ��������һ����Ҫ�������ء��������ϵĸɺ��Ͱ�ɺ�����,������С,��������,�����Լ���������ˮ�����ö��ٽ����η�в�ȵļӾ硣��в��ͨ�����Ӷ�����Ӫ��ʧ�����в��,����ֲ��������������лʧ��,����Ӱ��������������������Ʒ�ʡ��ܽ����η�в�ȶ�ֲ����������Ӱ����о���չ,�������ɻ�������Ĥ֬�������������˺������˺����ж����ʻ��۵ȷ���ϵͳ��������в�ȶ�ֲ����˺�����,��������ֲ����η�в�ȵ���Ӧ����,�ܽ�����Ҫ�Ŀ�������ָ�ꡣ
DOI:10.1080/01904167.2013.867980URL [��������: 1]
An experiment with factorial arrangement of treatments on a randomized complete block (RCB) design basis with three replications was conducted in a greenhouse during Spring 2010 to investigate changes in sodium ion (Na+), potassium ion (K+), Na+/K+ and to determine proline, protein content, and superoxide dismutase (SOD) of four wheat and four barley cultivars. Three salt levels {1, control (no salt), 7, and 13 dS m611 [2.5 and 502g salt [sodium chloride (NaCl) and sodium sulfate (Na2SO4) in 1:1 ratio] per kg of soil, respectively]} were used in this investigation. Salt stress treatments were applied 4 weeks after planting (at 2 leaf stage). Leaf samples were taken four weeks after imposition of salt treatment. The results showed that salinity caused an increased in proline and protein content, and SOD in all wheat and barley cultivars. The highest proline and protein content of barley and wheat cultivars at all salinity levels were observed in ��Nimrooz�� and ��Bam�� cultivars, respectively. At all salinity levels, wheat and barley cultivars ��Kavir�� and ��Nimrooz��, respectively, had the lowest Na+ content. Barley cultivar ��Kavir�� and wheat cultivar ��Bam�� had higher K+ and K+:Na+ ratios. This might be related to salt tolerance in these two cultivars. Wheat and barley cultivars showed differences with regard to proline, protein, and SOD content, Na+, K+, and K+:Na+ ratio, indicating existence of genetic diversity among the cultivars. These findings indicated that higher K+, K+:Na+ ratio, proline, protein, and SOD content could be the key factors, which offer advantage to barley over wheat for superior performance under saline conditions.
[��������: 2]
[��������: 2]
DOI:10.3969/j.issn.1002-1302.2012.12.028URL [��������: 3]
����0��50��100 mmol/L 3��Ũ��NaCl��Һ,�о���Բ�ͬ���������������ˮ���ȷ���Ӱ��.�������:A1Ʒϵ����ˮ���ȷ��ڵ�������������A2Ʒϵ,��ˮ�ʡ���ˮ���ʡ���ѿ�ʡ���ѿ�ơ���ѿָ�����߸����Ⱥ���ѿ���Ⱦ�����A2Ʒϵ.50 mmol/L NaCl��A1Ʒϵ����ˮ����һ���ٽ�����.��0 mmol/L NaCl���,50 mmol/L NaCl���������2��Ʒϵ�ķ�ѿ�ʡ���ѿ�ơ���ѿָ�����߸����Ⱥ���ѿ���ȣ���100 mmol/L NaC1������������2��Ʒϵ����ˮ�ʡ���ˮ���ʡ���ѿ�ʡ���ѿ�ơ���ѿָ��,�������߸�����ѿ������.��Ũ�ȵ���в�ȿ��ܶ����������(�ر��Ǿ���һ����������������)����ˮ���ȷ�����һ���Ĵٽ�����,����Ũ�ȵ���в����������������ӵ���ˮ���ȷ�.
DOI:10.3969/j.issn.1002-1302.2012.12.028URL [��������: 3]
����0��50��100 mmol/L 3��Ũ��NaCl��Һ,�о���Բ�ͬ���������������ˮ���ȷ���Ӱ��.�������:A1Ʒϵ����ˮ���ȷ��ڵ�������������A2Ʒϵ,��ˮ�ʡ���ˮ���ʡ���ѿ�ʡ���ѿ�ơ���ѿָ�����߸����Ⱥ���ѿ���Ⱦ�����A2Ʒϵ.50 mmol/L NaCl��A1Ʒϵ����ˮ����һ���ٽ�����.��0 mmol/L NaCl���,50 mmol/L NaCl���������2��Ʒϵ�ķ�ѿ�ʡ���ѿ�ơ���ѿָ�����߸����Ⱥ���ѿ���ȣ���100 mmol/L NaC1������������2��Ʒϵ����ˮ�ʡ���ˮ���ʡ���ѿ�ʡ���ѿ�ơ���ѿָ��,�������߸�����ѿ������.��Ũ�ȵ���в�ȿ��ܶ����������(�ر��Ǿ���һ����������������)����ˮ���ȷ�����һ���Ĵٽ�����,����Ũ�ȵ���в����������������ӵ���ˮ���ȷ�.
[��������: 1]
[��������: 4]
DOI:10.1007/S11099-005-0001-6URL [��������: 1]
Salt stress causes decrease in plant growth and productivity by disrupting physiological processes, especially photosynthesis. The accumulation of intracellular sodium ions at salt stress changes the ratio of K : Na, which seems to affect the bioenergetic processes of photosynthesis. Both multiple inhibitory effects of salt stress on photosynthesis and possible salt stress tolerance mechanisms in cyanobacteria and plants are reviewed.
[��������: 1]
URLMagsci [��������: 1]
<p>�������һ����Դ������п��������ԡ����ε��ص㡣���о���BJ-17��BJ-18��M-0011���������Ʒ�֣�ϵ��Ϊ���ϣ��о�����в�ȶ�����������ȷ�������������Ӱ�죬ͨ������в�����������������ָ���⣬�ȽϷ���������Ʒ�֣�ϵ���������ԣ�Ϊ�������������ѡ�����ṩ���ݡ������������ͬŨ��NaClв�������£���������ӵ��ȷ����ܵ��˲�ͬ�̶ȵ����ƣ���������NaClŨ�ȵ����ӣ��ȷ����Ƶij̶���֮��������ƣ�����в�������£���ͬƷ�֣�ϵ������������������������ָ����ֳ����¹��ɣ��������������غ���߾������½������������ҶƬ�������������ʡ�����ȩ���������Ậ�����������������������ҶƬҶ���غ����½�������Ҷ����b�����½���Ϊ���ԣ����������Ʒ�֣�ϵ��������������ָ����������BJ-18���н�ǿ�������ԣ�BJ-17��֮��M1-00110����������</p>
URLMagsci [��������: 1]
<p>�������һ����Դ������п��������ԡ����ε��ص㡣���о���BJ-17��BJ-18��M-0011���������Ʒ�֣�ϵ��Ϊ���ϣ��о�����в�ȶ�����������ȷ�������������Ӱ�죬ͨ������в�����������������ָ���⣬�ȽϷ���������Ʒ�֣�ϵ���������ԣ�Ϊ�������������ѡ�����ṩ���ݡ������������ͬŨ��NaClв�������£���������ӵ��ȷ����ܵ��˲�ͬ�̶ȵ����ƣ���������NaClŨ�ȵ����ӣ��ȷ����Ƶij̶���֮��������ƣ�����в�������£���ͬƷ�֣�ϵ������������������������ָ����ֳ����¹��ɣ��������������غ���߾������½������������ҶƬ�������������ʡ�����ȩ���������Ậ�����������������������ҶƬҶ���غ����½�������Ҷ����b�����½���Ϊ���ԣ����������Ʒ�֣�ϵ��������������ָ����������BJ-18���н�ǿ�������ԣ�BJ-17��֮��M1-00110����������</p>
DOI:10.1088/1009-0630/6/5/011URL [��������: 1]
Anti-oxidative mechanisms of Haloxylon ammodendron,Calligonum mongolicum and Elaeagnus angustifolia under drought stress were comparatively studied.The results indicate that the membranes of three plants are damaged to different degree in arid habitat,among which damage to the membranes of Elaeagnus angustifolia is stronger than to that of Haloxylon ammodendron and Calligonum mongolicum.MDA content shows certain daily changes with water potential in three plants.Concretely,MDA contents have an increasing tendency with the decrease of water potential before 10 A.M.,and then decrease continually till 6 P.M.except that in Elaeagnus angustifolia,which begin to increase at 2 P.M.Therefore anti-oxidative ability of Elaeagnus angustifolia is weaker than that of Haloxylon ammodendron,Calligonum mongolicum.Three plants show different means to cope with oxidation stress.SOD activity and ASA content in Calligonum mongolicum is much higher than that in Haloxylon ammodendron and Elaeagnus angustifolia,which result a lower O~(-)_2 content.With regard to the cleanse of toxic H_2O_2,antioxidant GSH works chiefly in Calligonum mongolicum,whereas CAT takes a major role in Haloxylon ammodendron among the antioxidant defense systems with antioxidant working auxiliary.AsA-POD is the main antioxidant enzyme that cleanses toxic H_2O_2 in Elaeagnus angustifolia with other antioxidants co-acting.A balance of all enzymes is kept in plant when they resist the adverse stresses.
[��������: 1]
DOI:10.1038/emboj.2010.261URL [��������: 1]
.
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}