 ,1,2,*, 陈其鲜,3,*, 孙万仓1,2, 方彦1, 李学才1,2, 武军艳1,2
,1,2,*, 陈其鲜,3,*, 孙万仓1,2, 方彦1, 李学才1,2, 武军艳1,2Cloning of RuBisCo Subunits Genes rbcL and rbcS from Winter Rapeseed (Brassica rapa) and Their Expression under Drought Stress
MI Chao2,**, ZHAO Yan-Ning2,**, LIU Zi-Gang,1,2,*, CHEN Qi-Xian,3,*, SUN Wan-Cang1,2, FANG Yan1, LI Xue-Cai1,2, WU Jun-Yan1,2通讯作者:
第一联系人:
收稿日期:2017-09-21接受日期:2018-08-20网络出版日期:2018-12-12
| 基金资助: |
Received:2017-09-21Accepted:2018-08-20Online:2018-12-12
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1744KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
米超, 赵艳宁, 刘自刚, 陈其鲜, 孙万仓, 方彦, 李学才, 武军艳. 白菜型冬油菜RuBisCo蛋白亚基基因rbcL和rbcS的克隆及其在干旱胁迫下的表达[J]. 作物学报, 2018, 44(12): 1882-1890. doi:10.3724/SP.J.1006.2018.01882
MI Chao, ZHAO Yan-Ning, LIU Zi-Gang, CHEN Qi-Xian, SUN Wan-Cang, FANG Yan, LI Xue-Cai, WU Jun-Yan.
干旱是农作物生长发育的主要影响因子。随着全球气温的上升, 水资源匮乏加剧, 世界范围内将面临持续干旱, 干旱将成为世界范围农业发展的主要限制因素。我国西北地区属于干旱、半干旱地区, 年降水量较少、降水分布不均, 干旱严重制约着该地区的农业发展。
蛋白质组(Proteome)是生物个体、某一组织或细胞在特定时空表达的全部蛋白质, 蛋白质组学分析生物体内、组织或者细胞内蛋白质成分、表达水平及修饰情况, 从而了解蛋白质间的相互作用, 以此研究蛋白质水平上对生长发育的调节机制。干旱胁迫下, 白菜型冬油菜的存活率依赖于其避旱或耐旱能力, 其中, 光合作用是重要的代谢过程[1]。植物感知干旱胁迫后, 通常表现出气孔关闭、光系统损伤、气孔限制以及光合作用关键酶的含量及活性变化。高等植物1,5-二磷酸核酮糖羧化酶/加氧酶(RuBisCo, Ribulose-1,5-bisphosphate carboxylase/oxygenase)由8个大亚基(rbcL)和8个小亚基(rbcS)组成, 是光合作用中决定碳同化速率的关键酶。干旱胁迫下, 植物RuBisCo活性及含量均下降, 并且, 抗旱性强的材料下降程度小于敏感材料[2], 因此, 抗旱材料在干旱条件下光合作用维持在较高的水平。拟南芥遭受逆境胁迫时, RuBisCo大亚基发生解聚的同时, 与之相互作用的蛋白质及结合量也发生了变化。Jeroni等[2]、Massacci等[3]、Flexas等[4]研究显示, 中度干旱胁迫造成光合作用相关酶RuBisCO、PEPase等活性及含量下降。研究显示, 菠菜中rbcL表达水平受到rbcS调控[5]。从甘薯[6]、大豆[7]、青稞[8]、菊花[9,10]等植物中已分离克隆了rbcL和rbcS基因。白菜型油菜(Brassica rapa)是芸薹属的栽培油料作物之一, 具有耐迟播、生育期短、耐贫瘠、抗逆性强等突出优点, 也是适宜北方干旱地区种植的冬油菜品种, 但其抗旱机制尚不明确, 并且与白菜型油菜光合作用相关的基因克隆及生物信息学分析鲜有报道, 分离和克隆与之相关基因对明确白菜型冬油菜在干旱胁迫反应中响应机制及抗旱机理具有重要意义。本研究应用双向电泳(two-dimensional electrophoresis, 2-DE)技术, 得到干旱诱导表达的蛋白RuBisCo, 并进一步对rbcL和rbcS基因进行克隆及生物信息学分析, 以全面了解该基因在白菜型冬油菜陇油7号中的生物学功能, 为培育新的抗旱品种以及转基因抗旱冬油菜的探究提供理论依据。
1 材料与方法
1.1 供试材料
以白菜型冬油菜抗旱品种陇油7号为试材, 选取籽粒饱满、大小一致的种子, 10%过氧化氢处理30 min, 无菌水冲洗2~3次, 置铺有2层滤纸的培养皿内催芽(光照14 h, 30°C; 黑暗10 h, 28°C), 待种子露白后, 播于装有育苗基质的花盆(14 cm×13 cm), 每盆4株幼苗, 于人工培养箱中(光照14 h, 25°C; 黑暗10 h, 20°C)培养45 d至六叶期。干旱胁迫各处理为5盆, 处理分为对照组(CK)和中度干旱组(MR), 各组对应田间最大持水量的70%~75%、40%~45%[11,12]。处理期间每日下午18:00, 用HM-S型(山东恒美电子科技有限公司)土壤水分仪测量每日土壤含水量, 并用量筒定量补充水分以控制土壤含水量。干旱胁迫7 d。待胁迫处理结束, 取第4片幼嫩叶片冷冻保存, 取MR及CK叶片提取总蛋白和总RNA。试验均设置3次重复。于处理第7天上午9:30至11:00, 选取完全展开叶, 采用LI-6400便携式光合仪测定叶片光合参数气孔导度(Gs)、净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(Tr); 测定时叶室(2 cm×3 cm)内温度为20℃, 光强为600 μmol m-2 s-1; 选每个处理5~10株, 每株读数3次; 参照龚富生等[13]的方法测定RuBPCase 活性。称取完全展开叶0.5 g, 加2 mL Tris-HCl (pH 7.4)冰浴研磨后置2 mL离心管。4℃下1317×g 离心15 min, 取上清液待测。根据NADPH的氧化或NADP+的还原, 340 nm 测定样品的吸光值, 计算酶活力。
1.2 白菜型冬油菜叶片蛋白质筛选与分析
1.2.1 总蛋白提取与蛋白质浓度测定 采用TCA-丙酮沉淀法[14]提取白菜型冬油菜叶片总蛋白, 并用Bradford[15]法测定蛋白质浓度。1.2.2 蛋白质双向电泳 对干旱胁迫处理组(MR)和对照(CK)使用pH 4~7、17 cm的IPG胶条, 按照GE Healthcare双向电泳操作手册说明操作, 略有修改。样品上样量为1000 μg, 将样品与水化液按体积比1∶4混匀, 总体积不超过500 μL, 按说明书程序进行第一向等电聚焦电泳, 采用12%酰胺凝胶进行第二向SDS-PAGE, 经考马斯亮蓝(R-250)染色、脱色后, 采用GS-900 Calibrated Densitometer图像扫描系统(Bio-Rad)对凝胶进行图像扫描, 用ImageMaster 2D Platium 7.0凝胶分析软件对图谱标准化处理, 蛋白质点匹配和生物统计, 确定差异表达蛋白点。
1.2.3 差异蛋白点的质谱鉴定 对差异表达蛋白点回收、酶解, 送往华大基因股份公司(北京)进行LC-MS鉴定和数据库检索。
1.3 总RNA的提取及反转录
按照Spectru植物试剂盒(Sigma公司)提取总RNA, 电泳检测后按PrimeScript RT reagent Kit with gDNA Eraser试剂盒(大连TaKaRa公司)说明书进行反转录, 得到单链cDNA, 测定其浓度后, 置-20°C冰箱保存备用。1.4 引物设计与rbcL和rbcS基因克隆
根据GenBank数据库中已报道的白菜型油菜rbcL (Gene ID: 106352579)和rbcS (Gene ID:103863779)基因的核苷酸序列, 利用Primer Premier 5.0软件设计引物, rbcL-F: 5'-ATGTCACCACAAACAGAGACT-3', rbcL-R: 5'-CTACTCTTGGCCATCTAATTT-3', rbcS-F: 5'-ATGGC TTCCTCTATGCTTTC-3', rbcS-R: 5'-TTAAGCACCGGT GAAGCTTG-3'按照十字花科首位序列上设计, 可扩增出编码区全序列。以未经干旱处理的陇油7号叶片反转录cDNA为模板, 利用引物进行RT-PCR扩增, 扩增程序为95.0°C预变性5 min; 94.0°C变性30 s, 退火57.0°C 30 s, 72.0°C延伸80 s, 共循环35次; 最后72°C延伸, 保持10 min; 4°C保存。反应结束后以1%琼脂糖凝胶电泳检测, 利用天根生化科技(北京)有限公司的普通琼脂糖凝DNA纯化回收试剂盒并回收目的条带。回收产物与pMD-19T载体连接, 转化大肠杆菌DH5a感受态细胞, 过夜培养, 蓝白斑筛选, 挑白斑进行菌液培养。PCR检测菌落后, 随机挑选3个阳性克隆送华大基因股份公司(北京)股份有限公司测序。1.5 rbcL和rbcS预测蛋白的生物信息学分析
蛋白质基本理化性质、跨膜结构、信号肽、二级结构、保守结构域等预测均参考齐国昌[8]; 采用SWISS- MODEL工具预测蛋白质的三级结构, 并采用PyMOL软件对三级结构查看处理。利用Blast从GenBank中挑选9个来源于十字花科植物的rbcL和rbcS基因编码的蛋白质的氨基酸序列; 利用DNAMAN软件进行多重序列比较和氨基酸同源性分析; 采用MEGA5.0软件构建系统发生树。1.6 白菜型冬油菜rbcL和rbcS在干旱胁迫下的表达量
根据白菜型油菜rbcL和rbcS的编码区序列设计定量表达引物rbcL-S: 5'-GCAACGGGAAACGGGTGTT-3', rbcL-A: 5'-AGCGACCTTGAAGATTCTGGAGT-3', rbcS-S: 5'-GCTGCTGTGGTTACCTCCC-3', rbcS-A: 5'-TGTCGTT GTTTGCTTTGCG-3'。以浓度一致的陇油7号叶片cDNA为模板, β-actin: Act-F: 5'-TGTGCCAATCTACGAGGG TTT-3'; Act-R: 5'-TTTCCCGCTCTGCTGTTGT-3'为内参, 进行荧光定量和半定量PCR。半定量RT-PCR采用同机分管扩增内参基因和目的基因, 电泳检测rbcL和rbcS基因对干旱胁迫的响应模式, PCR扩增程序同上, 退火温度为60°C。参考SYBR Premix Ex Taq II (TliRNaseH Plus)试剂盒(大连TaKaRa公司)的荧光定量PCR, 采用两步法, 扩增程序为94°C 2 min; 95°C 10 s, 60°C 35 s, 40个循环。96孔上样板目的基因与内参基因对应各3次重复, 避光操作。采用2-ΔΔCt方法计算[14]。2 结果与分析
2.1 干旱胁迫冬油菜差异蛋白质筛选
采用ImageMaster 2D Plantium图像分析软件分析陇油7号叶片中度胁迫组(MR)与对照组(CK)的差异蛋白斑点(图1), 进行自动匹配, 并结合肉眼观察及手动调整, 共检测到1135~1478个的蛋白点, 干旱胁迫前后, 共有343个蛋白点发生了具有统计学显著差异的丰度变化, 表达量变化在8倍以上的蛋白点为21个, 其中, 增量表达蛋白质点共9个(spots 1, 3, 7, 9~13, 15), 减量表达蛋白质点8个(spots 4, 6, 8, 14, 17, 18, 20, 21), 诱导蛋白点4个(spots 2, 5, 16, 19)。正常水分条件下这些蛋白丰度较低或不表达, 干旱胁迫下表达发挥功能, 植物体内发生一系列生理生化变化, 进而植物耐受或抵御干旱环境的胁迫。2.2 干旱胁迫冬油菜差异蛋白质质谱鉴定及功能注释
切取干旱胁迫后诱导表达的21个蛋白点(图1, spots 1~21), 用胰蛋白酶进行胶内酶解, 经LC-MS质谱分析和数据库检索, 最后成功鉴定到21个蛋白质(表1)。参与刺激响应蛋白6个(spots 1, 2, 7, 8, 13, 16)、能量代谢蛋白6个(spots 6, 9, 11, 15, 17, 20)、脂代谢蛋白1个(spot 14)、糖代谢蛋白1个(spot 19)、信号转导过程蛋白1个(spot 21)、伴侣蛋白2个(spots 12, 18)、蛋白质代谢蛋白1个(spot 5)、光合作用蛋白3个(spots 3, 4, 10)。spot 4为叶绿体RuRisCo F1小链, 表达量下调10倍(表1)。图1
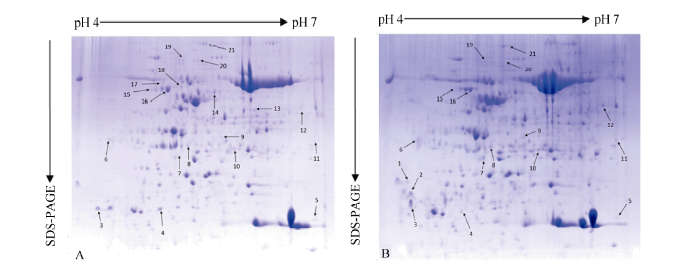 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1白菜型冬油菜陇油7号双向电泳图
A: pH 4~7对照; B: pH 4~7干旱处理。
Fig. 1Two-dimensional electrophoresis reference map of winter rapeseed Longyou 7
A: pH 4-7 contrast; B: pH 4-7 drought stress treatment.
Table 1
表1
表1LC-MS鉴定结果
Table 1
| 序号 Spot No. | 登录号 Accession | 蛋白名称 Protein name | 分子量/等电点 MW/pI | 得分 Score | 来源 Resource | 上调/下调 Up/down | |
|---|---|---|---|---|---|---|---|
| 1 | gi|541620 | 低温相关蛋白BN115 Low-temperature regulated protein BN115 | 14.8 kDa/7.75 | 81.8 | 甘蓝型油菜 Brassica napus | +10 | |
| 2 | gi|229893642 | 病毒抗性蛋白 Virus-resistance protein | 20.1 kDa/7.31 | 46.1 | 大白菜 Brassica rapa subsp. chinensis | +20 | |
| 3 | gi|109389998 | 叶绿体叶绿素a/b结合蛋白 Chloroplast chlorophyll a/b binding protein | 28.3 kDa/5.22 | 46.2 | 甘蓝型油菜 Brassica napus | +12 | |
| 4 | gi|132091 | 叶绿体Rubisco F1小链 Ribulose bisphosphate carboxylase small chain F1, chloroplastic; Short | 20.2 kDa/7.31 | 111 | 甘蓝型油菜 Brassica napus | -10 | |
| 5 | gi|171702843 | 半胱氨酸蛋白酶 Cysteine protease | 50.5 kDa/5.17 | 73.7 | 白菜型油菜 Brassica rapa | +18 | |
| 6 | gi|19570344 | 核苷二磷酸激酶1 Nucleoside diphosphate kinase 1 | 16.4 kDa/6.05 | 101 | 白菜型油菜 Brassica rapa | -11 | |
| 7 | gi|15225120 | 应激蛋白α-β结构域 Stress responsive alpha-beta barrel domain protein | 22.5 kDa/5.04 | 135 | 拟南芥 Arabidopsis thaliana | +13 | |
| 8 | gi|15241956 | 谷胱甘肽S-转移酶家族蛋白 Glutathione S-transferase family protein | 31.9 kDa/8.03 | 79.1 | 拟南芥 Arabidopsis thaliana | -15 | |
| 序号 Spot No. | 登录号 Accession | 蛋白名称 Protein name | 分子量/等电点 MW/pI | 得分 Score | 来源 Resource | 上调/下调 Up/down | |
| 9 | gi|356494238 | 磷酸甘露糖异构酶 Phosphomannomutase | 27.9 kDa/5.23 | 95.6 | 大白菜 Brassica rapa subsp. chinensis | +13 | |
| 10 | gi|29778743| | 叶绿体β-碳酸酐酶 Chloroplast beta-carbonic anhydrase | 37.7kDa/6.00 | 106 | 甘蓝型油菜 Brassica napus | +11 | |
| 11 | gi|14009294 | 假定的6-磷酸葡萄糖酸内酯酶 Putative 6-phosphogluconolactonase | 29.3 kDa/5.90 | 114 | 埃塞俄比亚芥 Brassica carinata | +14 | |
| 12 | gi|15234962 | DNAJ热激蛋白N-端结构域 DNAJ heat shock N-terminal domain-containing protein | 38.4 kDa/7.38 | 50.8 | 拟南芥 Arabidopsis thaliana | +12 | |
| 13 | gi|211905345 | 上皮硫特异蛋白 Epithiospecifier protein | 37.9kDa/5.71 | 73.8 | 白菜 Brassica rapa subsp. pekinensis | +15 | |
| 14 | gi|62321480 | 脂氧合酶 Lipoxygenase | 103.3 kDa/5.36 | 96.4 | 拟南芥 Arabidopsis thaliana | -14 | |
| 15 | gi|89257686 | 假定的乙酰鸟苷酸脱乙酰基酶 Acetylornithine deacetylase, putative | 48.0 kDa/5.62 | 129.0 | 甘蓝 Brassica oleracea | +12 | |
| 16 | gi|19553697| | β-1,3-葡聚糖酶 Beta-1,3-glucanase | 38.9 kDa/4.89 | 95.6 | 大白菜 Brassica rapa subsp. chinensis | +16 | |
| 17 | gi|356494248 | L-半乳糖脱氢酶 L-galactose dehydrogenase | 34.9 kDa/5.08 | 68 | 大白菜 Brassica rapa subsp. chinensis | -9 | |
| 18 | gi|41584275 | 热休克同源蛋白70 Heat shock cognate protein 70 | 28.1 kDa/5.17 | 65.7 | 山嵛菜 Eutrema halophilum | -16 | |
| 19 | gi|28540855 | 可溶性淀粉合成酶 Soluble starch synthase | 71.6 kDa/5.24 | 103 | 白菜 Brassica rapa subsp. pekinensis | +17 | |
| 20 | gi|15229519 | 二羟丙酮激酶 Dihydroxyacetone kinase | 61.9 kDa/5.22 | 111 | 拟南芥 Arabidopsis thaliana | -11 | |
| 21 | gi|197245081 | 玉米黄质环氧酶 Zeaxanthin epoxidase | 79.8 kDa/5.67 | 108 | 甘蓝型油菜 Brassica napus | -13 | |
新窗口打开|下载CSV
2.3 rbcL和rbcS基因克隆
本实验鉴定的21个差异表达蛋白质中3个蛋白点与光合作用相关, 其中spot 4为光合固碳酶Rubisco F1小链蛋白表达丰度下调10倍, 故克隆RuBisCo亚基rbcL和rbcS基因。以陇油7号cDNA为模板, 以1.4所述为引物进行RT-PCR扩增, 分别得到一条1550及600 bp的扩增产物, 回收纯化片段, 连接到T载体后转化, 利用菌液PCR筛选阳性克隆, 分别得到一条1500 bp及600 bp左右的目的片段, 说明克隆的基因已插入载体中。阳性克隆送华大基因(北京)股份有限公司测序得到碱基序列, 全长为1105 bp和552 bp。2.4 白菜型冬油菜rbcL和rbcS蛋白特性分析
采用NCBI ORF finder软件对白菜型冬油菜rbcL和rbcS基因编码蛋白质的序列与NCBI数据库中其他十字花科植物的对应基因编码蛋白质序列比对显示, 编码蛋白质序列基本一致。rbcL的完整开放阅读框1095 bp, 编码364个氨基酸(图2-A)。rbcS完整的开放阅读框546 bp, 编码181个氨基酸(图2-B)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2rbcL和rbcS基因的编码区核酸序列及编码氨基酸序列
上游的起始密码子ATG用方框表示; *为终止密码的位置。
Fig. 2Nucleotide sequence and deduced amino acid sequence of rbcL and rbcS cDNA
An upstream start codon ATG is boxed; * indicates the positions of termination codon.
预测rbcL蛋白质理化性质表明, 白菜型冬油菜rbcL蛋白由20种氨基酸组成, 相对分子量(MW)为40.29 kDa, 等电点(pI) 6.70; 预测结果显示rbcL是一个不稳定的蛋白质, 不稳定指数(instability index, II>40为不稳定蛋白质) 41.67; 脂融指数(aliphatic index, AI)为83.63, 平均亲水性值(grand average of hydropathicity, GRAVY)为-0.232, 为亲水性蛋白质。冬油菜rbcS蛋白主要由20种氨基酸组成, MW为20.32 kDa, pI为8.23, II为33.66, 为稳定蛋白质, 并且, AI为74.86, GRAVY为-0.142, 为亲水性蛋白质。
2.5 rbcL和rbcS蛋白结构预测
冬油菜rbcL跨膜结构存在3个跨膜结构域, 第1~18位氨基酸由里向外螺旋, 第145~169位氨基酸由外向里螺旋, 第247~267位氨基酸由里向外螺旋, N-端信号序列无信号肽断裂点, 二级结构包括38.74% α-螺旋、10.99%延伸链及50.27%自由卷曲, 新生rbcL肽链经过组装、加工后在细胞内与小亚基组成成熟的RuBisCo光合固碳酶。冬油菜rbcS存在一个跨膜的结构域, 第1~18位氨基酸由里向外螺旋, N-端信号序列不是一个信号多肽, 二级结构包含16.02% α-螺旋、28.73%延伸链及55.25%自由卷曲组成, 成熟的rbcS最终在绿色细胞内行使其功能。SWISS-MODEL预测rbcL三级结构, 模型序列区分度达到93.75, 构建模型的序列相似度达到0.60, 序列覆盖率达到0.99, 模型质量评估函数QMEAN达到-1.62, 为1,5-二磷酸核酮糖羧化/加氧酶(长链) (图3-A)。POCASA预测显示, rbcL蛋白共5个活性口袋。冬油菜rbcS三级结构, 模型序列覆盖率达到0.68, 序列区分度达到73.98, 序列相似性达到0.56, QMEAN达到-1.60, 为1,5-二磷酸核酮糖羧化/加氧酶(小链) (图3-B)。rbcS共4个活性口袋, 为rbcS底物结合区。
图3
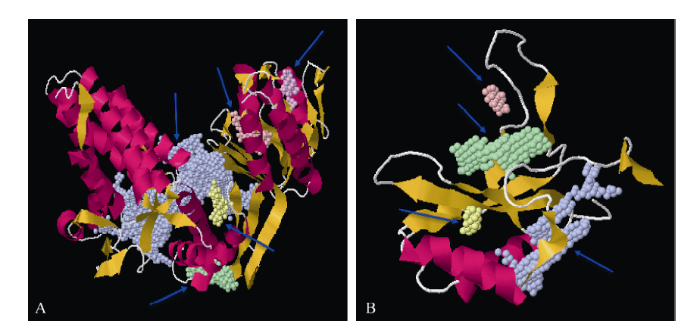 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3白菜型冬油菜rbcL和rbcS蛋白三级结构及活性口袋
A: rbcL蛋白质三级结构; B: rbcS蛋白质三级结构; 紫色箭头表示不同活性口袋。
Fig. 3rbcL and rbcS protein tertiary structure and active pocket in winter rapeseed
A : rbcL protein tertiary structure; B: rbcS protein tertiary structure; purple shows differential pockets.
rbcL蛋白保守区域属于RuBisCo Large superfamily, 存在第40~347位氨基酸的保守区域, 第84位氨基酸是一个催化位点, 第86、87位为2个金属离子结合位点, 均与光合作用有光。rbcL基因是由叶绿体DNA编码, 具有高的保守性, 冬油菜rbcL蛋白质保守域与NCBI数据库中其他植物有相似特征保守域。冬油菜rbcS蛋白保守区域存在20个多聚体结合位点, 在第2~44位氨基酸之间的rbcS superfamily保守区域及第65~175位氨基酸之间的RuBisCo small like superfamily的保守区域, 均与光合作用相关。
2.6 rbcL和rbcS氨基酸序列同源性及系统进化分析
rbcL亲缘关系分两类, 含大白菜(Brassica rapa subsp. chinensis, AHY18972.1)、甘蓝型油菜(Brassica napus, AMR44349.1)、白菜型油菜(Brassica rapa, AKG25031.1)、甘蓝(Brassica oleracea, P48686.1)所属的芸薹属为一类, 并与之亲缘关系较近, 氨基酸序列相似度达到99%; 萝卜(Raphanus sativus L., YP_009175683.1)所属的萝卜属、黑芥(Brassica nigra, AEK33921.1)、拟南芥(Arabidopsis thaliana, AMR44347.1)所属的鼠耳芥属、沙芥(Arabidopsis arenosa, YP_009230829.1)所属的沙芥属、亚麻荠 (Camelina sativa, YP_009231083.1)所属的荠菜属为一类, 序列相似度达到98% (图4-A)。rbcS的亲缘关系分两类, 含白菜型油菜(ABL97962.1)、甘蓝型油菜(ABB51649.1)、大白菜(BAJ08160.1)、甘蓝(XP_013635885.1)、芥菜型油菜(Brassica juncea, AEB00556.1)所属的芸薹属及亚麻荠(XP_010450911.1)所属的荠菜属为一类, 序列相似度达到95%以上, 其中陇油7号与甘蓝的关系最近(相似度99%); 拟南芥(CAA32701.1)所属的鼠耳芥属、萝卜(XP_006405 770.1)所属的萝卜属、山嵛菜(Eutrema salsugineum, XP_006405770.1)所属的山萮菜属为一类, 序列相似度达到94%以上(图4-B)。可见不同植物的rbcL和rbcS进化具有明显的种属特性。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4rbcL和rbcS蛋白与其他相关物种蛋白序列的系统进化树
Fig. 4Phylogenetic tree of rbcL and rbcS proteins with other related species
2.7 干旱胁迫下rbcL和rbcS基因的表达分析
实时荧光定量PCR结果显示, 冬油菜遭受干旱胁迫, 冬油菜rbcL和rbcS基因表达量分别下降到CK的51.90%、68.97%, 差异达到显著水平(P<0.05)(图5-A, 图5-B)。说明干旱胁迫严重影响冬油菜光合作用, 光合作用关键酶基因表达量下调, 是影响光合作用下降的因素。图5
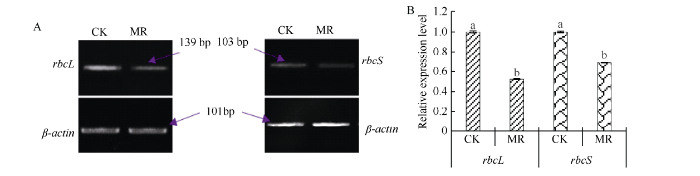 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5干旱胁迫下白菜型冬油菜rbcL和rbcS基因的半定量RT-PCR分析及相对表达量
CK: 对照组; MR: 中度干旱组。
Fig. 5Semi-quantitative RT-PCR analysis and relative expression of rbcL and rbcS genes in winter rapeseed under drought stress
CK: control; MR: middle drought stress.
2.8 干旱胁迫下白菜型冬油菜叶片光合气体交换参数及RuBPcase活性变化
随着干旱程度的加强, 白菜型冬油菜叶片Gs、Tr、Pn、Ci明显降低(表2)。表明叶片光合同化消耗CO2能力减弱, 通过气孔进入叶片的CO2在胞间积累, 导致Ci升高, 此时叶片Pn下降的主要原因是非气孔限制, 即光合系统效率降低所致。随着干旱胁迫的加强, 白菜型冬油菜叶片RuBPCase活性显著降低。与对照(CK)相比, 中度干旱下叶片RuBPCase活性下降了52.58%, 处理间差异达到显著水平。Table 2
表2
表2干旱胁迫对白菜型冬油菜‘陇油7号’光合气体交换参数及RuBPcase活性的影响
Table 2
| 处理 Treatment | 气孔导度 Gs (mol H2O m-2 s-1) | 胞间CO2浓度 Ci (μmol mol-1) | 蒸腾速率 Tr (mmol H2O m-2 s-1) | 净光合速率 Pn (μmol CO2 m-2 s-1) | RuBPCase activity (μmol m-2 s-1) |
|---|---|---|---|---|---|
| CK | 0.29±0.01 a | 330.4±6.65 b | 6.63±0.18 a | 0.90±0.04 a | 35.05±0.91 a |
| MR | 0.15±0.01 b | 395.7±3.33 a | 2.96±0.23 b | 0.47±0.11 b | 18.43±0.56 b |
新窗口打开|下载CSV
3 讨论
干旱条件下, 气孔关闭或部分关闭(气孔限制)及光合叶肉细胞光合能力的下降(非气孔限制)是光合速率下降的主要原因。RuBisCo是植物光合作用的关键酶, 既能催化1,5-二磷酸核酮糖(RuBP)与CO2形成3-磷酸甘油酸的反应, 又能催化RuBP与O2氧化裂解形成3-磷酸甘油酸、磷酸和磷酸乙醇酸的加氧反应, 具有双重作用, 其活性及细胞内含量受到逆境压力的影响[16]。抗旱品种在干旱条件下能维持相对较高的光合作用。本研究显示, 干旱胁迫下, 冬油菜叶片光合气孔参数Ci、Pn、Tr、Gs及光合作用关键酶RuBisCo活性显著下降, 叶片光合作用下降。前人研究显示, 非气孔限制下, RuBP羧化酶活性的降低、光合磷酸化活性的降低、RuBisCo及PEP羧化酶活性的降低等是影响植物光合作用下降的主要因素。作为光合作用固碳关键酶, RuBisCo活性降低亦是植物光合速率下降的非气孔限制因素之一[17,18,19,20]。高等植物叶绿体DNA是一个闭合的环形双链DNA, 进化过程中具有较高的保守性[7], 可以从系统进化上解释物种进化的多样性, rbcL是由叶绿体DNA编码, 并且RuBisCo酶的底物结合中心及活性中心均位于大亚基, 进化过程中保守性较高, 在系统发育研究中能确定种属间差异。rbcS是由核基因组编码, 能调节RuBisCo的蛋白质活性[11]。本实验从白菜型冬油菜叶片克隆到了具有完整ORF的rbcL和rbcS cDNA序列。rbcL编码区由1095 bp碱基组成, 编码364个氨基酸序列, 属于亲水性蛋白, 具有RuBisCo Large superfamily保守域, 该基因与十字花科大白菜、甘蓝型油菜、白菜型油菜、甘蓝等氨基酸序列具有较高的相似性, α-螺旋和自由卷曲是其二级结构的主要原件, 5个不同活性口袋区域、1个催化位点、2个金属结合位点是其三级结构重要结构区域; rbcS编码区由549 bp碱基组成, 编码181个氨基酸序列, 是亲水性蛋白, 具有rbcS superfamily和RuBisCo small like superfamily两个保守区域, 该蛋白与甘蓝rbcS同源关系最近(序列相似性达99%), 延伸链及自由卷曲是其二级结构的主要原件, 20个多聚体结合位点及4个活性口袋是其三级结构的重要区域。本研究中, 中度胁迫下, rbcL和rbcS基因表达量均下降, 说明干旱影响了植物基因表达, 光合关键酶RuBisCo合成受阻, 影响光合作用, 植物生长发育过程受到阻碍。
为了提高植物固定CO2的能力及光合作用效率, 研究者对多种植物RuBisCo大小亚基的结构及功能等方面进行了大量研究[20,21,22,23,24]。这些研究结果为采用生物技术的方法获得高光合利用率的作物品种提供了可能。本试验首先采用2-DE获得了干旱胁迫下白菜型冬油菜的差异表达蛋白RuBiSCo亚基, 以此克隆了RuBisCo的大小亚基的完整cDNA片段, 并预测了其结构及功能, 为获得高RuBisCo活性的白菜型冬油菜品种提供了依据。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3724/SP.J.1006.2015.00154URL [本文引用: 1]

assimilation (), stomatal conductance () and intercellular CO concentration () decreased during drought stress, showing the decreased photosynthesis due to stomatal factor. During moderate and severe drought stress, down-regulation in net CO fixation () was primarily mediated through non-stomatal limitation. The OJIP transients and other associated biophysical parameters elucidated the events of photoacclimatory changes in photosystem II (PSII) with progressive increase of drought stress, and the K-bands appeared in the stage of extreme drought severity indicating the imbalance between the electrons at the acceptor and donor sides of PSII, which suggested the oxygen-evolving complex (OEC) and electron transport were inhibited. In conclusion, the moderate and severedrought stress mainly damages the electron transfer from the plastoquinone (PQ) pool to the PSI terminal acceptors; this, along with constraints to both stomatal and non-stomatal components of photosynthesis, limits carbon assimilation. There are differences in cultivars with the response of PSII activity to drought stress.
DOI:10.3724/SP.J.1006.2015.00154URL [本文引用: 1]
assimilation (), stomatal conductance () and intercellular CO concentration () decreased during drought stress, showing the decreased photosynthesis due to stomatal factor. During moderate and severe drought stress, down-regulation in net CO fixation () was primarily mediated through non-stomatal limitation. The OJIP transients and other associated biophysical parameters elucidated the events of photoacclimatory changes in photosystem II (PSII) with progressive increase of drought stress, and the K-bands appeared in the stage of extreme drought severity indicating the imbalance between the electrons at the acceptor and donor sides of PSII, which suggested the oxygen-evolving complex (OEC) and electron transport were inhibited. In conclusion, the moderate and severedrought stress mainly damages the electron transfer from the plastoquinone (PQ) pool to the PSI terminal acceptors; this, along with constraints to both stomatal and non-stomatal components of photosynthesis, limits carbon assimilation. There are differences in cultivars with the response of PSII activity to drought stress.
[本文引用: 2]
DOI:10.1016/j.plaphy.2007.10.006URLPMID:18053735 [本文引用: 1]
The functioning of the photosynthetic apparatus of cotton ( Gossypium hirsutum) grown during the onset of water limitation was studied by gas-exchange and chlorophyll fluorescence to better understand the adaptation mechanisms of the photosynthetic apparatus to drought conditions. For this, cotton was grown in the field in Central Asia under well-irrigated and moderately drought-stressed conditions. The light and CO 2 responses of photosynthesis ( A G), stomatal conductance ( g s) and various chlorophyll fluorescence parameters were determined simultaneously. Furthermore, chlorophyll fluorescence images were taken from leaves to study the spatial pattern of photosystem II (PSII) efficiency and non-photochemical quenching parameters. Under low and moderate light intensity, the onset of drought stress caused an increase in the operating quantum efficiency of PSII photochemistry ( 01 PSII) which indicated increased photorespiration since photosynthesis was hardly affected by water limitation. The increase in 01 PSII was caused by an increase of the efficiency of open PSII reaction centers ( F v′/ F m′) and by a decrease of the basal non-photochemical quenching ( 01 NO). Using a chlorophyll fluorescence imaging system a low spatial heterogeneity of 01 PSII was revealed under both irrigation treatments. The increased rate of photorespiration in plants during the onset of drought stress can be seen as an acclimation process to avoid an over-excitation of PSII under more severe drought conditions.
DOI:10.1111/nph.2006.172.issue-1URL [本文引用: 1]
DOI:10.1093/abbs/36.9.644URL [本文引用: 1]
URL [本文引用: 1]
根据烟草、水稻和菠菜叶绿体的全基因组序列设计引物,以甘薯的叶绿体基因组DNA为模板,PCR扩增包舍 甘暑叶绿体rbcL完整基因(GenBank登录号为AY942199)在内的一段序列.序列分析表明:此片段的全长为1 627 bp,包括1 443bp的编码区序列在内.推测编码480个氨基酸,同时构建了此片段的限制性酶切图谱.相似性比较显示,此基因编码区序列与烟草、菠菜、小麦、水稻、 玉米、矮牵牛、紫花苜蓿、拟南芥、莨菪、葡萄以及甜菜的rbcL基因核苷酸的同源性为85%~98%,氨基酸的同源性为92%~95%.
URL [本文引用: 1]
根据烟草、水稻和菠菜叶绿体的全基因组序列设计引物,以甘薯的叶绿体基因组DNA为模板,PCR扩增包舍 甘暑叶绿体rbcL完整基因(GenBank登录号为AY942199)在内的一段序列.序列分析表明:此片段的全长为1 627 bp,包括1 443bp的编码区序列在内.推测编码480个氨基酸,同时构建了此片段的限制性酶切图谱.相似性比较显示,此基因编码区序列与烟草、菠菜、小麦、水稻、 玉米、矮牵牛、紫花苜蓿、拟南芥、莨菪、葡萄以及甜菜的rbcL基因核苷酸的同源性为85%~98%,氨基酸的同源性为92%~95%.
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
土地盐碱化是农业生产面临的生产难题之一,与此同时,随着生态环境的恶化和不合理的开发利用,使得这一问题变得更加突出。如何开发利用大量的盐碱地对现代农业的发展具有重要的意义。其中筛选培育优良的耐盐碱作物是对盐碱地进行经济高效改良的重要措施之一。青稞作为一种高原特色农作物具有生育期短、消耗土地肥力小、但产量并不低、同时又具有很好耐盐碱性的特点,因此,筛选鉴定一批耐盐碱较好的青稞品种(系)对于盐碱地进行改良或者是对植物耐盐机理的研究都具有比较重要的作用。此外,从分子生物学水平对植物的耐盐能力进行研究也是当前研究的热点,因为这将大大提高培育具有较强耐盐能力作物的速率。基于此,本实验对来自西藏、四川和甘肃三个地区的34份青稞品种(系)进行了芽期和苗期的耐盐性筛选鉴定,同时对和植物耐盐性相关的rbcS基因进行克隆,并对其在盐分胁迫条件下的表达情况进行了研究。得到的主要研究结果如下: 1.通过测定34份青稞材料的发芽势、发芽率、发芽指数等指标并应用主成分分析、隶属函数法等方法对青稞材料进行了多指标耐盐性综合评价。结果表明:耐盐性较强的材料是XQ0478、白玉青稞、XQ0792、XQ0198和N4-1等;耐盐性相对较弱的材料有矮黑六、矮六密、9718和9803-2。用综合指标值(D)进行聚类分析,可将34份青稞品种(系)聚为三类:A、B、C,A类的青稞材料有8份,其D>0.5,代表耐盐性较强的品种(系);B类的青稞材料有4份,其D<0.3,代表耐盐性较敏感的品种(系);C类的青稞材料有22份,其0.3≦D≦0.5,为中度耐盐性品种(系)。 2.用来自西藏、四川和甘肃的34份地方青稞品种(系)为实验材料,以叶绿素(Ch1)含量、细胞膜通透性和超氧化物歧化酶(SOD)含量等6个指标,运用相关分析、主成分分析和隶属函数法等分析方法,对青稞苗进行耐盐性综合分析。结果表明:耐盐性较强的材料是长芒红四棱、矮黑六和XQ0391;耐盐性相对较弱的材料是XQ0317、XQ0497和XQ0491等。对青稞材料综合评价结果(D)进行聚类分析,可将34份青稞品种(系)聚为3类,A类综合评价值D≧0.6,有3份材料是耐盐性相对较强的青稞材料。B类综合评价值0.44≦D<0.6,有18份材料是中度耐盐的青稞材料;C类的综合评价值D<0.44,有13份材料是相对盐敏感青稞材料。,并将评价结果与芽期耐盐性评价结果进行了比较,结果发现,在这34份青稞材料中有11份青稞材料在芽期和苗期耐盐性相同,其余材料在芽期和苗期表现出不同的耐盐能力。 3.根据大麦和小麦已知的1,5—二磷酸核酮糖羧化/加氧酶(Rubisco)小亚基基因rbcS的保守序列设计简并引物,通过同源PCR克隆技术扩增出了青稞rbcS基因的一条DNA序列和一条cDNA序列。经过测序和BLAST分析,结果显示:对比DNA序列和cDNA序列发现在DNA序列中含有一个长度为107bp的内含子。将得到的rbcS基因的核苷酸序列与其他物种的同源序列进行比对后发现,rbcS基因与小麦和大麦的rbcS基因之间亲缘关系最近相似性分别可以达到96.57%和91.0%;与土豆和番茄的rbcS基因之间亲缘关系较远相似性分别为59.12%和57.12%。 4.使用实时荧光定量PCR技术分析青稞材料rbcS基因在盐胁迫条件下不同时间点出的表达情况,结果显示:当青稞材料在受到盐分胁迫时rbcS基因表达量受到抑制,尤其是受到盐胁迫12h时其表达量相对于对照组有很大幅度的下降。此外,观察对照组后发现光照条件对其的表达有明显的调控作用。
URL [本文引用: 2]
土地盐碱化是农业生产面临的生产难题之一,与此同时,随着生态环境的恶化和不合理的开发利用,使得这一问题变得更加突出。如何开发利用大量的盐碱地对现代农业的发展具有重要的意义。其中筛选培育优良的耐盐碱作物是对盐碱地进行经济高效改良的重要措施之一。青稞作为一种高原特色农作物具有生育期短、消耗土地肥力小、但产量并不低、同时又具有很好耐盐碱性的特点,因此,筛选鉴定一批耐盐碱较好的青稞品种(系)对于盐碱地进行改良或者是对植物耐盐机理的研究都具有比较重要的作用。此外,从分子生物学水平对植物的耐盐能力进行研究也是当前研究的热点,因为这将大大提高培育具有较强耐盐能力作物的速率。基于此,本实验对来自西藏、四川和甘肃三个地区的34份青稞品种(系)进行了芽期和苗期的耐盐性筛选鉴定,同时对和植物耐盐性相关的rbcS基因进行克隆,并对其在盐分胁迫条件下的表达情况进行了研究。得到的主要研究结果如下: 1.通过测定34份青稞材料的发芽势、发芽率、发芽指数等指标并应用主成分分析、隶属函数法等方法对青稞材料进行了多指标耐盐性综合评价。结果表明:耐盐性较强的材料是XQ0478、白玉青稞、XQ0792、XQ0198和N4-1等;耐盐性相对较弱的材料有矮黑六、矮六密、9718和9803-2。用综合指标值(D)进行聚类分析,可将34份青稞品种(系)聚为三类:A、B、C,A类的青稞材料有8份,其D>0.5,代表耐盐性较强的品种(系);B类的青稞材料有4份,其D<0.3,代表耐盐性较敏感的品种(系);C类的青稞材料有22份,其0.3≦D≦0.5,为中度耐盐性品种(系)。 2.用来自西藏、四川和甘肃的34份地方青稞品种(系)为实验材料,以叶绿素(Ch1)含量、细胞膜通透性和超氧化物歧化酶(SOD)含量等6个指标,运用相关分析、主成分分析和隶属函数法等分析方法,对青稞苗进行耐盐性综合分析。结果表明:耐盐性较强的材料是长芒红四棱、矮黑六和XQ0391;耐盐性相对较弱的材料是XQ0317、XQ0497和XQ0491等。对青稞材料综合评价结果(D)进行聚类分析,可将34份青稞品种(系)聚为3类,A类综合评价值D≧0.6,有3份材料是耐盐性相对较强的青稞材料。B类综合评价值0.44≦D<0.6,有18份材料是中度耐盐的青稞材料;C类的综合评价值D<0.44,有13份材料是相对盐敏感青稞材料。,并将评价结果与芽期耐盐性评价结果进行了比较,结果发现,在这34份青稞材料中有11份青稞材料在芽期和苗期耐盐性相同,其余材料在芽期和苗期表现出不同的耐盐能力。 3.根据大麦和小麦已知的1,5—二磷酸核酮糖羧化/加氧酶(Rubisco)小亚基基因rbcS的保守序列设计简并引物,通过同源PCR克隆技术扩增出了青稞rbcS基因的一条DNA序列和一条cDNA序列。经过测序和BLAST分析,结果显示:对比DNA序列和cDNA序列发现在DNA序列中含有一个长度为107bp的内含子。将得到的rbcS基因的核苷酸序列与其他物种的同源序列进行比对后发现,rbcS基因与小麦和大麦的rbcS基因之间亲缘关系最近相似性分别可以达到96.57%和91.0%;与土豆和番茄的rbcS基因之间亲缘关系较远相似性分别为59.12%和57.12%。 4.使用实时荧光定量PCR技术分析青稞材料rbcS基因在盐胁迫条件下不同时间点出的表达情况,结果显示:当青稞材料在受到盐分胁迫时rbcS基因表达量受到抑制,尤其是受到盐胁迫12h时其表达量相对于对照组有很大幅度的下降。此外,观察对照组后发现光照条件对其的表达有明显的调控作用。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 2]
研究了长期不同干旱梯度胁迫条件下紫穗槐幼苗叶片各种生理生化指标变化.结果表明:轻度、中度、重度胁迫条件下SOD、POD活性明显高于对照,而且随着胁迫时间的延长而逐渐增高.重度胁迫下SOD、POD活性与轻度、中度差异显著.胁迫前期(1~60 d)SOD、POD活性变化具有波动现象,胁迫后期(60~120 d)SOD活性下降,POD活性上升.轻度、中度、重度胁迫条件下可溶性蛋白质、脯氨酸含量高于对照,而且随着胁迫时间的延长而增高.重度胁迫下可溶性蛋白质、脯氨酸含量与轻度、中度差异显著,轻度、中度胁迫条件下可溶性蛋白质、脯氨酸含量差异不显著;从第80天始脯氨酸含量下降,而可溶性蛋白质含量增高.轻度、中度、重度胁迫条件下,可溶性糖含量差异较小.轻度、中度、重度胁迫条件下MDA含量、细胞膜相对透性高于对照,且逐渐增高,重度与轻度、中度差异显著,细胞膜透性变化与MDA含量变化成明显的正相关,MDA含量变化与SOD活性变化成正相关.
Magsci [本文引用: 2]
研究了长期不同干旱梯度胁迫条件下紫穗槐幼苗叶片各种生理生化指标变化.结果表明:轻度、中度、重度胁迫条件下SOD、POD活性明显高于对照,而且随着胁迫时间的延长而逐渐增高.重度胁迫下SOD、POD活性与轻度、中度差异显著.胁迫前期(1~60 d)SOD、POD活性变化具有波动现象,胁迫后期(60~120 d)SOD活性下降,POD活性上升.轻度、中度、重度胁迫条件下可溶性蛋白质、脯氨酸含量高于对照,而且随着胁迫时间的延长而增高.重度胁迫下可溶性蛋白质、脯氨酸含量与轻度、中度差异显著,轻度、中度胁迫条件下可溶性蛋白质、脯氨酸含量差异不显著;从第80天始脯氨酸含量下降,而可溶性蛋白质含量增高.轻度、中度、重度胁迫条件下,可溶性糖含量差异较小.轻度、中度、重度胁迫条件下MDA含量、细胞膜相对透性高于对照,且逐渐增高,重度与轻度、中度差异显著,细胞膜透性变化与MDA含量变化成明显的正相关,MDA含量变化与SOD活性变化成正相关.
DOI:10.5846/stxb201412162507URL [本文引用: 1]
以山西道地药材黄芪一年生幼苗为试验材料,设置常规水分条件(CK)、轻度干旱胁迫(A1)、中度干旱胁迫(A2)、重度干旱胁迫(A3)4个不同处理,研究土壤干旱胁迫对黄芪生长及生理的影响.结果表明:黄芪茎叶快速生长集中在出苗后80-120d,以后生长减缓;当茎叶枯萎时,根中生物量短期快速积累.与常规水分条件相比,干旱胁迫处理显著降低了黄芪苗高及茎叶生物量,但对抗氧化能力、根系生长及次生代谢物积累产生了不同的影响.轻度干旱胁迫下SOD、POD、CAT 3种抗氧化酶活性升高,丙二醛(MDA)含量和细胞膜透性降低,同时根长与根生物量增加、多糖与皂苷两种次生代谢物积累增多,黄芪药材的质量得到显著提高(P<0.05);胁迫上升到中度、重度时,SOD酶活性逐渐降低,重度胁迫下低于对照,而POD及CAT酶活性、MDA含量、细胞膜透性均随胁迫增强而升高,相反,根长、根生物量、多糖与皂苷含量降低,导致黄芪药材的质量显著降低(P<0.01).综上表明,干旱胁迫下,SOD酶表现较为敏感,可能在清除活性氧中起主要作用;轻度水分胁迫能有效启动黄芪体内抗氧化酶系统和次生代谢,它们相互协作共同对抗胁迫对细胞产生的伤害,通过降低地上部分的生长,将营养物质优先运往根部,促进根产量及药材质量的提高.这一结论,可在黄芪多糖和皂苷次生代谢物定向培育的水分管理中加以利用.
DOI:10.5846/stxb201412162507URL [本文引用: 1]
以山西道地药材黄芪一年生幼苗为试验材料,设置常规水分条件(CK)、轻度干旱胁迫(A1)、中度干旱胁迫(A2)、重度干旱胁迫(A3)4个不同处理,研究土壤干旱胁迫对黄芪生长及生理的影响.结果表明:黄芪茎叶快速生长集中在出苗后80-120d,以后生长减缓;当茎叶枯萎时,根中生物量短期快速积累.与常规水分条件相比,干旱胁迫处理显著降低了黄芪苗高及茎叶生物量,但对抗氧化能力、根系生长及次生代谢物积累产生了不同的影响.轻度干旱胁迫下SOD、POD、CAT 3种抗氧化酶活性升高,丙二醛(MDA)含量和细胞膜透性降低,同时根长与根生物量增加、多糖与皂苷两种次生代谢物积累增多,黄芪药材的质量得到显著提高(P<0.05);胁迫上升到中度、重度时,SOD酶活性逐渐降低,重度胁迫下低于对照,而POD及CAT酶活性、MDA含量、细胞膜透性均随胁迫增强而升高,相反,根长、根生物量、多糖与皂苷含量降低,导致黄芪药材的质量显著降低(P<0.01).综上表明,干旱胁迫下,SOD酶表现较为敏感,可能在清除活性氧中起主要作用;轻度水分胁迫能有效启动黄芪体内抗氧化酶系统和次生代谢,它们相互协作共同对抗胁迫对细胞产生的伤害,通过降低地上部分的生长,将营养物质优先运往根部,促进根产量及药材质量的提高.这一结论,可在黄芪多糖和皂苷次生代谢物定向培育的水分管理中加以利用.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/0003-2697(76)90527-3URL [本文引用: 2]
DOI:10.1002/(ISSN)1097-0231URL [本文引用: 1]
DOI:10.1046/j.0016-8025.2001.00814.xURLPMID:11841670 [本文引用: 1]
Summary Experimental studies on CO 2 assimilation of mesophytic C3 plants in relation to relative water content (RWC) are discussed. Decreasing RWC slows the actual rate of photosynthetic CO 2 assimilation (A) and decreases the potential rate (A pot ). Generally, as RWC falls from c . 100 to c. 75%, the stomatal conductance (g s ) decreases, and with it A. However, there are two general types of relation of A pot to RWC, which are called Type 1 and Type 2. Type 1 has two main phases. As RWC decreases from 100 to c. 75%, A pot is unaffected, but decreasing stomatal conductance (g s ) results in smaller A, and lower CO 2 concentration inside the leaf (C i ) and in the chloroplast (C c ), the latter falling possibly to the compensation point. Down-regulation of electron transport occurs by energy quenching mechanisms, and changes in carbohydrate and nitrogen metabolism are considered acclimatory, caused by low C i and reversible by elevated CO 2 . Below 75% RWC, there is metabolic inhibition of A pot , inhibition of A then being partly (but progressively less) reversible by elevated CO 2 ; g s regulates A progressively less, and C i and CO 2 compensation point, rise. It is suggested that this is the true stress phase, where the decrease in A pot is caused by decreased ATP synthesis and a consequent decreased synthesis of RuBP. In the Type 2 response, A pot decreases progressively at RWC 100 to 75%, with A being progressively less restored to the unstressed value by elevated CO 2 . Decreased g s leads to a lower C i and C c but they probably do not reach compensation point: g s becomes progressively less important and metabolic limitations more important as RWC falls. The primary effect of low RWC on A pot is most probably caused by limited RuBP synthesis, as a result of decreased ATP synthesis, either through inhibition of Coupling Factor activity or amount due to increased ion concentration. Carbohydrate synthesis and accumulation decrease. Type 2 response is considered equivalent to Type 1 at RWC below c. 75%, with A pot inhibited by limited ATP and RuBP synthesis, respiratory metabolism dominates and C i and rise. The importance of inhibited ATP synthesis as a primary cause of decreasing A pot is discussed. Factors determining the Type 1 and Type 2 responses are unknown. Electron transport is maintained (but down-regulated) in Types 1 and 2 over a wide range of RWC, and a large reduced/oxidized adenylate ratio results. Metabolic imbalance results in amino acid accumulation and decreased and altered protein synthesis. These conditions profoundly affect cell functions and ultimately cause cell death. Type 1 and 2 responses may reflect differences in g s and in sensitivity of metabolism to decreasing RWC.
DOI:10.1038/44842URL [本文引用: 1]
Nature is the international weekly journal of science: a magazine style journal that publishes full-length research papers in all disciplines of science, as well as News and Views, reviews, news, features, commentaries, web focuses and more, covering all branches of science and how science impacts upon all aspects of society and life.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7666/d.y1832982URL [本文引用: 2]
RbcS (the small subunits of ribulose 1,5-bisphosphate carboxylase/oxygenase)基因是编码1,5-二磷酸核酮糖羧化/加氧酶(Rubisco)小亚基的基因。这种酶是叶绿体内含量最高的一种蛋白,位于叶绿体基质内,能催化光合作用中固定CO2的反应,其小亚基RbcS对全酶Rubisco的结构和功能起调控作用,且常以基因家族的形式定位于一条或多条染色体上,大多数高等植物中包含4~10个RbcS基因,各基因表达的氨基酸序列有差异,但是蛋白质在功能上基本是相似的。高等植物中的RbcS基因家族各成员在植物的各个器官及发育的各个时期的表达情况不完全一样,所受的调控机制也不同,这种表达调控上的不同在转录水平和转录后水平都存在,各个基因在DNA-蛋白质结合和mRNA稳定性方面都存在差异,也就是说以基因家族的形式存在不是单纯地为了增加基因产物的表达。 盐分是影响植物发育增殖的一个重要因素,高盐下,在多数植物以及蓝藻中,盐胁迫是抑制光合作用的,但盐藻却能增强光合作用,Liska等的研究表明当盐藻从低盐环境转移到高盐环境时,它能通过增强光合作用,增加C02的同化,从而产生大量甘油,达到适应高盐环境的目的。其中RbcS在C02的同化及淀粉合成方面发挥重要作用,但是盐藻的RbcS在盐分胁迫下的表达调控机制研究不多。本研究根据RbcS高度保守的氨基酸序列设计简并引物,采用RACE法扩增其5’和3’上下游未知序列,得到RbcS2新序列。再根据全长cDNA设计特异引物,利用荧光定量PCR方法分析光照和盐分对其表达的影响,这些为下一步研究RbcS基因家族中各成员的结构和功能,以及为探索盐胁迫下盐藻耐盐的分子机制提供实验依据。 实验方法 1简并引物扩增杜氏盐藻RbcS基因家族新成员的部分cDNA序列 根据GenBank上登录的杜氏盐藻、莱茵衣藻、红球藻、团藻等藻类的RbcS基因高度保守的氨基酸序列设计简并引物。采用TRIzol试剂提取杜氏盐藻细胞总RNA,用M-MuLV反转录酶合成cDNA第一链为模板,PCR扩增RbcS基因的部分cDNA片段,产物用T-A法亚克隆到pMD18-T载体上,酶切鉴定后测序,结果与GenBank上其他物种的RbcS基因比对分析。 2 RACE扩增杜氏盐藻RbcS2上下游序列 根据上述扩增出的RbcS2部分cDNA序列设计特异引物,以5’和3'RACE法扩增上下游未知序列,PCR产物用T-A法亚克隆到pMD18-T载体上,酶切鉴定后测序 3杜氏盐藻RbcS2全长cDNA的拼接和分析 将扩增出的简并cDNA,5’和3'RACE cDNA进行拼接,到cDNA全长序列,并利用DNAMEN和GenBank BLAST分析核苷酸和氨基酸的同源性及密码子偏好性。 4杜氏盐藻RbcS2功能分析 根据已克隆到的杜氏盐藻RbcS2的cDNA设计特异引物,以已知的杜氏盐藻保守基因GAPDH为内参照,Trizol试剂分别提取光照下和不同盐度下,不同时间培养的盐藻总RNA,反转录成cDNA,利用荧光定量PCR分析其光照和耐盐功能。 实验结果 1简并引物扩增杜氏盐藻RbcS基因家族新成员的部分cDNA序列 测序结果表明,简并引物扩增得到的部分cDNA片段长为209 bp,其氨基酸序列为69个氨基酸。BLAST结果表明,与GenBank中其他物种的RbcS有很高的同源性,且与杜氏盐藻RbcS1氨基酸的同源性为84%,说明所得的序列是杜氏盐藻RbcS的一个新片段。 2 RACE扩增杜氏盐藻RbcS2上下游序列 5'RACE得到一个有302个核苷酸的序列,除去部分已知序列及非编码区序列,共得到一个有168未知核苷酸的序列,由其推导的氨基酸序列有56个氨基酸残基,5'UTR部分有90个核苷酸。3'RACE得到一个有489个核苷酸的序列,除去部分已知序列及非编码区序列,共得到一个有271未知核苷酸的序列,由其推导的氨基酸序列有90个氨基酸残基,3'UTR部分有218个核苷酸。 3杜氏盐藻RbcS2全长cDNA的拼接和分析 将简并引物、5'RACE、3'RACE扩增得到的三段cDNA序列拼接起来,得到的cDNA全长为869 bp,包含561 bp的开放读码框,推导成多肽链包含187个氨基酸,BLAST结果显示,RbcS2的氨基酸序列与其他物种的RbcS的氨基酸序列有很高的同源性,且与杜氏盐藻的RbcS1(GenBank登录号AY739272)进行同源性分析,核苷酸和氨基酸序列的相似性分别为77.89%和77.19%,说明所得的序列是杜氏盐藻RbcS的一个新的片段,克隆得到了完整的杜氏盐藻RbcS基因家族的一个新成员RbcS2基因的cDNA序列,且与其它微藻的氨基酸序列同源性高些,与高等植物的氨基酸序列同源性低。对该基因密码子的使用偏好性进行分析,具有两种以上同义密码子的氨基酸偏好使用以C/G为结尾的密码子,表明杜氏盐藻和其他真核生物一样一般偏爱以C/G结尾的密码子。 4杜氏盐藻RbcS2功能分析 在黑暗条件下,RbcS2的表达量相对较低,RbcS2对光照的反应随时间推移,其转录表达先快速上升,达到平台期后逐步下降并趋于稳定,光照能强烈诱导该基因的表达,并具有时间效应。高盐下,RbcS2基因的转录表达高于正常水平,又由于有光照的影响,二者均随光暗有表达起伏变化。 结论与讨论 盐胁迫下,多数植物的光和作用都是受到抑制的,但盐藻却能增强光合作用。为分析盐藻高盐下促进光合作用独特的盐耐受性能,本研究从光合作用中固定CO2的关键基因RbcS入手,克隆到其基因家族中的一个新序列RbcS2的cDNA全长,并以此设计一对特异引物,利用相对荧光定量PCR分析其光照和耐盐功能。结果表明,与暗环境相比,光照能在一定时间内诱导RbcS2基因的转录表达;在高盐胁迫条件下,RbcS2的转录高于正常水平。高盐下,盐藻RbcS2的升高说明其可能是一个与盐胁迫相关的基因;这些结果为从分子水平上研究高盐下,盐藻的光合作用不是降低而是加强提供一些实验依据,及为高盐下盐藻分子水平的耐盐机制奠定基础。
DOI:10.7666/d.y1832982URL [本文引用: 2]
RbcS (the small subunits of ribulose 1,5-bisphosphate carboxylase/oxygenase)基因是编码1,5-二磷酸核酮糖羧化/加氧酶(Rubisco)小亚基的基因。这种酶是叶绿体内含量最高的一种蛋白,位于叶绿体基质内,能催化光合作用中固定CO2的反应,其小亚基RbcS对全酶Rubisco的结构和功能起调控作用,且常以基因家族的形式定位于一条或多条染色体上,大多数高等植物中包含4~10个RbcS基因,各基因表达的氨基酸序列有差异,但是蛋白质在功能上基本是相似的。高等植物中的RbcS基因家族各成员在植物的各个器官及发育的各个时期的表达情况不完全一样,所受的调控机制也不同,这种表达调控上的不同在转录水平和转录后水平都存在,各个基因在DNA-蛋白质结合和mRNA稳定性方面都存在差异,也就是说以基因家族的形式存在不是单纯地为了增加基因产物的表达。 盐分是影响植物发育增殖的一个重要因素,高盐下,在多数植物以及蓝藻中,盐胁迫是抑制光合作用的,但盐藻却能增强光合作用,Liska等的研究表明当盐藻从低盐环境转移到高盐环境时,它能通过增强光合作用,增加C02的同化,从而产生大量甘油,达到适应高盐环境的目的。其中RbcS在C02的同化及淀粉合成方面发挥重要作用,但是盐藻的RbcS在盐分胁迫下的表达调控机制研究不多。本研究根据RbcS高度保守的氨基酸序列设计简并引物,采用RACE法扩增其5’和3’上下游未知序列,得到RbcS2新序列。再根据全长cDNA设计特异引物,利用荧光定量PCR方法分析光照和盐分对其表达的影响,这些为下一步研究RbcS基因家族中各成员的结构和功能,以及为探索盐胁迫下盐藻耐盐的分子机制提供实验依据。 实验方法 1简并引物扩增杜氏盐藻RbcS基因家族新成员的部分cDNA序列 根据GenBank上登录的杜氏盐藻、莱茵衣藻、红球藻、团藻等藻类的RbcS基因高度保守的氨基酸序列设计简并引物。采用TRIzol试剂提取杜氏盐藻细胞总RNA,用M-MuLV反转录酶合成cDNA第一链为模板,PCR扩增RbcS基因的部分cDNA片段,产物用T-A法亚克隆到pMD18-T载体上,酶切鉴定后测序,结果与GenBank上其他物种的RbcS基因比对分析。 2 RACE扩增杜氏盐藻RbcS2上下游序列 根据上述扩增出的RbcS2部分cDNA序列设计特异引物,以5’和3'RACE法扩增上下游未知序列,PCR产物用T-A法亚克隆到pMD18-T载体上,酶切鉴定后测序 3杜氏盐藻RbcS2全长cDNA的拼接和分析 将扩增出的简并cDNA,5’和3'RACE cDNA进行拼接,到cDNA全长序列,并利用DNAMEN和GenBank BLAST分析核苷酸和氨基酸的同源性及密码子偏好性。 4杜氏盐藻RbcS2功能分析 根据已克隆到的杜氏盐藻RbcS2的cDNA设计特异引物,以已知的杜氏盐藻保守基因GAPDH为内参照,Trizol试剂分别提取光照下和不同盐度下,不同时间培养的盐藻总RNA,反转录成cDNA,利用荧光定量PCR分析其光照和耐盐功能。 实验结果 1简并引物扩增杜氏盐藻RbcS基因家族新成员的部分cDNA序列 测序结果表明,简并引物扩增得到的部分cDNA片段长为209 bp,其氨基酸序列为69个氨基酸。BLAST结果表明,与GenBank中其他物种的RbcS有很高的同源性,且与杜氏盐藻RbcS1氨基酸的同源性为84%,说明所得的序列是杜氏盐藻RbcS的一个新片段。 2 RACE扩增杜氏盐藻RbcS2上下游序列 5'RACE得到一个有302个核苷酸的序列,除去部分已知序列及非编码区序列,共得到一个有168未知核苷酸的序列,由其推导的氨基酸序列有56个氨基酸残基,5'UTR部分有90个核苷酸。3'RACE得到一个有489个核苷酸的序列,除去部分已知序列及非编码区序列,共得到一个有271未知核苷酸的序列,由其推导的氨基酸序列有90个氨基酸残基,3'UTR部分有218个核苷酸。 3杜氏盐藻RbcS2全长cDNA的拼接和分析 将简并引物、5'RACE、3'RACE扩增得到的三段cDNA序列拼接起来,得到的cDNA全长为869 bp,包含561 bp的开放读码框,推导成多肽链包含187个氨基酸,BLAST结果显示,RbcS2的氨基酸序列与其他物种的RbcS的氨基酸序列有很高的同源性,且与杜氏盐藻的RbcS1(GenBank登录号AY739272)进行同源性分析,核苷酸和氨基酸序列的相似性分别为77.89%和77.19%,说明所得的序列是杜氏盐藻RbcS的一个新的片段,克隆得到了完整的杜氏盐藻RbcS基因家族的一个新成员RbcS2基因的cDNA序列,且与其它微藻的氨基酸序列同源性高些,与高等植物的氨基酸序列同源性低。对该基因密码子的使用偏好性进行分析,具有两种以上同义密码子的氨基酸偏好使用以C/G为结尾的密码子,表明杜氏盐藻和其他真核生物一样一般偏爱以C/G结尾的密码子。 4杜氏盐藻RbcS2功能分析 在黑暗条件下,RbcS2的表达量相对较低,RbcS2对光照的反应随时间推移,其转录表达先快速上升,达到平台期后逐步下降并趋于稳定,光照能强烈诱导该基因的表达,并具有时间效应。高盐下,RbcS2基因的转录表达高于正常水平,又由于有光照的影响,二者均随光暗有表达起伏变化。 结论与讨论 盐胁迫下,多数植物的光和作用都是受到抑制的,但盐藻却能增强光合作用。为分析盐藻高盐下促进光合作用独特的盐耐受性能,本研究从光合作用中固定CO2的关键基因RbcS入手,克隆到其基因家族中的一个新序列RbcS2的cDNA全长,并以此设计一对特异引物,利用相对荧光定量PCR分析其光照和耐盐功能。结果表明,与暗环境相比,光照能在一定时间内诱导RbcS2基因的转录表达;在高盐胁迫条件下,RbcS2的转录高于正常水平。高盐下,盐藻RbcS2的升高说明其可能是一个与盐胁迫相关的基因;这些结果为从分子水平上研究高盐下,盐藻的光合作用不是降低而是加强提供一些实验依据,及为高盐下盐藻分子水平的耐盐机制奠定基础。
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.0517-6611.2011.22.020URL [本文引用: 1]
[目的]获得高纯度的苜蓿叶绿体DNA(cpDNA)和Rbcl基因片段.[方法]采用改进的高盐低pH法分离和纯化苜蓿叶绿体DNA;利用叶绿体基因组进化中高度保守的特点,设计引物,从苜蓿叶绿体基因组中扩增Rbcl基因,利用生物学软件对测序结果进行分析.[结果]经检测分析,获得了产率较高和纯度较好的苜蓿叶绿体DNA,并用PCR方法扩增获得了1130 bp的Rbcl基因片段.该基因片段有17个常用的限制性内切酶酶切位点,与烟草、水稻、菜豆、玉米、甘薯和甜菜中Rbcl基因核苷酸的同源性为85.6%~90.6%.[结论]为构建苜蓿叶绿体表达载体和通过叶绿体生物反应器生产药用蛋白等奠定了基础.
DOI:10.3969/j.issn.0517-6611.2011.22.020URL [本文引用: 1]
[目的]获得高纯度的苜蓿叶绿体DNA(cpDNA)和Rbcl基因片段.[方法]采用改进的高盐低pH法分离和纯化苜蓿叶绿体DNA;利用叶绿体基因组进化中高度保守的特点,设计引物,从苜蓿叶绿体基因组中扩增Rbcl基因,利用生物学软件对测序结果进行分析.[结果]经检测分析,获得了产率较高和纯度较好的苜蓿叶绿体DNA,并用PCR方法扩增获得了1130 bp的Rbcl基因片段.该基因片段有17个常用的限制性内切酶酶切位点,与烟草、水稻、菜豆、玉米、甘薯和甜菜中Rbcl基因核苷酸的同源性为85.6%~90.6%.[结论]为构建苜蓿叶绿体表达载体和通过叶绿体生物反应器生产药用蛋白等奠定了基础.
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}