 ,*西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室, 重庆 400715
,*西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室, 重庆 400715Phenotypic characterizing and gene mapping of a lesion mimic and premature senescence 1 (lmps1) mutant in rice (Oryza sativa L.)
XIA Sai-Sai, CUI Yu, LI Feng-Fei, TAN Jia, XIE Yuan-Hua, SANG Xian-Chun, LING Ying-Hua,*Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400715, China通讯作者:
收稿日期:2018-04-25接受日期:2018-08-20网络出版日期:2018-09-18
| 基金资助: |
Received:2018-04-25Accepted:2018-08-20Online:2018-09-18
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (10311KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
夏赛赛, 崔玉, 李凤菲, 谭佳, 谢园华, 桑贤春, 凌英华. 水稻类病斑早衰突变体lmps1的表型鉴定与基因定位[J]. 作物学报, 2019, 45(1): 46-54. doi:10.3724/SP.J.1006.2019.82022
XIA Sai-Sai, CUI Yu, LI Feng-Fei, TAN Jia, XIE Yuan-Hua, SANG Xian-Chun, LING Ying-Hua.
植物类病斑突变体(lesion mimic mutant, lmm)是指在没有病原菌侵染或明显逆境的情况下, 植物体自发形成类似于病原菌侵染产生坏死斑的一类突变体[1]。此类突变体首先在玉米中发现[1,2], 随后在大麦[3]、拟南芥[4]、水稻[5,6]、花生[7]等植物中相继被报道。植物类病斑突变体的坏死斑表型类似于过敏反应, 且大多数与过敏反应以及细胞程序性死亡有关, 一些类病斑突变体能提高植物抗病性并引发相关蛋白的表达。
国内外研究工作者通过各种物理化学诱变方法已鉴定到大量水稻类病斑突变体, 其多数性状受隐性基因调控, 只有少部分由显性或半显性基因控制[8]。在这些类病斑突变体中, 已被克隆的目标基因相对较少[9]。从报道看, 水稻的类病斑主要是由抗病相关基因突变、某些代谢途径中的相关基因突变、胞内运输异常以及转录因子所致。其中一类是导致lmm的抗病相关基因, 包括OsAT1[6]、OsNH1[10]、NLS1[11]、OsPti1a [12]、OsABA2[13]等。另一类基因, 包括FGL、SL、RLIN1等, 与叶绿素或其他细胞色素合成有关[14,15,16], OsHPL3参与脂氧物代谢过程[17], SPL29可能与糖类代谢有关[18]。此外, 细胞程序性死亡、热击蛋白、ROS相关基因的突变, 同样会导致水稻植株出现lmm表型。如SPL28[19]、SPL5[20]、LSD1[21]、NOE1[22]、SPL7[23]、SPL11[24]、LMR[25]、OsACD1[26]等。
众多水稻类病斑突变体中, 仅有少数伴随早衰现象。水稻类病斑早衰突变体既能自发形成坏死病斑, 又伴有叶早衰现象, 故研究该类突变体对了解叶片衰老过程中的各种生理变化及其影响因素、解析植物抗病机制、研究细胞程序性死亡及植物防御反应相关历程具有十分重要的意义。本研究在籼型恢复系缙恢10号的EMS诱变体库中, 鉴定到一个类病斑早衰突变体lmps1 (lesion mimic and premature senescence 1)。其前期表型正常, 从分蘖早期叶片开始出现褐色小斑点, 并随植株生长而数目增多直至布满叶片。至孕穗期植株叶片开始萎黄, 表现出早衰特征。本研究对该突变体进行表型鉴定、光合特性、超微结构、生理学特性、遗传特性、基因定位等研究, 为目标基因的克隆及后续的分子机制解析等奠定良好基础。
1 材料与方法
1.1 研究材料及其表型鉴定
类病斑早衰突变体lmps1经多代自交, 其表型稳定遗传。将缙恢10号作为该突变体的野生型(WT)参照, 自然条件下, 将WT与lmps1种植田间, 比较其全生育期植株表型。成熟期分别随机选取WT和lmps1植株各10株, 分别考察其株高、穗长、有效穗数、每穗总粒数、每穗实粒数、结实率、千粒重。1.2 遮光处理及光合色素含量与光合效率参数分析
在分蘖期, 用锡箔纸包裹田间尚未发生病斑的突变体lmps1叶片, 连续遮光1周后恢复光照, 期间持续观察叶片病斑的发生情况。在孕穗期用Beckman22S型分光光度计测定光合色素含量, 参照Wellburn[27]描述的方法, 于田间利用便携式光合测定仪Li-6400测定净光合速率(Pn, μmol m-2 s-1)、气孔导度(Gs, mmol m-2 s-1)、胞间CO2浓度(Ci, μmol mol-1)与蒸腾速率(Tr, mmol m-2 s-1)等光合参数。1.3 透射电镜观察及生理指标测定
参照Fang等[28]的方法, 用透射电子显微镜观察WT与lmps1叶肉细胞和叶绿体的结构。用相关试剂盒(南京建成科技有限公司)测定孕穗期倒一、倒二、倒三叶的过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)的活性和过氧化氢(H2O2)的含量, 每个样本重复3次, 用DPS软件进行数据分析。1.4 lmps1目标基因的精细定位
分别将西农1A、lmps1及西农1A×lmps1获得的F2群体种植于西南大学水稻研究所歇马试验基地。在分蘖早期选取该群体中叶片上有褐色斑点的突变单株, 采用改良的CTAB法[29]提取亲本和F2群体中突变表型植株与10个正常表型植株的DNA备用。参照刘宝玉等[9]的方法定位lmps1的基因, 所用引物由上海英骏生物技术公司合成。1.5 RNA提取及qRT-PCR
在分蘖期, 用RNA提取试剂盒(北京天根生化科技有限公司)提取野生型和突变体植株叶片总RNA, 以之为模板通过反转录得到相应cDNA (TaKaRa公司生产的PrimeScript 1st Strand cDNA Synthesis Kit)。用ACTIN作为内参基因, 进行相关基因cDNA扩增, 以2-ΔΔCT计算各基因的相对表达量, 对每个样品至少做3个重复。qRT-PCR主要用于定位区间内部分基因和一些防卫反应相关标记基因的表达分析。2 结果与分析
2.1 突变体lmps1的表型特性
与WT相比, 突变体lmps1苗期表型正常, 分蘖早期叶片开始出现褐色小斑点, 随着植株生长, 斑点数目持续增加(图1-A, B)。孕穗期开始出现早衰表型; 至成熟期, 斑点布满所有叶片, 衰老加速, 叶片严重黄化, 整个植株呈黄褐色(图1-C, D)。突变体lmps1的类病斑与早衰表型, 最早出现的时期不同, 但均由叶尖发展到叶片中部和基部, 并随叶龄增长而程度加深(图1-A, B, C, D)。成熟期突变体的株高、穗长、有效穗数、每穗总粒数、每穗实粒数、结实率和千粒重较野生型均显著或极显著降低(表1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1野生型(WT)与突变体lmps1的表型
A: 分蘖期WT与lmps1植株; B: 分蘖期WT与lmps1的叶片; C: 成熟期WT与lmps1植株; D: 成熟期WT与lmps1的叶片; 1: 倒一叶; 2: 倒二叶; 3: 倒三叶; 标尺=5 cm。
Fig. 1Phenotypes of the wild type (WT) and the lmps1 mutant
A: plants of the wild type and the lmps1 mutant at tillering stage; B: leaves of the wild type and the lmps1 mutant at tillering stage; C: plants of the wild type and the lmps1 mutant at mature period; D: leaves of the wild type and the lmps1 mutant at mature period; 1: the flag leaves; 2: the second leaves; 3: the third leaves; bar = 5 cm.
Table 1
表1
表1野生型与突变体lmps1之间的表型差异
Table 1
| 性状 Trait | 野生型 Wild type | 突变体 lmps1 | lmps1较WT减少率 Reduction in lmps1 than in WT (%) |
|---|---|---|---|
| 株高 Plant height (cm) | 108.50±2.29 | 93.00±2.00** | 14.3 |
| 穗长 Panicle length (cm) | 26.15±0.55 | 19.80±1.03** | 24.3 |
| 有效穗数 Effective panicle | 9.62±1.31 | 7.00±1.07** | 27.2 |
| 每穗总粒数 Grain number per panicle | 161.34±6.78 | 148.42±13.40* | 8.0 |
| 每穗实粒数 Filled grain number per panicle | 133.48±6.36 | 66.27±7.66** | 50.0 |
| 结实率 Seed setting rate (%) | 82.75±2.51 | 44.97±5.95** | 45.7 |
| 千粒重 1000-grain weight (g) | 25.74±0.44 | 22.01±0.43** | 14.5 |
新窗口打开|下载CSV
2.2 突变体lmps1对光的响应
分蘖期对突变体lmps1即将出现但尚未有类病斑表型的叶片进行遮光处理表明, 未遮光部位逐渐产生大量褐色小斑点, 而锡箔纸遮光部位没有斑点或仅产生少量斑点, 且叶片呈绿色, 与野生型基本一致(图2-A, C)。移去锡箔纸恢复光照后, 该部位又开始逐渐变黄, 并出现褐色小斑点(图2-D)。说明突变体lmps1表型受光诱导。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2遮光对野生型(WT)和突变体lmps1叶片的影响
A: WT叶片; B: lmps1叶片; C: lmps1遮光1周后; D: lmps1遮光处理后复光1周后; 标尺=6 cm。
Fig. 2Effects of shading on the wild type (WT) and the lmps1 mutant leaves
A: leaf of the wild type; B: leaf of the lmps1 mutant; C: leaf of the lmps1 mutant after one week shading; D: leaf of the lmps1 mutant illuminated for one week after shading; bar = 6 cm.
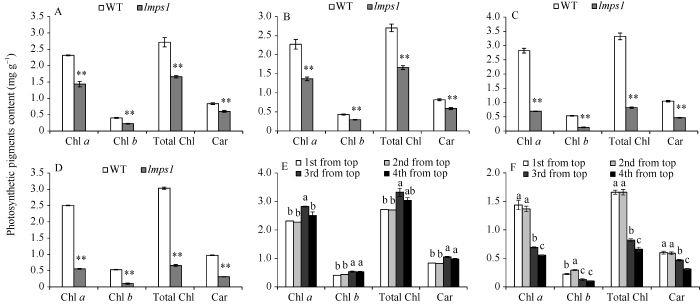
2.3 光合色素含量分析
孕穗期突变体lmps1光合色素含量均低于WT, 差异达到极显著水平(P<0.01)(图3-A, B, C, D), 并且突变体lmps1的倒三、倒四叶光合色素含量明显低于倒一叶、倒二叶, 其中倒四叶最低(图3-E, F)。表明在孕穗期突变体lmps1叶片已在生理水平上进入衰老阶段, 且衰老是从倒四叶向倒一叶发展。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3野生型(WT)与突变体lmps1孕穗期光合色素含量
A~D: 孕穗期WT与lmps1的倒一叶(A)、倒二叶(B)、倒三叶(C)和倒四叶(D)光合色素含量; E: WT光合色素含量; F: lmps1光合色素含量。条柱表示平均值, 误差线表示标准差; **表示在0.01水平上差异显著; E, F图中标的不同字母的柱值在0.05水平上差异显著。
Fig. 3Photosynthetic pigments contents of the wild type (WT) and the lmps1 mutant at booting stage
A-D: photosynthetic pigments contents of the flag leaves, the second leaves, the third leaves and the fourth leaves respectively in the wild type and the lmps1 mutant at booting stage; E: photosynthetic pigments contents of the wild type; F: photosynthetic pigments contents of the lmps1 mutant. The column shows the mean value and the error line shows the standard deviation; ** means significant difference at P < 0.01 by t-test; in Figs. E and F, bars superscripted by different letter are significantly different at P < 0.05.
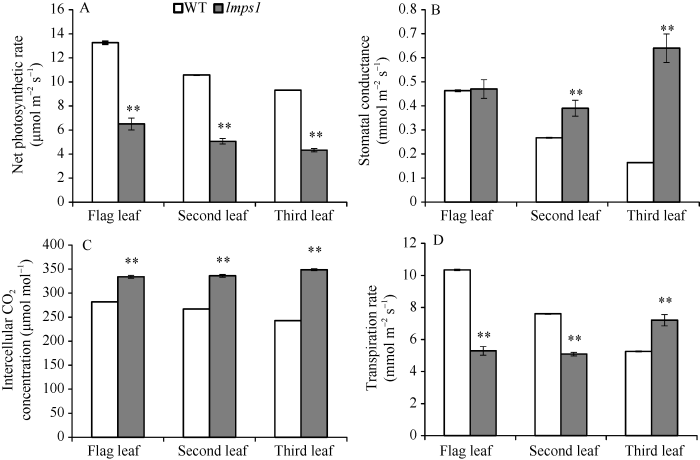
2.4 光合效率变化
孕穗期, 突变体lmps1除倒一叶的气孔导度(Gs)外, 其他光合效率参数与WT差异均极显著: lmps1的净光合速率(Pn)较WT极显著降低(P<0.01); 胞间CO2浓度(Ci)均极显著升高; 气孔导度(Gs)倒二叶、倒三叶极显著升高; lmps1的蒸腾速率(Tr)倒一叶、倒二叶均极显著降低, 而倒三叶则极显著升高(图4-A, B, C, D)。光合效率显著降低表明突变体lmps1光合能力受到严重影响。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4孕穗期野生型(WT)与突变体lmps1功能叶的光合特性
A: WT和lmps1的净光合速率; B: WT和lmps1的气孔导度; C: WT和lmps1的细胞间CO2浓度; D: WT和lmps1的蒸腾速率。条柱表示平均值, 误差线表示标准差; **表示在0.01水平上差异显著, *表示在0.05水平上差异显著。
Fig. 4Photosynthetic characteristics of the wild type (WT) and the lmps1 mutant at booting stage
A: net photosynthetic rate of the wild type and the lmps1 mutant; B: stomatal conductance of the wild type and the lmps1 mutant; C: intercellular CO2 concentration of the wild type and the lmps1 mutant; D: transpiration rate of the wild type and the lmps1 mutant. The column shows the mean value and the error line shows the standard deviation; ** means significant difference at P < 0.01 by t-test, * means significant difference at P < 0.05 by t-test.
2.5 细胞超微结构分析
利用透射电镜观察孕穗期WT和突变体lmps1倒四叶叶尖和叶中部的细胞超微结构表明, 突变体lmps1的叶绿体结构较WT出现明显变化: 细胞内叶绿体数目明显减少(图5-D, J), 嗜锇小体的数量和体积增加(图5-E, F, K, L)。叶尖部分存在结构完好的叶绿体, 但部分叶绿体严重受损, 类囊体片层结构损伤解体(图5-E, F); 叶片中部叶绿体基本完好, 基质片层结构相对叶尖部分而言较为完整, 但结构松散(图5-K, L)。表明突变体lmps1叶绿体结构损伤, 叶绿体开始降解, 致使其叶绿素含量和净光合速率Pn显著降低; 其叶绿体的完整程度与衰老进程一致。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5孕穗期野生型(WT)和突变体lmps1的叶肉细胞超微结构
A: WT倒四叶尖; B、C、D: WT叶尖细胞超微结构; E: lmps1倒四叶尖; F、G、H: lmps1叶尖细胞超微结构; I: WT倒四叶中部; J、K、L: WT叶中部细胞超微结构; M: lmps1倒四叶中部; N、O、P: lmps1叶中部细胞超微结构; Chl: 叶绿体; Os: 嗜锇小体; TM: 类囊体膜; N: 细胞核。标尺: 2 μm (A, D), 1 μm (B, E), 200 nm (C, F)。
Fig. 5Ultrastructure of mesophyll cells in the wild type (WT) and the lmps1 mutant at booting stage
A: tip of the fourth leaves of the wild type; B, C, D: ultrastructure of the blade tip of the wild type; E: tip of the fourth leaves of the lmps1 mutant; F, G, H: ultrastructure of the blade tip of the lmps1 mutant; I: middle part of the fourth leaves of the wild type; J, K, L: ultrastructure of the middle part of blade of the wild type; M: middle part of the fourth leaves of the lmps1 mutant; N, O, P: ultrastructure of the middle part of blade of the lmps1 mutant. Chl: chloroplast; Os: osmiophilic granule; TM: thylakoid membranes; N: ribosome. Bars: 2 μm (A, D), 1 μm (B, E), 200 nm (C, F).
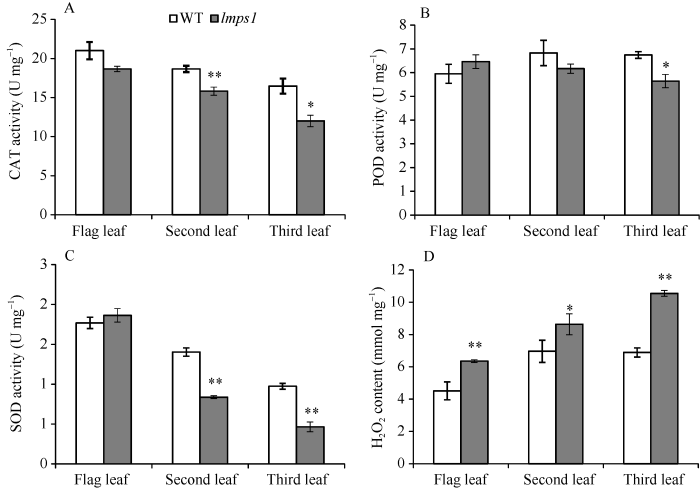
2.6 抗氧化酶活性及H2O2含量分析
在孕穗期, 除lmps1倒一叶外, lmps1倒二、倒三叶的CAT、SOD活性较WT均显著或极显著降低(图6-A, C)。lmps1的POD活性除倒三叶较WT显著降低外, 其倒一、倒二叶差异不显著(图6-B)。而lmps1的H2O2含量较WT均显著或极显著增加(图6-D)。抗氧化酶活性降低和活性氧H2O2的增加表明lmps1叶片发生衰老。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6孕穗期野生型(WT)和突变体lmps1的抗氧化酶活性及H2O2含量
A~C: WT和lmps1的过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)活性; D: WT和lmps1各功能叶过氧化氢(H2O2)含量。条柱表示平均值, 误差线表示标准差; **表示在0.01水平上差异显著, *表示在0.05水平上差异显著。
Fig. 6Antioxidant enzyme activity and the H2O2 content in the wild type (WT) and the lmps1 mutant at the booting stage
A-C: the enzyme activity of catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD) in functional leaves of the wild type and the lmps1 mutant; D: H2O2 content of different functional leaves of the wild type and the lmps1 mutant. The column shows the mean value and the error line shows the standard deviation; ** means significant difference at P < 0.01 by t-test, * means significant difference at P < 0.05 by t-test.
2.7 遗传分析与目标基因的定位
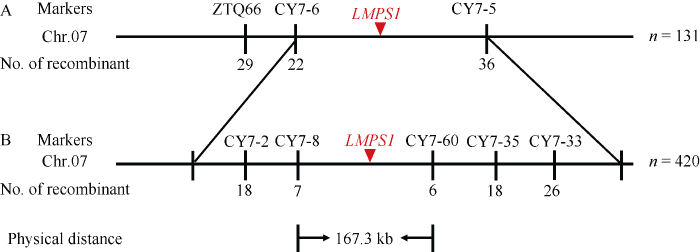
用籼稻西农1A与突变体lmps1构建的F1单株表型均与WT一致, F2代发生性状分离。共收获正常单株1214株、类病斑早衰突变单株420株, 其比例为2.89∶1。经卡平方测验, 符合3∶1的理论分离比例(χ2 = 0.3949, P = 0.5211), 表明突变体lmps1的类病斑早衰性状受1对隐性核基因控制。选取西南大学水稻研究所现存均匀分布于水稻12条染色体上的400个SSR和InDel标记, 在西农1A和突变体lmps1之间进行多态性筛选。用筛选到的106对多态性引物扩增正常叶片和类病斑早衰叶片基因池, 发现位于水稻第7染色体的InDel标记ZTQ66在两组基因池之间表现出差异, 利用F2群体中正常单株10株和类病斑早衰单株21 株进行单株验证, 初步确定ZTQ66与目标基因连锁。进一步在该标记上下游开发新的多态性标记, 初步将目标基因定位于第7染色体CY7-6和CY7-5之间(图7-A)。利用CY7-6和CY7-5之间的多态性标记, 最终将LMPS1基因定位在标记CY7-8和CY7-60之间, 遗传距离分别为0.83 cM和0.71 cM, 物理距离约为167.3 kb (图7-B)。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7LMPS1基因在第7染色体的分子定位
A: LMPS1基于F2群体的初定位; B: LMPS1的精细定位。
Fig. 7Molecular mapping of LMPS1 on chromosome 7
A: QTL mapping of LMPS1; B: fine mapping of LMPS1.
2.8 定位区间内基因及防卫反应相关基因表达量分析
根据Rice Genome Annotation Project网站上的信息预测, 定位区间内共25个注释基因。根据遗传距离挑选部分基因并利用网站提供序列信息设计qRT-PCR引物, 对部分基因进行表达分析发现, 与野生型相比, LOC_Os07g46390、LOC_Os07g46430、LOC_Os07g 46490 表达量没有明显变化, LOC_Os07g46410上调接近1.5倍, LOC_ Os07g46460上调2倍, 其他LOC_ Os07g46420、LOC_Os07g46440、LOC_Os07g46450分别下调37.3%、76.6%和44.4%, LOC_Os07g46480下调近20倍(图8-A)。图8
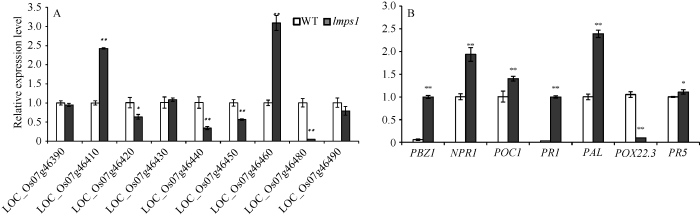 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8野生型(WT)和突变体lmps1相关基因的表达量
A: LMPS1精细定位物理区间内部分基因的表达量; B: 抗病相关基因相对表达量。条柱表示平均值, 误差线表示标准差; *表示0.05水平上差异显著; **表示在0.01水平上差异显著。
Fig. 8Relative expression level of genes in the wild type (WT) and the lmps1 mutant
A: relative expression level of annotated genes within the physical fine mapping region of LMPS1; B: relative expression level of genes related to disease resistance. The column shows the mean value and the error line shows the standard deviation; * refers to the significant difference at P < 0.05; ** denotes significant difference at P < 0.01.
类病斑突变体中常伴随防卫反应相关基因表达的变化, 这可能是类病斑突变体对病原菌抗性增强的原因之一。图8-B表明, lmps1除POX22.3相对WT下调9倍外, POC1、PAL、PBZ1、PR1、NPR1、PR5均为上调。POC1和PR5分别上调28.2%和9.8%, PAL上调约1.4倍、PBZ1上调15.8倍、PR1上调27倍、NPR1上调接近1倍。说明lmps1的突变可能激活部分病程相关因子, 引起水稻防卫反应。
3 讨论
目前, 通过图位克隆或其他手段, 已有多个控制水稻类病斑形成的基因被定位或克隆。已报道的类病斑突变体中, 仅有少数伴随叶片早衰现象, 包括已克隆的SPL28[19]、SPL29[18]、OsLMS[30]基因对应的突变体, 它们分别被定位在第1、第8和第2染色体。未克隆的lmes1[31]、lmes2[32]、lms1[33]等被定位在第7、第10和第11染色体。SPL28编码一个网格蛋白相关衔接蛋白复合物1, 其突变体苗期表型正常, 分蘖初期开始出现红褐色小斑点, 抽穗期斑点面积达到最大, 开花早期叶片开始黄化, 开花后期叶片枯萎, 成熟期植株死亡, 该突变体的籽粒干瘪, 产量严重下降[19]; SPL29编码UDP-N-乙酰葡萄糖胺焦磷酸化酶, 在种子萌发后23 d左右开始出现黑褐色类病斑, 30 d左右开始出现叶片早衰现象, 主要农艺性状指标均严重低于野生型[18]; OsLMS编码一个羧基末端磷酸酶结构域及2个双链的RNA结合区, 相应突变体在播种30~40 d左右出现红棕色类坏死斑点, 开花后叶片早衰, 籽粒形态与野生型类似[30]。LMES1和LMES2尚未被克隆, 前者分蘖早期出现褐色类病斑, 后者五叶期自发出现坏死斑, 都在孕穗期出现叶片黄化衰老[31,32]。本研究中的突变体lmps1, 从分蘖早期出现褐色类病斑, 之后类病斑数量逐渐增加, 直至整个植株叶片被斑点覆盖。至孕穗期lmps1开始衰老直至整个植株呈黄褐色, 此时野生型植株呈绿色。对lmps1进行相应的生理指标测定, 与野生型相比, lmps1的光合色素含量极显著降低, 光合效率显著降低, 抗氧化酶活性降低, 活性氧H2O2显著增加。此外, 突变体lmps1较野生型, 成熟期每穗总粒数显著降低而株高、穗长、有效穗数、每穗实粒数、结实率与千粒重极显著降低。类病斑突变体中常伴随防御反应相关基因表达的变化, 这可能是类病斑突变体对病原菌抗性增强原因之一。本研究中, 除POX22.3下调9倍外, 其余POC1、PAL、PBZ1、PR1、NPR1、PR5表达量均高于野生型。POX22.3为PR9家族基因, 编码过氧化物酶, 与H2O2代谢相关, 突变体lmps1的POX22.3表达量的下降符合叶片H2O2含量的变化[26]; POC1编码过氧化物酶[36]; PR5编码的蛋白属类甜蛋白质, 类甜蛋白质具有抗真菌活性[26]; PAL编码的苯丙氨酸解氨酶, 是植物防卫反应的一个关键蛋白, 涉及SA信号途径[36]; PBZ1为PR10家族基因, 编码一种噻菌灵诱导蛋白; PR1编码的蛋白属于类丝氨酸羧肽酶, 常被看作水稻抗病反应的标记基因[37]; NPR1为系统获得抗性中SA信号途径的标记[10]。结果表明, lmps1的突变可能激活部分病程相关因子, 引起水稻防卫反应。
目前在第7染色体已报道的类病斑突变体有spl5、lmes1。SPL5基因已被克隆, 定位在第7染色体的短臂, 与lmps1定位区域相距较远, 且表型不同, 即LMPS1基因与SPL5不是同一基因[20]; lmps1目标基因定位区间包含了已报道类病斑早衰突变体lmes1的定位区间, 两者表型极为相似, 细微区别之处在于lmes1 单株有效穗数以及每穗总粒数没有差异或差异不明显, 而lmps1表现出极显著的差异[31], 鉴于lmps1与lmes1的目的基因均尚不确定, 两者是否由同一基因突变有待进一步研究。
4 结论
lmps1是EMS诱变得到的类病斑早衰突变体, 早期表型正常, 分蘖早期出现病斑, 孕穗期开始衰老, 斑点数目随植株生长而增多。植株变矮, 结实率降低, 千粒重下降。叶片中光合色素含量降低, 细胞内叶绿体数目减少, 叶绿体的类囊体片层结构损伤降解。光合色素的减少和叶绿体的降解引起光合效率降低, 进而引起突变体lmps1植株表型的相应变化。突变体lmps1类病斑的产生受光照诱导, 并受ROS胁迫, 其突变能激活部分病程相关因子。该性状受1对隐性核基因控制, 被定位于第7染色体长臂端粒附近的CY7-8和CY7-60两个分子标记之间, 物理距离约为167.3 kb。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1002/bies.950170805URL [本文引用: 2]

A class of maize mutants, collectively known as disease lesion mimics, display discrete disease-like symptoms in the absence of pathogens. It is intriguing that a majority of these lesion mimics behave as dominant gain-of-function mutations. The production of lesions is strongly influenced by light, temperature, developmental state and genetic background. Presently, the biological significance of this lesion mimicry is not clear, although suggestions have been made that they may represent defects in the plants' recognition of, or response to, pathogens. One feature that is common to all lesion mimics is their association with cell death. In plants, as in animals, a number of developmental and pathological processes exist where controlled cell death, whether programmed or triggered in response to physiological or environmental stimuli, constitutes the normal aspect of life. Might disease lesion mimic mutations represent variants where regulation of desirable cell death has gone awry? In this paper we argue that this might be the case, and further conjecture that these mutants offer a unique opportunity for studying the genetic and cellular mechanisms of cell death in plants.
DOI:10.2307/3869149URL [本文引用: 1]
Sections of the elongation zones of coleoptiles of Zea mays L. were incubated in buffers containing various concentrations of human epidermal growth factor (EGF). The growth rate of sections incubated with 10, 1.0 or 0.1-mu-g liter-1 EGF was higher than that of control. EGF at 10-mu-g liter-1 increased the growth rate by 73% of the control rate.
[本文引用: 1]
DOI:10.1101/gad.218202URL [本文引用: 1]
DOI:10.1534/genetics.109.102871URLPMID:19506306 [本文引用: 1]
Abstract Rice blast, caused by Magnaporthe oryzae, is one of the most devastating diseases. The two major subspecies of Asian cultivated rice (Oryza sativa L.), indica and japonica, have shown obvious differences in rice blast resistance, but the genomic basis that underlies the difference is not clear. We performed a genomewide comparison of the major class of resistant gene family, the nucleotide-binding site-leucine-rich repeat (NBS-LRR) gene family, between 93-11 (indica) and Nipponbare (japonica) with a focus on their pseudogene members. We found great differences in either constitution or distribution of pseudogenes between the two genomes. According to this comparison, we designed the PCR-based molecular markers specific to the Nipponbare NBS-LRR pseudogene alleles and used them as cosegregation markers for blast susceptibility in a segregation population from a cross between a rice blast-resistant indica variety and a susceptible japonica variety. Through this approach, we identified a new blast resistance gene, Pid3, in the indica variety, Digu. The allelic Pid3 loci in most of the tested japonica varieties were identified as pseudogenes due to a nonsense mutation at the nucleotide position 2208 starting from the translation initiation site. However, this mutation was not found in any of the tested indica varieties, African cultivated rice varieties, or AA genome-containing wild rice species. These results suggest that the pseudogenization of Pid3 in japonica occurred after the divergence of indica and japonica.
DOI:10.1007/s11103-006-9130-yURLPMID:17273822 [本文引用: 2]
A lesion mimic mutant that we designated Spotted leaf 18 ( Spl18 ) was isolated from 13,000 activation-tagging lines of rice produced by our modified activation-tagging vector and further characterized. Spl18 was dominant and its phenotype was linked to the T-DNA insertion. An ORF was located about 500 bp downstream of the inserted T-DNA, and the deduced protein, designated OsAT1, showed sequence similarity to an acyltransferase whose expression is induced by hypersensitive reaction in tobacco. The transcriptional level of OsAT1 was very low in the WT leaf blade but high in Spl18 leaf blade. In wild-type rice, OsAT1 was transcribed mainly in the young panicle, in the panicle just after heading, and in the leaf sheath. In addition, transcription of the genes for PR protein was upregulated in Spl18 , accumulation of phytoalexins (both momilactone A and sakuranetin) was increased, and resistance to blast disease was improved. We then combined OsAT1 genomic DNA downstream of the modified 35S promoter and re-transformed it into rice. Lesion mimic and blast resistance phenotypes were detected in the transgenic lines produced, clearly indicating that overexpression of OsAT1 caused the Spl18 phenotypes. In addition, plants overexpressing OsAT1 showed resistance to bacterial blight.
DOI:10.1093/jhered/93.1.50URLPMID:12011176 [本文引用: 1]
Abstract In groundnut, two identical mutants with disease lesion mimic leaf trait were isolated independently from two different parents through induced mutagenesis and in vitro culture technique. The leaf chlorophyll content in both the mutants was found to be drastically reduced. The segregation pattern in the F(2) and F(3) generations for normal and mutant traits fitted a 13:3 ratio, indicating that the disease lesion mimic trait in the mutants was due to suppressive gene action. Both mutants were allelic for the disease mimic trait.
[本文引用: 1]
URL [本文引用: 2]
利用化学诱变剂EMS处理籼型水稻恢复系"缙恢10号",从其后代中筛选到1个遗传稳定的类病斑突变体spl34。该突变体于分蘖后期在下部叶片的叶鞘上开始出现褐色的类病斑,随后沿着中脉扩散至整个叶片,成熟期扩散至整个植株。相比于野生型,该突变体的株高显著变矮,穗长显著变短,穗粒数、结实率和千粒重极显著降低。遮光试验和组织化学分析表明,突变体类病斑的形成受光诱导,在类病斑形成部位发生大量过氧化氢沉积和细胞程序性死亡。荧光显微镜观察发现,在紫外光照射下突变体产生的荧光较野生型弱。与野生型相比,突变体spl34的H_2O_2和O_2~-含量较高,而CAT、POD和T-SOD等保护酶的活性显著降低;稻瘟病抗性无明显差异或略显降低。遗传分析表明,突变体spl34的表型受1对隐性核基因控制。基因定位结果表明,该基因定位于第4染色体的LR49和LR52两个分子标记之间,物理距离为200 kb。测序分析发现该区间内的候选基因LOC_Os04g56480的第3449位碱基发生突变(G3449T),导致色氨酸替换为半胱氨酸。qRT-PCR结果表明该基因在突变体内表达量降低,而部分病程相关基因的表达量则升高。
URL [本文引用: 2]
利用化学诱变剂EMS处理籼型水稻恢复系"缙恢10号",从其后代中筛选到1个遗传稳定的类病斑突变体spl34。该突变体于分蘖后期在下部叶片的叶鞘上开始出现褐色的类病斑,随后沿着中脉扩散至整个叶片,成熟期扩散至整个植株。相比于野生型,该突变体的株高显著变矮,穗长显著变短,穗粒数、结实率和千粒重极显著降低。遮光试验和组织化学分析表明,突变体类病斑的形成受光诱导,在类病斑形成部位发生大量过氧化氢沉积和细胞程序性死亡。荧光显微镜观察发现,在紫外光照射下突变体产生的荧光较野生型弱。与野生型相比,突变体spl34的H_2O_2和O_2~-含量较高,而CAT、POD和T-SOD等保护酶的活性显著降低;稻瘟病抗性无明显差异或略显降低。遗传分析表明,突变体spl34的表型受1对隐性核基因控制。基因定位结果表明,该基因定位于第4染色体的LR49和LR52两个分子标记之间,物理距离为200 kb。测序分析发现该区间内的候选基因LOC_Os04g56480的第3449位碱基发生突变(G3449T),导致色氨酸替换为半胱氨酸。qRT-PCR结果表明该基因在突变体内表达量降低,而部分病程相关基因的表达量则升高。
DOI:10.1094/MPMI-18-0511URL [本文引用: 2]
DOI:10.1111/j.1365-313X.2011.04557.xURLPMID:21418352 [本文引用: 1]
In this study, we characterized the semi-dominant mutant nls1-1D (necrotic leaf sheath 1) of rice, which displays spontaneous lesions, specifically on leaf sheaths, with a developmental pattern. nls1-1D plants also exhibited constitutively activated defense responses, including extensive cell death, excess hydrogen peroxide and salicylic acid (SA) accumulation, up-regulated expressions of pathogenesis-related genes, and enhanced resistance to bacterial pathogens. Map-based cloning revealed that NLS1 encodes a typical CC-NB-LRR-type protein in rice. The nls1-1D mutation causes a S367N substitution in the non-conserved region close to the GLPL motif of the NB domain. An adjacent S366T substitution was found in another semi-dominant mutant, nls1-2D, which exhibited the same phenotypes as nls1-1D. Combined analyses of wild-type plants transformed with the mutant NLS1 gene (nls1-1D), NLS1 RNAi and over-expression transgenic lines showed that nls1-2D is allelic to nls1-1D, and both mutations may cause constitutive auto-activation of the NLS1 R protein. Further real-time PCR analysis revealed that NLS1 is expressed constitutively in an age-dependent manner. In addition, because the morphology and constitutive defense responses of nls1-1D were not suppressed by blocking SA or NPR1 transcript accumulation, we suggest that NLS1 mediates both SA and NPR1-independent defense signaling pathways in rice.
[本文引用: 1]
DOI:10.3389/fpls.2018.00405URLPMID:29643863 [本文引用: 1]
Abstract Lesion mimic mutants display spontaneous cell death, and thus are valuable for understanding the molecular mechanism of cell death and disease resistance. Although a lot of such mutants have been characterized in rice, the relationship between lesion formation and abscisic acid (ABA) synthesis pathway is not reported. In the present study, we identified a rice mutant, lesion mimic mutant 9150 ( lmm9150 ), exhibiting spontaneous cell death, pre-harvest sprouting, enhanced growth, and resistance to rice bacterial and blast diseases. Cell death in the mutant was accompanied with excessive accumulation of H 2 O 2 . Enhanced disease resistance was associated with cell death and upregulation of defense-related genes. Map-based cloning identified a G-to-A point mutation resulting in a D-to-N substitution at the amino acid position 110 of OsABA2 (LOC_Os03g59610) in lmm9150 . Knock-out of OsABA2 through CRISPR/Cas9 led to phenotypes similar to those of lmm9150 . Consistent with the function of OsABA2 in ABA biosynthesis, ABA level in the lmm9150 mutant was significantly reduced. Moreover, exogenous application of ABA could rescue all the mutant phenotypes of lmm9150 . Taken together, our data linked ABA deficiency to cell death and provided insight into the role of ABA in rice disease resistance.
[本文引用: 1]
DOI:10.1074/jbc.M109.091371URLPMID:2857009 [本文引用: 1]
Serotonin is a well known neurotransmitter in mammals and plays an important role in various mental functions in humans. In plants, the serotonin biosynthesis pathway and its function are not well understood. The rice sekiguchi lesion (sl) mutants accumulate tryptamine, a candidate substrate for serotonin biosynthesis. We isolated the SL gene by map-based cloning and found that it encodes CYP71P1 in a cytochrome P450 monooxygenase family. A recombinant SL protein exhibited tryptamine 5-hydroxylase enzyme activity and catalyzed the conversion of tryptamine to serotonin. This pathway is novel and has not been reported in mammals. Expression of SL was induced by the N-acetylchitooligosaccharide (chitin) elicitor and by infection with Magnaporthe grisea, a causal agent for rice blast disease. Exogenously applied serotonin induced defense gene expression and cell death in rice suspension cultures and increased resistance to rice blast infection in plants. We also found that serotonin-induced defense gene expression is mediated by the RacGTPase pathway and by the G alpha subunit of the heterotrimeric G protein. These results suggest that serotonin plays an important role in rice innate immunity.
[本文引用: 1]
DOI:10.1371/journal.pone.0050089URLPMID:3510209 [本文引用: 1]
The allene oxide synthase (AOS) and hydroperoxide lyase (HPL) branches of the oxylipin pathway, which underlie the production of jasmonates and aldehydes, respectively, function in plant responses to a range of stresses. Regulatory crosstalk has been proposed to exist between these two signaling branches; however, there is no direct evidence of this. Here, we identified and characterized a jasmonic acid (JA) overproduction mutant,cea62, by screening a rice T-DNA insertion mutant library for lineages that constitutively express the AOS gene. Map-based cloning was used to identify the underlying gene as hydroperoxide lyaseOsHPL3.HPL3expression and the enzyme activity of its product, (E)-2-hexenal, were depleted in thecea62mutant, which resulted in the dramatic overproduction of JA, the activation of JA signaling, and the emergence of the lesion mimic phenotype. A time-course analysis of lesion formation and of the induction of defense responsive genes in thecea62mutant revealed that the activation of JA biosynthesis and signaling incea62was regulated in a developmental manner, as was OsHPL3 activity in the wild-type plant. Microarray analysis showed that the JA-governed defense response was greatly activated incea62and this plant exhibited enhanced resistance to the T1 strain of the bacterial blight pathogenXanthomonasoryzaepvoryzae(Xoo). The wounding response was attenuated incea62plants during the early stages of development, but partially recovered when JA levels were elevated during the later stages. In contrast, the wounding response was not altered during the different developmental stages of wild-type plants. These findings suggest that these two branches of the oxylipin pathway exhibit crosstalk with regards to biosynthesis and signaling and cooperate with each other to function in diverse stress responses.
DOI:10.1093/jxb/eru456URLPMID:4321554 [本文引用: 3]
This study identified the novel gene UAP1 in rice. UAP1 is involved in early leaf senescence and defence responses. Plant leaf senescence and defence responses are important biological processes, but the molecular mechanisms involved are not well understood. This study identified a new rice mutant, spotted leaf 29 (spl29). The SPL29 gene was identified by map-based cloning, and SPL29 was confirmed as UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) by enzymatic analysis. The mutant spl29 lacks UAP activity. The biological phenotypes for which UAP is responsible have not previously been reported in plants. The spl29 mutant displayed early leaf senescence, confirmed by chlorophyll loss and photosystem II decline as physiological indicators, chloroplast degradation as a cellular characteristic, and both upregulation of senescence transcription factors and senescence-associated genes, and downregulation of photosynthesis-related genes, as molecular evidence. Defence responses were induced in the spl29 mutant, shown by enhanced resistance to bacterial blight inoculation and upregulation of defence response genes. Reactive oxygen species, including O2 and H2O2, accumulated in spl29 plants; there was also increased malondialdehyde content. Enhanced superoxide dismutase activity combined with normal catalase activity in spl29 could be responsible for H2O2 accumulation. The plant hormones jasmonic acid and abscisic acid also accumulated in spl29 plants. ROS and plant hormones probably play important roles in early leaf senescence and defence responses in the spl29 mutant. Based on these findings, it is suggested that UAP1 is involved in regulating leaf senescence and defence responses in rice.
[本文引用: 3]
DOI:10.1186/s12284-015-0052-7URLPMID:26029330 [本文引用: 2]
Rice mutant,spl5(spotted leaf 5), has spontaneous hypersensitive-like lesions on its leaves and shows enhanced resistance to pathogens, indicating thatSPL5plays a role in programmed cell death (PCD) and disease resistance. To understand the molecular mechanism ofSPL5gene, we investigated the transcriptome profiles of thespl5mutant leaves with few lesions (FL) and leaves with many lesions (ML) compared to the wild-type (WT) leaves respectively by microarray. The data from microarray revealed that 243 and 896 candidate genes (Fold change 3.0) were up- or down-regulated in thespl5-FL andspl5-ML, respectively, and a large number of these genes involved in biotic defense responses or reactive oxygen species (ROS) metabolism. Interestingly, according to our microarray and real-time PCR assays, the expressions of a transcription factorOsWRKY14and genes responsible for the biosynthesis of serotonin, anthranilate synthase (AS), indole-3-glycerolphosphate synthase (IGPS), tryptophan synthase (TS) and tryptophan decarboxylase (TDC) were significantly up-regulated in thespl5mutant. It has been reported previously thatTSandTDCexpressions are regulated byOsWRKY14in rice, which raises the possibility thatOsWRKY14regulates serotonin production through the up-regulation ofTSandTDC. Our HPLC analysis further confirmed that serotonin levels were higher in the leaves ofspl5mutant than that in WT. Since the serotonin plays a critical role in inducing disease-resistance, the increased serotonin level may contribute, at least partly, to the disease resistance inspl5. TheSPL5gene may act as a negative regulatory factor activating the serotonin metabolic pathway, and these results might provide a new insight into thespl5-induced defense response mechanisms in plants. The online version of this article (doi:10.1186/s12284-015-0052-7) contains supplementary material, which is available to authorized users.
DOI:10.1094/MPMI-18-0375URLPMID:15915636 [本文引用: 1]
Abstract The Arabidopsis LSD1 and LOL1 proteins both contain three conserved zinc finger domains and have antagonistic effects on plant programmed cell death (PCD). In this study, a rice (Oryza sativa) functional homolog of LSD1, designated OsLSD1, was identified. The expression of OsLSD1 was light-induced or dark-suppressed. Overexpression of OsLSD1 driven by the cauliflower mosaic virus 35S promoter accelerated callus differentiation in transformed rice tissues and increased chlorophyll b content in transgenic rice plants. Antisense transgenic rice plants exhibited lesion mimic phenotype, increased expression of PR-1 mRNA, and an accelerated hypersensitive response when inoculated with avirulent isolates of blast fungus. Both sense and antisense transgenic rice plants conferred significantly enhanced resistance against a virulent isolate of blast fungus. Moreover, ectopic overexpression of OsLSD1 in transgenic tobacco (Nicotiana tabacum) enhanced the tolerance to fumonisins B1 (FB1), a PCD-eliciting toxin. OsLSD1 green fluorescent protein fusion protein was located in the nucleus of tobacco cells. Our results suggest that OsLSD1 plays a negative role in regulating plant PCD, whereas it plays a positive role in callus differentiation.
DOI:10.1104/pp.111.184531URLPMID:22106097 [本文引用: 1]
Nitric oxide (NO) is a key redox-active, small molecule involved in various aspects of plant growth and development. Here, we report the identification of an NO accumulation mutant, nitric oxide excessl (noel), in rice (Oryza sativa), the isolation of the corresponding gene, and the analysis of its role in NO-mediated leaf cell death. Map-based cloning revealed that NOE1 encoded a rice catalase, OsCATC. Furthermore, noe1 resulted in an increase of hydrogen peroxide (H60O60) in the leaves, which consequently promoted NO production via the activation of nitrate reduclase. The removal of excess NO reduced cell death in both leaves and suspension cultures derived from noe1 plants, implicating NO as an important endogenous mediator of H60O60-induced leaf cell death. Reduction of intracellular S-nitrosothiol (SNO) levels, generated by overexpression of rice S-nitrosoglutathione reducíase gene (GSNOR1), which regulates global levels of protein S-nitrosylation, alleviated leaf cell death in noe1 plants. Thus, S-nitrosylation was also involved in light-dependent leaf cell death in noe1. Utilizing the biotin-switch assay, nanoliquid chromatography, and tandem mass spectrometry, S-nitrosylated proteins were identified in both wild-type and noe1 plants. NO targets identified only in noe1 plants included glyceraldehyde 3-phosphate dehydrogenase and thioredoxin, which have been reported to be involved in S-nitrosylation-regulated cell death in animals. Collectively, our data suggest that both NO and SNOs are important mediators in the process of H60O60-induced leaf cell death in rice.
DOI:10.1073/pnas.112209199URLPMID:12032317 [本文引用: 1]
A rice spotted leaf (lesion-mimic) gene, Spl7, was identified by map-based cloning. High-resolution mapping with cleaved amplified polymorphic sequence markers enabled us to define a genomic region of 3 kb as a candidate for Spl7. We found one ORF that showed high similarity to a heat stress transcription factor (HSF). Transgenic analysis verified the function of the candidate gene for Spl7: leaf spot development was suppressed in spl7 mutants with a wild-type Spl7 transgene. Thus, we conclude that Spl7 encodes the HSF protein. The transcript of spl7 was observed in mutant plants. The levels of mRNAs (Spl7 in wild type and spl7 in mutant) increased under heat stress. Sequence analysis revealed only one base substitution in the HSF DNA-binding domain of the mutant allele, causing a change from tryptophan to cysteine.
DOI:10.1105/tpc.104.025171URLPMID:15377756 [本文引用: 1]
The rice (Oryza sativa) spotted leaf11 (spl11) mutant was identified from an ethyl methanesulfonate-mutagenized indica cultivar IR68 population and was previously shown to display a spontaneous cell death phenotype and enhanced resistance to rice fungal and bacterial pathogens. Here, we have isolated Spl11 via a map-based cloning strategy. The isolation of the Spl11 gene was facilitated by the identification of three additional spl11 alleles from an IR64 mutant collection. The predicted SPL11 protein contains both a U-box domain and an armadillo (ARM) repeat domain, which were demonstrated in yeast and mammalian systems to be involved in ubiquitination and protein-protein interactions, respectively. Amino acid sequence comparison indicated that the similarity between SPL11 and other plant U-box-ARM proteins is mostly restricted to the U-box and ARM repeat regions. A single base substitution was detected in spl11, which results in a premature stop codon in the SPL11 protein. Expression analysis indicated that Spl11 is induced in both incompatible and compatible rice-blast interactions. In vitro ubiquitination assay indicated that the SPL11 protein possesses E3 ubiquitin ligase activity that is dependent on an intact U-box domain, suggesting a role of the ubiquitination system in the control of plant cell death and defense.
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
DOI:10.1007/s11434-010-4013-7URL [本文引用: 1]
Leaf senescence as an active process is essential for plant survival and reproduction. However, premature senility is harmful to agricultural production. In this study, a rice mutant, named as psl3 ( presescing leaf 3 ) isolated from EMS-treated Jinhui 10, displays obvious premature senility features both in morphological and physiological level. Genetic analysis showed that mutant trait was controlled by a single dominant gene ( PSL3 ), which was located on rice chromosome 7 between SSR marker c7sr1 and InDel marker ID10 with an interval of 53.5 kb. The result may be useful for the isolation of the PSL3 gene.
DOI:10.1007/BF02172406URLPMID:8914521 [本文引用: 1]
Microsatellite markers containing simple sequence repeats (SSR) are a valuable tool for genetic analysis. Our objective is to augment the existing RFLP map of rice with simple sequence length polymorphisms ( SSLP ). In this study, we describe 20 new microsatellite markers that have been assigned to positions along the rice chromosomes, characterized for their allelic diversity in cultivated and wild rice, and tested for amplification in distantly related species. Our results indicate that the genomic distribution of microsatellites in rice appears to be random, with no obvious bias for, or clustering in particular regions, that mapping results are identical in intersubspecific and interspecific populations, and that amplification in wild relatives of Oryza sativa is reliable in species most closely related to cultivated rice but becomes less successful as the genetic distance increases. Sequence analysis of SSLP alleles in three related indica varieties demonstrated the clustering of complex arrays of SSR motifs in a single 300-bp region with independent variation in each. Two microsatellite markers amplified multiple loci that were mapped onto independent rice chromosomes, suggesting the presence of duplicated regions within the rice genome. The availability of increasing numbers of mapped SSLP markers can be expected to increase the power and resolution of genome analysis in rice.
DOI:10.1266/ggs.87.169URLPMID:22976392 [本文引用: 2]
The rice (Oryza sativa L.) lesion mimic and senescence (lms) EMS-mutant, identified in a japonica cultivar Hitomebore, is characterized by a spontaneous lesion mimic phenotype during its vegetative growth, an accelerated senescence after flowering, and enhanced resistance to rice blast (Magnaporthe oryzae). To isolate the OsLMS gene, we crossed the lms mutant to Kasalath (indica), and used mutant F(2) plants to initially map the candidate region to about 322-kb on the long arm of chromosome 2. Illumina whole-genome re-sequencing of the mutant and aligning the reads to Hitomebore reference sequence within the candidate region delineated by linkage analysis identified a G to A nucleotide substitution. The mutation corresponded to the exon-intron splicing junction of a novel gene that encodes a carboxyl-terminal domain (CTD) phosphatase domain and two double stranded RNA binding motifs (dsRBM) containing protein. By PCR amplification, we confirmed that the mutation causes splicing error that is predicted to introduce a premature stop codon. RNA interference (RNAi) transgenic lines with suppressed expression of LMS gene exhibited the lesion mimic phenotype, confirming that the mutation identified in LMS is responsible for the mutant phenotype. OsLMS shares a moderate amino-acid similarity to the Arabidopsis FIERY2/CPL1 gene, which is known to control many plant processes such as stress response and development. Consistence with this similarity, the lms mutant shows sensitivity to cold stress at the early growth stage, suggesting that LMS is a negative regulator of stress response in rice.
[本文引用: 3]
[本文引用: 2]
DOI:10.3969/j.issn.1008-0384.2014.01.007URL [本文引用: 1]
通过筛选籼稻恢复系明恢86的组培变异后代,获得1个类病斑及早衰突变体 lms1(lesion mimic and senescence 1)。 lms1植株生长至拔节期开始在叶片上出现黄褐色小斑点,随后斑点逐渐扩展至大部分叶片和茎组织;生长至抽穗期后呈现早衰,穗、茎、叶明显干枯,并快速衰亡。RT-PCR分析表明,在呈现类病斑性状叶片组织的 lsm1中,病程相关基因 PBZ1、 PA L1表达明显高于其在野生型叶片组织中的水平。遗传分析表明, lms1的突变性状受单隐性核基因控制。利用9311与 lms1配置的F2及F3群体进行基因定位,将 lms1基因定位在水稻第11染色体长臂末端。
DOI:10.3969/j.issn.1008-0384.2014.01.007URL [本文引用: 1]
通过筛选籼稻恢复系明恢86的组培变异后代,获得1个类病斑及早衰突变体 lms1(lesion mimic and senescence 1)。 lms1植株生长至拔节期开始在叶片上出现黄褐色小斑点,随后斑点逐渐扩展至大部分叶片和茎组织;生长至抽穗期后呈现早衰,穗、茎、叶明显干枯,并快速衰亡。RT-PCR分析表明,在呈现类病斑性状叶片组织的 lsm1中,病程相关基因 PBZ1、 PA L1表达明显高于其在野生型叶片组织中的水平。遗传分析表明, lms1的突变性状受单隐性核基因控制。利用9311与 lms1配置的F2及F3群体进行基因定位,将 lms1基因定位在水稻第11染色体长臂末端。
DOI:10.1016/j.plaphy.2006.06.010URLPMID:16806959
Rice, a first cereal crop whose draft genome sequence from two subspecies (japonica-type cv. Nipponbare and indica-type 93-11) was available in 2002, along with its almost complete genome sequence in 2005, has drawn the attention of researchers worldwide because of its immense impact on human existence. One of the most critical research areas in rice is to discern the self-defense mechanism(s), an innate property of all living organisms. The last few decades have seen scattered research into rice responses to diverse environmental stimuli and stress factors. Our understanding on rice self-defense mechanism has increased considerably with accelerated research during recent years mainly due to identification and characterization of several defense/stress-related components, genes, proteins and secondary metabolites. As these identified components have been used to study the defense/stress pathways, their compilation in this review will undoubtedly help rice (and others) researchers to effectively use them as a potential marker for better understanding, and ultimately, in defining rice (and plant) self-defense response pathways.
DOI:10.1007/s001220051178URL
A 1.1-kb DNA fragment containing the coding region of a thaumatin-like protein (TLP-D34), a member of the PR-5 group, was cloned into the rice transformation vector pGL2, under the control of the CaMV 35S promoter. The Indica rice cultivars, ‘Chinsurah Boro II’, ‘IR72’, and ‘IR51500’ were transformed with the tlp gene construct by PEG-mediated direct gene transfer to protoplasts and by biolistic transformation using immature embryos. The presence of the chimeric gene in T 0 , T 1 , and T 2 transgenic plants was detected by Southern blot analysis. The presence of the expected 23-kDa TLP in transgenic plants was confirmed by Western blot analysis and by staining with Coomassie Brilliant Blue. Bioassays of transgenic plants challenged with the sheath blight pathogen, Rhizoctonia solani , indicated that over-expression of TLP resulted in enhanced resistance compared to control plants.
[本文引用: 2]
DOI:10.1105/tpc.8.10.1809URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}