 ,**, 崔翠,**, 叶桑**, 崔明圣**, 赵愉风**, 林呐**, 唐章林**, 李加纳**, 周清元,*西南大学农学与生物科技学院, 重庆400715
,**, 崔翠,**, 叶桑**, 崔明圣**, 赵愉风**, 林呐**, 唐章林**, 李加纳**, 周清元,*西南大学农学与生物科技学院, 重庆400715Screening and Comprehensive Evaluation of Germplasm Resources with Tribenuron-methyl Tolerance at Germination Stage in Rapeseed (Brassica napus L.)
WANG Qian,**, CUI Cui,**, YE Sang**, CUI Ming-Sheng**, ZHAO Yu-Feng**, LIN Na**, TANG Zhang-Lin**, LI Jia-Na**, ZHOU Qing-Yuan,*College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China通讯作者:
第一联系人:
收稿日期:2017-11-20接受日期:2018-03-26网络出版日期:2018-04-18
| 基金资助: |
Received:2017-11-20Accepted:2018-03-26Online:2018-04-18
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (426KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王倩, 崔翠, 叶桑, 崔明圣, 赵愉风, 林呐, 唐章林, 李加纳, 周清元. 甘蓝型油菜种子萌发期耐苯磺隆种质筛选与综合评价[J]. 作物学报, 2018, 44(8): 1169-1184. doi:10.3724/SP.J.1006.2018.01169
WANG Qian, CUI Cui, YE Sang, CUI Ming-Sheng, ZHAO Yu-Feng, LIN Na, TANG Zhang-Lin, LI Jia-Na, ZHOU Qing-Yuan.
自1942年除草剂2,4-D问世以来, 人类使用化学除草剂防除农田杂草已有70多年的历史。西方发达国家已经在85%~100%的作物上使用除草剂[1], 我国除草剂生产和使用量也逐年增加[2]。但是, 在所施用的除草剂中, 绝大部分都残留于土壤或淋溶于水中, 真正起作用的很少[3], 极容易导致对除草剂敏感的作物死亡[4]。不科学地使用半衰期长的除草剂还会毒害下茬作物[5]。因此, 既要高效地消灭杂草, 又不使作物受到药害, 可以通过新型除草剂产品的研发和耐除草剂作物品种的培育两种途径实现, 其中后者相对比较经济, 费用一般不超过研制新除草剂的5%[6]。发掘农作物除草剂耐性基因、培育耐除草剂农作物新品种成为作物遗传育种的重要方向之一。
油菜是当今世界的主要油料作物之一, 我国油菜的种植面积、总产量均约占世界的1/3[7]。田间杂草危害可使油菜产量下降15%, 更甚者可减产50%以上[8,9]。我国油菜主产区的杂草类型大致可分为以禾本科杂草为主、以阔叶杂草为主以及禾本科与阔叶草混生3种[10]。油菜为阔叶型植物, 所以阔叶杂草就成为油菜田杂草防除的一个关键瓶颈[11]。苯磺隆作为阔叶类杂草除草剂, 在小麦田除草取得良好效果[12], 若要选育出耐苯磺隆的油菜品种, 将苯磺隆和现有油菜田单子叶除草剂混合使用, 可为油菜田杂草的有效防除提供一条新的途径[13]。因此, 培育和推广耐苯磺隆除草剂油菜品种可以有效控制阔叶草害, 减少中耕除草用工,提高油菜产量, 增加效益[14]。另外, 苯磺隆作为化学杀雄剂在油菜杂种优势利用研究中效果良好, 已经引起广泛重视[15,16], 选育具有苯磺隆除草剂耐性的甘蓝型油菜作为父本,有利于简化制种程序, 降低制种成本[16,17]。信晓阳等[14]使用苯磺隆溶液处理49个不同基因型油菜幼苗, 筛选出2个在苗期低敏感材料。曲高平等[18]用甲基磺酸乙酯(EMS)溶液对甘蓝型油菜中双9号种子诱变处理, 在M2突变体库中筛选到3株苯磺隆耐性突变体。华中农业大学报道从华双5号油菜EMS突变后代群体中鉴定出几株苯磺隆耐性突变体, 测序表明它们的突变位点均为BnALS3的Pro197Ser/ Leu突变[16]。吴学莉等[17]从一个耐苯磺隆除草剂的播娘蒿天然突变体中克隆了乙酰乳酸合成酶(ALS)基因DsALS-108, 并通过转基因发现其表达使甘蓝型油菜对苯磺隆的耐性提高至野生型致死浓度的3倍。汪亚琴[19]以甘蓝型油菜品系甲572为受体, 转化水稻细胞色素P450基因家族成员CYP81A6基因, 获得了抗苯磺隆的油菜株系。有研究表明植物耐受苯磺隆除草剂主要与ALS基因突变和非靶标酶代谢解毒能力有关[1,20-24]。同时, 田间杂草对苯磺隆耐性突变体的机制研究也相继报道, 其中黑草和看麦娘的AmGSTF1基因在拟南芥中表达, 获得的转基因植物可表现抗多种除草剂[25,26,27,28]。在耐除草剂作物的培育中, 鉴定并获得具有天然耐药性的植物是研究的关键[29], 因此, 从现有油菜中筛选耐苯磺隆种质资源, 并挖掘内源耐苯磺隆基因源对于油菜育种具有重要的理论和应用价值。
虽然在4种磺酰脲类除草剂中, 苯磺隆的植物毒性最低[30], 且苯磺隆为短残留除草剂[31], 但是也有报道指出小麦地土壤中苯磺隆降解半衰期为5~12 d [32,33], 因此, 苯磺隆仍存在短期土壤残留现象。目前, 关于苯磺隆土壤残留对作物种子, 尤其是对油菜种子萌发的影响未见报道。本研究拟综合鉴定与评价供试品种(系)的发芽率、发芽势、鲜重、干重、根长和芽长, 筛选出萌发期耐性较强的油菜种质及相关性较强的测定指标, 研究播种期苯磺隆残留对油菜种子萌发的影响, 为苗期、蕾薹期、花期筛选耐苯磺隆油菜品种(系)提供基础, 也为进一步在遗传和育种研究中利用这些优异种质提供理论依据。
1 材料与方法
1.1 供试油菜种质资源
试验材料为国内外各农业高校、科研院所选育或收集的, 具有不同遗传背景和广泛地理来源的241份甘蓝型油菜种质(附表1), 该群体包含部分自交系和常规品种, 种子均由重庆市油菜工程技术研究中心提供。处理药剂为10%苯磺隆除草剂(江苏瑞东农药有限公司), 有效成分含量100 g L-1, 生产用量每公顷150 g兑水600 kg (250 mg kg-1)。
1.2 方法
1.2.1 萌发期最适苯磺隆处理浓度筛选 参照陈新等[34]的培养皿纸上发芽法并略有改动。以遗传背景不同的11份甘蓝型油菜品种(系)为材料(附表1), 对每个品种设置蒸馏水对照和不同浓度(250、25、2.5、0.25 mg kg-1)苯磺隆胁迫共5种处理, 各处理3次重复。在铺有2层滤纸的培养皿中加入3βmL不同浓度的苯磺隆溶液, 对照中加入相同体积蒸馏水, 将已清洗的饱满种子均匀放在滤纸上, 每皿20粒, 盖上皿盖, 于人工培养箱中培养, 设置温度为25℃, 相对湿度为85%, 光照和黑暗时间为16 h/8 h。参照汪梦竹等[35]的研究, 于第7天测定油菜主根根长, 分析并确定苯磺隆最佳胁迫浓度。1.2.2 萌发期耐苯磺隆鉴定 用1.2.1中确定的最佳处理浓度, 以蒸馏水为对照, 依照上述方法培养241份甘蓝型油菜(附表1), 3次重复。于第3天统计种子发芽势(germination vigor, GV), 第7天统计种子发芽率(germination rate, GR), 随机选取每个重复10株幼苗分别测定其茎长(shoot length, SL)、根长(root length, RL)、鲜重(fresh weight, FW)和干重(dry weight, DW)形态指标。
1.3 数据处理与分析
利用Microsoft Excel 2013、IBM SPSS19.0[36]和DPS 2006[37]统计软件整理分析数据。参考闫峰等[38]、Upadhyaya[39]、汪灿等[40]的方法, 以各品种6个性状测定值作为基础数据, 分析其平均数差异显著性, 按公式(1)计算对苯磺隆单项耐性系数(tribenuron-methyl tolerance coefficient, TC), 式中Xij、CKij分别表示第i个指标下第j个品种(系)苯磺隆胁迫和对照处理的指标测定值。针对各指标TC值, 进行简单相关分析、连续次数分布统计分析和主成分分析。
TC =$\frac{{{X}_{ij}}}{\text{C}{{\text{K}}_{ij}}}$ i =1, 2, 3, …, n; j=1, 2, 3, …, m (1)
按公式(2)、(3)和(4)计算取各种质各指标的隶属函数值[R(Xij)]和平均隶属函数值(ASF, average subordinative function value)。
ωi = Pi /$\sum{_{i=1}^{k}}Pi$ i = 1, 2, 3, …, k (2)
R(Xij) = (Xij - Xij,min) /( Xij,max - Xij,min) i = 1, 2, 3…, n; j = 1, 2, 3…, m (3)
ASF =$\frac{1}{n}\sum{_{i=1}^{k}}R({{X}_{i}})$ i = 1, 2, 3, …, n (4)
式中, Pi为第i个综合指标贡献率, 表示第i个指标在所有指标中的重要程度。Xij、Xij,min、Xij,max分别表示第i个指标下第j个品种的TC值及第i个指标下的最小值和最大值。其中k为选取的主成分个数。
根据因子权重系数(ωi)和各主成分得分值[R’(Xi)], 按公式(5)计算苯磺隆耐性度量值(T, tribenuron-methyl tolerance comprehensive evaluation value)。
T =$\sum{_{i=1}^{k}}[{R}'({{X}_{i}})\times ({{P}_{i}}\div \sum{_{i=1}^{k}}{{P}_{i}})]$ i = 1, 2, 3, …, k (5)
按公式(6)、(7)计算关联系数(ξi)及等权关联度(γi)。
ξi = $\frac{\min {{\Delta }_{ij}}+p\max {{\Delta }_{ij}}}{{{\Delta }_{ij}}+p\max {{\Delta }_{ij}}}$ i, j = 1, 2, 3, …, n(6)
γi =$\frac{1}{n}\sum{_{i=1}^{n}{{\xi }_{i}}}$ i= 1, 2, 3, …, n (7)
式中, ξi为关联系数, Δij为品种最优性状与第j个品种(系)第i个性状的绝对差值。minΔij为最小二级绝对差值, maxΔij为最大二级绝对差值, p为分辨系数(取0.5)。
分别按公式(8)和(9)计算各指标权重系数[ωi (γ)]和加权关联度(weight tribenuron-methyl tolerance coefficient, WTC)。
ωi (γ) = γi /$\sum{_{i=1}^{n}{{\gamma }_{i}}}$ i=1, 2, 3,…, n (8)
WTC =$\sum{_{i=1}^{n}[\text{TC}\times ({{\gamma }_{i}}/\sum{_{i=1}^{n}{{\gamma }_{i}}})]}$ i = 1, 2, 3,…, n
(9)
式中, γi为各指标关联度, TC为对苯磺隆单项耐性系数。
以各指标TC值为比较序列, 分别以T值和WTC值为参考序列进行灰色关联度分析, 获得各指标TC值与T值间的关联度(γT)和TC值与WTC值间的关联度(γWTC)。
最后基于供试甘蓝型油菜种质T值, 采用加权配对算术平均法(weighted pair group method average, WPGMA)和欧氏距离进行聚类分析, 划分苯磺隆耐性等级, 并分别以T值、ASF值和WTC值为参考序列, 对各指标TC值进行逐步回归分析, 获得回归方程。
2 结果与分析
2.1 苯磺隆胁迫浓度筛选
11份甘蓝型油菜品种在不同苯磺隆浓度处理条件下, 根长均受到不同程度的抑制(表1)。在0、0.25、2.5、25和250 mg kg-1浓度下, 根长均值分别为7.73、4.44、1.62、1.26和1.15 cm, 处理较对照分别下降36.12%、79.01%、83.64%和85.15%, 根长在2.5 mg kg-1处理时下降接近最大值, 到25 mg kg-1和250 mg kg-1处理时下降趋势变缓。11份品种在5个处理中差异均具有统计学意义, 但是25 mg kg-1胁迫处理时, 更能够区分材料本身的差异; 在25 mg kg-1具有差异的材料, 在0.25、2.5和250 mg kg-1胁迫处理均没有差异(表1)。如25 mg kg-1胁迫下, P18与所有品种(系)间差异均显著, 甲预31棚与中双12等5个品种差异显著。2.5 mg kg-1胁迫下, P18与至尊和B265品种(系)间差异不显著, 甲预31棚与6个品种(系)差异不显著。250 mg kg-1胁迫下, P18与至尊品种(系)间差异不显著, 甲预31棚与9个品种差异不显著。另外, 250 mg kg-1胁迫处理7 d后, 各品种根部萎缩生长普遍变差。因此, 25 mg kg-1浓度可作为鉴定甘蓝型油菜萌发期耐苯磺隆的适宜浓度。Table 1
表1
表1不同浓度下11份品种根长相对值的显著性分析
Table 1
| 品种(系) Variety (line) | 苯磺隆浓度 Concentration of tribenuron-methyl | |||||||
|---|---|---|---|---|---|---|---|---|
| 0.25 mg kg-1 | 2.50 mg kg-1 | 25.00 mg kg-1 | 250.00 mg kg-1 | |||||
| 炎81-2 Yan 81-2 | 70.3±0.007 | c | 17.8±0.015 | de | 10.4±0.003 | f | 10.3±0.003 | de |
| SWU92 | 82.5±0.030 | b | 19.3±0.009 | de | 14.5±0.002 | def | 11.7±0.011 | de |
| SWU101 | 64.5±0.077 | cd | 15.3±0.017 | de | 12.5±0.001 | ef | 9.3±0.012 | e |
| 782 | 48.5±0.003 | e | 18.5±0.004 | de | 13.5±0.006 | ef | 12.9±0.002 | cde |
| 509 | 50.0±0.008 | e | 19.8±0.021 | de | 15.4±0.004 | cdef | 14.5±0.009 | bcde |
| P18 | 106.3±0.091 | a | 36.0±0.031 | a | 31.1±0.002 | a | 28.8±0.016 | a |
| 甲预31棚 Jiayu 31 peng | 24.6±0.010 | f | 22.4±0.013 | cd | 19.6±0.029 | bcd | 15.8±0.006 | bcde |
| 苏油1号 Suyou 1 | 95.7±0.039 | a | 26.1±0.008 | bc | 23.8±0.012 | b | 20.0±0.004 | bc |
| 至尊 Zhizun | 85.2±0.002 | b | 29.3±0.030 | ab | 22.3±0.001 | bc | 21.4±0.019 | ab |
| B265 | 96.4±0.044 | a | 31.1±0.054 | ab | 18.5±0.011 | bcde | 18.0±0.006 | bcd |
| 中双12 Zhongshuang 12 | 60.1±0.010 | d | 13.3±0.017 | e | 13.2±0.038 | ef | 13.5±0.059 | cde |
新窗口打开|下载CSV
2.2 耐苯磺隆油菜种质资源筛选
2.2.1 241份甘蓝型油菜萌发期指标 对照和处理(25 mg kg-1苯磺隆胁迫)的6个性状在品种间差异均具有统计学意义, 各性状的变异系数分别为16.45%~54.56%和20.01%~54.83%, 说明试验中的241份种质在两个条件下均表现出广泛的遗传变异。在苯磺隆胁迫下, 各性状受到影响程度不同。胁迫处理下根长均值仅为0.94 cm, 较对照下降87.99%, 受抑制最严重。胁迫处理下的发芽势、发芽率、鲜重均值分别为75.77%、83.22%、0.36 g, 较对照分别下降6.81%、5.37%、5.26%。胁迫处理下芽长均值为2.37 cm, 较对照增加4.61%。在胁迫条件下, 根长、芽长、发芽势、发芽率的变异系数均增加, 其中根长的变异系数较对照增加最明显(表2)。Table 2
表2
表2苯磺隆胁迫下油菜萌发期各指标的变化
Table 2
| 项目 Item | 参数 Parameter | 根长 RL (cm) | 芽长 SL (cm) | 发芽势 GV (%) | 发芽率 GR (%) | 鲜重 FW (g) | 干重 DW (g) | |
|---|---|---|---|---|---|---|---|---|
| 对照 Control | 最大值 Max. | 13.13 | 5.81 | 100.00 | 100.00 | 0.79 | 0.08 | |
| 最小值 Min. | 0.55 | 0.76 | 10.00 | 15.00 | 0.08 | 0.01 | ||
| 均值 Average | 7.38** | 2.27** | 81.31** | 87.94** | 0.38** | 0.04** | ||
| 标准差 SE | 1.61 | 1.24 | 19.87 | 14.47 | 0.11 | 0.01 | ||
| 变异系数 CV (%) | 21.77 | 54.56 | 24.44 | 16.45 | 28.91 | 26.74 | ||
| 胁迫 Stress | 最大值 Max. | 2.97 | 6.24 | 100.00 | 100.00 | 0.79 | 0.06 | |
| 最小值 Min. | 0.34 | 0.71 | 5.00 | 15.00 | 0.12 | 0.01 | ||
| 均值 Average | 0.94** | 2.37** | 75.77** | 83.22** | 0.36** | 0.04** | ||
| 标准差 SE | 0.38 | 1.30 | 21.47 | 16.66 | 0.10 | 0.01 | ||
| 变异系数 CV (%) | 40.00 | 54.83 | 28.34 | 20.01 | 28.00 | 25.91 | ||
| 较对照变化 | 均值 Average | -6.44 | 0.10 | -5.54 | -4.72 | -0.02 | 0 | |
| Comparison with the control | 变异系数 CV (%) | 18.23 | 0.27 | 3.90 | 3.56 | -0.91 | -0.83 | |
新窗口打开|下载CSV
2.2.2 单项指标分析 供试种质在苯磺隆胁迫处理后, 与对照相比各指标均发生不同程度的变化(表3)。同一指标下不同品种对苯磺隆单项耐性系数(TC)的最大值和最小值之间差异较大, 说明不同品种间对苯磺隆的耐性不同。不同指标下TC值存在明显差异, 变异系数介于10.3%~49.0%之间, 表明各指标对苯磺隆胁迫反应的敏感性不同。此外对各单项指标进行单因素方差分析, 表明不同种质间在根长、芽长、发芽率、发芽势、鲜重5个指标下均存在具有统计学意义的差异。相关分析表明(表4), 各指标间存在一定程度的相关性。其中干重与根长呈显著负相关, 与发芽率呈显著正相关, 而与其余指标相关不显著。发芽势与发芽率呈极显著正相关, 与根长呈极显著负相关, 相关系数分别为0.687和-0.150。发芽率与根长呈显著负相关, 鲜重与芽长呈极显著正相关, 其余指标间无显著相关性。除发芽势与发芽率相关系数较高, 其余相关系数绝对值均在0.1~0.3之间。此外, 各指标TC值分布次数相差较大(图1)。根长的TC值在0.095~0.215之间的品种最多。TC>0.90的鲜重、发芽势、发芽率、芽长、干重的分布频率分别为68.0%、70.5%、80.5%、81.3%和96.3%, 各指标对苯磺隆胁迫的敏感性由强到弱依次是根长、鲜重、发芽势、发芽率、芽长、干重。因此, 如果直接采用各个指标, 由于指标间信息的叠加重合, 很难准确客观地评价各种质的耐性, 从而影响对苯磺隆耐性鉴定结果。
图1
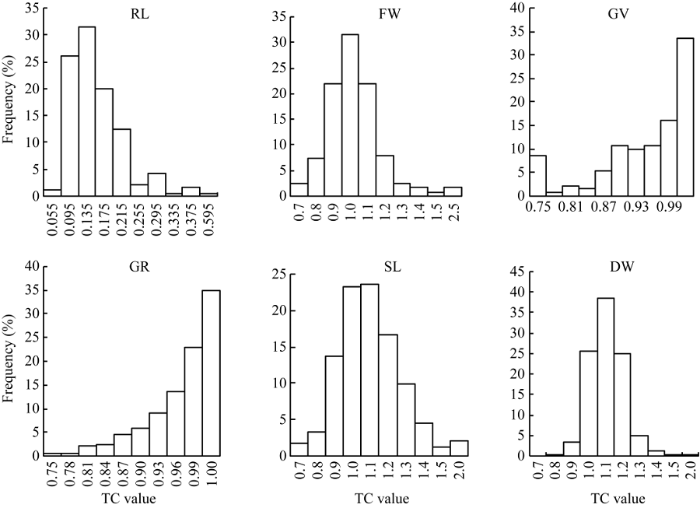 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1241份甘蓝型油菜不同指标耐性系数(TC)的频次分布
RL: 根长; FW: 鲜重; GV: 发芽势; GR: 发芽率; SL: 芽长; DW: 干重。
Fig. 1Frequency distribution of tribenuron-methyl tolerance coefficients (TC) of different indexes in 241 Brassica napus
RL: root length; SL: shoot length; GV: germination vigor; GR: germination rate; FW: fresh weight; DW: dry weight.
Table 3
表3
表3供试油菜种质各指标的苯磺隆耐性系数
Table 3
| 参数 Parameter | 根长 RL | 芽长 | 发芽势 | 发芽率 | 鲜重 | 干重 |
|---|---|---|---|---|---|---|
| SL | GV | GR | FW | DW | ||
| 最大值 Max. | 0.588 | 2.005 | 1.000 | 1.000 | 2.357 | 1.946 |
| 最小值 Min. | 0.050 | 0.494 | 0.143 | 0.333 | 0.411 | 0.333 |
| 均值 Average | 0.136** | 1.052** | 0.926** | 0.942** | 0.980** | 1.055 |
| 标准差 SE | 0.066 | 0.187 | 0.123 | 0.097 | 0.202 | 0.136 |
| 变异系数 CV (%) | 48.950 | 17.800 | 13.269 | 10.276 | 20.593 | 12.856 |
新窗口打开|下载CSV
Table 4
表4
表4供试油菜种质各指标耐性系数的相关性
Table 4
| 指标 Index | 根长 RL | 芽长 SL | 鲜重 FW | 发芽率 GR | 发芽势 GV | 干重 DW |
|---|---|---|---|---|---|---|
| RL | 1 | |||||
| SL | 0.046 | 1 | ||||
| FW | 0.042 | 0.215** | 1 | |||
| GR | -0.150* | -0.059 | -0.018 | 1 | ||
| GV | -0.171** | 0.031 | 0.022 | 0.687** | 1 | |
| DW | -0.128* | 0.031 | 0.062 | 0.133* | 0.097 | 1 |
新窗口打开|下载CSV
2.2.3 萌发期主成分分析 由表5可知, 第1、第2、第3和第4主成分的贡献率分别为30.17%、21.83%、18.27%和13.16%, 累计贡献率达到83.42%, 已基本代表了所测指标的信息, 可反映影响萌发期油菜耐苯磺隆性主导因素。从各个指标在综合指标的贡献率及主成分特征向量值可以看出, 第1主成分中发芽率、发芽势所占的比率较大, 第2主成分中鲜重所占的比率较大, 第3主成分中根长所占的比率较大, 第4主成分中芽长所占比率较大。
Table 5
表5
表5各综合指标的特征值、贡献率和主成分特征向量值
Table 5
| 主成分 Principal component | 特征值 Eigen value | 贡献率 Variance contribution (%) | 累计贡献率 Accumulated variance contribution (%) | 所测指标的特征向量值 Eigen vector of measured indicators | |||||
|---|---|---|---|---|---|---|---|---|---|
| 发芽率 GR | 发芽势GV | 鲜重 FW | 根长 RL | 芽长 SL | 干重 DW | ||||
| 1 | 1.810 | 30.167 | 30.167 | 0.978 | 0.945 | 0.279 | 0.011 | 0.261 | 0.617 |
| 2 | 1.310 | 21.825 | 51.992 | -0.358 | -0.263 | 0.841 | 0.396 | 0.769 | 0.258 |
| 3 | 1.096 | 18.267 | 70.259 | 0.115 | 0.311 | -0.263 | 0.904 | 0.195 | -0.637 |
| 4 | 0.790 | 13.159 | 83.418 | -0.044 | -0.057 | 0.316 | 0.508 | -0.755 | 0.323 |
新窗口打开|下载CSV
2.2.4 甘蓝型油菜品种资源耐苯磺隆性综合评价
供试种质平均隶属函数值(ASF值)介于0.155~ 0.687之间, 平均值均为0.485, 变异系数为15.3%, 根据ASF值大小对供试种质进行耐性排序, 其结果见表6。其中, 对苯磺隆耐性强的品种(系)有SWU95、SWU113、希望106和WH-33, 耐性弱的品种(系)有10-804、中双7号、中油589和10-1043, 其余种质介于两者之间。供试种质加权关联系数(WTC值)介于0.360~0.615之间, 平均值均为0.508, 变异系数为8.14%。根据WTC值大小对供试种质进行耐性排序, 对苯磺隆耐性强的品种(系)有希望106、WH-33、SWU113和SWU95, 耐性弱的品种(系)有中双7号、中油589和10-1043, 其余种质介于两者之间(表6)。供试种质苯磺隆耐性度量值(T值)介于-2.58~ +2.22之间, 根据T值大小对供试种质进行耐性排序, 对苯磺隆耐性强的品种(系)有希望106、SWU95和WH-33, 耐性弱的品种(系)有中油589和10-1043, 其余种质介于两者之间(表6)。三者之间的鉴定结果有所区别, 但供试种质耐性较强较弱的鉴定结果基本一致。为方便比较, 将ASF值、WTC值和T值的排序数值相加, 再根据累加值从小到大进行排序, 从241份种质资源的综合排序看, 其中排名前五的依次为希望106、SWU95、WH-33、SWU113和97097。从4种排序方法看, 希望106、SWU95、WH-33均表现为对苯磺隆耐性强。
Table 6
表6
表6苯磺隆胁迫下油菜品种资源的ASF值、WTC值、T值和综合排序
Table 6
| 编号 Code | ASF值 ASF value | WTC值 WTC value | T值 T value | 综合排序 Final rank | |||
|---|---|---|---|---|---|---|---|
| 数值Value | 排序Order | 数值Value | 排序Order | 数值Value | 排序Order | ||
| 1 | 0.55 | 35 | 0.55 | 30 | 0.28 | 73 | 40 |
| 2 | 0.46 | 167 | 0.53 | 67 | -0.30 | 178 | 141 |
| 3 | 0.49 | 127 | 0.55 | 32 | 0.07 | 108 | 81 |
| 4 | 0.48 | 138 | 0.54 | 62 | -0.16 | 154 | 124 |
| 5 | 0.44 | 192 | 0.50 | 164 | -0.55 | 201 | 192 |
| 6 | 0.57 | 24 | 0.55 | 26 | 0.65 | 31 | 25 |
| 7 | 0.48 | 141 | 0.48 | 194 | -0.29 | 177 | 175 |
| 8 | 0.42 | 213 | 0.46 | 208 | -0.53 | 199 | 212 |
| 9 | 0.47 | 148 | 0.52 | 101 | -0.29 | 176 | 148 |
| 10 | 0.54 | 49 | 0.55 | 33 | 0.55 | 35 | 35 |
| 11 | 0.47 | 147 | 0.52 | 114 | -0.14 | 145 | 138 |
| 12 | 0.52 | 78 | 0.52 | 121 | 0.05 | 111 | 103 |
| 13 | 0.45 | 177 | 0.52 | 106 | -0.62 | 207 | 165 |
| 14 | 0.51 | 84 | 0.53 | 84 | 0.23 | 80 | 74 |
| 15 | 0.42 | 206 | 0.52 | 100 | -0.66 | 215 | 178 |
| 16 | 0.47 | 150 | 0.49 | 174 | -0.01 | 121 | 153 |
| 17 | 0.47 | 154 | 0.50 | 145 | -0.15 | 151 | 154 |
| 18 | 0.50 | 98 | 0.53 | 83 | 0.10 | 105 | 85 |
| 19 | 0.42 | 203 | 0.49 | 178 | -0.72 | 218 | 209 |
| 20 | 0.50 | 110 | 0.52 | 99 | 0.13 | 98 | 100 |
| 21 | 0.45 | 184 | 0.50 | 144 | -0.55 | 203 | 181 |
| 22 | 0.44 | 194 | 0.51 | 132 | -0.52 | 198 | 179 |
| 23 | 0.44 | 196 | 0.48 | 183 | -0.54 | 200 | 202 |
| 24 | 0.49 | 122 | 0.53 | 65 | -0.14 | 146 | 116 |
| 25 | 0.46 | 164 | 0.50 | 163 | -0.12 | 141 | 161 |
| 26 | 0.47 | 155 | 0.51 | 123 | -0.30 | 179 | 158 |
| 27 | 0.40 | 218 | 0.46 | 212 | -0.83 | 227 | 218 |
| 28 | 0.49 | 134 | 0.52 | 94 | 0.05 | 110 | 118 |
| 29 | 0.60 | 11 | 0.57 | 7 | 0.79 | 24 | 11 |
| 30 | 0.42 | 209 | 0.48 | 191 | -0.60 | 206 | 210 |
| 31 | 0.52 | 68 | 0.54 | 57 | 0.02 | 118 | 70 |
| 32 | 0.53 | 58 | 0.52 | 98 | 0.22 | 81 | 67 |
| 33 | 0.59 | 14 | 0.56 | 13 | 0.96 | 15 | 12 |
| 34 | 0.48 | 143 | 0.52 | 104 | -0.21 | 163 | 140 |
| 35 | 0.33 | 235 | 0.41 | 236 | -1.28 | 234 | 235 |
| 36 | 0.16 | 241 | 0.36 | 241 | -2.57 | 241 | 241 |
| 37 | 0.50 | 92 | 0.50 | 141 | -0.01 | 119 | 123 |
| 38 | 0.25 | 238 | 0.38 | 238 | -1.77 | 238 | 238 |
| 39 | 0.48 | 140 | 0.53 | 66 | -0.10 | 138 | 122 |
| 40 | 0.47 | 157 | 0.54 | 49 | -0.07 | 133 | 119 |
| 41 | 0.46 | 169 | 0.53 | 82 | -0.43 | 191 | 152 |
| 42 | 0.50 | 107 | 0.53 | 63 | 0.03 | 116 | 86 |
| 43 | 0.41 | 215 | 0.49 | 166 | -0.57 | 204 | 206 |
| 编号 Code | ASF值 ASF value | WTC值 WTC value | T值 T value | 综合排序 Final rank | |||
| 数值Value | 排序Order | 数值Value | 排序Order | 数值Value | 排序Order | ||
| 44 | 0.46 | 162 | 0.50 | 147 | -0.13 | 143 | 156 |
| 45 | 0.44 | 197 | 0.49 | 175 | -0.39 | 186 | 194 |
| 46 | 0.49 | 120 | 0.54 | 47 | -0.02 | 122 | 87 |
| 47 | 0.39 | 223 | 0.45 | 223 | -0.74 | 219 | 223 |
| 48 | 0.51 | 89 | 0.54 | 40 | 0.35 | 64 | 54 |
| 49 | 0.54 | 43 | 0.53 | 81 | 0.35 | 63 | 52 |
| 50 | 0.53 | 52 | 0.55 | 25 | 0.21 | 82 | 46 |
| 51 | 0.42 | 204 | 0.47 | 206 | -0.36 | 185 | 208 |
| 52 | 0.64 | 5 | 0.57 | 9 | 1.13 | 6 | 5 |
| 53 | 0.50 | 106 | 0.50 | 156 | -0.07 | 135 | 134 |
| 54 | 0.50 | 108 | 0.51 | 137 | 0.36 | 60 | 99 |
| 55 | 0.44 | 187 | 0.48 | 186 | -0.31 | 181 | 191 |
| 56 | 0.46 | 163 | 0.49 | 167 | -0.17 | 157 | 164 |
| 57 | 0.45 | 182 | 0.47 | 197 | -0.24 | 167 | 189 |
| 58 | 0.50 | 103 | 0.54 | 58 | -0.15 | 152 | 106 |
| 59 | 0.46 | 168 | 0.48 | 185 | -0.31 | 180 | 183 |
| 60 | 0.54 | 50 | 0.55 | 27 | 0.46 | 46 | 37 |
| 61 | 0.47 | 160 | 0.53 | 75 | -0.33 | 182 | 144 |
| 62 | 0.51 | 83 | 0.53 | 86 | 0.33 | 67 | 66 |
| 63 | 0.32 | 236 | 0.42 | 235 | -1.41 | 236 | 236 |
| 64 | 0.53 | 60 | 0.54 | 55 | 0.20 | 86 | 58 |
| 65 | 0.54 | 46 | 0.54 | 52 | 0.53 | 39 | 39 |
| 66 | 0.55 | 34 | 0.56 | 14 | 0.53 | 38 | 26 |
| 67 | 0.49 | 124 | 0.53 | 89 | -0.27 | 172 | 131 |
| 68 | 0.50 | 104 | 0.53 | 73 | -0.11 | 140 | 107 |
| 69 | 0.45 | 175 | 0.52 | 105 | -0.39 | 188 | 162 |
| 70 | 0.52 | 71 | 0.52 | 102 | 0.18 | 88 | 78 |
| 71 | 0.39 | 222 | 0.44 | 224 | -0.81 | 225 | 225 |
| 72 | 0.57 | 21 | 0.54 | 42 | 1.00 | 12 | 20 |
| 73 | 0.29 | 237 | 0.41 | 237 | -1.47 | 237 | 237 |
| 74 | 0.53 | 51 | 0.54 | 38 | 0.13 | 99 | 53 |
| 75 | 0.45 | 174 | 0.54 | 61 | -0.26 | 169 | 136 |
| 76 | 0.52 | 69 | 0.50 | 142 | 0.04 | 114 | 112 |
| 77 | 0.51 | 85 | 0.51 | 133 | 0.12 | 100 | 109 |
| 78 | 0.45 | 173 | 0.49 | 181 | -0.17 | 155 | 172 |
| 79 | 0.52 | 72 | 0.52 | 118 | 0.16 | 92 | 84 |
| 80 | 0.40 | 219 | 0.45 | 222 | -0.77 | 223 | 221 |
| 81 | 0.37 | 228 | 0.42 | 234 | -0.75 | 221 | 229 |
| 82 | 0.54 | 42 | 0.51 | 124 | 0.11 | 101 | 82 |
| 83 | 0.42 | 211 | 0.47 | 198 | -0.64 | 212 | 213 |
| 84 | 0.47 | 152 | 0.50 | 152 | 0.02 | 117 | 147 |
| 85 | 0.49 | 133 | 0.50 | 158 | -0.08 | 136 | 150 |
| 86 | 0.45 | 179 | 0.45 | 215 | -0.10 | 137 | 182 |
| 87 | 0.21 | 239 | 0.38 | 239 | -1.99 | 239 | 239 |
| 88 | 0.56 | 31 | 0.54 | 59 | 0.44 | 48 | 41 |
| 89 | 0.53 | 63 | 0.52 | 96 | 0.52 | 40 | 57 |
| 90 | 0.45 | 172 | 0.49 | 169 | -0.15 | 149 | 166 |
| 91 | 0.43 | 202 | 0.46 | 209 | -0.22 | 164 | 201 |
| 92 | 0.50 | 101 | 0.53 | 74 | 0.32 | 69 | 73 |
| 93 | 0.51 | 90 | 0.51 | 139 | 0.35 | 61 | 89 |
| 94 | 0.52 | 77 | 0.56 | 22 | 0.42 | 55 | 45 |
| 95 | 0.47 | 156 | 0.49 | 165 | -0.13 | 144 | 160 |
| 96 | 0.55 | 33 | 0.56 | 16 | 0.43 | 49 | 31 |
| 97 | 0.52 | 75 | 0.53 | 70 | 0.04 | 115 | 77 |
| 98 | 0.53 | 54 | 0.53 | 76 | 0.54 | 37 | 47 |
| 99 | 0.50 | 102 | 0.54 | 45 | 0.15 | 96 | 71 |
| 100 | 0.38 | 226 | 0.45 | 219 | -0.92 | 231 | 227 |
| 101 | 0.45 | 171 | 0.50 | 161 | -0.43 | 192 | 180 |
| 102 | 0.57 | 25 | 0.53 | 72 | 0.90 | 17 | 34 |
| 103 | 0.45 | 176 | 0.51 | 131 | -0.35 | 183 | 167 |
| 104 | 0.53 | 62 | 0.50 | 143 | 0.15 | 95 | 95 |
| 105 | 0.42 | 212 | 0.47 | 205 | -0.68 | 216 | 216 |
| 106 | 0.55 | 36 | 0.52 | 110 | 0.76 | 26 | 48 |
| 107 | 0.59 | 13 | 0.56 | 15 | 0.97 | 14 | 13 |
| 108 | 0.50 | 93 | 0.53 | 79 | -0.15 | 150 | 111 |
| 109 | 0.54 | 44 | 0.54 | 53 | 0.43 | 50 | 44 |
| 110 | 0.51 | 88 | 0.51 | 136 | 0.20 | 83 | 101 |
| 111 | 0.48 | 137 | 0.50 | 148 | -0.04 | 127 | 142 |
| 112 | 0.49 | 119 | 0.54 | 60 | -0.04 | 129 | 102 |
| 113 | 0.52 | 79 | 0.50 | 159 | 0.37 | 59 | 91 |
| 114 | 0.52 | 80 | 0.53 | 64 | 0.20 | 84 | 64 |
| 115 | 0.49 | 125 | 0.52 | 97 | 0.27 | 75 | 92 |
| 116 | 0.49 | 131 | 0.51 | 127 | -0.19 | 162 | 146 |
| 117 | 0.52 | 74 | 0.53 | 85 | 0.40 | 57 | 61 |
| 118 | 0.47 | 151 | 0.52 | 92 | -0.16 | 153 | 133 |
| 119 | 0.49 | 121 | 0.52 | 117 | -0.01 | 120 | 127 |
| 120 | 0.45 | 180 | 0.49 | 180 | -0.51 | 197 | 193 |
| 121 | 0.50 | 99 | 0.52 | 95 | 0.08 | 107 | 96 |
| 122 | 0.42 | 207 | 0.47 | 201 | -0.65 | 213 | 214 |
| 122 | 0.42 | 207 | 0.47 | 201 | -0.65 | 213 | 214 |
| 123 | 0.38 | 225 | 0.43 | 227 | -0.62 | 208 | 219 |
| 124 | 0.47 | 159 | 0.50 | 155 | -0.40 | 189 | 171 |
| 125 | 0.44 | 190 | 0.46 | 207 | -0.19 | 161 | 195 |
| 126 | 0.53 | 65 | 0.51 | 140 | 0.42 | 54 | 75 |
| 127 | 0.44 | 189 | 0.48 | 190 | -0.63 | 209 | 207 |
| 128 | 0.51 | 87 | 0.52 | 109 | -0.05 | 131 | 114 |
| 129 | 0.39 | 221 | 0.44 | 225 | -0.77 | 224 | 224 |
| 130 | 0.42 | 208 | 0.51 | 135 | -0.44 | 193 | 185 |
| 131 | 0.42 | 210 | 0.49 | 176 | -0.17 | 156 | 188 |
| 132 | 0.40 | 217 | 0.46 | 210 | -0.76 | 222 | 217 |
| 133 | 0.61 | 7 | 0.56 | 21 | 0.97 | 13 | 10 |
| 134 | 0.54 | 47 | 0.54 | 51 | 0.51 | 42 | 42 |
| 135 | 0.43 | 200 | 0.48 | 195 | -0.36 | 184 | 203 |
| 136 | 0.41 | 214 | 0.45 | 220 | -0.90 | 230 | 222 |
| 137 | 0.42 | 205 | 0.48 | 192 | -0.71 | 217 | 211 |
| 138 | 0.50 | 111 | 0.53 | 88 | 0.10 | 102 | 97 |
| 139 | 0.50 | 94 | 0.54 | 56 | -0.11 | 139 | 88 |
| 140 | 0.51 | 82 | 0.55 | 34 | 0.17 | 91 | 59 |
| 141 | 0.49 | 126 | 0.52 | 103 | 0.13 | 97 | 113 |
| 142 | 0.58 | 19 | 0.55 | 24 | 0.72 | 27 | 17 |
| 143 | 0.49 | 123 | 0.51 | 128 | -0.04 | 128 | 129 |
| 144 | 0.50 | 109 | 0.53 | 87 | -0.04 | 125 | 110 |
| 145 | 0.47 | 146 | 0.52 | 112 | -0.17 | 159 | 145 |
| 146 | 0.55 | 39 | 0.57 | 10 | 0.80 | 22 | 18 |
| 147 | 0.44 | 198 | 0.49 | 182 | -0.55 | 202 | 204 |
| 148 | 0.55 | 37 | 0.56 | 17 | 0.52 | 41 | 30 |
| 149 | 0.39 | 220 | 0.44 | 226 | -0.81 | 226 | 226 |
| 150 | 0.49 | 129 | 0.52 | 116 | -0.25 | 168 | 143 |
| 151 | 0.50 | 96 | 0.50 | 153 | 0.08 | 106 | 125 |
| 152 | 0.49 | 132 | 0.50 | 162 | -0.07 | 134 | 151 |
| 153 | 0.50 | 113 | 0.52 | 120 | 0.07 | 109 | 120 |
| 154 | 0.49 | 130 | 0.50 | 154 | -0.24 | 166 | 155 |
| 155 | 0.43 | 201 | 0.45 | 217 | -0.65 | 214 | 215 |
| 156 | 0.49 | 128 | 0.52 | 119 | 0.17 | 90 | 117 |
| 157 | 0.59 | 16 | 0.57 | 6 | 0.66 | 30 | 15 |
| 158 | 0.57 | 20 | 0.56 | 23 | 0.60 | 33 | 22 |
| 159 | 0.56 | 26 | 0.56 | 20 | 0.40 | 58 | 32 |
| 160 | 0.50 | 95 | 0.54 | 39 | 0.10 | 103 | 68 |
| 161 | 0.45 | 183 | 0.48 | 184 | -0.45 | 194 | 198 |
| 162 | 0.63 | 6 | 0.56 | 11 | 1.25 | 5 | 6 |
| 163 | 0.44 | 195 | 0.50 | 149 | -0.46 | 195 | 187 |
| 164 | 0.56 | 28 | 0.55 | 29 | 0.89 | 18 | 21 |
| 165 | 0.49 | 117 | 0.46 | 211 | 0.71 | 28 | 126 |
| 166 | 0.35 | 232 | 0.42 | 230 | -1.29 | 235 | 233 |
| 167 | 0.46 | 170 | 0.51 | 130 | -0.49 | 196 | 170 |
| 168 | 0.49 | 135 | 0.53 | 69 | -0.04 | 126 | 115 |
| 169 | 0.59 | 12 | 0.57 | 8 | 0.83 | 20 | 9 |
| 170 | 0.53 | 56 | 0.54 | 50 | 0.30 | 70 | 49 |
| 171 | 0.50 | 114 | 0.50 | 150 | 0.23 | 79 | 121 |
| 172 | 0.47 | 158 | 0.53 | 77 | -0.26 | 170 | 137 |
| 173 | 0.52 | 73 | 0.49 | 173 | 0.35 | 65 | 104 |
| 174 | 0.53 | 64 | 0.54 | 43 | 0.30 | 71 | 50 |
| 175 | 0.51 | 81 | 0.51 | 129 | 0.43 | 51 | 79 |
| 176 | 0.58 | 18 | 0.57 | 5 | 0.54 | 36 | 16 |
| 177 | 0.53 | 55 | 0.55 | 36 | 0.15 | 94 | 51 |
| 178 | 0.49 | 115 | 0.52 | 93 | 0.10 | 104 | 105 |
| 179 | 0.44 | 191 | 0.52 | 108 | -0.64 | 211 | 173 |
| 180 | 0.68 | 1 | 0.58 | 4 | 2.16 | 2 | 2 |
| 181 | 0.52 | 66 | 0.52 | 107 | 0.41 | 56 | 65 |
| 182 | 0.57 | 23 | 0.55 | 31 | 0.79 | 23 | 23 |
| 183 | 0.52 | 70 | 0.51 | 125 | 0.32 | 68 | 80 |
| 184 | 0.44 | 193 | 0.47 | 200 | -0.12 | 142 | 184 |
| 185 | 0.68 | 2 | 0.60 | 2 | 1.12 | 7 | 4 |
| 186 | 0.53 | 53 | 0.51 | 126 | 0.93 | 16 | 55 |
| 187 | 0.48 | 142 | 0.49 | 171 | -0.26 | 171 | 163 |
| 188 | 0.54 | 45 | 0.49 | 179 | 0.86 | 19 | 72 |
| 189 | 0.48 | 139 | 0.48 | 187 | -0.05 | 130 | 157 |
| 190 | 0.37 | 229 | 0.45 | 221 | -1.19 | 233 | 230 |
| 191 | 0.17 | 240 | 0.37 | 240 | -2.14 | 240 | 240 |
| 192 | 0.53 | 61 | 0.52 | 115 | 0.46 | 45 | 63 |
| 193 | 0.38 | 224 | 0.43 | 229 | -0.63 | 210 | 220 |
| 194 | 0.41 | 216 | 0.45 | 218 | -0.14 | 148 | 205 |
| 195 | 0.48 | 145 | 0.53 | 78 | 0.28 | 74 | 93 |
| 196 | 0.44 | 186 | 0.47 | 202 | -0.28 | 173 | 199 |
| 197 | 0.51 | 86 | 0.48 | 188 | 0.51 | 43 | 108 |
| 198 | 0.49 | 118 | 0.50 | 157 | 0.04 | 112 | 132 |
| 199 | 0.67 | 3 | 0.62 | 1 | 2.21 | 1 | 1 |
| 200 | 0.46 | 165 | 0.46 | 213 | -0.28 | 175 | 190 |
| 201 | 0.50 | 97 | 0.52 | 91 | 0.18 | 89 | 83 |
| 202 | 0.45 | 178 | 0.48 | 193 | -0.39 | 187 | 196 |
| 203 | 0.60 | 9 | 0.56 | 18 | 1.03 | 10 | 8 |
| 204 | 0.60 | 8 | 0.53 | 68 | 1.34 | 4 | 24 |
| 205 | 0.46 | 161 | 0.49 | 177 | -0.28 | 174 | 176 |
| 206 | 0.36 | 230 | 0.42 | 233 | -0.89 | 229 | 232 |
| 207 | 0.59 | 15 | 0.56 | 12 | 1.04 | 9 | 7 |
| 208 | 0.56 | 30 | 0.55 | 37 | 0.82 | 21 | 27 |
| 209 | 0.45 | 185 | 0.45 | 216 | -0.23 | 165 | 200 |
| 210 | 0.49 | 116 | 0.50 | 146 | -0.14 | 147 | 139 |
| 211 | 0.59 | 17 | 0.56 | 19 | 1.01 | 11 | 14 |
| 212 | 0.47 | 149 | 0.47 | 203 | -0.17 | 158 | 174 |
| 213 | 0.47 | 153 | 0.46 | 214 | -0.02 | 123 | 168 |
| 214 | 0.56 | 29 | 0.55 | 28 | 0.56 | 34 | 28 |
| 215 | 0.67 | 4 | 0.60 | 3 | 1.85 | 3 | 3 |
| 216 | 0.43 | 199 | 0.47 | 199 | -0.19 | 160 | 197 |
| 217 | 0.36 | 231 | 0.43 | 228 | -0.75 | 220 | 228 |
| 218 | 0.52 | 67 | 0.52 | 90 | 0.20 | 85 | 69 |
| 219 | 0.50 | 105 | 0.51 | 122 | 0.29 | 72 | 94 |
| 220 | 0.45 | 181 | 0.49 | 168 | 0.26 | 76 | 149 |
| 221 | 0.54 | 48 | 0.54 | 44 | 0.43 | 52 | 43 |
| 222 | 0.50 | 112 | 0.49 | 172 | 0.24 | 78 | 128 |
| 223 | 0.57 | 22 | 0.54 | 46 | 0.79 | 25 | 29 |
| 224 | 0.56 | 27 | 0.54 | 48 | 0.67 | 29 | 33 |
| 225 | 0.50 | 91 | 0.51 | 138 | 0.35 | 66 | 90 |
| 226 | 0.48 | 136 | 0.50 | 151 | 0.04 | 113 | 135 |
| 227 | 0.55 | 40 | 0.52 | 111 | 0.35 | 62 | 60 |
| 228 | 0.48 | 144 | 0.48 | 189 | -0.03 | 124 | 159 |
| 229 | 0.60 | 10 | 0.54 | 54 | 1.07 | 8 | 19 |
| 230 | 0.52 | 76 | 0.51 | 134 | 0.16 | 93 | 98 |
| 231 | 0.46 | 166 | 0.48 | 196 | -0.06 | 132 | 169 |
| 232 | 0.44 | 188 | 0.50 | 160 | -0.41 | 190 | 186 |
| 233 | 0.50 | 100 | 0.47 | 204 | 0.25 | 77 | 130 |
| 234 | 0.53 | 57 | 0.49 | 170 | 0.63 | 32 | 76 |
| 235 | 0.37 | 227 | 0.42 | 231 | -0.87 | 228 | 231 |
| 236 | 0.55 | 38 | 0.55 | 35 | 0.48 | 44 | 36 |
| 237 | 0.55 | 32 | 0.52 | 113 | 0.42 | 53 | 56 |
| 238 | 0.54 | 41 | 0.54 | 41 | 0.44 | 47 | 38 |
| 239 | 0.53 | 59 | 0.53 | 71 | 0.19 | 87 | 62 |
| 240 | 0.33 | 234 | 0.53 | 80 | -0.57 | 205 | 177 |
| 241 | 0.34 | 233 | 0.42 | 232 | -1.18 | 232 | 234 |
新窗口打开|下载CSV
2.2.5 灰色关联度分析 各指标TC值与T值间的关联度大小依次为根长、鲜重、发芽率、发芽势、干重和芽长, 反映了各指标TC值与T值的密切程度, 这与各指标对苯磺隆胁迫反应的敏感性基本吻合。各指标TC值与WTC值间的关联度大小依次为根长、鲜重、芽长、发芽势、发芽率和干重(表7)。
Table 7
表7
表7供试油菜种质各指标TC值与T值和WTC值的关联度及各指标权重
Table 7
| 指标 Index | 等权关联度γT | 权重系数 ωi | 加权关联度γWTC | ||
|---|---|---|---|---|---|
| 数值 Value | 排序 Order | 数值 Value | 排序 Order | ||
| 发芽率 GR | 0.649 | 3 | 0.163 | 0.669 | 5 |
| 发芽势 GV | 0.634 | 4 | 0.159 | 0.671 | 4 |
| 鲜重 FW | 0.658 | 2 | 0.165 | 0.745 | 2 |
| 干重 DW | 0.633 | 5 | 0.159 | 0.615 | 6 |
| 根长 RL | 0.781 | 1 | 0.196 | 0.763 | 1 |
| 芽长 SL | 0.630 | 6 | 0.158 | 0.673 | 3 |
新窗口打开|下载CSV
2.2.6 聚类分析及耐性级别的划分 基于T值, 在欧氏距离为0.75处将241份供试种质分为5类(附图1)。其中第I类为高度耐苯磺隆的品种(系), 有SWU95、希望106和WH-33共3份; 第II类为耐性品种(系), 共30份, 占总数的12.4%; 第III类为耐性一般品种(系), 共198份, 占总数的82.2%; 第IV类为不耐苯磺隆的品种(系)共6份; 第V类为高度不耐苯磺隆的品种(系), 有10-1043、10-804、中双7号和中油589共4份。根据供试材料对苯磺隆的耐性聚类分析及耐性级别划分结果, 对其耐性评价指标分级统计表明, 除芽长外, 其余单项指标的隶属函数值、ASF值、WTC值和T值均随对苯磺隆耐性级别的升高而增大(表8)。
Table 8
表8
表8供试油菜种质苯磺隆耐性评价指标的分级
Table 8
| 指标 Index | 隶属函数值 Subordinate function value | ||||
|---|---|---|---|---|---|
| I | II | III | V | IV | |
| 发芽率 GR | 0.94 | 0.94 | 0.93 | 0.84 | 0.48 |
| 发芽势 GV | 0.98 | 0.96 | 0.92 | 0.83 | 0.43 |
| 鲜重 FW | 0.71 | 0.28 | 0.21 | 0.16 | 0.15 |
| 干重 DW | 0.50 | 0.43 | 0.41 | 0.35 | 0.22 |
| 根长 RL | 0.44 | 0.29 | 0.15 | 0.11 | 0.09 |
| 芽长 SL | 0.40 | 0.47 | 0.40 | 0.32 | 0.28 |
| ASF值 ASF value | 0.67 | 0.57 | 0.50 | 0.43 | 0.28 |
| WTC值 WTC value | 0.60 | 0.54 | 0.52 | 0.48 | 0.40 |
| T值 T value | 2.08 | 0.77 | 0.12 | -0.47 | -1.63 |
新窗口打开|下载CSV
2.2.7 回归方程 分别以ASF值、WTC值和T值为参考序列, 对供试种质各指标TC值进行逐步回归分析, 得到的3个回归方程的相关系数r ≈ 1, F检验均达极显著水平(表9), 说明回归方程最优, 模型拟合度好, 用这3个方程进行甘蓝型油菜种质资源萌发期对苯磺隆耐性评价的效果较好。根据T值与各指标TC值的回归方程可知, 在甘蓝型油菜种质资源萌发期耐性鉴定中, 有选择性地测定与T值密切相关的指标, 如根长、鲜重、发芽率可有效鉴定油菜种质资源对苯磺隆的耐性。
Table 9
表9
表9供试油菜种质耐性模型预测
Table 9
| 因变量 Dependent variable | 多元逐步回归方程 Multiple stepwise regression equation | 相关系数 r | F值 F-value | P值 P-value |
|---|---|---|---|---|
| ASF值 ASF value | y = -0.4505+0.6436x1+0.1260x3+0.4108x5+0.1421x6 | 0.9555 | 620 | 0.0001 |
| WTC值 WTC value | y = -0.0506+0.2568x1+0.1867x2+0.3401x3-0.0606x4+0.0910x5+0.0320x6 | 0.9578 | 433 | 0.0001 |
| T值 T value | y = -6.5511+4.5564x1+1.5790x3+5.2021x5 | 0.9247 | 466 | 0.0001 |
新窗口打开|下载CSV
3 讨论
种子萌发是植物生长发育的前提, 是对环境因素最为敏感的阶段。苯磺隆作为一种化学除草剂, 在杀死杂草的同时, 对作物也是一种非生物逆境胁迫。随着化学除草剂在农业生产中的广泛应用, 杂草对化学除草剂的耐性增强, 化学除草剂的使用量大幅度增加, 非合理使用导致化学除草剂在土壤中的残留越来越多, 对种子的萌发影响会越来越大。苯磺隆在现阶段油菜生产中使用量较少, 但播种前结合其他化学除草剂防除田间杂草时, 苯磺隆残留就可能影响油菜种子萌发, 目前未见关于油菜种子萌发过程中受苯磺隆影响的公开报道。本研究认为萌发期最佳苯磺隆胁迫浓度为25 mg kg-1, 为生产上除草剂浓度的1/10, 这一结果与前人[14]筛选浓度不同, 可能与研究对象、处理方式以及测定指标的不同有关。不同作物或同一作物的不同品种对除草剂敏感性不同[14,24,41-46], 植物存在天然耐性基因, 通过除草剂的选择, 可筛选出耐除草剂作物[47]。本研究中25 mg kg-1苯磺隆胁迫条件下, 所有供试油菜种质指标均受到不同程度的影响, 且其影响差异均具统计学意义。牛志峰等[42]研究表明不同小麦品种种芽对苯磺隆的耐药性明显较种根强, 娄国强等[43]研究发现苯磺隆对不同小麦品种安全性存在较大差异, 且对根长的抑制作用明显大于株高。本研究单项耐性系数表明, 根长受抑制最严重, 这与前人研究结果基本一致[29,42,48]。另外, 频次分析表明, 发芽势、发芽率指标在0.90<TC≤1区间的品种分别占供试总材料的70.5%和80.5%, 根长指标在0.095<TC≤ 0.215区间的的品种数最多, 与杜小娟等[49]研究表明的不同苗前除草剂对黄芩种子幼苗生长抑制的结论相符。关联度反映的是各性状组成的比较数列和参考数列间的密切程度[38]。灰色关联度分析表明, TC值与T值间的关联度大小依次为根长、鲜重、发芽率、发芽势、干重、芽长。综合回归方程与T值密切相关, 本研究认为根长、鲜重和发芽率可作为有效鉴定油菜种质资源萌发期苯磺隆耐性的指标。
近年来, 随着作物耐逆性研究的发展, 采用多种综合评价, 已在燕麦[34]、花生[39]、薏苡[40]等作物耐逆性鉴定中得到初步应用, 可以避免单一指标的片面性和不稳定性, 而目前作物耐除草剂研究中评价方法较单一。本研究采用ASF值、T值和WTC值等综合评价其对苯磺隆耐性, 确定T值为主要综合评价指标, 以ASF值和WTC值作为辅助综合评价指标, 进行较确切的聚类分析和耐药型划分, 并结合灰色关联分析、逐步回归分析筛选关键性状指标, 建立了拟合度较好的回归方程。采用ASF值、T值和WTC值对供试油菜种质综合排序, 可消除因各指标单位不同带来的影响。据此综合评价, 选出耐性强的品种(系) SWU95、希望106和WH-33, 耐性弱的品种(系) 10-1043、10-804、中双7号和中油589。种质资源的鉴定结果为油菜其他生育时期耐苯磺隆种质资源筛选和鉴定奠定了基础, 也为油菜耐苯磺隆遗传和育种研究提供了理论依据。
4 结论
采用苯磺隆耐性度量值(T值)、平均隶属函数值(ASF值)和加权耐性系数(WTC值)为综合评价指标, 结合相关分析、频数分析、主成分分析、灰色关联度分析、聚类分析和逐步回归分析等方法, 对241份甘蓝型油菜品种的萌发期耐性鉴定发现, 根长、鲜重、发芽率可作为油菜品种的萌发期耐苯磺隆能力和品种选育时优先考虑指标。根据T值进行聚类分析, 可将供试苯磺隆种质划分为5个耐性级别, 其中I级3份、II级30份、III级198份、IV级6份、V级4份。希望106、SWU95和WH-33为苯磺隆耐性较强的种质, 可作为甘蓝型油菜的苯磺隆耐性育种和耐性机理研究的重要资源。附图和附表 请见网络版: 1) 本刊网站
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]

农药的大量使用,已造成了许多生态环境问题,其中对生物多样性的影响尤为重要.农药的不合理使用,对生物群落的结构与功能产生了严重影响,降低了生物多样性.从昆虫群落结构、土壤中无脊椎动物种群数量、微生物区系和植物群落等方面论述了农药对农业区域生物多样性的影响,并提出了合理使用农药。保护农业区域生物多样性的具体措施.
URL [本文引用: 1]
农药的大量使用,已造成了许多生态环境问题,其中对生物多样性的影响尤为重要.农药的不合理使用,对生物群落的结构与功能产生了严重影响,降低了生物多样性.从昆虫群落结构、土壤中无脊椎动物种群数量、微生物区系和植物群落等方面论述了农药对农业区域生物多样性的影响,并提出了合理使用农药。保护农业区域生物多样性的具体措施.
DOI:10.3969/j.issn.1002-5480.2007.09.017URL [本文引用: 1]
农田杂草与农作物在农田共生,除草剂的选择性效应受多种因素影响而有严格的要求,使用不当必然对非靶植物(农作物)产生一定的影响,伤及作物而表现为药害.除草剂对作物的药害可以是当季作物,而长残效除草剂也可能对后茬敏感作物产生药害,轻者抑制作物生长、减产,重者可致作物死亡,乃至绝产.
DOI:10.3969/j.issn.1002-5480.2007.09.017URL [本文引用: 1]
农田杂草与农作物在农田共生,除草剂的选择性效应受多种因素影响而有严格的要求,使用不当必然对非靶植物(农作物)产生一定的影响,伤及作物而表现为药害.除草剂对作物的药害可以是当季作物,而长残效除草剂也可能对后茬敏感作物产生药害,轻者抑制作物生长、减产,重者可致作物死亡,乃至绝产.
DOI:10.1590/S0100-83582002000200016URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/erh098URL [本文引用: 1]
DOI:10.1111/wre.2005.45.issue-6URL [本文引用: 1]
DOI:10.3969/j.issn.0528-9017.2010.01.048URL [本文引用: 1]
文系统综述近年来油菜田的杂草种类、危害特点以及防治措施的最新研究进展,着重介绍农作防除、化学防除、综合防除、转基因和非转基因抗除草剂油菜选育技术在杂草防治上的研究和应用动态,分析化学防除措施存在的问题和不足。此外,还展望了利用基因工程技术和诱变筛选技术选育创制抗除草剂油菜新品种的发展趋势与应用前景。
DOI:10.3969/j.issn.0528-9017.2010.01.048URL [本文引用: 1]
文系统综述近年来油菜田的杂草种类、危害特点以及防治措施的最新研究进展,着重介绍农作防除、化学防除、综合防除、转基因和非转基因抗除草剂油菜选育技术在杂草防治上的研究和应用动态,分析化学防除措施存在的问题和不足。此外,还展望了利用基因工程技术和诱变筛选技术选育创制抗除草剂油菜新品种的发展趋势与应用前景。
DOI:10.3969/j.issn.1002-5480.2003.12.013URL [本文引用: 1]
本文综述了百草枯抗性杂草的发生、发展情况以及抗性机理方面的报道,并就抗性杂草的治理提出几点建议。
DOI:10.3969/j.issn.1002-5480.2003.12.013URL [本文引用: 1]
本文综述了百草枯抗性杂草的发生、发展情况以及抗性机理方面的报道,并就抗性杂草的治理提出几点建议。
DOI:10.3969/j.issn.1674-098X.2008.15.201URL [本文引用: 1]
本文概述了目前我国油菜生产、种植情况,油菜田发生、危害的主要杂草,我国油菜田登记的除草剂品种和产品情况,并分析了在草害控制过程中存在几个问题.衡量我国油菜上目前登记的除草剂品种、产品、剂型和防治谱,可基本满足我国油菜生产的需要.如使用科学、合理,可避免当茬和后茬药害问题,保障农产品安全,促进我国油菜生产的可持续发展.
DOI:10.3969/j.issn.1674-098X.2008.15.201URL [本文引用: 1]
本文概述了目前我国油菜生产、种植情况,油菜田发生、危害的主要杂草,我国油菜田登记的除草剂品种和产品情况,并分析了在草害控制过程中存在几个问题.衡量我国油菜上目前登记的除草剂品种、产品、剂型和防治谱,可基本满足我国油菜生产的需要.如使用科学、合理,可避免当茬和后茬药害问题,保障农产品安全,促进我国油菜生产的可持续发展.
[本文引用: 1]
[本文引用: 1]
DOI:10.7606/j.issn.1004-1389.2014.07.012URL [本文引用: 4]
首先以中双9号油菜为试材,探索适宜油菜筛选的苯磺隆药液质量浓度和鉴定指标。叶片药害指数、叶片数减少率、叶夹角减少率与植株死亡率间均存在显著或极显著相关关系,说明这3个指标可作为早期鉴定油菜对苯磺隆耐药性的指标。用上述试验筛选出的2.0μg·mL^-1苯磺隆溶液处理49个不同基因型油菜幼苗,根据叶片药害指数、叶片数减少率、叶夹角减少率、植株死亡率4个指标计算出的耐苯磺隆平均隶属函数值(AS)将参试材料分为3个等级,低度敏感材料2份(AS〈0.33)、中度敏感材料22份(0.33≤AS〈0.67)、高度敏感材料25份(AS≥0.67)。参试材料对苯磺隆抗性存在一定差异,但与小麦田推荐的苯磺隆使用质量浓度相比,油菜对苯磺隆抗性水平还很低,说明利用油菜种内基因资源,进行抗苯磺隆油菜育种潜力有限。
DOI:10.7606/j.issn.1004-1389.2014.07.012URL [本文引用: 4]
首先以中双9号油菜为试材,探索适宜油菜筛选的苯磺隆药液质量浓度和鉴定指标。叶片药害指数、叶片数减少率、叶夹角减少率与植株死亡率间均存在显著或极显著相关关系,说明这3个指标可作为早期鉴定油菜对苯磺隆耐药性的指标。用上述试验筛选出的2.0μg·mL^-1苯磺隆溶液处理49个不同基因型油菜幼苗,根据叶片药害指数、叶片数减少率、叶夹角减少率、植株死亡率4个指标计算出的耐苯磺隆平均隶属函数值(AS)将参试材料分为3个等级,低度敏感材料2份(AS〈0.33)、中度敏感材料22份(0.33≤AS〈0.67)、高度敏感材料25份(AS≥0.67)。参试材料对苯磺隆抗性存在一定差异,但与小麦田推荐的苯磺隆使用质量浓度相比,油菜对苯磺隆抗性水平还很低,说明利用油菜种内基因资源,进行抗苯磺隆油菜育种潜力有限。
DOI:10.1111/j.1439-0523.2006.01180.xURL [本文引用: 1]
A sulphonylurea herbicide, tribenuron-methyl, methyl 2-[[[[(4-methoxy-6-methyl-1,3,5-triazin-2-yl) methylamino] carbonyl] amino] sulphonyl] benzoate, was used to induce male sterility in rapeseed. Application of 0.2 μ g tribenuron-methyl per plant at the bolting stage with the longest floral bud <2 mm and repeated 15 days afterwards, resulted in 94.5–100% plants being male sterile in six different breeding lines, but combined with low phytotoxicity. However, excessive double application of the chemical (>0.2 μ g per plant) did have some significant impact on rapeseed, including stunting, fading leaves and petals, reduction in the size of floral parts and a shortened duration of flowering. The percentage of hybrid seeds from '84004' treated with 0.2 μ g × 2 tribenuron-methyl per plant and pollinated by a male parent 'Huaye' was 92.7%, which met the hybridity requirement in China. The results suggest that tribenuron-methyl could be used as an efficient chemical hybridizing agent.
[本文引用: 3]
DOI:10.3969/j.issn.1674-7968.2016.04.001URL [本文引用: 2]
利用化学杀雄剂如苯磺隆(tribenuron-methyl,TBM)除草剂诱导雄性不育,是甘蓝型油菜(Brassica napus)杂种优势应用的主要途径之一。选育具有TBM除草剂抗性的甘蓝型油菜作为父本,有利于简化制种程序,降低制种成本,对提高甘蓝型油菜的产量和品质具有重要意义。乙酰乳酸合成酶(acetolactate synthase,ALS)是合成支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)的关键酶。ALS抑制类除草剂以乙酰乳酸合成酶为靶标,通过抑制其活性阻碍支链氨基酸的合成,导致植株死亡。TBM属于一种磺酰脲类(sulfonylureas,SU)ALS抑制类除草剂。为了改良甘蓝型油菜抗除草剂的遗传特性,本研究首先从一个抗TBM除草剂的播娘蒿(Descurainia sophia)天然突变体(#108)中克隆了ALS基因(Gen Bank登录号:EU520490),命名为Ds ALS-108。和拟南芥(Arabidopsis thaliana)乙酰乳酸合成酶At ALS蛋白序列的比对结果显示,保守氨基酸位点脯氨酸197在突变型蛋白Ds ALS-108中替换为苏氨酸(即P197T)。为了验证含有P197T突变的Ds ALS-108基因是否使植株产生TBM除草剂抗性的原因,构建表达载体PBI121-Ds ALS-108,并通过农杆菌(Agrobacterium tumefaciens)介导法将该载体转化拟南芥,共获得12个阳性转基因株系。喷洒浓度大于等于1.0×10-4g/L TBM后,野生型植株停止生长,趋于死亡,而拟南芥转化植株均能正常生长,说明表达突变基因Ds ALS-108致使拟南芥植株抗TBM除草剂。在甘蓝型油菜转化中,以下胚轴为外植体,通过农杆菌介导法将载体p CAMBIA3301-Ds ALS-108导入甘蓝型油菜中,共获得23个阳性转基因株系。结果发现,Ds ALS-108的表达使甘蓝型油菜对苯磺隆的抗性提高至野生型致死浓度的3倍,即7.5×10-3g/L。而且,当用化学杀雄使用剂量(0.05 mg/L TBM)处理甘蓝型油菜转化植株后,其花粉育性正常,说明该转化植株可作为父本应用于化学诱导的雄性不育
DOI:10.3969/j.issn.1674-7968.2016.04.001URL [本文引用: 2]
利用化学杀雄剂如苯磺隆(tribenuron-methyl,TBM)除草剂诱导雄性不育,是甘蓝型油菜(Brassica napus)杂种优势应用的主要途径之一。选育具有TBM除草剂抗性的甘蓝型油菜作为父本,有利于简化制种程序,降低制种成本,对提高甘蓝型油菜的产量和品质具有重要意义。乙酰乳酸合成酶(acetolactate synthase,ALS)是合成支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)的关键酶。ALS抑制类除草剂以乙酰乳酸合成酶为靶标,通过抑制其活性阻碍支链氨基酸的合成,导致植株死亡。TBM属于一种磺酰脲类(sulfonylureas,SU)ALS抑制类除草剂。为了改良甘蓝型油菜抗除草剂的遗传特性,本研究首先从一个抗TBM除草剂的播娘蒿(Descurainia sophia)天然突变体(#108)中克隆了ALS基因(Gen Bank登录号:EU520490),命名为Ds ALS-108。和拟南芥(Arabidopsis thaliana)乙酰乳酸合成酶At ALS蛋白序列的比对结果显示,保守氨基酸位点脯氨酸197在突变型蛋白Ds ALS-108中替换为苏氨酸(即P197T)。为了验证含有P197T突变的Ds ALS-108基因是否使植株产生TBM除草剂抗性的原因,构建表达载体PBI121-Ds ALS-108,并通过农杆菌(Agrobacterium tumefaciens)介导法将该载体转化拟南芥,共获得12个阳性转基因株系。喷洒浓度大于等于1.0×10-4g/L TBM后,野生型植株停止生长,趋于死亡,而拟南芥转化植株均能正常生长,说明表达突变基因Ds ALS-108致使拟南芥植株抗TBM除草剂。在甘蓝型油菜转化中,以下胚轴为外植体,通过农杆菌介导法将载体p CAMBIA3301-Ds ALS-108导入甘蓝型油菜中,共获得23个阳性转基因株系。结果发现,Ds ALS-108的表达使甘蓝型油菜对苯磺隆的抗性提高至野生型致死浓度的3倍,即7.5×10-3g/L。而且,当用化学杀雄使用剂量(0.05 mg/L TBM)处理甘蓝型油菜转化植株后,其花粉育性正常,说明该转化植株可作为父本应用于化学诱导的雄性不育
DOI:10.7505/j.issn.1007-9084.2014.01.004URL [本文引用: 1]
为了利用诱变技术快速创造变异 类型丰富的油菜新材料用于油菜遗传改良,分别用0.4%、0.8%、1.0%和1.2%的EMS(甲基磺酸乙酯)溶液对甘蓝型油菜中双9号种子进行诱变处 理。结果发现:(1)1.0%的EMS溶液是处理中双9号干种子的最适浓度,4种不同浓度处理M2表型变异率依次为11.26%、14.82%、 27.19%和12.38%;(2)EMS诱变具有器官特异性;(3)M2突变体库中筛选到3株苯磺隆抗性突变体,突变率约为10-4。现已创建了包括子 叶、叶片、花器、株型、角果等多器官变异类型的突变体库,为油菜遗传研究提供了丰富的种质资源。
DOI:10.7505/j.issn.1007-9084.2014.01.004URL [本文引用: 1]
为了利用诱变技术快速创造变异 类型丰富的油菜新材料用于油菜遗传改良,分别用0.4%、0.8%、1.0%和1.2%的EMS(甲基磺酸乙酯)溶液对甘蓝型油菜中双9号种子进行诱变处 理。结果发现:(1)1.0%的EMS溶液是处理中双9号干种子的最适浓度,4种不同浓度处理M2表型变异率依次为11.26%、14.82%、 27.19%和12.38%;(2)EMS诱变具有器官特异性;(3)M2突变体库中筛选到3株苯磺隆抗性突变体,突变率约为10-4。现已创建了包括子 叶、叶片、花器、株型、角果等多器官变异类型的突变体库,为油菜遗传研究提供了丰富的种质资源。
URL [本文引用: 1]
油菜(Brassica napus)是我国主要的油料作物,是食用油、蛋白质和工业原料的重要来源。杂草极大地影响了油菜的产量和品质,因此,培育抗除草剂油菜品种对油菜的生产具有十分重要的意义。苯达松和磺酰脲是两种不同的除草剂,在生产上被广泛用于清除稻田的阔叶杂草。细胞色素P450基因CYP81A6编码一种单加氧酶,能使水稻对苯达松和磺酰脲产生抗性。转入了CYP81A6基因的拟南芥和烟草也能获得对这两种除草剂的抗性。本研究以甘蓝型油菜品系甲572为受体,通过农杆菌(Agrobacterium tumefaciens)介导转化水稻CYP81A6基因,获得了抗苯达松和苯磺隆的油菜株系,为油菜的转基因育种提供了种质资源。主要研究结果如下: (1)以油菜下胚轴为外植体的转化获得了135个bastar抗性植株,PCR检测结果表明其中的37株为阳性,阳性率为27.4%。 (2)利用Southern杂交分析了阳性转基因植株,结果显示目的基因在油菜基因组中的拷贝数为1-4个,在6个植株中是单拷贝插入的。 (3)以β-actin为内参的半定量RT-PCR结果表明CYP81A6基因在所有转基因株系中均能表达,且不同株系之间的表达量有差异。其中CYP81A6-1的表达量最低,株系CYP81A6-2的表达量最高。 (4)待油菜长至4-5片真叶时,分别用不同浓度的苯达松和苯磺隆喷洒T1代植株,进行抗药性鉴定。结果显示非转基因植株用浓度0.24g/L苯达松或1.0μg/ml苯磺隆处理时全部死亡,而转基因植株用1.6g/L苯达松或5.0μg/ml苯磺隆处理时仍能正常生长。当用4.8g/L苯达松或10.0μg/ml苯磺隆处理时,所有的转基因植株和非转基因植株全部死亡。这些结果表明CYP81A6基因能使转基因油菜对苯达松和苯磺隆产生抗性,苯达松的的安全浓度范围为0.24-1.6g/L,苯磺隆的安全浓度范围为1.0-5.0μg/ml.
URL [本文引用: 1]
油菜(Brassica napus)是我国主要的油料作物,是食用油、蛋白质和工业原料的重要来源。杂草极大地影响了油菜的产量和品质,因此,培育抗除草剂油菜品种对油菜的生产具有十分重要的意义。苯达松和磺酰脲是两种不同的除草剂,在生产上被广泛用于清除稻田的阔叶杂草。细胞色素P450基因CYP81A6编码一种单加氧酶,能使水稻对苯达松和磺酰脲产生抗性。转入了CYP81A6基因的拟南芥和烟草也能获得对这两种除草剂的抗性。本研究以甘蓝型油菜品系甲572为受体,通过农杆菌(Agrobacterium tumefaciens)介导转化水稻CYP81A6基因,获得了抗苯达松和苯磺隆的油菜株系,为油菜的转基因育种提供了种质资源。主要研究结果如下: (1)以油菜下胚轴为外植体的转化获得了135个bastar抗性植株,PCR检测结果表明其中的37株为阳性,阳性率为27.4%。 (2)利用Southern杂交分析了阳性转基因植株,结果显示目的基因在油菜基因组中的拷贝数为1-4个,在6个植株中是单拷贝插入的。 (3)以β-actin为内参的半定量RT-PCR结果表明CYP81A6基因在所有转基因株系中均能表达,且不同株系之间的表达量有差异。其中CYP81A6-1的表达量最低,株系CYP81A6-2的表达量最高。 (4)待油菜长至4-5片真叶时,分别用不同浓度的苯达松和苯磺隆喷洒T1代植株,进行抗药性鉴定。结果显示非转基因植株用浓度0.24g/L苯达松或1.0μg/ml苯磺隆处理时全部死亡,而转基因植株用1.6g/L苯达松或5.0μg/ml苯磺隆处理时仍能正常生长。当用4.8g/L苯达松或10.0μg/ml苯磺隆处理时,所有的转基因植株和非转基因植株全部死亡。这些结果表明CYP81A6基因能使转基因油菜对苯达松和苯磺隆产生抗性,苯达松的的安全浓度范围为0.24-1.6g/L,苯磺隆的安全浓度范围为1.0-5.0μg/ml.
DOI:10.3969/j.issn.1003-935X.2015.02.001URL [本文引用: 1]
综述了杂草抗药性产生机理、杂草抗药性演化影响因素,并对其治理途径进行阐述,为杂草抗药性治理提供参考。除草剂抗药机制分为靶标抗性、非靶标抗性,其中靶标抗性包括除草剂作用位点改变、基因倍增及过量表达;非靶标抗性主要包括代谢解毒能力增强、屏蔽作用或与作用位点的隔离作用等。杂草抗药性演化受多种因素共同影响,不仅包括抗药性突变频率、除草剂选择压、杂草适合度及杂草种子库寿命四大因素,还与基因突变和遗传特征直接相关。在未来的杂草治理中,要经常进行田间杂草调查与鉴定,正确使用除草剂(交替使用、混用),并辅以合理的农艺管理措施来减缓杂草抗药性的演化速度。同时,应加强植物间化感作用的基础研究。
DOI:10.3969/j.issn.1003-935X.2015.02.001URL [本文引用: 1]
综述了杂草抗药性产生机理、杂草抗药性演化影响因素,并对其治理途径进行阐述,为杂草抗药性治理提供参考。除草剂抗药机制分为靶标抗性、非靶标抗性,其中靶标抗性包括除草剂作用位点改变、基因倍增及过量表达;非靶标抗性主要包括代谢解毒能力增强、屏蔽作用或与作用位点的隔离作用等。杂草抗药性演化受多种因素共同影响,不仅包括抗药性突变频率、除草剂选择压、杂草适合度及杂草种子库寿命四大因素,还与基因突变和遗传特征直接相关。在未来的杂草治理中,要经常进行田间杂草调查与鉴定,正确使用除草剂(交替使用、混用),并辅以合理的农艺管理措施来减缓杂草抗药性的演化速度。同时,应加强植物间化感作用的基础研究。
URL
随着我国国民经济的快速发展,农村青壮年劳动力大量向城市转移。轻简化、机械化已成为油菜产业发展的主要方向,杂草的综合有效治理是实现油菜机械化、轻简化的重要技术环节。培育和推广抗除草剂的油菜品种是防除田间草害,提高油菜产量和质量最为经济有效的措施。 本研究首先以中双9号油菜为试材,探索了适宜油菜筛选的苯磺隆药液浓度和鉴定指标。结果表明,叶片药害指数、叶片数减少率、叶夹角减少率与植株死亡率之间都存在显著或极显著的相关关系,说明这3个指标可考虑作为早期鉴定油菜对苯磺隆耐药性的指标。用筛选出的适宜浓度苯磺隆溶液(2.0μg mL-1),处理了49个不同基因型油菜的幼苗,根据基于叶片药害指数、叶片数减少率、叶夹角减少率、植株死亡率4个指标计算出的耐苯磺隆平均隶属函数值(AS)将参试材料分为3个等级,低度敏感材料2份(AS 0.33)、中度敏感材料22份(0.33≤AS 0.67)、高度敏感材料25份(AS≥0.67)。虽然参试材料对苯磺隆抗性存在一定的差异,然而与小麦田推荐的苯磺隆使用浓度相比,所有参试油菜材料对苯磺隆抗性水平还很低,说明利用油菜种内遗传资源,进行抗苯磺隆油菜育种潜力有限。 初步研究了乙酰乳酸合成酶基因与油菜对苯磺隆抗性之间的关系。结果表明,不同甘蓝型油菜品种的ALS1、ALS3基因扩增特性具有一定保守性, ALS1、ALS3基因结构与油菜对苯磺隆耐药性强弱的关系有待进一步研究。 苯磺隆对油菜乙酰乳酸合成酶(ALS)和谷胱甘肽转移酶(GSTs)活性变化有影响:苯磺隆溶液处理后,3个苯磺隆抗性不同的油菜品种叶片ALS活体活性均受到抑制,GSTs活性先下降然后回升。低度敏感品种17号相对ALS活性高于其他2个品种,并且GSTs活性下降较快,回升后峰值也高于其他2个品种。不同品种油菜GSTs对苯磺隆代谢能力差异可能是对苯磺隆敏感性不同的原因之一。 以C5(C9)为抗除草剂草胺膦(草甘膦)基因bar(gox)的供体亲本,采用回交育种策略,试图将上述2个抗除草剂性状转移到优良油菜品系2011Gr-1和2011Gr-2中。在回交育种中,利用目的基因PCR检测和喷施除草剂相结合的方法,进行抗性基因的“前景”选择,同时利用分子标记对回交后代进行“背景”选择,获得了抗草胺膦(草甘膦)基因的回交3代油菜,初步选育出除草剂抗性稳定、农艺性状优良的油菜品系。 上述试验结果为抗除草剂油菜品种的选育,利用抗除草剂品种防除油菜田间杂草奠定了初步基础。
URL
随着我国国民经济的快速发展,农村青壮年劳动力大量向城市转移。轻简化、机械化已成为油菜产业发展的主要方向,杂草的综合有效治理是实现油菜机械化、轻简化的重要技术环节。培育和推广抗除草剂的油菜品种是防除田间草害,提高油菜产量和质量最为经济有效的措施。 本研究首先以中双9号油菜为试材,探索了适宜油菜筛选的苯磺隆药液浓度和鉴定指标。结果表明,叶片药害指数、叶片数减少率、叶夹角减少率与植株死亡率之间都存在显著或极显著的相关关系,说明这3个指标可考虑作为早期鉴定油菜对苯磺隆耐药性的指标。用筛选出的适宜浓度苯磺隆溶液(2.0μg mL-1),处理了49个不同基因型油菜的幼苗,根据基于叶片药害指数、叶片数减少率、叶夹角减少率、植株死亡率4个指标计算出的耐苯磺隆平均隶属函数值(AS)将参试材料分为3个等级,低度敏感材料2份(AS 0.33)、中度敏感材料22份(0.33≤AS 0.67)、高度敏感材料25份(AS≥0.67)。虽然参试材料对苯磺隆抗性存在一定的差异,然而与小麦田推荐的苯磺隆使用浓度相比,所有参试油菜材料对苯磺隆抗性水平还很低,说明利用油菜种内遗传资源,进行抗苯磺隆油菜育种潜力有限。 初步研究了乙酰乳酸合成酶基因与油菜对苯磺隆抗性之间的关系。结果表明,不同甘蓝型油菜品种的ALS1、ALS3基因扩增特性具有一定保守性, ALS1、ALS3基因结构与油菜对苯磺隆耐药性强弱的关系有待进一步研究。 苯磺隆对油菜乙酰乳酸合成酶(ALS)和谷胱甘肽转移酶(GSTs)活性变化有影响:苯磺隆溶液处理后,3个苯磺隆抗性不同的油菜品种叶片ALS活体活性均受到抑制,GSTs活性先下降然后回升。低度敏感品种17号相对ALS活性高于其他2个品种,并且GSTs活性下降较快,回升后峰值也高于其他2个品种。不同品种油菜GSTs对苯磺隆代谢能力差异可能是对苯磺隆敏感性不同的原因之一。 以C5(C9)为抗除草剂草胺膦(草甘膦)基因bar(gox)的供体亲本,采用回交育种策略,试图将上述2个抗除草剂性状转移到优良油菜品系2011Gr-1和2011Gr-2中。在回交育种中,利用目的基因PCR检测和喷施除草剂相结合的方法,进行抗性基因的“前景”选择,同时利用分子标记对回交后代进行“背景”选择,获得了抗草胺膦(草甘膦)基因的回交3代油菜,初步选育出除草剂抗性稳定、农艺性状优良的油菜品系。 上述试验结果为抗除草剂油菜品种的选育,利用抗除草剂品种防除油菜田间杂草奠定了初步基础。
DOI:10.1002/(ISSN)1526-4998URL
[本文引用: 2]
[本文引用: 2]
DOI:10.1017/S0890037X00025252URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1221179110URLPMID:23530204 [本文引用: 1]
Abstract Multiple-herbicide resistance (MHR) in black-grass (Alopecurus myosuroides) and annual rye-grass (Lolium rigidum) is a global problem leading to a loss of chemical weed control in cereal crops. Although poorly understood, in common with multiple-drug resistance (MDR) in tumors, MHR is associated with an enhanced ability to detoxify xenobiotics. In humans, MDR is linked to the overexpression of a pi class glutathione transferase (GSTP1), which has both detoxification and signaling functions in promoting drug resistance. In both annual rye-grass and black-grass, MHR was also associated with the increased expression of an evolutionarily distinct plant phi (F) GSTF1 that had a restricted ability to detoxify herbicides. When the black-grass A. myosuroides (Am) AmGSTF1 was expressed in Arabidopsis thaliana, the transgenic plants acquired resistance to multiple herbicides and showed similar changes in their secondary, xenobiotic, and antioxidant metabolism to those determined in MHR weeds. Transcriptome array experiments showed that these changes in biochemistry were not due to changes in gene expression. Rather, AmGSTF1 exerted a direct regulatory control on metabolism that led to an accumulation of protective flavonoids. Further evidence for a key role for this protein in MHR was obtained by showing that the GSTP1- and MDR-inhibiting pharmacophore 4-chloro-7-nitro-benzoxadiazole was also active toward AmGSTF1 and helped restore herbicide control in MHR black-grass. These studies demonstrate a central role for specific GSTFs in MHR in weeds that has parallels with similar roles for unrelated GSTs in MDR in humans and shows their potential as targets for chemical intervention in resistant weed management.
URL [本文引用: 2]
大豆(Glycine max(L.) Merr)起源于中国,是重要的植物蛋白及食用油的主要来源。因我国大豆产量偏低、农村劳动力转移和草害严重等因素导致大豆种植面积逐年下降、国内大豆综合竞争力不足等严重问题,国内大豆供不应求的市场现状迫使我国大量进口大豆以满足市场需求。因此,培育抗草甘膦大豆新品种对提高农田杂草防治效率、降低防除成本,推动免耕少耕等新型耕作方式的推广及促进我国大豆生产自给自足等均有重要意义。尽管国际转基因大豆早已商业化,但转基因大豆的食品安全问题还有待进一步的验证。 草甘膦是是美国孟山都公司开发的非选择性灭生除草...
URL [本文引用: 2]
大豆(Glycine max(L.) Merr)起源于中国,是重要的植物蛋白及食用油的主要来源。因我国大豆产量偏低、农村劳动力转移和草害严重等因素导致大豆种植面积逐年下降、国内大豆综合竞争力不足等严重问题,国内大豆供不应求的市场现状迫使我国大量进口大豆以满足市场需求。因此,培育抗草甘膦大豆新品种对提高农田杂草防治效率、降低防除成本,推动免耕少耕等新型耕作方式的推广及促进我国大豆生产自给自足等均有重要意义。尽管国际转基因大豆早已商业化,但转基因大豆的食品安全问题还有待进一步的验证。 草甘膦是是美国孟山都公司开发的非选择性灭生除草...
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7666/d.y2050365URL [本文引用: 1]
麦田草害成为影响和制约小麦稳产高产的限制性因子之一,全世界小 麦因草害引起产量损失达总产的10%左右。如何合理的使用麦田除草剂来提高小麦产量与品质及资源利用效率是我国农业发展面临的重要课题之一。本研究以麦田 常用的5种除草剂苯磺隆、使它隆、异丙隆、骠马、绿麦隆为研究对象,以弱筋小麦扬麦13为供试小麦品种,于2008-2010年在扬州大学江苏省作物遗传 生理重点实验室试验场进行,在评价麦田除草剂不同施用剂量(0.5倍推荐剂量,推荐剂量,2倍推荐剂量,4倍推荐剂量)、不同施药时期(冬季施药,春季施 药)对麦田杂草防效及小麦安全生产技术的基础上,重点...
DOI:10.7666/d.y2050365URL [本文引用: 1]
麦田草害成为影响和制约小麦稳产高产的限制性因子之一,全世界小 麦因草害引起产量损失达总产的10%左右。如何合理的使用麦田除草剂来提高小麦产量与品质及资源利用效率是我国农业发展面临的重要课题之一。本研究以麦田 常用的5种除草剂苯磺隆、使它隆、异丙隆、骠马、绿麦隆为研究对象,以弱筋小麦扬麦13为供试小麦品种,于2008-2010年在扬州大学江苏省作物遗传 生理重点实验室试验场进行,在评价麦田除草剂不同施用剂量(0.5倍推荐剂量,推荐剂量,2倍推荐剂量,4倍推荐剂量)、不同施药时期(冬季施药,春季施 药)对麦田杂草防效及小麦安全生产技术的基础上,重点...
DOI:10.7666/d.y1597372URL [本文引用: 1]
本文研究建立了小麦植株、籽粒和土壤中苯磺隆与氯氟吡氧乙酸的残留分析方法,评价了方法的准确度、灵敏度和精密度,进行了苯磺隆与氯氟吡氧乙酸在小麦植株和土壤中残留田间消解动态和最终残留试验,为该两种除草剂在小麦地的合理使用提供了科学依据。主要研究结果如下: 1.小麦植株、籽粒和土壤中苯磺隆残留分析方法的建立 土壤样品以磷酸盐缓冲液-甲醇混合提取,机械振荡提取2次,合并滤液,以SPE柱净化;小麦植株样品以甲醇为提取液,经石油醚液-液分配后再采用SPE柱净化;小麦籽粒样品采用二氯甲烷提取后直接进样分析。采用配有紫外检测器的高效液相色谱仪检测。 检测条件:Agilent 1200高效液相色谱仪,具紫外检测器;色谱柱:Agilent ZORBAX-ODS(250mm×4.6mm)色谱柱;检测波长:240nm;流动相:乙腈/0.5%冰乙酸(45/55,v/v);流速:1.0mL/min;柱温:30℃;进样量:20μL。该检测条件下,苯磺隆的保留时间为10.4min,仪器对苯磺隆的最小检出量为1.0×10-9 g。 提取体系:比较了机械振荡法和超声波振荡法不同提取时间的提取效率,确定了机械振荡30 min + 15 min为苯磺隆优化后的提取方法;比较了二氯甲烷、甲醇﹑乙腈、0.2 mol/L磷酸盐缓冲液-甲醇(8:2,v/v)等4种提取溶剂对苯磺隆的提取效率,确定了甲醇、二氯甲烷和0.2mol/L磷酸盐缓冲液-甲醇(8:2,v/v)分别为苯磺隆在小麦植株、籽粒和土壤中的提取溶剂。 净化方法:采用SPE小柱进行样品的净化处理,比较了土壤样品SPE小柱淋洗液的种类以及配比对净化效果和添加回收率的影响,最终确定先用4mL乙腈pH6.0的0.2mol/L磷酸缓冲液3/7(v/v,85%磷酸调节pH值至6.0左右)将杂质洗脱,再用3mL乙腈- pH7.8的0.2mol/L磷酸缓冲液(1/1,v/v)淋洗,收集淋出液检测。植株样品在基础上,先分别用石油醚和二氯甲烷液液分配萃取后,再经SPE小柱净化。 优化后方法的添加回收试验结果表明:在0.01mg/kg~0.80mg/kg添加浓度范围内,小麦植株中苯磺隆的平均回收率为73.1~85.3%,变异系数为5.88~8.53%;土壤中苯磺隆平均回收率为80.1~95.2 % ,变异系数为3.41~7.90% ;在0.025mg/kg~0.80mg/kg添加浓度范围内,籽粒中苯磺隆的平均回收率为73.3~87.5%,变异系数为3.21~6.49%。该残留分析方法的准确性、精确性均达到农药残留分析的要求。 2.小麦植株、籽粒和土壤中氯氟吡氧乙酸残留分析方法的建立 样品以碱性甲醇混合提取液机械振荡提取后,液液分配净化,采用浓硫酸做为催化剂,甲醇做为衍生化试剂,反应后经石油醚提取,GC-ECD法检测。 检测条件的确立:Agilent 6890气相色谱仪具ECD检测器;色谱柱:HP-5毛细管柱(30.0m×250um×0.25um);检测温度:柱温起始温度,70℃,保持1min,以20℃/min至240℃,保持6min;进样口温度250℃,检测器温度300℃;载气:高纯氮气(99.999%),载气流速为1mL/min;进样方式:不分流方式;进样量为2uL。在此条件下氯氟吡氧乙酸的保留时间为10.5 min左右,仪器对氯氟吡氧乙酸的最小检出量为1.0×10-11 g。 提取体系:比较了机械振荡法和超声波振荡法两种提取方式不同提取时间的提取效率,确定了机械振荡30min为氯氟吡氧乙酸优化后的提取方法;比较了乙腈、乙酸乙酯、碱性甲醇等3种提取溶剂对氯氟吡氧乙酸提取效率,确定碱性甲醇为氯氟吡氧乙酸在小麦植株、籽粒、土壤中的提取溶剂。 衍生化方法:比较了不同甲醇用量、酯化时间和酯化温度等因素对衍生化结果的影响,结果表明,甲醇用量为2 mL,浓H2SO4 1.5 mL,93~98℃水浴条件下酯化时间10 min,较好。 优化后方法的添加回收试验结果表明:在0.01mg/kg~0.80mg/kg的添加浓度范围内,小麦植株中氯氟吡氧乙酸的平均回收率为72.3~86.7%,变异系数为3.02~8.59%;籽粒中氯氟吡氧乙酸的平均回收率为77.7~87.3%,变异系数为2.75~7.61%;土壤中的氯氟吡氧乙酸平均回收率为83.6~95.8%,变异系数为2.87~8.46%。该残留分析方法的准确性、精确性均达到农药残留分析的要求。 3.小麦植株和土壤中苯磺隆残留消解动态 2008年在安徽、山东两地的田间残留试验结果表明,苯磺隆的消解动态符合一级反应动力学方程。在合肥试验点,小麦植株上苯磺隆的田间消解动态方程为C = 0.0593e-0.1393t,半衰期为4.97d;土壤中苯磺隆消解动态方程为C= 0.0434e-0.1250t,半衰期为5.54d。在青岛试验点,小麦植株上苯磺隆的田间消解动态方程为C = 0.0826e-0.1659t,半衰期为4.18d;土壤中苯磺隆消解动态方程为C = 0.0572e-0.1306t,半衰期为5.31d。在合肥和青岛两地最终残留试验的小麦籽粒和土壤样品中均未有苯磺隆检出。 4.小麦植株和土壤中氯氟吡氧乙酸残留消解动态 2008年在安徽、山东两地的田间残留试验结果表明,氯氟吡氧乙酸的消解动态符合一级反应动力学方程。在合肥试验点,小麦植株上氯氟吡氧乙酸田间消解动态方程为C= 0.1226e-0.1171t,半衰期为5.92d;土壤中氯氟吡氧乙酸田间消解动态方程为C = 0.0861e-0.0828t,半衰期为8.37d。在青岛试验点,小麦植株上氯氟吡氧乙酸田间消解动态方程为C= 0.2149e-0.1368t,半衰期为5.07 d;土壤中氯氟吡氧乙酸田间消解动态方程为C = 0.1478e-0.0893t,半衰期为7.76d。在合肥和青岛两地最终残留试验的小麦籽粒和土壤样品中均未有氯氟吡氧乙酸检出。
DOI:10.7666/d.y1597372URL [本文引用: 1]
本文研究建立了小麦植株、籽粒和土壤中苯磺隆与氯氟吡氧乙酸的残留分析方法,评价了方法的准确度、灵敏度和精密度,进行了苯磺隆与氯氟吡氧乙酸在小麦植株和土壤中残留田间消解动态和最终残留试验,为该两种除草剂在小麦地的合理使用提供了科学依据。主要研究结果如下: 1.小麦植株、籽粒和土壤中苯磺隆残留分析方法的建立 土壤样品以磷酸盐缓冲液-甲醇混合提取,机械振荡提取2次,合并滤液,以SPE柱净化;小麦植株样品以甲醇为提取液,经石油醚液-液分配后再采用SPE柱净化;小麦籽粒样品采用二氯甲烷提取后直接进样分析。采用配有紫外检测器的高效液相色谱仪检测。 检测条件:Agilent 1200高效液相色谱仪,具紫外检测器;色谱柱:Agilent ZORBAX-ODS(250mm×4.6mm)色谱柱;检测波长:240nm;流动相:乙腈/0.5%冰乙酸(45/55,v/v);流速:1.0mL/min;柱温:30℃;进样量:20μL。该检测条件下,苯磺隆的保留时间为10.4min,仪器对苯磺隆的最小检出量为1.0×10-9 g。 提取体系:比较了机械振荡法和超声波振荡法不同提取时间的提取效率,确定了机械振荡30 min + 15 min为苯磺隆优化后的提取方法;比较了二氯甲烷、甲醇﹑乙腈、0.2 mol/L磷酸盐缓冲液-甲醇(8:2,v/v)等4种提取溶剂对苯磺隆的提取效率,确定了甲醇、二氯甲烷和0.2mol/L磷酸盐缓冲液-甲醇(8:2,v/v)分别为苯磺隆在小麦植株、籽粒和土壤中的提取溶剂。 净化方法:采用SPE小柱进行样品的净化处理,比较了土壤样品SPE小柱淋洗液的种类以及配比对净化效果和添加回收率的影响,最终确定先用4mL乙腈pH6.0的0.2mol/L磷酸缓冲液3/7(v/v,85%磷酸调节pH值至6.0左右)将杂质洗脱,再用3mL乙腈- pH7.8的0.2mol/L磷酸缓冲液(1/1,v/v)淋洗,收集淋出液检测。植株样品在基础上,先分别用石油醚和二氯甲烷液液分配萃取后,再经SPE小柱净化。 优化后方法的添加回收试验结果表明:在0.01mg/kg~0.80mg/kg添加浓度范围内,小麦植株中苯磺隆的平均回收率为73.1~85.3%,变异系数为5.88~8.53%;土壤中苯磺隆平均回收率为80.1~95.2 % ,变异系数为3.41~7.90% ;在0.025mg/kg~0.80mg/kg添加浓度范围内,籽粒中苯磺隆的平均回收率为73.3~87.5%,变异系数为3.21~6.49%。该残留分析方法的准确性、精确性均达到农药残留分析的要求。 2.小麦植株、籽粒和土壤中氯氟吡氧乙酸残留分析方法的建立 样品以碱性甲醇混合提取液机械振荡提取后,液液分配净化,采用浓硫酸做为催化剂,甲醇做为衍生化试剂,反应后经石油醚提取,GC-ECD法检测。 检测条件的确立:Agilent 6890气相色谱仪具ECD检测器;色谱柱:HP-5毛细管柱(30.0m×250um×0.25um);检测温度:柱温起始温度,70℃,保持1min,以20℃/min至240℃,保持6min;进样口温度250℃,检测器温度300℃;载气:高纯氮气(99.999%),载气流速为1mL/min;进样方式:不分流方式;进样量为2uL。在此条件下氯氟吡氧乙酸的保留时间为10.5 min左右,仪器对氯氟吡氧乙酸的最小检出量为1.0×10-11 g。 提取体系:比较了机械振荡法和超声波振荡法两种提取方式不同提取时间的提取效率,确定了机械振荡30min为氯氟吡氧乙酸优化后的提取方法;比较了乙腈、乙酸乙酯、碱性甲醇等3种提取溶剂对氯氟吡氧乙酸提取效率,确定碱性甲醇为氯氟吡氧乙酸在小麦植株、籽粒、土壤中的提取溶剂。 衍生化方法:比较了不同甲醇用量、酯化时间和酯化温度等因素对衍生化结果的影响,结果表明,甲醇用量为2 mL,浓H2SO4 1.5 mL,93~98℃水浴条件下酯化时间10 min,较好。 优化后方法的添加回收试验结果表明:在0.01mg/kg~0.80mg/kg的添加浓度范围内,小麦植株中氯氟吡氧乙酸的平均回收率为72.3~86.7%,变异系数为3.02~8.59%;籽粒中氯氟吡氧乙酸的平均回收率为77.7~87.3%,变异系数为2.75~7.61%;土壤中的氯氟吡氧乙酸平均回收率为83.6~95.8%,变异系数为2.87~8.46%。该残留分析方法的准确性、精确性均达到农药残留分析的要求。 3.小麦植株和土壤中苯磺隆残留消解动态 2008年在安徽、山东两地的田间残留试验结果表明,苯磺隆的消解动态符合一级反应动力学方程。在合肥试验点,小麦植株上苯磺隆的田间消解动态方程为C = 0.0593e-0.1393t,半衰期为4.97d;土壤中苯磺隆消解动态方程为C= 0.0434e-0.1250t,半衰期为5.54d。在青岛试验点,小麦植株上苯磺隆的田间消解动态方程为C = 0.0826e-0.1659t,半衰期为4.18d;土壤中苯磺隆消解动态方程为C = 0.0572e-0.1306t,半衰期为5.31d。在合肥和青岛两地最终残留试验的小麦籽粒和土壤样品中均未有苯磺隆检出。 4.小麦植株和土壤中氯氟吡氧乙酸残留消解动态 2008年在安徽、山东两地的田间残留试验结果表明,氯氟吡氧乙酸的消解动态符合一级反应动力学方程。在合肥试验点,小麦植株上氯氟吡氧乙酸田间消解动态方程为C= 0.1226e-0.1171t,半衰期为5.92d;土壤中氯氟吡氧乙酸田间消解动态方程为C = 0.0861e-0.0828t,半衰期为8.37d。在青岛试验点,小麦植株上氯氟吡氧乙酸田间消解动态方程为C= 0.2149e-0.1368t,半衰期为5.07 d;土壤中氯氟吡氧乙酸田间消解动态方程为C = 0.1478e-0.0893t,半衰期为7.76d。在合肥和青岛两地最终残留试验的小麦籽粒和土壤样品中均未有氯氟吡氧乙酸检出。
[本文引用: 2]
[本文引用: 2]
DOI:10.13802/j.cnki.zwbhxb.2017.2015145URL [本文引用: 1]
为揭示油菜和小麦根系对乙草胺耐药性差异的原因,采用水培法研究了梯度浓度乙草胺对油菜和小麦种苗根系形态、根尖生理代谢和解剖结构的影响。结果表明,1 mg/L乙草胺对油菜根长抑制率为33.63%,而对小麦根长抑制率可达55.22%;100 mg/L乙草胺对油菜侧根抑制率为63.03%,而对小麦侧根抑制率达100.00%;经0.01 mg/L乙草胺处理后的油菜根尖细胞膜透性高于小麦,当乙草胺浓度高于0.1 mg/L后,小麦根尖细胞膜透性剧烈增加且高于油菜;在较高浓度乙草胺胁迫下,小麦根尖抗氧化酶活性均低于油菜;10 mg/L乙草胺处理下,小麦根尖的解剖结构变异较油菜明显,表现为细胞排列松散、混乱,根冠变形,分生组织细胞染色程度变浅,伸长区细胞分化提前,中柱鞘细胞木质化。研究表明,小麦的根系建成比油菜更容易受乙草胺抑制,且侧根数比根长更敏感;油菜和小麦对乙草胺耐药性差异可能与细胞膜透性、抗氧化酶活性以及根尖细胞组织分化等差异有关。
DOI:10.13802/j.cnki.zwbhxb.2017.2015145URL [本文引用: 1]
为揭示油菜和小麦根系对乙草胺耐药性差异的原因,采用水培法研究了梯度浓度乙草胺对油菜和小麦种苗根系形态、根尖生理代谢和解剖结构的影响。结果表明,1 mg/L乙草胺对油菜根长抑制率为33.63%,而对小麦根长抑制率可达55.22%;100 mg/L乙草胺对油菜侧根抑制率为63.03%,而对小麦侧根抑制率达100.00%;经0.01 mg/L乙草胺处理后的油菜根尖细胞膜透性高于小麦,当乙草胺浓度高于0.1 mg/L后,小麦根尖细胞膜透性剧烈增加且高于油菜;在较高浓度乙草胺胁迫下,小麦根尖抗氧化酶活性均低于油菜;10 mg/L乙草胺处理下,小麦根尖的解剖结构变异较油菜明显,表现为细胞排列松散、混乱,根冠变形,分生组织细胞染色程度变浅,伸长区细胞分化提前,中柱鞘细胞木质化。研究表明,小麦的根系建成比油菜更容易受乙草胺抑制,且侧根数比根长更敏感;油菜和小麦对乙草胺耐药性差异可能与细胞膜透性、抗氧化酶活性以及根尖细胞组织分化等差异有关。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
采用灰色关联度分析法对9个绿 豆品种的7个农艺性状进行综合评价,结果表明:吉绿6号、辽绿6号、大鹦哥绿935综合表现最好,等权关联度与加权关联度的分析结果基本一致。不同农艺性 状对小区产量的关联度不同,6个农艺性状与小区产量的关联度排序为:单株粒重百粒重单株荚数单荚粒数荚长株高。
URL [本文引用: 2]
采用灰色关联度分析法对9个绿 豆品种的7个农艺性状进行综合评价,结果表明:吉绿6号、辽绿6号、大鹦哥绿935综合表现最好,等权关联度与加权关联度的分析结果基本一致。不同农艺性 状对小区产量的关联度不同,6个农艺性状与小区产量的关联度排序为:单株粒重百粒重单株荚数单荚粒数荚长株高。
DOI:10.2135/cropsci2004.0389URL [本文引用: 2]
DOI:10.13430/j.cnki.jpgr.2017.05.006URL [本文引用: 2]
干旱是影响薏苡生产的主要因素之一,鉴定薏苡种质资源的抗旱性,筛选抗旱指标,培育抗旱品种,对薏苡产业的发展具有重要意义。本研究首先以抗旱性不同的6份薏苡种质为材料,通过调查5个不同浓度的PEG-6000水溶液模拟干旱胁迫下的发芽率,确定了薏苡萌发期模拟干旱胁迫的最适PEG-6000水溶液渗透势为-0.1 MPa。然后以-0.1 MPa的PEG-6000水溶液模拟干旱胁迫,探讨50份薏苡种质发芽势、发芽率、萌发指数、芽长、芽粗、芽鲜重、芽干重、根长、根粗、根鲜重和根干重的变化,利用综合评价法对50份薏苡种质进行萌发期抗旱性鉴定及抗旱指标筛选。结果表明,干旱胁迫对薏苡萌发期各指标均有极显著影响。筛选出萌发期抗旱性强的薏苡种质分别为yy18-1、yy14-3和yy13-1,可为薏苡抗旱育种、抗旱机理及干旱调控缓解机制的研究提供基础材料。芽长、芽干重、根长、根鲜重和根干重可作为薏苡种质资源萌发期简单、直观的抗旱性评价指标。
DOI:10.13430/j.cnki.jpgr.2017.05.006URL [本文引用: 2]
干旱是影响薏苡生产的主要因素之一,鉴定薏苡种质资源的抗旱性,筛选抗旱指标,培育抗旱品种,对薏苡产业的发展具有重要意义。本研究首先以抗旱性不同的6份薏苡种质为材料,通过调查5个不同浓度的PEG-6000水溶液模拟干旱胁迫下的发芽率,确定了薏苡萌发期模拟干旱胁迫的最适PEG-6000水溶液渗透势为-0.1 MPa。然后以-0.1 MPa的PEG-6000水溶液模拟干旱胁迫,探讨50份薏苡种质发芽势、发芽率、萌发指数、芽长、芽粗、芽鲜重、芽干重、根长、根粗、根鲜重和根干重的变化,利用综合评价法对50份薏苡种质进行萌发期抗旱性鉴定及抗旱指标筛选。结果表明,干旱胁迫对薏苡萌发期各指标均有极显著影响。筛选出萌发期抗旱性强的薏苡种质分别为yy18-1、yy14-3和yy13-1,可为薏苡抗旱育种、抗旱机理及干旱调控缓解机制的研究提供基础材料。芽长、芽干重、根长、根鲜重和根干重可作为薏苡种质资源萌发期简单、直观的抗旱性评价指标。
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1002-2481.2008.02.011URL [本文引用: 2]
通过敏感性测定,明确了不同小麦品种对麦田磺酰脲类除草剂苯磺隆的耐药性.结果表明:河东TX耐药性较强;晋麦67、晋农66、晋麦47、长5456、农大3608耐药性居中;长006219、乐39耐药性较弱.长006219对苯磺隆药剂的敏感性最强,用药不当,易产生药害.对品种耐药性以及药剂抑制力的进行了综合分析.
DOI:10.3969/j.issn.1002-2481.2008.02.011URL [本文引用: 2]
通过敏感性测定,明确了不同小麦品种对麦田磺酰脲类除草剂苯磺隆的耐药性.结果表明:河东TX耐药性较强;晋麦67、晋农66、晋麦47、长5456、农大3608耐药性居中;长006219、乐39耐药性较弱.长006219对苯磺隆药剂的敏感性最强,用药不当,易产生药害.对品种耐药性以及药剂抑制力的进行了综合分析.
DOI:10.3969/j.issn.1000-6850.2005.10.089URLMagsci [本文引用: 1]
室内土壤滴浇法测定了苯磺隆、苄嘧磺隆对不同小麦品种的安全性和叶绿素含量的变化,结果表明:两种药剂对不同小麦品种的安全性存在较大差异,且对根长的抑制作用明显大于株高;对叶绿素含量的影响与对株高的影响结果一致。苯磺隆对周麦13的根和芽敏感,使用不当易产生药害;对偃师4110根长最安全,株高最敏感。苄嘧磺隆对高优503根长最安全,株高最敏感;对周麦13株高最安全,根长最敏感。因此,小麦田化学除草应根据不同小麦品种来选择除草剂品种、有效用量和最佳施药方法。
DOI:10.3969/j.issn.1000-6850.2005.10.089URLMagsci [本文引用: 1]
室内土壤滴浇法测定了苯磺隆、苄嘧磺隆对不同小麦品种的安全性和叶绿素含量的变化,结果表明:两种药剂对不同小麦品种的安全性存在较大差异,且对根长的抑制作用明显大于株高;对叶绿素含量的影响与对株高的影响结果一致。苯磺隆对周麦13的根和芽敏感,使用不当易产生药害;对偃师4110根长最安全,株高最敏感。苄嘧磺隆对高优503根长最安全,株高最敏感;对周麦13株高最安全,根长最敏感。因此,小麦田化学除草应根据不同小麦品种来选择除草剂品种、有效用量和最佳施药方法。
DOI:10.13430/j.cnki.jpgr.2015.05.003URL
以萌发初期大豆弯曲的子叶节为靶点,利用草甘膦(Roundup)点施鉴定法,分析了221份大豆微核心种质的草甘膦耐受性及其与抗草甘膦转CP4-EPSPS基因大豆AG5601的差异,结果表明,草甘膦对不同大豆种质的抑制程度与点施的草甘膦浓度呈正相关,Roundup稀释浓度为1/1000、1/10000时,草甘膦对大豆生长几乎无影响;Roundup稀释浓度为1/10时,草甘膦显著抑制大豆生长,导致植株死亡;Roundup稀释浓度为1/100时,不同大豆种质对草甘膦耐受性差异显著,并鉴定出对草甘膦具有较好耐受性的种质10份。虽然大豆微核心种质对草甘膦的耐受程度远远低于AG5601,但不同大豆种质对草甘膦耐受性存在显著差异,这为利用转基因和杂交转育技术培育抗草甘膦转基因大豆的受体或轮回亲本的选择提供了理论依据。
DOI:10.13430/j.cnki.jpgr.2015.05.003URL
以萌发初期大豆弯曲的子叶节为靶点,利用草甘膦(Roundup)点施鉴定法,分析了221份大豆微核心种质的草甘膦耐受性及其与抗草甘膦转CP4-EPSPS基因大豆AG5601的差异,结果表明,草甘膦对不同大豆种质的抑制程度与点施的草甘膦浓度呈正相关,Roundup稀释浓度为1/1000、1/10000时,草甘膦对大豆生长几乎无影响;Roundup稀释浓度为1/10时,草甘膦显著抑制大豆生长,导致植株死亡;Roundup稀释浓度为1/100时,不同大豆种质对草甘膦耐受性差异显著,并鉴定出对草甘膦具有较好耐受性的种质10份。虽然大豆微核心种质对草甘膦的耐受程度远远低于AG5601,但不同大豆种质对草甘膦耐受性存在显著差异,这为利用转基因和杂交转育技术培育抗草甘膦转基因大豆的受体或轮回亲本的选择提供了理论依据。
DOI:10.7666/d.Y2787485URL [本文引用: 1]
自1940年第一例除草剂2,4-D问世至今,不断出现各种作用机制的除草剂,目前种类繁多。1974年商业化的草甘膦具有广谱、灭生性,杀草能力强,副作用小,在除草剂中独树一帜,且在全世界范围内被广泛使用。发展转基因抗草甘膦作物很大程度上方便了草甘膦除草,但存在转基因安全隐患。因此,在植棉过程中,从现有棉花资源挖掘内源抗草甘膦基因源,并用于棉花育种,可有效解决诸多问题。棉花种质资源蕴含许多优异基因,鉴于此,本文旨在对野生棉、半野生棉和亚洲棉种质材料草甘膦自然抗性进行鉴定和评价,分析半野生棉极端反应材料的epsps基因序列及其表达量,阐明抗性与epsps基因的关系,并通过全基因组关联分析方法,从数量性状思路挖掘棉花中抗草甘膦的基因源。研究结果如下:1.棉花种质材料草甘膦自然抗性评价及比较分析分别采用嫩枝喷洒法和棉花叶片棉球定位法,对26个野生棉种共134株和7个半野生棉种系共403份,及收集自5个地区的亚洲棉共101份材料,进行了草甘膦自然抗性的鉴定和评价,并对其抗性水平进行了比较分析。通过田间鉴定试验,获得了一个棉花除草剂自然抗性鉴定的方法,拟定了相应抗性水平的评价参考标准,并分别筛选出1份、2份和5份高抗的野生棉、半野生棉和亚洲棉材料,16份和1份高感的野生棉和半野生棉材料。分别以苗期、蕾期和花铃期常规栽培棉中棉所12的草甘膦致死浓度0.1%、0.15%和0.25%为标准,采用喷洒法对高抗和高感半野生棉材料进行田间抗性鉴定,最终得到稳定的抗和敏感材料各1份。同时,比较分析发现鉴定材料抗性水平多样性依次是:野生棉半野生棉亚洲棉;鉴定材料的整体抗性水平依次是:亚洲棉半野生棉野生棉。对棉花种质资源的草甘膦抗性进行评价,有助于全面系统地了解棉花的草甘膦抗性,为研究棉花的草甘膦抗性机制及内源抗性基因源的挖掘奠定材料基础,同时也指导抗草甘膦棉花育种。2.半野生棉内源epsps基因序列及表达量分析以sad1基因为参考基因,采用孟山都公司转基因抗草甘膦棉花的基因序列特异引物,对半野生棉极端材料进行转基因成分检测,证明筛选出的极端材料不含转抗草甘膦基因成分。采用陆地棉(珂字棉312)中epsps基因全长引物对半野生棉材料DNA进行扩增,扩增产物经测序并翻译成氨基酸序列,比对分析发现半野生棉epsps基因没有特异的氨基酸突变位点,说明半野生棉材料抗性的产生不是由epsps基因突变而致。对材料喷施0.2%的草甘膦溶液,48 h后取样,采用RT-PCR方法检测epsps基因表达量变化,发现草甘膦处理后,epsps基因表达量无明显变化,这表明半野生棉草甘膦抗性可能不是由epsps基因引起,而是其它抗性机制而致,有待以新的思路去研究。3.关联分析方法挖掘半野生棉草甘膦抗性基因源2013年,在三个不同环境下,对202份半野生棉材料进行与草甘膦抗性相关表型的测定,包括莽草酸含量和地上部分相对干重,并与覆盖全基因组的182对SSR多态性标记的基因型数据进行全基因组关联分析,共检测到72个与草甘膦抗性相关的位点,其中,48个和41个位点分别与莽草酸含量和地上部分相对干重显著相关。基于成功关联的SSR引物序列和中国农业科学院棉花研究所公布的雷蒙德氏棉(G.raimondii)、亚洲棉(G.arboreum L)和陆地棉(G.hirsutum)全基因组测序数据,共得到87条候选基因,包括亚洲棉中的21条,雷蒙德氏棉中的24条和陆地棉中的42条。以可可(Theobroma cacao)为模式植物,将这些基因在KEGG数据库中进行注释,探索相关的代谢通路,其中,ABC转运蛋白家族可能与草甘膦抗性相关,且已被前人证明,另外注释结果也推测组氨酸代谢、氨基酸生物合成和次生代谢都与草甘膦抗性有关。
DOI:10.7666/d.Y2787485URL [本文引用: 1]
自1940年第一例除草剂2,4-D问世至今,不断出现各种作用机制的除草剂,目前种类繁多。1974年商业化的草甘膦具有广谱、灭生性,杀草能力强,副作用小,在除草剂中独树一帜,且在全世界范围内被广泛使用。发展转基因抗草甘膦作物很大程度上方便了草甘膦除草,但存在转基因安全隐患。因此,在植棉过程中,从现有棉花资源挖掘内源抗草甘膦基因源,并用于棉花育种,可有效解决诸多问题。棉花种质资源蕴含许多优异基因,鉴于此,本文旨在对野生棉、半野生棉和亚洲棉种质材料草甘膦自然抗性进行鉴定和评价,分析半野生棉极端反应材料的epsps基因序列及其表达量,阐明抗性与epsps基因的关系,并通过全基因组关联分析方法,从数量性状思路挖掘棉花中抗草甘膦的基因源。研究结果如下:1.棉花种质材料草甘膦自然抗性评价及比较分析分别采用嫩枝喷洒法和棉花叶片棉球定位法,对26个野生棉种共134株和7个半野生棉种系共403份,及收集自5个地区的亚洲棉共101份材料,进行了草甘膦自然抗性的鉴定和评价,并对其抗性水平进行了比较分析。通过田间鉴定试验,获得了一个棉花除草剂自然抗性鉴定的方法,拟定了相应抗性水平的评价参考标准,并分别筛选出1份、2份和5份高抗的野生棉、半野生棉和亚洲棉材料,16份和1份高感的野生棉和半野生棉材料。分别以苗期、蕾期和花铃期常规栽培棉中棉所12的草甘膦致死浓度0.1%、0.15%和0.25%为标准,采用喷洒法对高抗和高感半野生棉材料进行田间抗性鉴定,最终得到稳定的抗和敏感材料各1份。同时,比较分析发现鉴定材料抗性水平多样性依次是:野生棉半野生棉亚洲棉;鉴定材料的整体抗性水平依次是:亚洲棉半野生棉野生棉。对棉花种质资源的草甘膦抗性进行评价,有助于全面系统地了解棉花的草甘膦抗性,为研究棉花的草甘膦抗性机制及内源抗性基因源的挖掘奠定材料基础,同时也指导抗草甘膦棉花育种。2.半野生棉内源epsps基因序列及表达量分析以sad1基因为参考基因,采用孟山都公司转基因抗草甘膦棉花的基因序列特异引物,对半野生棉极端材料进行转基因成分检测,证明筛选出的极端材料不含转抗草甘膦基因成分。采用陆地棉(珂字棉312)中epsps基因全长引物对半野生棉材料DNA进行扩增,扩增产物经测序并翻译成氨基酸序列,比对分析发现半野生棉epsps基因没有特异的氨基酸突变位点,说明半野生棉材料抗性的产生不是由epsps基因突变而致。对材料喷施0.2%的草甘膦溶液,48 h后取样,采用RT-PCR方法检测epsps基因表达量变化,发现草甘膦处理后,epsps基因表达量无明显变化,这表明半野生棉草甘膦抗性可能不是由epsps基因引起,而是其它抗性机制而致,有待以新的思路去研究。3.关联分析方法挖掘半野生棉草甘膦抗性基因源2013年,在三个不同环境下,对202份半野生棉材料进行与草甘膦抗性相关表型的测定,包括莽草酸含量和地上部分相对干重,并与覆盖全基因组的182对SSR多态性标记的基因型数据进行全基因组关联分析,共检测到72个与草甘膦抗性相关的位点,其中,48个和41个位点分别与莽草酸含量和地上部分相对干重显著相关。基于成功关联的SSR引物序列和中国农业科学院棉花研究所公布的雷蒙德氏棉(G.raimondii)、亚洲棉(G.arboreum L)和陆地棉(G.hirsutum)全基因组测序数据,共得到87条候选基因,包括亚洲棉中的21条,雷蒙德氏棉中的24条和陆地棉中的42条。以可可(Theobroma cacao)为模式植物,将这些基因在KEGG数据库中进行注释,探索相关的代谢通路,其中,ABC转运蛋白家族可能与草甘膦抗性相关,且已被前人证明,另外注释结果也推测组氨酸代谢、氨基酸生物合成和次生代谢都与草甘膦抗性有关。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1004-1389.2012.04.040URL [本文引用: 1]
通过室内和大田小区试验初步测定8种常用除草剂对黄芩种子萌发及 幼苗生长的影响.结果表明,4种苗前处理除草剂乙草胺、金都尔、上格747和农达在室内试验中对黄芩种子萌发均无影响,但都使黄芩幼苗生长受到抑制;在大 田条件下乙草胺和金都尔对黄芩出苗和幼苗生长均无影响.4种苗后茎叶处理除草剂中除莎阔丹使黄芩幼苗产生药害外,其余3种除草剂在室内条件下对黄芩幼苗安 全;烟嘧·莠去津在大田条件下对黄芩产生药害,高效盖草能对黄芩的安全性较好.
DOI:10.3969/j.issn.1004-1389.2012.04.040URL [本文引用: 1]
通过室内和大田小区试验初步测定8种常用除草剂对黄芩种子萌发及 幼苗生长的影响.结果表明,4种苗前处理除草剂乙草胺、金都尔、上格747和农达在室内试验中对黄芩种子萌发均无影响,但都使黄芩幼苗生长受到抑制;在大 田条件下乙草胺和金都尔对黄芩出苗和幼苗生长均无影响.4种苗后茎叶处理除草剂中除莎阔丹使黄芩幼苗产生药害外,其余3种除草剂在室内条件下对黄芩幼苗安 全;烟嘧·莠去津在大田条件下对黄芩产生药害,高效盖草能对黄芩的安全性较好.

{kind=link}
{kind=link}