 ,*农学基础实验教学中心 / 西南大学农学与生物科技学院, 重庆400715
,*农学基础实验教学中心 / 西南大学农学与生物科技学院, 重庆400715Identification and Gene Mapping of sdb1 Mutant with a Semi-dwarfism and Bigger Seed in Rice
TAO Yi-Ran, XIONG Yu-Zhen, XIE Jia, TIAN Wei-Jiang, ZHANG Xiao-Qiong, ZHANG Xiao-Bo, ZHOU Qian, SANG Xian-Chun, WANG Xiao-Wen,*Basic Experiment Teaching Center of Agronomy / College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China通讯作者:
第一联系人:
收稿日期:2018-03-9接受日期:2018-07-20网络出版日期:2018-08-07
| 基金资助: |
Received:2018-03-9Accepted:2018-07-20Online:2018-08-07
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (5817KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
陶怡然, 熊毓贞, 谢佳, 田维江, 张晓琼, 张孝波, 周倩, 桑贤春, 王晓雯. 水稻矮化大粒突变体sdb1的鉴定与基因定位[J]. 作物学报, 2018, 44(11): 1621-1630. doi:10.3724/SP.J.1006.2018.01621
TAO Yi-Ran, XIONG Yu-Zhen, XIE Jia, TIAN Wei-Jiang, ZHANG Xiao-Qiong, ZHANG Xiao-Bo, ZHOU Qian, SANG Xian-Chun, WANG Xiao-Wen.
水稻是世界上重要的粮食作物, 也是发育和遗传研究的模式植物。随着世界人口的逐年增加及耕地面积的连续减少, 选育高产品种对维护世界粮食安全具有重要的意义。水稻育种经历了3次绿色革命, 每一次绿色革命都伴随着株型的改良。第一次绿色革命始于20世纪60年代, 高秆变矮秆, 增加了水稻的抗倒伏性, 进而极大提高了单产; 2002年, 图位克隆了调控基因SD1, 编码赤霉素(GA)合成途径关键酶GA20ox-2[1,2,3]; 第二次绿色革命始于20世纪70年代野败型细胞质雄性不育(CMS-WA)的发掘, 2013年克隆了调控基因WA352[4]; 杂种优势结合株型改良实现了水稻单产的第二次飞跃。第三次绿色革命始于理想株型概念, 利用分子设计育种手段选育符合理想株型理念的水稻品种, 从而达到定向改良水稻的目的, 目前已克隆了IPA1等水稻理想株型调控基因[5,6]。
水稻株高由穗长、节间数量和节间长度三部分构成, 除少部分因穗长和节间数量发育缺陷外, 大多数矮秆是由于节间长度变短造成的。目前已经报道了80多个水稻矮化突变体, 其中基因克隆的有30多个, 多通过赤霉素、油菜素内酯、生长素、细胞分裂素等激素途径调控茎秆的发育[7]。部分通过调控细胞壁、微管微丝、角质层等的发育控制水稻株叶型的发育。如, OsFH5编码一种类甲精蛋白, 通过调节肌动蛋白动力学微管微丝的正确空间结构, 进而调控细胞的扩展和水稻植株的形态[8,9]; 此外, OsFH5/RMD也是生长素-肌动蛋白从细胞核到细胞质中自我组织调控环的关键组分[10]; OsFH5/RMD功能缺失突变体表现为植株矮化、籽粒变小等性状[8,9,10,11]。
矮化基因一般具有“一因多效”的特点。相关突变体除植株矮化外, 一般还伴随分蘖、叶片和籽粒等组织器官的变化, 如矮秆多蘖突变体d53、矮化卷叶突变体nrl1、矮秆小粒突变体tid、矮化淡叶突变体yld、矮秆花器官变异突变体ddf2等。NRL1编码类纤维素合酶基因, 突变体nrl1主要表现为叶片变窄、半卷以及不同程度的矮化[12], 等位突变体cd1还表现为谷粒减少、穗变短, 灌浆不充分、种子干扁细小[13]。TID编码α微管蛋白, 调控细胞的分裂和伸长, 突变体表现为植株矮化、地上部扭曲生长、根变短但不扭曲及籽粒变小等性状[14,15]。突变体yld叶色淡黄、植株矮化, 并伴随穗粒数、千粒重和结实率的显著降低[16]。ddf2节间的长和茎粗均极显著降低, 叶片极显著变短、变窄, 同时花序也极显著变短, 花器官特征发育受到了严重干扰[17]。
由于植株矮化性状的复杂性, 其分子机制还远不清晰, 此外, 鉴定的矮秆突变体多伴随小籽粒、致死等不良性状, 制约了这些矮源的育种利用[18]。通过筛选缙恢10号的EMS诱变体库, 我们鉴定到一个植株半矮化、籽粒显著变大的突变体sdb1 (Semi- Dwarf and Big seed mutant 1)。本研究对其开展了形态鉴定、细胞学观察和基因定位等研究, 为水稻分子设计育种提供了新的矮源, 也为调控基因的图位克隆和功能研究奠定了基础。
1 材料与方法
1.1 实验材料
sdb1来源于籼型水稻恢复系缙恢10号的EMS诱变, 连续多代自交, 突变表型已稳定遗传。配制中花11/sdb1杂交组合, 利用F1和F2群体进行遗传分析, 并利用F2隐性单株进行基因定位。所有材料均种植于西南大学水稻研究所歇马基地, 常规管理。1.2 形态鉴定
突变体和野生型种植于水稻田内, 3个小区, 每个小区50株, 全生育期调查株高和叶片大小; 成熟后, 每个小区取中间10株, 进行农艺性状考察, 性状包括株高、节间长、穗长、有效穗、穗粒数、穗实粒数、结实率、千粒重、粒长、粒宽、粒厚等。1.3 冷冻切片观察
取水稻成熟叶片中部, OTC冷冻切片包埋剂包埋, -20℃冰箱完全冰冻后, 利用冷冻切片机切片, 切片厚度为5~10 μm。于NIKON E6000显微镜下观察切片并照相, 统计分析细胞的大小和数量。1.4 石蜡切片分析
取水稻倒二节间中部及刚发育完整的小花, 以FAA固定液固定后, 依次进行乙醇脱水、二甲苯透明、石蜡包埋、切片、番红固绿染色。用1︰1的二甲苯和中性树胶封片后, 置42℃烘箱至少48 h, 然后用NIKON E6000显微镜观察并照相, 统计细胞大小和数量。1.5 光合色素含量测定
开花期, 早上9:00左右取sdb1和野生型植株的倒一叶、倒二叶和倒三叶, 参照王晓雯等[19]描述的方法测定光合色素含量。1.6 基因组DNA提取
采用改良的CTAB法提取亲本、基因池及F2定位群体DNA, 即取水稻叶片0.1 g左右, 剪成小块, 于2 mL离心管中, 加入2个直径2 mm钢珠, 液氮速冻后利用组织研磨机粉碎, 再加入0.7 mL 65℃预热的CTAB溶液, 放入65℃烘箱中, 每15 min上下颠倒3次, 45 min后取出, 再加入0.7 mL的氯仿, 颠倒混合后12 000 ×g离心5 min, 将0.5 mL上清液转移至1.5 mL的离心管中, 加入2倍的无水乙醇, 轻轻上下颠倒(期间出现白色絮状物), 12 000 ×g离心2 min, 弃上清液, 真空干燥至无酒精气味后, 加入200 μL ddH2O充分溶解, 即得基因组DNA。1.7 PCR分析
根据日本晴和9311的基因组序列, 利用Vector 10软件设计InDel标记, 分析中花11和sdb1之间的多态性, 多态性引物序列见表1, RM系列SSR引物则来源于http://www.gramene.org/网站。PCR反应总体系12.5 μL, 包括1.25 μL的10 × PCR buffer, 0.65 μL的25 mmol L-1 MgC12、0.5 μL 2.5 mmol L-1 dNTPs、8.0 μL的ddH2O, 1.0 μL的10 μmol L-1引物、1.0 μL的模板DNA和0.1 μL的5 U μL-1Taq DNA聚合酶。PCR反应程序为, 94℃预变性5 min, 94℃变性20 s, 55℃退火20 s, 72℃延伸20 s, 35个循环, 最后再72℃充分延伸10 min。PCR产物经10%非变性聚丙烯酰胺凝胶电泳, 0.1%质量浓度的AgNO3染色12 min, 去离子水漂洗2次, 1%质量浓度的NaOH和0.1%体积浓度的甲醛混合液染色, 直至条带清晰可见, 观察照相。Table 1
表1
表1基因定位引物序列
Table 1
| 引物名称 Primer name | 上游引物序列 Upstream primer sequence (5'-3') | 下游引物序列 Downstream primer sequence (5'-3') |
|---|---|---|
| J50-3 | TGCCTCCTTGTGTGTGGAGT | TCCAAGCACTGACTTCATAGC |
| J50-2 | CCACCAGTGATCCCTCATACA | TCCGAGCGATTCTACTGGTC |
| J50-7 | CCTCGTGTGGAATCTCGTTC | GTAACAGTAGGTGCGAAGATGG |
| J50-53 | GCGTTGTGTGCTCACTGG | CCACTTGTCAGGTCTCCATC |
| J50-65 | CCGGACGAGCCAATCAG | CGGTGGCTTCTCCCAAGG |
新窗口打开|下载CSV
1.8 连锁图谱构建
中花11/sdb1群体中的F2隐性单株中, 具有sdb1带型的单株记为B, 具有中花11带型的单株记为A, 具有中花11/sdb1 F1带型的单株记为H, 利用公式[(2A+H)/2n]×100%计算重组率, 其中, n表示总的F2隐性单株数, A和H分别表示具有A带和H带的单株数量。2 结果与分析
2.1 sdb1植株矮化
田间种植条件下, 苗期sdb1的株高略矮于野生型, 根少而短(图1-A); 抽穗期sdb1的各节间发育迟缓, 植株与野生型相比明显变矮(图1-B); 灌浆成熟期, sdb1的株高76.66 cm, 仅为野生型117.43 cm的65.28%, 差异达极显著水平(图1-C)。突变体5个节间和稻穗的长度均极显著短于野生型, 其中穗长由野生型的28.33 cm降至sdb1的22.14 cm, 下降了21.85% (图1-D, E, F)。因此, sdb1是一个全生育期矮化的水稻突变体, 其矮化性状是穗和各节间均变短造成的。图1
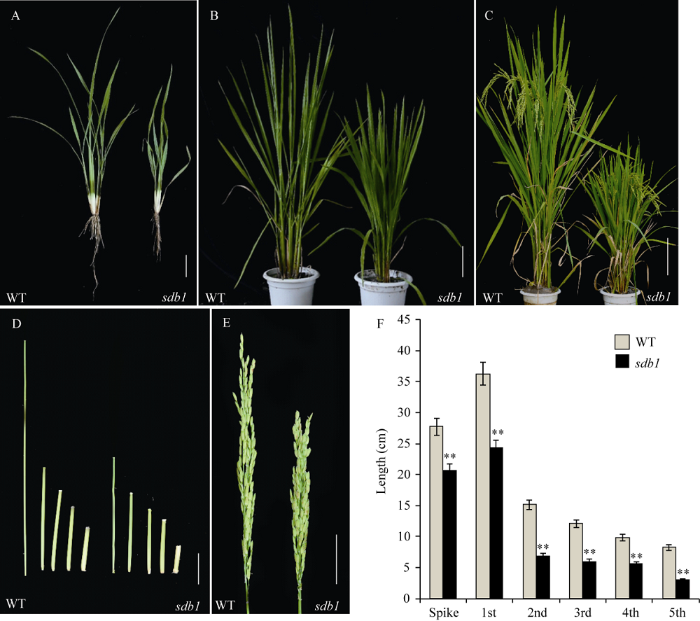 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1突变体sdb1和野生型WT的表型鉴定
A: 苗期野生型WT和突变体sdb1, Bar=5 cm; B: 分蘖期野生型WT和突变体sdb1, Bar=10 cm; C: 灌浆期野生型WT和突变体sdb1, Bar=15 cm; D: 灌浆期野生型WT和突变体sdb1的茎秆, Bar=5 cm; E: 灌浆期野生型WT和突变体sdb1的穗, Bar=5 cm; F: 野生型WT和突变体sdb1穗长及各节间长的统计分析。**在0.01水平上差异显著。
Fig. 1Plant phenotype detection of wild type (WT) and sdb1 mutant
A: phenotype of the wild type and the sdb1 at the seedling stage, Bar=5 cm; B: phenotype of the wild type and the sdb1 at the heading stage, Bar=10 cm; C: phenotype of the wild type and the sdb1 at the filling stage, Bar=15 cm; D: internodes of the wild type and the sdb1 at the filling stage, Bar=5 cm; E: panicle of the wild type and the sdb1 at the filling stage, Bar=5 cm; F: statistical analysis of panicle and internodes length of the wild type and the sdb1. **Significantly different at P<0.01.
从倒二节间的纵向石蜡切片中发现, 与野生型相比, sdb1的细胞数目增多, 但尺寸明显变小(图2-A, B, C, D); 单位面积内sdb1的细胞数目增加了1.24倍(图2-E), 细胞面积下降为野生型的44.64% (图2-F); sdb1细胞的长与野生型相比无显著变化(图2-G), 宽度则仅为野生型的41.18% (图2-H), 细胞宽度的极显著降低导致了sdb1细胞变小。倒二节间横切, 可更直观地看到sdb1横向细胞数目的增多和尺寸的变小(图2-I, J), sdb1单位面积细胞数量增加了2.79倍(图2-O), 细胞尺寸下降了73.60% (图2-P)。此外, sdb1维管束的面积也变小(图2-K, L), 但厚壁组织增多(图2-M, N)。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2野生型与突变体sdb1茎石蜡切片
A, B: 野生型WT和突变体sdb1茎秆纵切, Bar=100 μm; C: 图A红色小框的放大图, Bar=25 μm; D: 图B红色小框的放大图, Bar=25 μm; E:野生型WT和突变体sdb1茎秆纵切同一面积的细胞数目; F: 野生型WT和突变体sdb1茎秆纵切的细胞大小比较; G, H: 野生型WT和突变体sdb1茎秆纵切的细胞长度和宽度统计; I, J: 野生型WT和突变体sdb1茎秆横切, Bar=100 μm; K: 图I中红色小框放大图, Bar=25 μm; L: 图J的红色小框放大图, Bar=25 μm; M: 图I的黑色小框放大图, Bar=50 μm; N: 图J的黑色小框放大图, Bar=50 μm; O: 野生型WT和突变体sdb1茎秆横切同一面积的细胞数目统计; P: 野生型WT和突变体sdb1茎秆横切的细胞大小比较。**在0.01水平上差异显著。
Fig. 2Stems paraffin sections of the wild type (WT) and sdb1 mutant
A and B: longitudinal section of internodes of the wild type and the sdb1, Bar=100 μm; C: enlarged views of the red box in A, Bar=100 μm; D: enlarged views of the red box in B, Bar=100 μm; E:statistics of the cells number of the wild type and the sdb1 in the same area of longitudinal section; F: comparison of the cells size of the wild type and the sdb1 in longitudinal section; G and H: statistics of the cell length and width of the wild type and the sdb1in longitudinal section; I and J: gross section of internodes of the wild type and the sdb1, Bar=100 μm; K: enlarged views of the red box in I, Bar=25 μm; L: enlarged views of the red box in J, Bar=25 μm; M: enlarged views of the black box in I, Bar=50 μm; N: enlarged views of the black box in J, Bar=50 μm; O: statistics of the cells number of the wild type and the sdb1 in the same area of cross section; P:comparison of the cells size of the wild type and the sdb1 in cross section. **Significantly different at P < 0.01.
2.2 sdb1籽粒变大
成熟期sdb1的籽粒明显大于野生型WT (图3-A), 千粒重由24.83 g增加到29.00 g (图3-F), 达极显著差异水平。进一步分析发现sdb1籽粒的长、宽、厚均极显著增大, 其中, sdb1的籽粒长度为10.50 mm, 与WT的9.53 mm相比, 增长了10.18% (图3-G); sdb1的籽粒宽度和厚度分别为2.98 mm和2.12 mm, 与WT的2.75 mm和1.95 mm相比, 分别增加了8.36%和8.72% (图3-H)。进一步利用石蜡切片观察纵切小花, 发现sdb1和WT之间颖壳内细胞的大小无显著变化, 但sdb1中的细胞数量极显著增多(图3-B, C, D, E), 其薄壁细胞由WT的666.30个增加到sdb1的813.21个(图3-B, C), 增加了22.05% (图3-I)。细胞数量增多是导致sdb1籽粒变大的主要原因。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3野生型WT与突变体sdb1籽粒的比较
A: 野生型WT与突变体sdb1的籽粒, Bar=0.5 cm; B, C: 野生型WT和突变体sdb1横切籽粒的石蜡切片, Bar=500 μm; D: 图B红色小框的放大, Bar=50 μm; E: 图C红色小框的放大, Bar=50 μm; F: 野生型WT和突变体sdb1千粒重统计; G, H: 野生型WT和突变体sdb1细胞长度 (G)、宽度(H)、厚度(H)数据统计; I: 野生型WT(B)和突变体sdb1(C)颖壳薄壁细胞数目统计。**在0.01水平上差异显著。
Fig. 3Comparison of the grains between the wild-type and the sdb1 mutant
A: grains of WT and the sdb1, Bar=0.5 cm; B and C: cross section of the grains of WT and the sdb1, Bar=500 μm; D: enlarged views of the red box in B, Bar=50 μm; E: enlarged views of the red box in C, Bar=50 μm; F: statistical chart of 1000-grain weight of WT and the sdb1; G and H: statistical chart of cell length (G), width (H) and thickness (H) of WT and the sdb1; I: statistical the number of parenchyma cells of WT (B) and the sdb1 (C). **Significantly different at P < 0.01.
2.3 sdb1叶片变厚
田间种植条件下, 全生育期内sdb1与WT相比, 叶色更深绿(图4-A)。冷冻切片后荧光显微镜下观察发现, WT的叶肉细胞仅为4层(图4-B), 而sdb1有7层(图4-C), 叶肉细胞增多导致叶片变厚, 进而造成了sdb1叶色深绿。开花期, 测量倒一、倒二和倒三叶的光合色素含量发现, sdb1叶绿素a和类胡萝卜素的含量均极显著高于野生型, 倒一叶的叶绿素b含量与野生型相比无差异(图4-D)、倒二和倒三叶的叶绿素b的含量显著和极显著小于野生型(图4-E, F)。图4
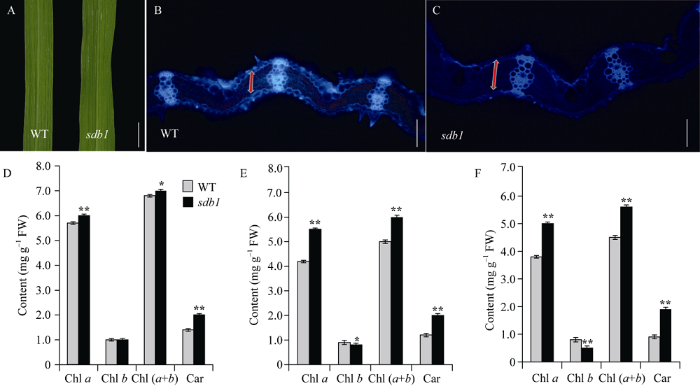 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4冷冻切片和开花期光合色素含量分析
A: 成熟期野生型和突变体sdb1倒一叶中部, Bar=2 cm; B, C: 野生型和突变体sdb1叶片中部冷冻切片横切, Bar=50 μm ; D: 野生型与突变体sdb1倒一叶光合色素含量; E: 野生型与突变体sdb1倒二叶光合色素含量; F: 野生型与突变体sdb1倒三叶光合色素含量。Chl a: 叶绿素a; Chl b: 叶绿素b; Car: 类胡萝卜素。**在0.01水平上差异显著; *在0.05水平上差异显著。
Fig. 4Frozen sections and photosynthetic pigment contents at flowering stage
A: the middle of flag leaf of the wild type (WT) and the mutant sdb1 at heading stage; B and C: frozen section of crosscutting leaf of the wild type and the mutant sdb1, Bar=50 μm; D: chlorophyll content of the flag leaf blade; E:chlorophyll content of the second leaf from the top; F: chlorophyll content of the third leaf from the top; Chl a: chlorophyll a; Chl b: chlorophyll b; Car: carotenoid. **Significantly different at P<0.01; *Significantly different at P<0.05.
2.4 sdb1受单隐性核基因调控
中花11和sdb1杂交组合的F1植株形态正常, F2群体中出现半矮化大籽粒和正常两种植株形态, 且χ2测验正常植株(2268)与半矮化大籽粒植株(756)的分离比符合3:1 (χ20.05=0.48<χ20.05=3.84), 暗示sdb1矮化大籽粒性状受1对隐性核基因调控。2.5 SDB1的分子定位
平均分布在水稻12条染色体上的460对RM系列SSR引物, 123对在中花11和sdb1之间具有多态性。从中花11/sdb1杂交组合F2群体中取10株正常单株和10株突变单株, 构成正常池和突变池, 利用亲本间具有多态性的引物进一步分析基因池, 并对可能连锁的标记用70株F2隐性单株验证, 初步将SDB1定位在第4染色体标记RM8212与J50-3之间, 交换株分别有16个和4个。为进一步定位SDB1, 在初步定位的基础上又合成了15对标记, 其中6对在亲本中花11和sdb1间有差异。利用这6对差异引物进一步分析756株中花11/sdb1的F2隐性单株发现, 标记J50-53、J50-65、RM16632、J50-2、J50-7和RM3317的交换株分别为63、13、1、0、2和56个, 且前3个标记的交换株和后2个标记的交换株不同, 从而最终将SDB1限定在第4染色体标记RM16632和J50-7之间(图5), 且与J50-2共分离, 物理距离约406 kb。图5
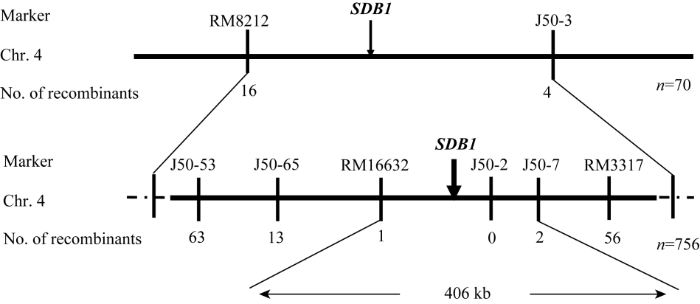 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5SDB1在水稻第4 染色体上的分子定位
Fig. 5Molecular mapping of SDB1 gene on rice chromosome 4
利用Gramene和RGAP等网站进行生物信息学分析发现, SDB1的定位区间位于着丝粒附近, 包含42个注释基因, 其中, 13个编码转座子蛋白、10个编码反转座子蛋白、2个编码假定蛋白; 另外17个基因中有11个编码表达蛋白、2个编码甘露糖结合凝集素家族蛋白、2个编码碱性螺旋-环-螺旋(bHLH)蛋白、1个编码木糖基转移酶、1个编码阳离子外排家族(CEF)蛋白(表2)。
Table 2
表2
表2定位区间 RGAP 注释的表达蛋白和功能基因
Table 2
| 基因名称 Locus name | 基因注释 Gene annotation |
|---|---|
| LOC_Os04g23170 | Expressed protein |
| LOC_Os04g23180 | Cation efflux family protein, putative, expressed |
| LOC_Os04g23200 | Expressed protein |
| LOC_Os04g23210 | Expressed protein |
| LOC_Os04g23220 | Expressed protein |
| LOC_Os04g23230 | Expressed protein |
| LOC_Os04g23319 | Expressed protein |
| LOC_Os04g23330 | Expressed protein |
| LOC_Os04g23420 | Expressed protein |
| LOC_Os04g23440 | Helix-loop-helix DNA-binding domain containing protein, expressed |
| LOC_Os04g23460 | Expressed protein |
| LOC_Os04g23550 | Basic helix-loop-helix family protein |
| LOC_Os04g23580 | Xylosyltransferase, putative, expressed |
| LOC_Os04g23600 | D-mannose binding lectin family protein, expressed |
| LOC_Os04g23610 | Expressed protein |
| LOC_Os04g23620 | D-mannose binding lectin family protein, expressed |
| LOC_Os04g23630 | Expressed protein |
新窗口打开|下载CSV
3 讨论
矮秆突变体通常伴随着籽粒的变化, 根据籽粒发育形态, 可分为畸形粒、小粒、正常粒和大粒4类。目前报道的矮秆突变体, 多表现为籽粒畸形或小籽粒, 如d11[20]、d2[21,22]、ddf1[23]等; 这些资源的鉴定极大促进了水稻株型发育的分子机制研究, 但不良农艺性状制约了其育种利用。株高适度矮化、籽粒大小不变或略大是培育超高产水稻的良好载体, 在水稻生产中具有更高的应用价值, 如第一次绿色革命利用的半矮秆资源sd1, 其籽粒大小就基本无变化[1]。本文鉴定的sdb1, 植株半矮化且千粒重极显著增大, 暗示该资源在水稻理想株型育种中可能具有潜在的应用价值, 这对避免单一矮源引起的水稻生产风险具有重要的意义。植株矮化一般受细胞分裂和扩展的影响, 即与细胞数目和大小的发育有关。如BC12/GDD1编码一个双靶向驱动蛋白-4, 即通过减少细胞数目导致bc12的植株矮化[24]; 水稻矮化剑叶卷曲突变体dcfl1则是细胞变短造成的[25]。DLT和SMOS1是油菜素内酯BR、赤霉素GA、生长素IAA和细胞分裂素CTK等激素调控途径中的重要节点基因, 其功能缺失突变体均表现为植株半矮化、叶片直立色深、穗型直立等, 二者均通过细胞的分裂和扩展调控水稻发育[26,27,28,29,30,31]。与突变体dlt和smos1类似, sdb1中的细胞数量显著增多, 但细胞变窄导致细胞变小, 推测SDB1功能缺陷促进了细胞横向分裂而抑制了细胞的横向生长和纵向分裂, 进而导致sdb1植株半矮化和籽粒变大, 但具体机制还有待进一步研究。此外, 我们还发现sdb1的叶色深绿, 叶肉细胞增多导致叶片变厚。叶肉细胞增厚有利于提高叶片的光能利用效率, 如nal1等位突变体lschl4就表现叶肉细胞增多和光能利用率显著增高, 在水稻高光效育种中具有重要的应用价值[32]。
本研究利用SSR等分子标记, 最终将SDB1定位在水稻第4染色体着丝粒附近406 kb的物理范围内。目前, 在水稻第4染色体上克隆的矮秆基因有2个。HTD1编码胡萝卜素裂解双加氧酶OsCCD7, 是独角金内酯生物合成过程中的重要参与酶之一, 功能缺失突变体表现矮秆多蘖[33]; D11则编码油菜素内酯生物合成过程中的一种关键酶, 隐性突变导致BR生物合成受阻, 植株矮化[20]。矮泰引-3植株半矮化, 受2对基因调控, 一个位点可能是SD1的等位突变, 另一个SD2定位在第4染色体上, 位于SSR标记RM1305和RM5633之间[34]。tddl(t)来源于籼稻IR64的EMS诱变, 是一个多蘖矮秆且叶色变深的突变体, 突变表型也是细胞变小增多造成的, 基因被定位在第4染色体SSR标记 RM5511和STS标记4-1082之间85.51 kb的物理范围内[35]。尽管SD2和TDDL(T)都被定位在第4染色体上, 但突变表型和定位区间均不同于SDB1, 这表明sdb1可能受一个新的尚未鉴定的功能基因调控。
bHLH蛋白是一类多功能蛋白, 如PGL2通过与典型bHLH蛋白APG互作调控水稻籽粒长度和重量[36]; RERJ1则可作为一个转录激活子调控胁迫下的诱导基因表达, 并与胁迫区域的茉莉酸积累相关[37]。RCN11编码一个β-1,2-木糖基转移酶OsXylT, 参与水稻对非生物胁迫和激素的应答, 功能缺失突变体在低温条件下株高变矮、分蘖减少、地上部和地下部干重严重降低[38,39]; CEF蛋白在金属离子的运输方面发挥重要作用, 如OsMTP9与锰元素的吸收有关[40], OsMTP11则与Cd、Zn、Ni和Mn等金属离子的转运有关[41]; Lectin receptor-like kinases (LecRLKs)在植物逆境响应中调控植株的生长和发育[42]。在SDB1定位区间内的42个注释基因中, 除转座子、反转座子、假定蛋白外, 还有11个表达蛋白和6个功能基因, 6个功能基因分别编码甘露糖结合凝集素家族蛋白、bHLH蛋白、木糖基转移酶和CEF蛋白, 除对RERJ1 (LOC_Os04g23550)进行了初步研究外[37], 其余功能均未知, 但具体哪一个是SDB1的目的基因, 还需进一步研究。
4 结论
全生育期内, sdb1的株高都矮于野生型, 成熟期仅为野生型的65.28%, 差异达极显著水平。茎秆中纵向细胞长度无显著变化, 但细胞宽度明显变窄。横向细胞数目增多、纵向细胞变少是导致sdb1株高矮化的主要原因。sdb1的千粒重极显著高于野生型, 其籽粒变大是由于颖壳中薄壁细胞数量极显著增多。此外, sdb1还表现叶色深绿、叶片较厚等特性。该突变性状受1对隐性核基因调控, 被定位在第4染色体SSR标记RM16632和J50-7之间约406 kb的物理范围内。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/416701aURL [本文引用: 2]
[本文引用: 1]
.
[本文引用: 1]
DOI:10.1038/ng.2570URLPMID:23502780 [本文引用: 1]

Plant cytoplasmic male sterility (CMS) results from incompatibilities between the organellar and nuclear genomes and prevents self pollination, enabling hybrid crop breeding to increase yields1–6. The Wild Abortive CMS (CMS-WA) has been exploited in the majority of ‘three-line’ hybrid rice production since the 1 1970s, but the molecular basis of this trait remains unknown. Here we report that a new mitochondrial gene, WA352, which originated recently in wild rice, confers CMS-WA because the protein it encodes interacts with the nuclear-encoded mitochondrial protein COX1111. In CMS-WA lines, WA352 accumulates preferentially in the anther tapetum, thereby inhibiting COX11 11 11 function in peroxide metabolism and triggering premature tapetal programmed cell death and consequent pollen abortion. WA352-induced sterility can be suppressed by two restorer-of-fertility (Rf) genes, suggesting the existence of different mechanisms to counteract deleterious cytoplasmic factors. Thus, CMS-related cytoplasmic-nuclear incompatibility is driven by a detrimental interaction between a newly evolved mitochondrial gene and a conserved, essential nuclear gene.
DOI:10.1038/ng.591URLPMID:20495565 [本文引用: 1]
Abstract Increasing crop yield is a major challenge for modern agriculture. The development of new plant types, which is known as ideal plant architecture (IPA), has been proposed as a means to enhance rice yield potential over that of existing high-yield varieties. Here, we report the cloning and characterization of a semidominant quantitative trait locus, IPA1 (Ideal Plant Architecture 1), which profoundly changes rice plant architecture and substantially enhances rice grain yield. The IPA1 quantitative trait locus encodes OsSPL14 (SOUAMOSA PROMOTER BINDING PROTEIN-LIKE 14) and is regulated by microRNA (miRNA) OsmiR156 in vivo. We demonstrate that a point mutation in OsSPL14 perturbs OsmiR156-directed regulation of OsSPL14, generating an 'ideal' rice plant with a reduced tiller number, increased lodging resistance and enhanced grain yield. Our study suggests that OsSPL14 may help improve rice grain yield by facilitating the breeding of new elite rice varieties.
DOI:10.1038/ng.592URLPMID:20495564 [本文引用: 1]
Identification of alleles that improve crop production and lead to higher-yielding varieties are needed for food security. Here we show that the quantitative trait locus WFP (WEALTHY FARMER'S PANICLE) encodes OsSPL14 (SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 14, also known as IPA1). Higher expression of OsSPL14 in the reproductive stage promotes panicle branching and higher grain yield in rice. OsSPL14 controls shoot branching in the vegetative stage and is affected by microRNA excision. We also demonstrate the feasibility of using the OsSLP14WFP allele to increase rice crop yield. Introduction of the high-yielding OsSPL14WFP allele into the standard rice variety Nipponbare resulted in increased rice production.
DOI:10.3724/SP.J.1006.2016.01261URL [本文引用: 1]
株高是影响水稻产量的一个重要性状。本研究从水稻稻瘟病普感品种丽江新团黑谷(LTH)经甲基磺酸乙酯(EMS)诱变群体中分离出一个遗传稳定的小粒矮化突变体LTH-m3。该突变体是赤霉素(GA)和油菜素内酯(BR)相关突变体,它对外源GA(GA_3)不敏感,对外源BR(eBL)的敏感性较野生型显著降低。遗传分析、基因克隆和转基因互补实验确认,该突变体是一个新的d1基因等位突变体,其D1基因在第6个外显子与内含子接合处发生单碱基突变(G(2522)→A(2522)),导致第6外显子被选择性剪切及Gα蛋白翻译提前终止,从而造成LTH-m3小粒矮化突变表型。进一步的研究表明,该突变体D1基因突变引起SD1和SLR1等基因表达的显著改变,因而影响植株细胞内GA和BR反馈调节功能和信号传递。突变体LTH-m3弥补了LTH植株过高、茎秆软和极易倒伏等缺陷,可作为LTH的改良系在今后水稻稻瘟病研究中加以利用,其功能突变基因的鉴定为深入研究水稻Gα蛋白的功能及激素信号途径提供了新的材料。
.
DOI:10.3724/SP.J.1006.2016.01261URL [本文引用: 1]
株高是影响水稻产量的一个重要性状。本研究从水稻稻瘟病普感品种丽江新团黑谷(LTH)经甲基磺酸乙酯(EMS)诱变群体中分离出一个遗传稳定的小粒矮化突变体LTH-m3。该突变体是赤霉素(GA)和油菜素内酯(BR)相关突变体,它对外源GA(GA_3)不敏感,对外源BR(eBL)的敏感性较野生型显著降低。遗传分析、基因克隆和转基因互补实验确认,该突变体是一个新的d1基因等位突变体,其D1基因在第6个外显子与内含子接合处发生单碱基突变(G(2522)→A(2522)),导致第6外显子被选择性剪切及Gα蛋白翻译提前终止,从而造成LTH-m3小粒矮化突变表型。进一步的研究表明,该突变体D1基因突变引起SD1和SLR1等基因表达的显著改变,因而影响植株细胞内GA和BR反馈调节功能和信号传递。突变体LTH-m3弥补了LTH植株过高、茎秆软和极易倒伏等缺陷,可作为LTH的改良系在今后水稻稻瘟病研究中加以利用,其功能突变基因的鉴定为深入研究水稻Gα蛋白的功能及激素信号途径提供了新的材料。
DOI:10.1105/tpc.110.081349URL [本文引用: 2]
DOI:10.1105/tpc.110.081802URL [本文引用: 2]
DOI:10.1073/pnas.1401680111URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2017.00522URL [本文引用: 1]
水稻叶色突变体是研究植物光合作用、叶绿素代谢和叶绿体发育的重要材料。本研究从籼稻品种蜀恢527经EMS(甲基磺酸乙酯)诱变处理后代中筛选出一个淡黄叶矮化突变体Yellow leaf and dwarf(yld)。与野生型蜀恢527相比,该突变体全生育期都表现出淡黄叶矮化性状,其剑叶的淡黄色表型最为明显,倒二叶次之,倒三叶最弱,其中剑叶的叶绿素及类胡萝卜素含量降低最为明显;并且伴随着穗粒数、千粒重、结实率、株高等主要农艺性状的显著降低,但有效穗显著增多。透射电镜观察结果显示,与野生型相比,该突变体多数叶绿体结构基本完整,但基粒模糊,基质片层大量减少且排列疏松。遗传分析表明,该突变性状受一对隐性核基因控制。在yld突变体与粳稻武运粳7号杂交的F2群体中分离出323个突变单株,最终将YLD基因定位在第11染色体的L5和L7两标记之间,物理距离为115.7 kb。本研究为YLD基因的克隆和功能分析奠定了基础。
DOI:10.3724/SP.J.1006.2017.00522URL [本文引用: 1]
水稻叶色突变体是研究植物光合作用、叶绿素代谢和叶绿体发育的重要材料。本研究从籼稻品种蜀恢527经EMS(甲基磺酸乙酯)诱变处理后代中筛选出一个淡黄叶矮化突变体Yellow leaf and dwarf(yld)。与野生型蜀恢527相比,该突变体全生育期都表现出淡黄叶矮化性状,其剑叶的淡黄色表型最为明显,倒二叶次之,倒三叶最弱,其中剑叶的叶绿素及类胡萝卜素含量降低最为明显;并且伴随着穗粒数、千粒重、结实率、株高等主要农艺性状的显著降低,但有效穗显著增多。透射电镜观察结果显示,与野生型相比,该突变体多数叶绿体结构基本完整,但基粒模糊,基质片层大量减少且排列疏松。遗传分析表明,该突变性状受一对隐性核基因控制。在yld突变体与粳稻武运粳7号杂交的F2群体中分离出323个突变单株,最终将YLD基因定位在第11染色体的L5和L7两标记之间,物理距离为115.7 kb。本研究为YLD基因的克隆和功能分析奠定了基础。
[本文引用: 1]
[本文引用: 1]
DOI:10.11983/CBB16010URL [本文引用: 1]
水稻(Oryza sativa)矮化是与光合效率及产量等密切相关的重要农艺性状。发掘更多的水稻矮秆资源,不仅能够进一步加深对水稻株高分子遗传机制的认识,而且还能为水稻新品种培育提供新的种质资源。在水稻T-DNA插入突变体库中筛选到1个矮化、宽叶小粒突变体(wld1)。经图位克隆将WLD1基因定位在第5号染色体长臂,位于分子标记In Del37与InDel48之间,基因编号为LOC_Os05g32270,属于AP2转录因子家族。该基因第6外显子处胸腺嘧啶缺失,造成转录提前终止。石蜡切片观察结果显示,茎部第2节间横向细胞数目增加,而纵向细胞数目未变。RT-PCR检测结果表明,LOC_Os05g32270在突变体wld1中不表达,造成功能缺失。该基因与已报道的水稻OsSMOS1(SMALL ORGAN SIZE1)为等位基因。水稻突变体wld1的矮秆遗传效应可直接应用于育种中。该研究结果进一步明确了突变体wld1的表型特征与遗传基础,为解析其参与的信号途径提供参考。
.
DOI:10.11983/CBB16010URL [本文引用: 1]
水稻(Oryza sativa)矮化是与光合效率及产量等密切相关的重要农艺性状。发掘更多的水稻矮秆资源,不仅能够进一步加深对水稻株高分子遗传机制的认识,而且还能为水稻新品种培育提供新的种质资源。在水稻T-DNA插入突变体库中筛选到1个矮化、宽叶小粒突变体(wld1)。经图位克隆将WLD1基因定位在第5号染色体长臂,位于分子标记In Del37与InDel48之间,基因编号为LOC_Os05g32270,属于AP2转录因子家族。该基因第6外显子处胸腺嘧啶缺失,造成转录提前终止。石蜡切片观察结果显示,茎部第2节间横向细胞数目增加,而纵向细胞数目未变。RT-PCR检测结果表明,LOC_Os05g32270在突变体wld1中不表达,造成功能缺失。该基因与已报道的水稻OsSMOS1(SMALL ORGAN SIZE1)为等位基因。水稻突变体wld1的矮秆遗传效应可直接应用于育种中。该研究结果进一步明确了突变体wld1的表型特征与遗传基础,为解析其参与的信号途径提供参考。
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1093/jxb/erw002URL [本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2017.09.001URL [本文引用: 1]
【目的】对一个水稻矮化剑叶卷曲突变体进行鉴定与基因定位,为水稻等禾谷类作物剑叶形态发育及分子改良奠定基础。【方法】在籼型水稻恢复系缙恢10号的甲基磺酸乙酯(EMS)突变库中筛选到一个隐性矮化剑叶卷曲突变体,命名为dcfl1(dwarf and curled flag leaf 1)。田间小区种植,全生育期内观察dcfl1和野生型的株型变化。苗期利用扫描电镜观察叶鞘内表皮细胞大小;孕穗期和抽穗期利用石蜡切片观察剑叶基部形态;开花期测定剑叶、倒2叶和倒3叶的叶绿素含量;成熟期考查株高、有效穗数、穗实粒数、结实率和千粒重等主要农艺性状。配制西农1A/dcfl1杂交组合,利用F1和F2群体进行遗传分析,并利用F2隐性群体进行基因定位。【结果】生育期内,突变体dcfl1都表现出矮化性状。dcfl1叶鞘内表皮细胞长度明显比野生型要短,达到了极显著水平。与野生型相比,穗长、倒1节间和倒2节间均显著变短,倒3节间和倒4节间无显著变化。抽穗期dcfl1剑叶的叶片和叶鞘连接处硬化,剑叶基部展开受阻,半边叶片向内卷曲,剑叶上部和中部正常,其他叶片也正常。农艺性状调查发现,dcfl1的有效穗数为14.24,极显著高于野生型的11.62,穗粒数、实粒数、结实率和千粒重等则无显著变化。此外,dcfl1的叶色略深,剑叶、倒2叶和倒3叶的叶绿素a含量均极显著高于野生型,类胡萝卜素含量也略有升高,但仅剑叶达到极显著差异水平,叶绿素b的含量则无显著变化。西农1A/dcfl1的F1群体中,株高和剑叶表型与野生型一致。F2群体中分离出正常和突变两种表型,突变表型与dcfl1类似,植株株高变矮,剑叶基部特异卷曲,说明矮化和剑叶基部特异卷曲是一对共分离性状。且两种表型分离比符合3﹕1,表明dcfl1突变型受1对隐性核基因控制。利用620株F2隐性单株,最终将DCFL1精细定位在第3染色体短臂In Del标记Ind03-11和In
DOI:10.3864/j.issn.0578-1752.2017.09.001URL [本文引用: 1]
【目的】对一个水稻矮化剑叶卷曲突变体进行鉴定与基因定位,为水稻等禾谷类作物剑叶形态发育及分子改良奠定基础。【方法】在籼型水稻恢复系缙恢10号的甲基磺酸乙酯(EMS)突变库中筛选到一个隐性矮化剑叶卷曲突变体,命名为dcfl1(dwarf and curled flag leaf 1)。田间小区种植,全生育期内观察dcfl1和野生型的株型变化。苗期利用扫描电镜观察叶鞘内表皮细胞大小;孕穗期和抽穗期利用石蜡切片观察剑叶基部形态;开花期测定剑叶、倒2叶和倒3叶的叶绿素含量;成熟期考查株高、有效穗数、穗实粒数、结实率和千粒重等主要农艺性状。配制西农1A/dcfl1杂交组合,利用F1和F2群体进行遗传分析,并利用F2隐性群体进行基因定位。【结果】生育期内,突变体dcfl1都表现出矮化性状。dcfl1叶鞘内表皮细胞长度明显比野生型要短,达到了极显著水平。与野生型相比,穗长、倒1节间和倒2节间均显著变短,倒3节间和倒4节间无显著变化。抽穗期dcfl1剑叶的叶片和叶鞘连接处硬化,剑叶基部展开受阻,半边叶片向内卷曲,剑叶上部和中部正常,其他叶片也正常。农艺性状调查发现,dcfl1的有效穗数为14.24,极显著高于野生型的11.62,穗粒数、实粒数、结实率和千粒重等则无显著变化。此外,dcfl1的叶色略深,剑叶、倒2叶和倒3叶的叶绿素a含量均极显著高于野生型,类胡萝卜素含量也略有升高,但仅剑叶达到极显著差异水平,叶绿素b的含量则无显著变化。西农1A/dcfl1的F1群体中,株高和剑叶表型与野生型一致。F2群体中分离出正常和突变两种表型,突变表型与dcfl1类似,植株株高变矮,剑叶基部特异卷曲,说明矮化和剑叶基部特异卷曲是一对共分离性状。且两种表型分离比符合3﹕1,表明dcfl1突变型受1对隐性核基因控制。利用620株F2隐性单株,最终将DCFL1精细定位在第3染色体短臂In Del标记Ind03-11和In
DOI:10.1111/jipb.12062URLPMID:23650998 [本文引用: 1]
Grain size is an important yield-related trait in rice. Intensive artificial selection for grain size during domestication is evidenced by the larger grains of most of today's cultivars compared with their wild relatives. However, the molecular genetic control of rice grain size is still not well characterized. Here, we report the identification and cloning of Grain Size 6 (GS6), which plays an important role in reducing grain size in rice. A premature stop at the +348 position in the coding sequence (CDS) of GS6 increased grain width and weight significantly. Alignment of the CDS regions of GS6 in 90 rice materials revealed three GS6 alleles. Most japonica varieties (95%) harbor the Type I haplotype, and 62.9% of indica varieties harbor the Type II haplotype. Association analysis revealed that the Type I haplotype tends to increase the width and weight of grains more than either of the Type II or Type III haplotypes. Further investigation of genetic diversity and the evolutionary mechanisms of GS6 showed that the GS6 gene was strongly selected in japonica cultivars. In addition, a 090008ggc090009 repeat region identified in the region that encodes the GRAS domain of GS6 played an important historic role in the domestication of grain size in rice. Knowledge of the function of GS6 might aid efforts to elucidate the molecular mechanisms that control grain development and evolution in rice plants, and could facilitate the genetic improvement of rice yield.
[本文引用: 1]
DOI:10.1105/tpc.112.097394URL [本文引用: 1]
DOI:10.1038/nplants.2017.43URLPMID:28394310 [本文引用: 1]
Abstract Grain size is a major determinant of grain yield in cereal crops. qSW5/GW5, which exerts the greatest effect on rice grain width and weight, was fine-mapped to a 2,263-bp/21-kb genomic region containing a 1,212-bp deletion, respectively. Here, we show that a gene encoding a calmodulin binding protein, located 5 b downstream of the 1,212-bp deletion, corresponds to qSW5/GW5. GW5 is expressed in various rice organs, with highest expression level detected in young panicles. We provide evidence that the 1,212-bp deletion affects grain width most likely through influencing the expression levels of GW5. GW5 protein is localized to the plasma membrane and can physically interact with and repress the kinase activity of rice GSK2 (glycogen synthase kinase 2), a homologue of Arabidopsis BIN2 (BRASSINOSTEROID INSENSITIVE2) kinase, resulting in accumulation of unphosphorylated OsBZR1 (Oryza sativa BRASSINAZOLE RESISTANT1) and DLT (DWARF AND LOW-TILLERING) proteins in the nucleus to mediate brassinosteroid (BR)-responsive gene expression and growth responses (including grain width and weight). Our results suggest that GW5 is a novel positive regulator of BR signalling and a viable target for genetic manipulation to improve grain yield in rice and perhaps in other cereal crops as well.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
矮泰引-3的矮生性状受两对独立遗传的半矮秆基因控制,利用 SSR标记将这两个矮秆基因分别定位到第1和第4染色体上.等位性测交的结果表明,位于第1染色体上的矮秆基因与sd1是等位的,所以仍然称其为sd1; 而位于第4染色体上的矮秆基因是一个新基因,暂命名为sdt2.利用SSR标记将sd1定位于RM297、RM302和RM212的同一侧,而与OSR3 共分离,它们之间的位置关系可能是RM297-RM302-RM212-OSR3-sd1,遗传距离分别为4.7cM、0cM、0.8cM和0cM,这与 sd1在第1染色体长臂上的确切位置是基本一致的.利用已有的SSR标记和拓展的SSR标记将sdt2定位于SSR332、RM1305和RM5633、 RM307、RM401之间,它们的排列位置可能是SSR332-RM1305-sdt2-RM5633-RM307-RM401,它们之间的遗传距离分 别为11.6cM、3.8cM、0.4cM、0cM和0.4cM.
URL [本文引用: 1]
矮泰引-3的矮生性状受两对独立遗传的半矮秆基因控制,利用 SSR标记将这两个矮秆基因分别定位到第1和第4染色体上.等位性测交的结果表明,位于第1染色体上的矮秆基因与sd1是等位的,所以仍然称其为sd1; 而位于第4染色体上的矮秆基因是一个新基因,暂命名为sdt2.利用SSR标记将sd1定位于RM297、RM302和RM212的同一侧,而与OSR3 共分离,它们之间的位置关系可能是RM297-RM302-RM212-OSR3-sd1,遗传距离分别为4.7cM、0cM、0.8cM和0cM,这与 sd1在第1染色体长臂上的确切位置是基本一致的.利用已有的SSR标记和拓展的SSR标记将sdt2定位于SSR332、RM1305和RM5633、 RM307、RM401之间,它们的排列位置可能是SSR332-RM1305-sdt2-RM5633-RM307-RM401,它们之间的遗传距离分 别为11.6cM、3.8cM、0.4cM、0cM和0.4cM.
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
.
[本文引用: 2]
.
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nplants.2015.170URLPMID:27251715 [本文引用: 1]
Abstract Manganese is an essential metal for plant growth. A number of transporters involved in the uptake of manganese from soils, and its translocation to the shoot, have been identified in Arabidopsis and rice. However, the transporter responsible for the radial transport of manganese out of root exodermis and endodermis cells and into the root stele remains unknown. Here, we show that metal tolerance protein 9 (MTP9), a member of the cation diffusion facilitator family, is a critical player in this process in rice (Oryza sativa). We find that MTP9 is mainly expressed in roots, and that the resulting protein is localized to the plasma membrane of exo- and endodermis cells, at the proximal side of these cell layers (opposite the manganese uptake transporter Nramp5, which is found at the distal side). We demonstrate that MTP9 has manganese transport activity by expression in proteoliposomes and yeast, and show that knockout of MTP9 in rice reduces manganese uptake and its translocation to shoots. We conclude that at least in rice MTP9 is required for manganese translocation to the root stele, and thereby manganese uptake.
DOI:10.1371/journal.pone.0174987URL [本文引用: 1]
.
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}