 ,1,*
,1,*Identification of fertility restoration and molecular mapping of restorer genes in two maize restore lines of CMS-C
MOU Bi-Tao1,2, ZHAO Zhuo-Fan1, YUE Ling1, LI Chuan1, ZHANG Jun3, LI Zhang-Bo3, SHEN Han3, CAO Mo-Ju,1,*通讯作者:
第一联系人:
收稿日期:2018-04-16接受日期:2018-10-8网络出版日期:2018-11-05
| 基金资助: |
Corresponding authors:
Received:2018-04-16Accepted:2018-10-8Online:2018-11-05
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (4269KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
牟碧涛, 赵卓凡, 岳灵, 李川, 张钧, 李章波, 申汉, 曹墨菊. 两份玉米CMS-C恢复系的育性恢复力测定及恢复基因的分子标记定位[J]. 作物学报, 2019, 45(2): 225-234. doi:10.3724/SP.J.1006.2019.083033
MOU Bi-Tao, ZHAO Zhuo-Fan, YUE Ling, LI Chuan, ZHANG Jun, LI Zhang-Bo, SHEN Han, CAO Mo-Ju.
细胞质雄性不育(cytoplasmic male sterility, CMS)作为“三系”配套杂交制种的重要工具, 在杂交种子的生产以及杂种优势利用上具有重要应用价值。根据恢复专效性, 可将玉米雄性不育胞质分为C型(CMS-C)、T型(CMS-T)和S型(CMS-S) 3种类型[1]。Pring等[2]根据线粒体基因组的酶切图谱将C型胞质分为CI (C)、CII (RB、BB及E)和CIII (ES) 3个亚组。而对于恢复基因(restorer gene, Rf)及恢复系的研究既有助于CMS的不育化制种应用, 也有助于对育性恢复机制的探究。迄今, 国内外****对玉米细胞质雄性不育的育性恢复进行了大量研究, CMS-T的育性恢复由基因Rf1和Rf2控制, Duvick等[3]将Rf1定位于玉米第3染色体短臂上, Snyder等[4]将Rf2定位于玉米第9染色体靠近Wx基因座位的附近区域。Wise等[5]的进一步研究将Rf1定位于第3染色体的umc97与umc92之间, 距离umc97为1.1 cM, 将Rf2定位在距离umc153为3.8 cM的位点上, Cui等[6]利用转座子标签法成功克隆得到Rf2基因。CMS-S的恢复主基因为Rf3, Zhang等[7]利用AFLP、SCAR标记将Rf3定位在第2染色体的长臂上, 位于标记E7P6与E12M7之间, 遗传距离分别为0.9 cM和1.8 cM。李鹏等[8]利用BC1F1同质群体将Rf3精细定位在第2染色体的长臂上, 位于SSR标记A165与CG2之间, 两标记的物理距离为1.4 Mb。此外, Feng等[9]通过全基因组关联分析发现, 除主效恢复基因Rf3外, 在染色体的其他位置上还有近30个位点与CMS-S的育性恢复有关。CMS-C的育性恢复机制较为复杂, Kheyr-Pour等[10]研究表明, C型细胞质雄性不育的育性恢复受显性基因Rf4控制; 陈伟程等[11]则认为, C型不育系的育性恢复受2对具有重叠效应的基因控制; Vidakovic等[12,13,14]提出, 玉米CMS-C的育性恢复可能受3对甚至更多的互补基因控制。Sisco等[15]利用RFLP标记将玉米自交系A619中的恢复基因Rf4定位在玉米第8染色体的短臂上, 与RFLP标记Npi114a连锁; 汤继华等[16]利用SSR标记同样将A619的恢复基因Rf4定位于玉米第8染色体的短臂, 与SSR标记bnlg2307的遗传距离为12.3 cM, 同时通过对玉米自交系凤可1号的研究, 将恢复基因Rf5定位在玉米第5染色体上, 与SSR标记bnlg1346、phi058、bnlg1711连锁, 遗传距离分别为1.68 cM、9.87 cM、7.51 cM。此外, Kohls等[17]通过对玉米C型不育系B37C与恢复系K55的研究, 分别在bin 2.09、bin 3.06和bin 7.03 3个染色体区段上检测到控制CMS-C育性部分恢复基因的主效QTL。
S型不育系属于配子体不育, 败育时期晚, 育性不稳定[18,19], 一定程度上阻碍其在生产上的利用; T型不育系属于孢子体不育, 败育时期早, 败育彻底, 但由于玉米小斑病T小种的专化侵染, 致使T型不育系的利用被迫停止[20]; 作为孢子体不育的C型不育系不仅败育早, 且败育彻底[21], 长期以来受到育种家的广泛关注, 但目前生产上因缺乏对CMS-C具有强恢复力的恢复系, 致使CMS-C的生产利用受到限制, 因此发掘、鉴定玉米CMS-C新恢复源并对其进行基因定位及克隆研究无疑具有十分重要的意义。本课题组在前期的研究中发现玉米自交系Z16、7250-14-1对C黄早四和C478表现为育性完全恢复, 因此本研究一方面通过广泛测交鉴定2份自交系的恢复力; 另一方面通过杂交、自交结合回交进行恢复基因的遗传分析, 同时利用F2群体进行恢复基因的分子标记定位, 为2份自交系在不育化制种中的生产利用及恢复基因的克隆提供重要参考。
1 材料与方法
1.1 试验材料
以同质异核、同核异质的CMS-C不育系以及CMS-T、CMS-S不育系为母本(表1), 分别与玉米自交系Z16、7250-14-1 (由内蒙古真金种业科技有限公司提供)杂交得到F1, 用于恢保关系的测定。以不育系C黄早四、C478为母本, 自交系Z16、7250-14-1为父本, 构建相应的F2群体; 同时以黄早四、478作为轮回亲本与F1回交, 得到相应的回交群体BC1。所有F2和BC1群体用于遗传分析, (C黄早四×Z16)F2和(C黄早四×7250-14-1)F2作为基因定位群体。Table 1
表1
表1玉米同核异质、同质异核不育系
Table 1
| C型不育系 CMS-C | T型不育系 CMS-T | S型不育系 CMS-S | C型亚组不育系 CMS-C subgroup |
|---|---|---|---|
| CMo17 | TMo17 | SMo17 | G48-2 |
| C698-3 | T698-3 | S698-3 | EC48-2 |
| C黄早四 C Huangzaosi | ES48-2 | ||
| C478 | RB48-2 | ||
| C48-2 | 类48-2 Lei 48-2 |
新窗口打开|下载CSV
1.2 育性鉴定
采用Duvick 5级分类标准鉴定育性[22]。对所有实验材料单株挂牌, 严格按照Duvick的方法逐株进行育性调查, 调查时间为植株整个散粉期, 每隔1 d调查1次, 每株至少调查3次; 采用I2-IK染色法, 于植株散粉期, 在每株雄穗主穗的上、中、下3个部位分别取1对小穗, 用固定液FAA固定, 进行花粉镜检。1.3 恢复基因的遗传分析
于不同年份不同地点种植F2、BC1群体, 统计各群体中可育株与不育株的分离比例, 并进行卡方检验。1.4 恢复基因的分子标记定位
采用CTAB法[23]提取叶片总DNA, 从F2分离群体中随机选取10株完全可育株和10株完全不育株, 等量混合其DNA, 分别构建可育基因池和不育基因池, 参照网站https://www.maizegdb.org/上的引物序列信息以及Qu等[24]设计的InDel引物信息, 由生工生物工程(上海)股份有限公司合成引物。利用分布于玉米10条染色体上的1062对SSR引物及29对InDel引物在亲本间进行多态性引物筛选, 然后经混合群体分离分析法(bulked segregant analysis, BSA)对筛选到的多态性引物进一步筛选, 最后利用获得的多态性引物对F2群体的完全不育单株进行基因分型, 寻找与目的基因连锁的分子标记。基于初步定位的结果, 从https://www.maizegdb. org/网站上下载定位区间内DNA序列信息, 用SSRHUNTER搜索SSR位点, 在https://www.ncbi.nlm. nih.gov/上设计SSR引物; 同时, 根据亲本间的DNA序列差异, 利用Oligo 7设计InDel引物, 并由生工生物工程(上海)股份有限公司合成。扩大F2群体, 利用新开发的多态性标记及初定位的分子标记对F2群体的所有完全不育单株进行基因分型。PCR体系为15 μL, 包括7.5 μL 2×Taq PCR Master Mix (由成都擎科梓熙生物技术有限公司提供)、1.5 μL DNA模板、5.0 μL ddH2O、10 μmol L-1前后引物各0.5 μL。PCR反应程序为95℃ 5 min; 95℃ 50 s, 57℃ 30 s, 72℃ 40 s, 35个循环; 72℃ 10 min。PCR产物经6%非变性聚丙烯酰胺凝胶电泳, 银染后, 观察记录扩增条带并拍照。
1.5 遗传连锁图谱构建
选取F2群体中完全不育单株的分离单株为作图群体, 将与母本带型一致的单株记为A, 与父本带型一致的单株记为B, 杂合带型的单株记为H。利用Kosambi函数将重组率转化为遗传距离[25], 利用MapMaker 3.0进行连锁数据分析, 绘制遗传图谱。2 结果与分析
2.1 恢保关系测定
以同质异核、同核异质的CMS-C不育系以及CMS-T、CMS-S不育系为母本, 自交系Z16、7250-14-1为父本, 分别测交, 2017年夏季, 将所有测交组合种植于四川温江, 每个组合2行, 每行14株, 对所有测交F1进行育性鉴定, 部分测交组合的雄穗育性表现及花粉染色情况如图1。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1部分测交组合的育性表现
I: 花药不外露; II: 花药外露比例为0~25%; III: 花药外露比例为25%~50%; IV: 花药外露比例为50%~75%; V: 花药外露比例为75%~100%。
Fig. 1Fertility phenotype of some F1 combinations
I: no emerged of anthers; II: from 0 to 25% anthers emerged; III: more than 25% to 50% anthers emerged; IV: more than 50% to 75% anthers emerged; V: more than 75% anthers emerged.
2.1.1 自交系Z16的恢保关系测定 将同质异核及同核异质的CMS-C不育系与Z16测交后代的育性鉴定结果列入表2, 由表2可知, Z16对CMo17、C698-3、C黄早四、C478均表现为育性恢复, 各测交组合的雄穗育性等级均为V级, 且花粉可染率高达96.7%以上, 说明育性恢复比较彻底。组合C48-2×Z16的雄穗育性等级除少数为II级外, 大多数为III级, 同时花粉可染率也仅为39.0%, 说明Z16对C48-2表现为育性部分恢复。此外, 所有CMS-C亚组胞质测交组合的雄穗育性等级均为I级, 其中组合G48-2×Z16、EC48-2×Z16能产生不可染的花粉, 而组合ES48-2×Z16、RB48-2×Z16以及类48-2×Z16为无花粉型, 即不能产生花粉。说明Z16对48-2核背景下的C胞质其他亚组胞质均不具有育性恢复能力。
Table 2
表2
表2CMS-C不育系与Z16测交后代的育性鉴定结果
Table 2
| 组合 Combination | 雄穗育性等级 Fertility grade of tassel | 花粉可染率 Pollen staining rate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | 可染 Fully staining (%) | 半染 Semi-staining (%) | 不染 Not staining (%) | ||
| CMo17×Z16 | 0 | 0 | 0 | 0 | 28 | 98.6 | 0 | 1.4 | |
| C698-3×Z16 | 0 | 0 | 0 | 0 | 25 | 96.7 | 0 | 3.3 | |
| C黄早四×Z16 C Huangzaosi×Z16 | 0 | 0 | 0 | 0 | 29 | 97.4 | 0 | 2.6 | |
| C478×Z16 | 0 | 0 | 0 | 0 | 28 | 97.8 | 0 | 2.2 | |
| C48-2×Z16 | 0 | 4 | 24 | 0 | 0 | 39.0 | 26.9 | 34.1 | |
| G48-2×Z16 | 28 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | |
| EC48-2×Z16 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | |
| ES48-2×Z16 | 29 | 0 | 0 | 0 | 0 | - | - | - | |
| RB48-2×Z16 | 28 | 0 | 0 | 0 | 0 | - | - | - | |
| 类48-2×Z16 Lei48-2×Z16 | 28 | 0 | 0 | 0 | 0 | - | - | - | |
新窗口打开|下载CSV
由表3可知, Z16对TMo17、T698-3均表现为不育性保持, 测交组合均为无花粉型。测交组合SMo17×Z16的雄穗育性等级多为III级, 花粉可染率仅为22.0%; 而组合S698-3×Z16的雄穗育性等级为I级, 但花粉可染率达到29.2%, 表明Z16对CMS-S表现育性部分恢复。
Table 3
表3
表3CMS-T、CMS-S不育系与Z16测交后代的育性鉴定结果
Table 3
| 组合 Combination | 雄穗育性等级Fertility grade of tassel | 花粉可染率Pollen staining rate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | 可染 Fully staining (%) | 半染 Semi-staining (%) | 不染 Not staining (%) | ||
| TMo17×Z16 | 28 | 0 | 0 | 0 | 0 | — | — | — | |
| T698-3×Z16 | 29 | 0 | 0 | 0 | 0 | — | — | — | |
| SMo17×Z16 | 0 | 5 | 24 | 0 | 0 | 22.0 | 12.1 | 65.9 | |
| S698-3×Z16 | 30 | 0 | 0 | 0 | 0 | 29.2 | 33.3 | 37.5 | |
新窗口打开|下载CSV
2.1.2 自交系7250-14-1的恢保关系测定 由表4可知, 7250-14-1对CMo17、C698-3、C黄早四以及C478均表现为育性恢复, 各测交组合的雄穗育性等级均为V级, 且花粉可染率均超过91%, 说明恢复彻底; 而测交组合C48-2×7250-14-1的雄穗育性等级为III级, 花粉可染率高达83.3%, 说明7250-14-1对C48-2表现为育性部分恢复。同时, 7250-14-1对G48-2、EC48-2及ES48-2均表现为育性部分恢复, 虽然3个测交组合的雄穗育性等级均为III级, 但组合G48-2×7250-14-1、ES48-2×7250- 14-1的花粉可染率超过94%远高于组合EC48-2× 7250-14-1(22.6%)。7250-14-1对RB48-2和类48-2表现为不育性保持, 测交组合的雄穗育性等级均为I级, 且花粉均完全不可染。表明7250-14-1对48-2背景下不同亚组胞质的育性恢复能力不同。
Table 4
表4
表4CMS-C不育系与7250-14-1测交后代的育性鉴定结果
Table 4
| 组合 Combination | 雄穗育性等级Fertility grade of tassel | 花粉可染率Pollen staining rate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | 可染 Fully staining (%) | 半染 Semi-staining (%) | 不染 Not staining (%) | ||
| CMo17×7250-14-1 | 0 | 0 | 0 | 0 | 28 | 96.6 | 3.3 | 0 | |
| C698-3×7250-14-1 | 0 | 0 | 0 | 0 | 29 | 91.9 | 2.7 | 5.4 | |
| C黄早四×7250-14-1 C Huangzaosi×7250-14-1 | 0 | 0 | 0 | 0 | 30 | 98.8 | 0 | 1.2 | |
| C478×7250-14-1 | 0 | 0 | 0 | 0 | 29 | 97.8 | 0 | 2.2 | |
| C48-2×7250-14-1 | 0 | 0 | 29 | 0 | 0 | 83.3 | 11.1 | 5.6 | |
| G48-2×7250-14-1 | 0 | 0 | 28 | 0 | 0 | 94.1 | 0 | 5.9 | |
| EC48-2×7250-14-1 | 0 | 0 | 28 | 0 | 0 | 22.6 | 25.0 | 52.4 | |
| ES48-2×7250-14-1 | 0 | 2 | 26 | 0 | 0 | 96.6 | 0 | 3.4 | |
| RB48-2×7250-14-1 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | |
| 类48-2×7250-14-1 Lei48-2×7250-14-1 | 28 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | |
新窗口打开|下载CSV
由表5可知, 7250-14-1对TMo17、T698-3均表现为不育性保持, 且测交组合均为无花粉型。对SMo17、S698-3均表现为育性部分恢复, 测交组合的雄穗育性等级均为IV级, 其中组合S698-3× 7250-14-1的花粉可染率为43.9%, 而SMo17×7250- 14-1的花粉可染率仅为14.1%。
Table 5
表5
表5CMS-T、CMS-S不育系与7250-14-1测交后代的育性鉴定结果
Table 5
| 组合 Combination | 雄穗育性等级Fertility grade of tassel | 花粉可染率Pollen staining rate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | 可染 Fully staining (%) | 半染 Semi-staining (%) | 不染 Not staining (%) | ||
| TMo17×7250-14-1 | 29 | 0 | 0 | 0 | 0 | — | — | — | |
| T698-3×7250-14-1 | 29 | 0 | 0 | 0 | 0 | — | — | — | |
| SMo17×7250-14-1 | 0 | 0 | 0 | 29 | 0 | 14.1 | 15.3 | 70.6 | |
| S698-3×7250-14-1 | 0 | 0 | 0 | 28 | 0 | 43.9 | 43.9 | 12.2 | |
新窗口打开|下载CSV
2.2 遗传分析
将不育系C黄早四、C478与恢复系Z16、7250-14-1组配的F2及BC1群体于2016—2017年分别在云南西双版纳(景洪)、四川温江及崇州进行育性鉴定, 育性调查结果见表6和表7。Table 6
表6
表6不育系与自交系Z16杂交后代的育性调查结果
Table 6
| 组合 Combination | 可育株 No. of fertile plants | 不育株 No. of sterile plants | 总株数 Total No. of plants | 理论比例 Theoretical ratio | χ2 | 年份和地点 Year and place |
|---|---|---|---|---|---|---|
| (C黄早四×Z16)F2 (C Huangzaosi×Z16) F2 | 287 | 115 | 402 | 3:1 | 1.33 | 2016景洪 Jinghong |
| 299 | 106 | 405 | 3:1 | 0.15 | 2017温江 Wenjiang | |
| 1820 | 616 | 2436 | 3:1 | 0.05 | 2017崇州 Chongzhou | |
| (C黄早四×Z16)×黄早四 (C Huangzaosi×Z16)×Huangzaosi | 33 | 36 | 69 | 1:1 | 0.06 | 2016温江 Wenjiang |
| 98 | 107 | 205 | 1:1 | 0.20 | 2017温江 Wenjiang | |
| (C478×Z16)F2 | 251 | 107 | 358 | 3:1 | 2.15 | 2017温江 Wenjiang |
| (C478×Z16)×478 | 134 | 135 | 269 | 1:1 | 0 | 2017温江 Wenjiang |
新窗口打开|下载CSV
Table 7
表7
表7不育系与自交系7250-14-1杂交后代的育性调查结果
Table 7
| 组合 Combination | 可育株 No. of fertile plants | 不育株 No. of sterile plants | 总株数 Total No. of plants | 期望比例 Theoretical ratio | χ2 | 年份、地点 Year and place |
|---|---|---|---|---|---|---|
| (C黄早四×7250-14-1)F2 (C Huangzaosi×7250-14-1) F2 | 271 | 85 | 356 | 3:1 | 0.12 | 2016景洪 Jinghong |
| 319 | 104 | 423 | 3:1 | 0.02 | 2017温江 Wenjiang | |
| 1386 | 475 | 1861 | 3:1 | 0.14 | 2017崇州 Chongzhou | |
| (C黄早四×7250-14-1)×黄早四 (C Huangzaosi×7250-14-1) ×Huangzaosi | 23 | 22 | 45 | 1:1 | 0 | 2016温江 Wenjiang |
| 124 | 141 | 265 | 1:1 | 0.55 | 2017温江 Wenjiang | |
| (C478×7250-14-1)F2 | 163 | 110 | 273 | 9:7 | 1.19 | 2016温江 Wenjiang |
| 243 | 159 | 392 | 9:7 | 1.44 | 2017温江 Wenjiang | |
| (C478×7250-14-1)×478 | 59 | 191 | 247 | 1:3 | 0.37 | 2017温江 Wenjiang |
新窗口打开|下载CSV
由表6可知, (C黄早四×Z16)F2及[(C黄早四×Z16)×黄早四]群体中可育单株与不育单株的比例分别符合3:1和1:1 (P>0.05), (C478×Z16)F2及[(C478×Z16)×478]群体中可育单株与不育单株的比例也分别符合3:1和1:1 (P>0.05), 说明Z16对C黄早四和C478的育性恢复均受1对基因控制。
由表7可知, (C黄早四×7250-14-1)F2和[(C黄早四×7250-14-1)×黄早四]群体中可育单株与不育单株的比例分别符合3:1和1:1 (P>0.05), 说明7250-14-1对C黄早四的育性恢复受1对基因控制;与此同时, (C478×7250-14-1)F2和[(C478×7250-14-1)×478]群体可育单株与不育单株的比例分别符合9:7和1:3 (P>0.05), 说明7250-14-1对C478的育性恢复受2对基因控制, 且呈显性互补关系。
2.3 恢复基因的分子标记定位
2.3.1 自交系Z16恢复基因的分子标记定位 选用位于玉米10条染色体上的1062对SSR引物以及29对InDel引物, 对亲本C黄早四与Z16进行多态性分析, 共有404对引物在两亲本间呈现多态性。利用BSA法对筛选到的多态性引物进一步筛选, 发现位于第8染色体短臂上的InDel引物IDP8573、Chr8-1330080、TIDP5557、Chr8-86080在可育基因池与不育基因池之间呈现多态性(引物信息见附表1)。利用这些多态性引物对(C黄早四×Z16)F2群体中的156株完全不育单株进行基因分型发现, TIDP5557检测到4个交换单株, Chr8-86080未检测到交换单株, 因此初步将Z16的恢复基因定位于标记TIDP5557至第8染色体短臂末端的区域(图2)。基于初定位结果, 在标记TIDP5557至第8染色体短臂末端区域内新开发了4对分子标记(附表1), 利用新开发的分子标记以及初定位的连锁标记对(C黄早四×Z16)F2群体中的798株完全不育单株进行基因分型。结果发现, 标记B-1检测到2个交换单株, 而B-6-1、B-2以及Chr8-86080均未检测到交换单株, 于是将Z16的恢复基因定位于分子标记B-1至第8染色体短臂末端的区域, 该区域的物理范围为494 kb (图2)。Supplementary table 1
附表1
附表1用于Z16恢复基因定位的多态性引物
Supplementary table 1
| 引物 Primer | 引物类型 Primer type | 正向序列 Forward sequence (5'-3') | 反向序列 Reverse sequence (5'-3') |
|---|---|---|---|
| Chr8-86080 | InDel | CGTCGTTGAGGTGAGAAGAG | CTCCGAACCTGATCCGAGTA |
| B-2 | InDel | ACGAATACGATACGTAGCCA | GTGAATCTGCGGTGAACAAA |
| B-6-1 | InDel | GGATGGAATATATAAAGTTTGCT | GGCTCATTACCTTGGTGTCA |
| B-1 | InDel | GATCGTTCCGGCCCAAGAAG | TAGCCGTGGAGTTGGTAGCC |
| m-1 | SSR | CATTGACCGGGGTAGGAAGT | CATTGACCGGGGTAGGAAGT |
| TIDP5557 | InDel | CATGAGATCAACGGGATGC | AGTAGAGATCCGGGAGGTGG |
| Chr8-1330080 | InDel | CCAAGTTGGATACAACGACAGA | AGAAGCAACGTCTGCAGGAT |
| IDP8573 | InDel | CGAGTCAGTTGCTTACGGG | AATTGCCGAGTGGATACAGG |
新窗口打开|下载CSV
图2
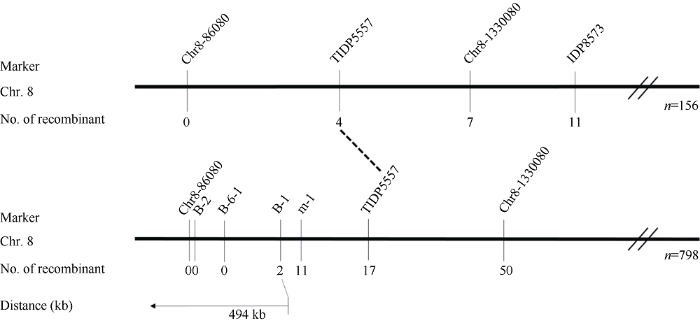 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2自交系Z16所含Rf基因在第8染色体上的连锁图谱
Fig. 2Linkage map of the Rf on chromosome 8 in Z16
2.3.2 自交系7250-14-1恢复基因的分子标记定位
选用位于玉米10条染色体上的1062对SSR引物和29对InDel引物在亲本C黄早四与7250-14-1之间进行多态性分析, 共有389对引物在两者间存在多态性。利用BSA法对筛选到的多态性引物进一步筛选发现, 位于第8染色体短臂上的InDel引物Chr8-1330080、IDP7866、IDP500、IDP8319、Chr8- 398180及Chr8-86080在可育基因池与不育基因池之间呈现多态性(引物信息见附表2)。利用这些多态性引物对(C黄早四×7250-14-1)F2群体中的143株完全不育单株进行基因分型发现, 引物IDP8319检测到1个交换单株, 而Chr8-398180、Chr8-86080均未检测到交换单株, 因此初步将7250-14-1的恢复基因定位于IDP8319至第8染色体短臂末端的区域(图3)。基于初定位结果, 在标记IDP8319至第8染色体短臂末端区域内新开发了4对分子标记(附表2), 利用新开发的分子标记以及初定位的连锁标记对(C黄早四×7250-14-1)F2群体中的432株完全不育单株进行基因分型。发现标记B-1检测到4个交换单株, Chr8-86080检测到1个交换单株, 且利用B-1和Chr8-86080检测到的交换单株不相同, 因此将7250-14-1的恢复基因定位于标记B-1与Chr8-86080之间, 物理距离为249 kb (图3)。
图3
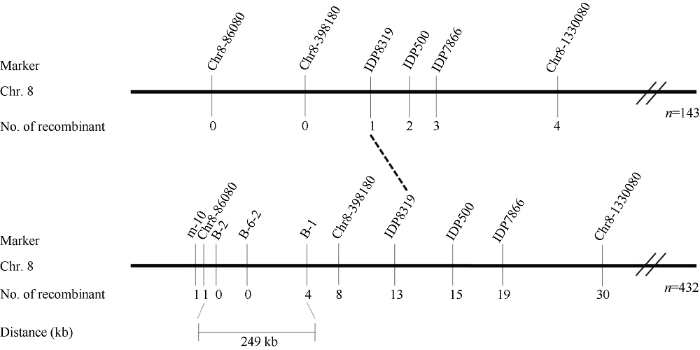 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3自交系7250-14-1所含Rf基因在第8染色体上的连锁图谱
Fig. 3Linkage map of the Rf on chromosome 8 in 7250-14-1
Supplementary table 2
附表2
附表2用于7250-14-1恢复基因定位的多态性引物
Supplementary table 2
| 引物 Primer | 引物类型 Primer type | 正向序列 Forward sequence (5'-3') | 反向序列 Reverse sequence (5'-3') |
|---|---|---|---|
| m-10 | SSR | AGCGCTCGATTCCTGTAGTG | GGGTGTCGTTGGTTGGGATT |
| Chr8-86080 | InDel | CGTCGTTGAGGTGAGAAGAG | CTCCGAACCTGATCCGAGTA |
| B-2 | InDel | ACGAATACGATACGTAGCCA | GTGAATCTGCGGTGAACAAA |
| B-6-2 | InDel | CCAATGTTTTGATGGAAGTCCT | AATTGCCATGTTCTTACCTGT |
| B-1 | InDel | GATCGTTCCGGCCCAAGAAG | TAGCCGTGGAGTTGGTAGCC |
| Chr8-398180 | InDel | GCCAGTTCGGAGACAGGAT | ACCGCCATCCAATTAACAAG |
| IDP8319 | InDel | TTGACCCTCCTGTTACGTGC | GAGCATGGACCACATGACC |
| IDP500 | InDel | CACTGCCGTAGAGTAGTGCG | GGCTTCAAGATCAGTCCG |
| IDP7866 | InDel | GGACGAAGCGATCGAGTACC | AGATGAGGGAAGTGAGCAGC |
| Chr8-1330080 | InDel | CCAAGTTGGATACAACGACAGA | AGAAGCAACGTCTGCAGGAT |
新窗口打开|下载CSV
3 讨论
“三系”配套是植物细胞质雄性不育应用于不育化制种的前提。而其中恢复系的选育及恢复力的表现又直接关系到不育化制种的应用成效。本研究发现, Z16和7250-14-1除了对玉米CMS-C的CI亚组中的C48-2表现为部分恢复外, 对CI亚组的C478、C黄早四、C698-3等均表现为完全恢复。自交系A619是含有主效恢复基因Rf4的强恢复系, 它不仅对C478、C黄早四、C698-3表现为强恢复, 对C48-2也同样表现为强恢复[26]。可见, Z16和7250-14-1对CMS-C不育系的育性恢复能力不同于A619。通过对玉米CMS-C不同亚组胞质的恢保关系测定发现, Z16不能恢复RB48-2、ES48-2、G48-2、EC48-2、类48-2的育性; 7250-14-1能够部分恢复ES48-2、G48-2、EC48-2的育性, 但对RB48-2、类48-2则表现为不育性保持。从这个意义上来说Z16与7250-14-1对玉米CMS-C的育性恢复能力也存在差异。本研究基于2个不育系背景下的F2及BC1群体的遗传分析发现, Z16对C黄早四和C478的育性恢复均受1对基因控制, 而7250-14-1对C黄早四的育性恢复受1对基因控制, 但对C478则表现为受2对基因控制。于是我们选择不育系C黄早四做母本, Z16及7250-14-1分别做父本, 配制F2作为基因定位群体, 分别对Z16及7250-14-1中的主效恢复基因进行分子标记定位。最终将Z16和7250-14-1的主效恢复基因均定位于第8染色体短臂末端, 且定位区间包含了Rf4候选基因GRMZM2G021276[27]。2份自交系的定位区间存在较大的重叠区域, 参照B73序列(http://ensembl.gramene.org/Zea_mays/Info/Index), 发现重叠区间内共有5个注释基因, 其中Zm00001 d008173编码过氧化物酶1前体蛋白, Zm00001d00 8174编码bHLH转录因子, Zm00001d008175编码S-腺苷甲硫氨酸合酶1, Zm00001d008176编码SBP结构域转录因子家族蛋白, Zm00001d008177编码类PTI1酪氨酸蛋白激酶3。而且, Zm00001d008174为已经报道的玉米核雄性不育基因ms23[28]。分子标记定位结果显示3个恢复材料Z16、7250-14-1及A619的恢复基因均被定位于第8染色体的短臂上, 但3个恢复材料对C型不育系的恢复能力存在差别。基于以上结果我们推测, 3个自交系都含有Rf4基因, 但在不同核背景下其发挥作用的效应不同; 或者它们分别含有不同的恢复基因, 但却在第8染色体短臂上成簇存在。关于恢复基因在染色体上成簇存在的现象在其他作物上已有报道[29,30]。
本研究新鉴定出的玉米CMS-C强恢复系Z16和7250-14-1, 虽然对玉米CMS-C的CI亚组具有相同的恢复力, 但对CII和CIII亚组的恢复能力却存在差异。Z16、7250-14-1对玉米CMS-C的恢复能力也有别于自交系A619。可见玉米细胞质雄性不育系的育性恢复既受测验系的核基因影响, 也受不育系的核背景及不育细胞质类型影响。因此在将不育化制种应用于生产时, 对恢复系的选育, 既要考虑到不育系的核背景, 也要考虑所用C型不育的亚组胞质类型。
4 结论
自交系Z16、7250-14-1均能恢复不同核背景下的CI胞质, 其中对大多数核背景下的CI胞质表现为育性完全恢复, 仅对48-2背景下的CI胞质表现为育性部分恢复。Z16对C黄早四、C478的育性恢复受1对基因控制; 7250-14-1对C黄早四的育性恢复受1对基因控制, 而对C478的育性恢复则受2对基因控制, 且显现互补关系。2份自交系的恢复基因均被定位于第8染色体短臂上, 其中Z16所含恢复基因被定位于分子标记B-1至第8染色体短臂末端区域, 物理距离为494 kb; 7250-14-1所含恢复基因被定位于分子标记B-1与Chr8-86080之间, 物理距离为249 kb。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.2135/cropsci1980.0011183X002000020002xURL [本文引用: 1]

Restriction endonuclease fragment analysis showed three mitochondrial DNA types to exist. On the basis of this, the five cytoplasms studied were grouped as follows: CI (C cytoplasm), CII (Rancho Barrieto, BB and E cytoplasms) and CIII (El Salvador cytoplasm).
DOI:10.2135/cropsci1969.0011183X000900020012xURLPMID:13888759 [本文引用: 1]
DUVICK DN, SNYDER RJ, ANDERSON EG.
[本文引用: 1]
DOI:10.1007/bf01253987URLPMID:24186179 [本文引用: 1]
Abstract There are three major groups of cytoplasmic male-sterile cytoplasms in maize; C (Charrua), S (USDA), and T (Texas). These cytoplasms can be classified by the unique nuclear genes that suppress the male-sterility ef- fects of these cytoplasms and restore pollen fertility. Typ- ically, plants that carry Texas (T) cytoplasm are male fer- tile only if they carry dominant alleles at two unlinked nu- clear restorer loci, rfl and rf2. To facilitate analysis of T- cytoplasm-mediated male sterility and fertility restoration, we have mapped rfl and rf2 relative to closely-linked RFLP markers using five populations. The rfl locus and/or linked visible markers were mapped in four populations; the rf2 locus was mapped in two of the populations. Data from the individual populations were joined with the aid of JoinMap software. The resulting consensus maps place rfl between umc97 and umc92 on chromosome 3 and rf2 between umc153 and susl on chromosome 9. Markers that flank the rfl and rf2 loci have been used to identify alleles at rfl and rf2 in segregating populations. These analyses demonstrate the possibility of tracking separate fertility re- storer loci that contribute to a single phenotype.
[本文引用: 1]
DOI:10.1007/s00438-006-0131-yURLPMID:16705419 [本文引用: 1]
The Rf3 gene restores the pollen fertility disturbed by S male sterile cytoplasm. In order to develop molecular markers tightly linked to Rf3 , we used amplified fragment length polymorphism (AFLP) technique with near isogenic lines (NILs) and bulk segregant analysis (BSA). A BC 1 F 1 population from a pair of NILs with different Rf3 locus was constructed and 528 primer combinations was screened. A linkage map was constructed around the Rf3 locus, which was mapped on the distal region of chromosome 2 long arm with the help of SSR marker UMC2184. The closest marker E7P6 was 0.902cM away from Rf3 . Marker E3P1, 2.402cM from Rf3 , and E12M7, 1.802cM from Rf3 , were converted into a codominant CAPS and a dominant SCAR marker, and designated as CAPSE3P1 and SCARE12M7, respectively. These markers are useful for marker-assisted selection and map-based cloning of the Rf3 gene.
DOI:10.3969/j.issn.1001-4942.2014.08.001URL [本文引用: 1]
细胞质雄性不育(Cytoplasmic male sterility,CMS)广泛存在于高等植物,CMS-S是玉米CMS的一种,其育性可以被恢复基因Rf3完全恢复。目前Rf3基因尚未成功克隆。本试验根据玉米CMS-S配子体不育特点,提出一种全新的同质群体构建方法,通过该方法获得群体的所有个体均携带Rf3恢复基因,省去田间表型鉴定工作,并可在同一世代获得足够的重组个体。该方法可直接用种子提取基因组DNA,大大提高Rf3定位的效率。利用该同质定位群体,将恢复基因Rf3精细定位到分子标记A165与CG2之间,参照玉米自交系B73的基因组序列,两个标记之间的物理距离约为1.4 Mb。
DOI:10.3969/j.issn.1001-4942.2014.08.001URL [本文引用: 1]
细胞质雄性不育(Cytoplasmic male sterility,CMS)广泛存在于高等植物,CMS-S是玉米CMS的一种,其育性可以被恢复基因Rf3完全恢复。目前Rf3基因尚未成功克隆。本试验根据玉米CMS-S配子体不育特点,提出一种全新的同质群体构建方法,通过该方法获得群体的所有个体均携带Rf3恢复基因,省去田间表型鉴定工作,并可在同一世代获得足够的重组个体。该方法可直接用种子提取基因组DNA,大大提高Rf3定位的效率。利用该同质定位群体,将恢复基因Rf3精细定位到分子标记A165与CG2之间,参照玉米自交系B73的基因组序列,两个标记之间的物理距离约为1.4 Mb。
[本文引用: 1]
DOI:10.1007/BF00135050URLPMID:17249089 [本文引用: 1]
The genetics of fertility restoration of cms-C group cytoplasm of maize was studied using crosses involving stable maintainer lines and lines that restored full pollen fertility. Pollen fertility in the sources of cms-C sterile cytoplasms studied was restored by a single dominant restorer (Rf4) gene. The fertility restoration was sporophytic. Allelism tests among five restorer lines showed that they all apparently carried the same alleles (Rf4 Rf4). Similar tests also demonstrated that seven nonrestoring maintainer lines had apparently the same genotype (rf4 rf4), although a partial "late break" of fertility was observed at low levels in some maintainer crosses. Comparative studies among different cms-C sources (C, Bb, ES, PR and RB) indicated that similar inheritance of fertility restoration was involved. The data indicated that a single, dominant Rf gene is involved in the restoration of several C-group cytoplasms, at least in the lines studied here. This is the first single-gene, sporophytic restorer system described in maize to date.
URL [本文引用: 1]
67 inbred lines of maize had been tested for their restoringreaction to the C—type cytoplasmic male—sterility.They were clas-sified into four groups according to the degree of pollen restoringability.Both the male—sterility and pollen restoration manifested inthe tested lines were found to be fai
URL [本文引用: 1]
67 inbred lines of maize had been tested for their restoringreaction to the C—type cytoplasmic male—sterility.They were clas-sified into four groups according to the degree of pollen restoringability.Both the male—sterility and pollen restoration manifested inthe tested lines were found to be fai
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.2135/cropsci1991.0011183X003100050036xURL [本文引用: 1]
Previous studies of the genetics of fertility restoration for C-type cytoplasmic male sterility (cms-C) in corn (Zea mays L.) have produced conflicting results. The objective of this study was to localize the restorer gene(s) for cms-C using restriction fragment length polymorphism (RFLP) analysis. Rf4, a single dominant restorer gene for cms-C found in inbred A619, was localized to Chromosome 8, almost-equal-to 2 centimorgans from an RFLP marker locus, NPI114A. The presence in corn of extensive nucleotide duplication complicated the analysis. Because the region of Chromosome 8 near Rf4 is duplicated on Chromosome 3, an additional restorer gene for cms-C may eventually be found on Chromosome 3.
DOI:10.3321/j.issn:0578-1752.2001.06.003URL [本文引用: 1]
通过对恢复系凤可 1、A619与 Cms-C237、Cms-CMo17组配的 F2和 BC1分离群体的研究结果表明:凤可 1有两对重叠恢复基因 Rf4和Rf5。用微卫星标记(SSR)将凤可1中的恢复主基因Rf5定位在第 5染色体长臂上,与引物 bnlg1711、bnlg1346和 phi058紧密连锁,距 3个引物的遗传距离分别为 7.51cM、1.68cM、9.87cM;恢复主基因 Rf4与第8染色体短臂上的引物bnlg2307连锁。
DOI:10.3321/j.issn:0578-1752.2001.06.003URL [本文引用: 1]
通过对恢复系凤可 1、A619与 Cms-C237、Cms-CMo17组配的 F2和 BC1分离群体的研究结果表明:凤可 1有两对重叠恢复基因 Rf4和Rf5。用微卫星标记(SSR)将凤可1中的恢复主基因Rf5定位在第 5染色体长臂上,与引物 bnlg1711、bnlg1346和 phi058紧密连锁,距 3个引物的遗传距离分别为 7.51cM、1.68cM、9.87cM;恢复主基因 Rf4与第8染色体短臂上的引物bnlg2307连锁。
DOI:10.1007/s00122-011-1586-8URLPMID:21479555 [本文引用: 1]
Partial restoration of male fertility limits the use of C-type cytoplasmic male sterility (C-CMS) for the production of hybrid seeds in maize. Nevertheless, the genetic basis of the trait is still unknown. Therefore, the aim to this study was to identify genomic regions that govern partial restoration by means of a QTL analysis carried out in an F 2 population ( n = 180). This population was derived from the Corn Belt inbred lines B37C and K55. F 2 BC 1 progenies were phenotyped at three locations in Switzerland. Male fertility was rated according to the quality and number of anthers as well as the anthesis-silking interval. A weak effect of environment on the expression of partial restoration was reflected by high heritabilities of all fertility-related traits. Partial restoration was inherited like an oligogenic trait. Three major QTL regions were found consistently across environments in the chromosomal bins 2.09, 3.06 and 7.03. Therefore, a marker-assisted counter-selection of partial restoration is promising. Minor QTL regions were found on chromosomes 3, 4, 5, 6 and 8. A combination of partial restorer alleles at different QTL can lead to full restoration of fertility. The maternal parent was clearly involved in the partial restoration, because the restorer alleles at QTL in bins 2.09, 6.04 and 7.03 originated from B37. The three major QTL regions collocated with other restorer genes of maize, a phenomenon, which seems to be typical for restorer genes. Therefore, a study of the clusters of restorer genes in maize could lead to a better understanding of their evolution and function. In this respect, the long arm of chromosome 2 is particularly interesting, because it harbors restorer genes for the three major CMS systems (C, T and S) of maize.
DOI:10.1002/j.1537-2197.1979.tb06269.xURL [本文引用: 1]
Anther development of the C-cytoplasmic male-sterile (cms C) and the normal cytoplasm version (N) in the W182BN corn inbred was studied by light and electron microscopy. Deviation from normal pollen development was first observed in the tapetal cells at the tetrad stage of development Two types of tapetal abnormalities were observed in plants with C cytoplasm. The first behaved like the N anther until the tetrad stage, when numerous small vacuoles appeared in the tapetal cells. Inner and radial tapetal cell walls broke down normally, but irregular Ubisch body deposition was observed, and exine development was inhibited and delayed. The tapetum and microspores disintegrated at the intermediate microspore stage. The second type of tapetum was highly vacuolated at the early tetrad stage, with dense inner and radial cell walls that remained intact and enlarged when the tetrads aborted. No organellar abnormalities, such as the mitochondrial changes observed in cms T, were observed in C anthers.
DOI:10.1093/oxfordjournals.aob.a086145URL [本文引用: 1]
Examination of the structural changes occurring during microsporogenesis in cms-T, -C and -S Zea mays L. with different nuclear backgrounds revealed considerable variations in the stages at which abortion of the sporocytes occurred within each cytoplasmic genotype. This variation was not associated with the nuclear background. Significantly, we found that the mitochondria in cms-T plants did not always follow the pattern of degeneration described previously. We conclude that these observations are not consistent with a proposed mechanism of cytoplasmic male sterility involving the direct, antagonistic action of an anther produced substance on an altered mitochondrion or plastid, that is functionally normal in the rest of the plant. The value of microscopic studies in detecting variation in apparently uniform material is emphasised, though it is recognized that other methods will be required to elucidate the mechanism of male sterility.
DOI:10.1126/science.171.3976.1113URLPMID:17777595 [本文引用: 1]
In "The sotuthern corn leaf blight epidemic" by L. A. Tatum (19 Mar., p. 1113), the first sentence of the third paragraph under the sidehead "History of southern corn leaf blight" on page 1114 should read "The loss in 1969 rather than "1968."
DOI:10.2135/cropsci2007.12.0694URL [本文引用: 1]
Cytoplasmic male sterility (CMS) is a maternally transmitted trait, whereby a plant is unable to produce viable pollen. Studies have revealed that this trait is a tool for enabling effi cient and reliable coexistence between genetically modified (GM) and non-GM cultivation by biocontainment of GM maize (Zea mays L.) pollen. Maize has three types of male-sterile cytoplasm (T, S, and C), the fertility of which can be restored by nuclear rf genes or by interactions with the environment. Twenty-two CMS versions of modern European maize hybrids were evaluated in 17 environments in Switzerland, France, and Bulgaria, with two or three sowing dates, in 2005 and 2006. Stable and unstable male sterility occurred in all three CMS types. T-cytoplasm hybrids were the most stable, while S-cytoplasm hybrids often showed partial restoration of fertility. C-cytoplasm was similar to T-cytoplasm with regard to maintaining male sterility. Climatic factors, especially air temperature, evapotranspiration, and water vapor, during the 10 d before anthesis as well as during anthesis, were correlated positively or negatively with the partial reversion to male fertility of CMS hybrids, indicating an interaction between genetic and climatic factors. This study illustrates that T- and C-cytoplasm in particular open up viable prospects for containing transgenic pollen, especially for Bt-maize.
DOI:10.1016/S0065-2660(08)60046-2URL [本文引用: 1]
DOI:10.1007/BF02772108URL [本文引用: 1]
A relatively quick, inexpensive and consistent protocol for extraction of DNA from expanded leaf material containing large quantities of polyphenols, tannins and polysaccharides is described. Mature strawberry leaves, which contain high levels of these secondary components, were used as a study group. The method involves a modified CTAB extraction, employing high salt concentrations to remove polysaccharides, the use of polyvinyl pyrrolidone (PVP) to remove polyphenols, an extended RNase treatment and a phenol-chloroform extraction. Average yields range from 20 to 84 g/g mature leaf tissue for both wild and cultivated octoploid and diploid Fragaria species. Results from 60 plants were examined, and were consistently amplifiable in the RAPD reaction with as little as 0.5 ng DNA per 25- L reaction. Presently, this is the first procedure for the isolation of DNA from mature strawberry leaf tissue that produces consistent results for a variety of different species, both octoploid and diploid, and is both stable and PCR amplifiable before and after extended storage. This procedure may prove useful for other difficult species in the family Rosaceae.
DOI:10.1186/1756-0500-6-1URL [本文引用: 1]
DOI:10.1111/j.1469-1809.1943.tb02321.xURL [本文引用: 1]
The articles published by the Annals of Eugenics (1925–1954) have been made available online as an historical archive intended for scholarly use. The work of eugenicists was often pervaded by prejudice against racial, ethnic and disabled groups. The online publication of this material for scholarly research purposes is not an endorsement of those views nor a promotion of eugenics in any way.
URL [本文引用: 1]
玉米是最早利用细胞质雄性不育系生产杂交种的作物之一,C型细胞质雄性不育系(C-type cytoplasmic male sterile,CMS-C)在杂交种生产中具有重要的作用,育性恢复的稳定性直接影响其应用价值。然而,玉米CMS-C的育性恢复机理复杂,且至今仍不明确。为进一步探究玉米CMS-C育性恢复的影响因素,本研究以玉米CMS-C同质异核不育系C48-2、C黄早四和C478为母本,分别与测验系18白、自330、5022以及恢复系A619组配杂交获得F1。其中育性恢复F1通过自交获得F2,并以育性恢复F1为父本分别给育性保持F1授粉,组配双交群体,共获得4个F2群体,6个双交群体。同时以不育系C48-2、C黄早四和C478为母本,各自的保持系48-2、黄早四和478为父本杂交组配不完全双列杂交F1。将所有杂交组合的F1、F2以及双交组合群体分别在不同年份不同地点种植观察,通过植株田间育性调查并结合室内花粉镜检鉴定育性表现。结果表明:1)同一测验系对玉米CMS-C同质异核不育系的恢保关系不同,暗示不育系的核背景参与调控育性恢复表现;2)在不同年份不同地点对(C48-2×A619)F2群体进行种植观察,发现不同环境下F2群体可育株与不育株的分离比均符合15∶1,但在云南种植的可育株的育性级别主要为Ⅲ和Ⅳ级,而在四川种植的可育株的育性级别主要为Ⅴ级,表明环境对恢复系A619恢复后代的育性表现有影响;3)通过恢保关系测定发现18白不能恢复C478,48-2也不能恢复C478,但双交群体[(C478×18白)F1S×(C48-2×18白)F1F]后代却出现了可育株与不育株的分离;同理,双交群体[(C48-2×自330)F1S×(C478×自330)F1F]的后代也出现了可育株与不育株的分离。因此,本文推测C48-2、C478核背景中存在微效恢复基因,这些微效基因与18白、自330中的微效恢复基因通过杂交聚合后能使C478、C48-2的育性恢复,暗示玉米CMS-C的育性恢复呈现一定的剂量效应。这些结果为进一步认识玉米CMS-C育性恢复的复杂性和多样性奠定了基础,为深入研究玉米CMS-C育性恢复机理以及加快CMS-C在不育化制种中的应用提供重要参考。
URL [本文引用: 1]
玉米是最早利用细胞质雄性不育系生产杂交种的作物之一,C型细胞质雄性不育系(C-type cytoplasmic male sterile,CMS-C)在杂交种生产中具有重要的作用,育性恢复的稳定性直接影响其应用价值。然而,玉米CMS-C的育性恢复机理复杂,且至今仍不明确。为进一步探究玉米CMS-C育性恢复的影响因素,本研究以玉米CMS-C同质异核不育系C48-2、C黄早四和C478为母本,分别与测验系18白、自330、5022以及恢复系A619组配杂交获得F1。其中育性恢复F1通过自交获得F2,并以育性恢复F1为父本分别给育性保持F1授粉,组配双交群体,共获得4个F2群体,6个双交群体。同时以不育系C48-2、C黄早四和C478为母本,各自的保持系48-2、黄早四和478为父本杂交组配不完全双列杂交F1。将所有杂交组合的F1、F2以及双交组合群体分别在不同年份不同地点种植观察,通过植株田间育性调查并结合室内花粉镜检鉴定育性表现。结果表明:1)同一测验系对玉米CMS-C同质异核不育系的恢保关系不同,暗示不育系的核背景参与调控育性恢复表现;2)在不同年份不同地点对(C48-2×A619)F2群体进行种植观察,发现不同环境下F2群体可育株与不育株的分离比均符合15∶1,但在云南种植的可育株的育性级别主要为Ⅲ和Ⅳ级,而在四川种植的可育株的育性级别主要为Ⅴ级,表明环境对恢复系A619恢复后代的育性表现有影响;3)通过恢保关系测定发现18白不能恢复C478,48-2也不能恢复C478,但双交群体[(C478×18白)F1S×(C48-2×18白)F1F]后代却出现了可育株与不育株的分离;同理,双交群体[(C48-2×自330)F1S×(C478×自330)F1F]的后代也出现了可育株与不育株的分离。因此,本文推测C48-2、C478核背景中存在微效恢复基因,这些微效基因与18白、自330中的微效恢复基因通过杂交聚合后能使C478、C48-2的育性恢复,暗示玉米CMS-C的育性恢复呈现一定的剂量效应。这些结果为进一步认识玉米CMS-C育性恢复的复杂性和多样性奠定了基础,为深入研究玉米CMS-C育性恢复机理以及加快CMS-C在不育化制种中的应用提供重要参考。
US2012090047 .
URL [本文引用: 1]
The present disclosure provides a method for selecting a plant comprising a functional restorer gene for maize S-type cytoplasmic male sterility comprising the steps of (a) screening a population of plants for at least one marker nucleic acid, wherein the marker nucleic acid comprises an allele linked to the functional restorer gene for maize S-type cytoplasmic male sterility; (b) detecting the marker nucleic acid; (c) identifying a plant comprising the marker nucleic acid; and (d) selecting the plant comprising the marker nucleic acid, wherein the plant comprising the marker nucleic acid further comprises the functional restorer gene for maize S-type cytoplasmic male sterility. The present disclosure also provides methods for restoring fertility in a progeny of an S-type cytoplasmic male sterile plant and methods for transferring an Rf3 gene into a progeny plant.
DOI:10.1242/dev.140673URLPMID:27913638 [本文引用: 1]
Successful male gametogenesis involves orchestration of sequential gene regulation for somatic differentiation in pre-meiotic anthers. We report here the cloning of Male Sterile23 (Ms23), encoding an anther-specific predicted basic helix-loop-helix (bHLH) transcription factor required for tapetal differentiation; transcripts localize initially to the precursor secondary parietal cells then predominantly to daughter tapetal cells. In knockout ms23-ref mutant anthers, five instead of the normal four wall layers are observed. Microarray transcript profiling demonstrates a more severe developmental disruption in ms23-ref than in ms32 anthers, which possess a different bHLH defect. RNA-seq and proteomics data together with yeast two-hybrid assays suggest that MS23 along with MS32, bHLH122 and bHLH51 act sequentially as either homo- or heterodimers to choreograph tapetal development. Among them, MS23 is the earliest-acting factor, upstream of bHLH51 and bHLH122, controlling tapetal specification and maturation. By contrast, MS32 is constitutive and independently regulated and is required later than MS23 in tapetal differentiation.
[本文引用: 1]
DOI:10.1105/tpc.105.038240URLPMID:16489123 [本文引用: 1]
Cytoplasmic male sterility (CMS) and nucleus-controlled fertility restoration are widespread plant reproductive features that provide useful tools to exploit heterosis in crops. However, the molecular mechanism underlying this kind of cytoplasmic-nuclear interaction remains unclear. Here, we show in rice (Oryza sativa) with Boro II cytoplasm that an abnormal mitochondrial open reading frame, orf79, is cotranscribed with a duplicated atp6 (B-atp6) gene and encodes a cytotoxic peptide. Expression of orf79 in CMS lines and transgenic rice plants caused gametophytic male sterility. Immunoblot analysis showed that the ORF79 protein accumulates specifically in microspores. Two fertility restorer genes, Rf1a and Rf1b, were identified at the classical locus Rf-1 as members of a multigene cluster that encode pentatricopeptide repeat proteins. RF1A and RF1B are both targeted to mitochondria and can restore male fertility by blocking ORF79 production via endonucleolytic cleavage (RF1A) or degradation (RF1B) of dicistronic B-atp6/orf79 mRNA. In the presence of both restorers, RF1A was epistatic over RF1B in the mRNA processing. We have also shown that RF1A plays an additional role in promoting the editing of atp6 mRNAs, independent of its cleavage function.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}