 ,1,*1
,1,*1 2
3
4
Molecular Cloning, Location and Expression Analysis of Brasscia oleracea Zinc Finger Protein Transcription Factor BoC2H2
LUO Shao-Lan1,**, LIAN Xiao-Ping2,**, PU Min1, BAI Xiao-Jing1, WANG Yu-Kui1, ZENG Jing4, SHI Song-Mei3, ZHANG He-Cui1, ZHU Li-Quan,1,*12
3
4
通讯作者:
第一联系人:
收稿日期:2018-02-12接受日期:2018-07-20网络出版日期:2018-07-30
| 基金资助: |
Received:2018-02-12Accepted:2018-07-20Online:2018-07-30
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (5045KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
罗绍兰, 廉小平, 蒲敏, 白晓璟, 王玉奎, 曾静, 施松梅, 张贺翠, 朱利泉. 甘蓝锌指蛋白转录因子BoC2H2的克隆、定位与表达分析[J]. 作物学报, 2018, 44(11): 1650-1660. doi:10.3724/SP.J.1006.2018.01650
LUO Shao-Lan, LIAN Xiao-Ping, PU Min, BAI Xiao-Jing, WANG Yu-Kui, ZENG Jing, SHI Song-Mei, ZHANG He-Cui, ZHU Li-Quan.
锌指蛋白(Zinc-finger protein, ZFP)是一种具有手指状结构域的转录因子, 研究表明, 锌指蛋白不仅在非生物胁迫例如低温、干旱、高盐以及病原菌防御的调控中发挥作用, 更是在形态建成、花粉发育、胚发育等过程参与调控[1,2,3,4]。最早在非洲爪蟾(Xenopus laevis)卵母细胞的转录因子中被发现[5]。目前已对拟南芥(Arabidopsis thaliana)、大豆(Glycine max)、矮牵牛(Petunia hybrida)、小麦(Triticum aestivum)、水稻(Oryzas ativa)、棉花(Anemone vitifolia)等植物中的一些锌指蛋白做了研究[6]。已将锌指蛋白按照半胱氨酸(C)和组氨酸(H)的数目及位置分为C2C2、C2H2、C2HC、C2HC5、CCCH、C3HC4、C4、C4HC3、C6、C8等多种类型[7,8,9,10,11,12]。其中C2H2型(Cys2/His2)成员最多, 也是研究最为深入的一类锌指蛋白[13,14]。
目前报道的植物C2H2型锌指蛋白主要参与植物各个时期的生长发育、病原菌防御及环境胁迫下基因的表达调控[15,16]。例如CgZFP1是菊花中的一个C2H2型锌指蛋白, 过表达该基因的拟南芥能提高抗盐和抗旱能力[17]; 甘蓝型油菜中大量的BnWRKY蛋白参与防御相关基因的转录调控, 以响应真菌病原体和激素刺激[18], 同时WRKY转录因子在对环境应激刺激的反应中也起重要作用[19]; 甘蓝中的部分C2H2型锌指蛋白与耐盐胁迫相关[20]; 在拟南芥中, ZAT6不仅参与调节磷酸盐的代谢和根的发育, 还介导盐和渗透胁迫的响应[21,22]。AZF2在种子萌发过程中是ABA信号的一个负调控因子[23]; 甘蓝型油菜中编码锌指蛋白转录因子的基因调控种皮发育和种皮色素合成与积累[24]。ZAT11调控百草枯诱导的细胞程序性死亡[25]。番茄中的SlZHD12和SlZHD21分别在花蕾和完全开放的花中高表达[26]; 白菜中的大多数BraZF-HD都在花中表达[27]; 甘蓝型油菜中C2H2型锌指蛋白部分基因在根、初花期雌蕊和角果中特异表达[28]; 矮牵牛中有7个锌指蛋白主要在花药中表达[29]; 在TAZ1被抑制表达的转基因植株中, 出现大量的败育花粉粒, 少数未败育花粉粒也由于不能形成正常花粉壁其萌发率极低[30]; 十字花科植物中的C2H2型锌指蛋白参与花粉发育[31]; 拟南芥SUPERMAN锌指蛋白在花发育中具有重要的作用[32]; BnVRN2a参与开花相关途径[33]。甘蓝中共鉴定了148个WRKY基因, 这些基因在各种组织, 即根、茎、叶、芽、花和长角果均有表达, 并且其中大部分表现出差异和组织特异性表达, 表明这些基因可能在植物发育过程中发挥重要作用[34]。
自交不亲和性(self-incompatibility, SI)是植物限制自交衰败和促进杂交优势, 在长期进化过程中形成的一种复杂而完善的重要遗传机制。甘蓝属于典型的孢子体自交不亲和植物, 迄今为止, 有关甘蓝自交不亲和相关的锌指蛋白还未见报道。本文以高代自交不亲和系甘蓝‘A4’和‘F1’为材料, 对处于花期的‘A4’材料进行自花和异花授粉处理, 取未授粉和授粉15、30和60 min后的柱头进行转录组测序分析, 筛选到1个表达差异显著的C2H2型锌指蛋白基因, 命名为BoC2H2 (Brassica oleracea Cys2/His2), 说明该基因可能参与了柱头响应花粉刺激的分子过程。到目前为止, 关于C2H2型锌指蛋白是否参与自交不亲和信号传导过程还是未知的。因此进一步对BoC2H2基因进行了克隆、序列分析、亚细胞定位、表达分析及启动子活性分析, 以期为研究C2H2型转录因子在甘蓝自交不亲和过程中的调控机制提供参考依据。
1 材料与方法
1.1 试验材料和菌种
以西南大学十字花科蔬菜研究所选育的自交不亲和甘蓝高代自交系‘A4’和‘F1’植株为材料。2017年3月底至4月初, 选取开花前1~2 d长势一致的‘A4’花蕾, 于开花当天人工去雄, 用成熟的‘A4’和‘F1’花粉对‘A4’柱头进行自花和异花授粉处理。分别取未授粉柱头和自花、异花授粉15、30和60 min的柱头, 立刻放入液氮速冻保存。同时取萼片、花瓣、花丝、花粉和柱头于-80℃保存备用。大肠杆菌DH5α购自北京康为世纪公司, 农杆菌EHA105、GV3101购自北京全式金生物技术有限公司。1.2 SI甘蓝自花、异花授粉后柱头转录组数据分析
将未授粉柱头, 自花授粉15、30和60 min和异花授粉15、30和60 min的甘蓝柱头送北京百迈克生物科技有限公司基于Illumina HiSeq 2000测序技术平台测序, 获得转录组数据。采用FPKM (Fragments Per Kilobase of transcript per Million fragment mapped)量化基因的表达水平[35]。使用EBSeq进行差异表达分析, 以差异倍数Fold Change≥2或≤0.5及错误发现率(False Discovery Rate) FDR<0.01作为筛选标准[36]。根据基因在不同授粉处理后的相对表达量, 筛选在自花授粉后表达差异明显、异花授粉后表达稳定的基因。1.3 差异基因的克隆验证
根据转录组测序获得的差异基因的cDNA序列和芸薹属数据库(http://brassicadb.org/brad/), 并且结合GenBank已登录植物的C2H2核苷酸序列进行多序列比对, 发现该类基因在不同植物中高度保守, 根据3°端和5°端的保守序列用Primer primer 6.0软件以及GFP融合表达载体1300-GFP (酶切位点为Xba I和Sac I)序列设计引物1300-GFP-F和1300-GFP-R (表1)。分别以甘蓝柱头gDNA和自花授粉30 min柱头cDNA为模板, 利用PrimerSTAR Max DNA Polymerase (Takara, 北京)进行扩增。PCR产物经1×104 mg L-1琼脂糖凝胶电泳, 胶回收后运用快速克隆技术将目的片段连接到pET-15b-1载体上后转化大肠杆菌DH5α, 经菌落PCR鉴定后挑取阳性克隆送北京华大基因公司进行测序。Table 1
表1
表1基因克隆及其荧光定量 PCR 分析所用引物
Table 1
| 引物名称 Primer | 引物序列 Primer sequence (5°-3°) | 引物说明 Primer annotation |
|---|---|---|
| 1300-GFP-F | GAGAACACGGGGGACTCTAGAATGAGTGATCCCGAGAAAACAAAAG | 基因的亚细胞定位 |
| 1300-GFP-R | GCCCTTGCTCACCATGAGCTCCTCGGCTTTGTCCTCTTTTGC | Subcellular localization |
| 35S-R | CCGATCTAGTAACATAGATGACACCG | 通用引物 Universal primer |
| Gus-F | CAAGCTTGGCTGCAGGTCGACTTCGACTCAGCGTGTTATG | 基因的启动子活性分析 |
| Gus-R | GGTGGACTCCTCTTAGAATTCGATTTTGACTTTGTTGAGAG | Promoter activity analysis |
| 1391-F | GAACTGATCGTTAAAACTGC | 通用引物 |
| 1391-R | TGGTCTTCTGAGACTGTATC | Universal primer |
| RT-PCR-F | GTGAACACTGAGGAAAGAATTAATG | 荧光定量PCR引物 |
| RT-PCR-R | CAACCTCTTGTTCCATGTTGTC | Primers for real-time PCR |
| Actin3-F | GAGTAGAAAATGGCTGATGGTGAAG | 扩增内参基因 |
| Actin3-R | TCATCTTCTCACGGTTAGCCTTTG | For the internal control |
新窗口打开|下载CSV
1.4 生物信息学分析
利用Seqman软件对基因全长进行拼接, 用Editseq进行ORF查询; 用GSDS分析基因结构; 用ProtParam分析基因编码蛋白质的理化性质; 用Signalp 4.1 server预测编码蛋白质的信号肽; 用TMHMM Server v.2.0进行跨膜结构预测; 用SOPMA软件分析蛋白质的二级结构; 用Netphos 2.0 server预测磷酸化位点; 用ProtScale预测疏水性/亲水性; 用SMART网站分析蛋白质的结构域; 用Cell-PLoc预测亚细胞定位; 通过PlantCARE分析启动子的顺式作用元件。在NCBI数据库中分别用Blast N和Blast X对核酸序列和氨基酸序列进行同源性分析; 用Version 6.0、ClustalX1.8和MEGA 6进行多序列比对并构建系统进化树。1.5 亚细胞定位表达分析
将测序正确的GFP融合表达菌株扩大培养后抽提重组质粒, 一方面采用热激法将重组质粒转至EHA105感受态细胞中[37], 用引物1300-GFP-F和通用引物35S-R进行PCR检测, 并将阳性转化子单克隆按1︰100扩大培养。选取四至六叶期健壮的本氏烟, 将含阳性克隆的农杆菌重悬液采用注射法侵染本氏烟叶片[38], 48 h后将注射孔部位的叶片表皮组织制作成临时玻片, 用激光共聚焦荧光显微镜通过观察本氏烟表皮细胞内绿色荧光来确定BoC2H2- 1300-GFP融合蛋白的定位情况。另一方面将重组质粒用PEG法转入拟南芥原生质体并融合表达, 在培养箱黑暗培养16 h后用激光共聚焦荧光显微镜观察绿色荧光来确定BoC2H2-1300-GFP融合蛋白的定位情况。1.6 表达分析
通过半定量PCR检测BoC2H2在萼片、花瓣、花丝、花粉和柱头等组织中的表达量, 依据Samuel[39]和Chandna[40]对候选内参基因的评价分析结果, 以Actin3作为内参, 引物为Actin3-F/R、RT-PCR-F/R (表1), 按照DNA聚合酶PrimeSTAR说明书操作。PCR扩增产物经1×104 mg L-1琼脂糖凝胶电泳检测, 分析BoC2H2基因在萼片、花瓣、花丝、花粉和柱头5种组织中的表达情况。根据获得的BoC2H2基因序列设计荧光定量PCR特异性引物Actin3-F/R、RT-PCR-F/R (表1), 以未授粉柱头, 自花授粉15、30、60 min和异花授粉15、30、60 min的柱头cDNA为模板, Actin3为内参。Bio-Rad SsoFast EvaGreen Supermix为荧光定量PCR染料, 反应体系为10 μL, 在Bio-Rad CFX-1000荧光定量PCR仪上对目的基因进行荧光定量PCR反应。反应参数95℃ 30 s; 95℃ 10 s, 57℃ 10 s, 72℃ 10 s, 40个循环, 每个循环结束时采集荧光信号。40个循环后对PCR扩增产物进行溶解曲线分析。反应完成后利用Bio-Rad CFX Manager Software和Microsoft Excel软件分析数据。
1.7 BoC2H2启动子活性分析
在NCBI选取BoC2H2基因起始密码子上游2000 bp左右的核苷酸序列, 载体为pCAMBIA 1391, 酶切位点为SalI和EcoRI, 设计引物为GUS-F/R (表1)。按照同源重组的方法构建BoC2H2-GUS融合表达载体。将构建好的融合表达载体转化农杆菌GV3101感受态细胞, 用通用引物1391-F/R (表1)进行PCR检测, 并将阳性转化子单克隆按1︰100扩大培养, 并通过遗传转化获得阳性拟南芥植株。通过筛选将得到的纯合转基因拟南芥种子播种, 分别取幼苗、茎和花, 置GUS染色液中, 37℃培养箱中温育过夜。将侵染过的样品转入70%酒精中脱色2~3次, 然后于体视显微镜下观察照相。2 结果与分析
2.1 BoC2H2基因筛选
为分离甘蓝SI相关的调控基因, 本研究以甘蓝‘A4’未授粉柱头和自花、异花授粉处理的柱头为材料, 进行转录组测序和分析。共获得60.25 Gb Clean Data, 各样品的Clean Data均达到7.31 Gb, Q30碱基百分比在93.06%以上, 可认为测序质量可靠。从中筛选差异表达基因, 进而筛选到1个受自花授粉诱导大量上调表达的基因。通过GO基因功能注释结果显示, 该基因对应的蛋白为锌指蛋白, 进一步通过NCBI序列比对发现, 其拟南芥同源基因AT4G35700为C2H2型锌指蛋白基因。由此将该基因命名为BoC2H2 (Brassica oleraceaCys2/His2)。BoC2H2基因在0 min的表达量约为0.90, 在自花授粉15、30、60 min后该基因表达量分别约为7.44、21.50、46.24, 显著上调表达; 而在异花授粉15、30、60 min后其表达量分别约为3.46、3.74、4.06, 其表达量趋于稳定(图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1柱头内BoC2H2响应自花和异花授粉后表达模式
0: 未授粉柱头。0: unpollinated stigma.
Fig. 1Expression pattern of BoC2H2 in stigma in response to self-pollination and cross-pollination
2.2 甘蓝BoC2H2基因的克隆与生物信息学分析
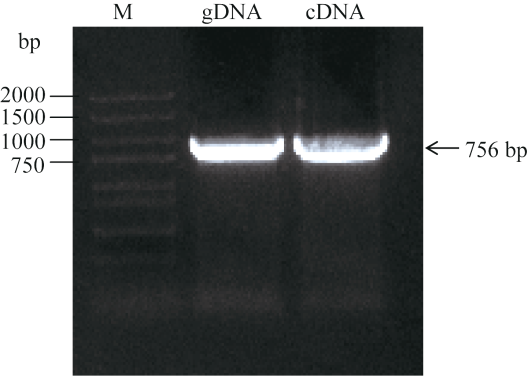
根据转录组测序获得的BoC2H2基因的cDNA序列和芸薹属数据库, 并且结合GenBank已登录植物的C2H2核苷酸序列进行多序列比对设计引物。以‘A4’柱头cDNA和gDNA为模板, 扩增获得的甘蓝BoC2H2基因。结果显示, 扩增得到的BoC2H2基因的gDNA和cDNA全长均为756 bp (图2), 包含了BoC2H2基因完整的开放阅读框, 编码251个氨基酸残基, 登录号为XP_013591069, 其编码蛋白分子量为26.7 kDa, 等电点为4.62。结合GSDS在线工具分析基因组结构结果说明, 该基因没有内含子, 结果与转录组测序获得的cDNA序列一致。在NCBI数据库中进行Blast比对, 发现BoC2H2基因序列与白菜BrC2H2、油菜BnC2H2和萝卜RsC2H2基因的序列高度保守, 同源性88%以上。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2甘蓝BoC2H2基因gDNA和cDNA PCR序列扩增
M: DL2000; gDNA: BoC2H2基因gDNA全长扩增条带; cDNA: BoC2H2基因cDNA全长扩增条带。
Fig. 2Amplification of BoC2H2gene from gDNA and cDNA of the stigma of Brassica oleracea
M: DL2000; gDNA: products of BoC2H2 gene gDNA; cDNA: products of BoC2H2 gene cDNA.
通过分析发现BoC2H2蛋白质无跨膜结构域和信号肽, 属于亲水性蛋白, 定位于细胞核和质膜。二级结构分析表明含有α-螺旋(alpha helix) 37.05%、β折叠(beta turn) 9.16%、无规则卷曲(random coil) 41.43%和延伸链(extended strand) 12.35%。在氨基酸组成中, Glu、Ala和Gly出现频率较高, 分别占氨基酸总数的11.55%、11.16%和9.96%, 而氨基酸Met、Cys和Trp、Tyr、His分别仅占氨基酸总数的1.59%和0.80%。带负电荷的氨基酸(Asp+Glu)残基有49个, 带正电荷的氨基酸(Arg+Lys)残基有31个。BoC2H2蛋白质包含16个丝氨酸磷酸化位点、6个苏氨酸磷酸化位点、1个酪氨酸磷酸化位点。BoC2H2蛋白质含有C2H2型锌指蛋白家族高度保守的ZnF_ C2H2结构域(图3)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3BoC2H2核苷酸序列及对应的氨基酸序列
实线框表示ZnF_C2H2结构域。The box was ZnF_C2H2 domain.
Fig. 3Nucleotide sequence of BoC2H2 and its amino acid sequence
将推导的BoC2H2蛋白的氨基酸序列与8种已知植物C2H2进行多序列比对, 并采用NJ法进行BoC2H2氨基酸序列系统进化树分析。由图4可知, 在已知的8种植物C2H2型锌指蛋白中, 甘蓝BoC2H2与白菜和油菜C2H2型锌指蛋白的进化同源性最近, 其序列相似度分别为90%和89%。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4BoC2H2与其他物种C2H2氨基酸序列的系统进化树
Fig. 4Phylogenetic tree of BoC2H2 and C2H2 amino acid sequence in other species
利用甘蓝基因组(http://brassicadb.org/brad/index. php)及PlantCARE启动子分析软件在位于BoC2H2基因起始密码子上游2000 bp的核苷酸序列中预测到光响应、昼夜节律、茉莉酸响应、生长素应答、蛋白代谢、胚乳表达、防御和应激反应等多种顺式作用元件(表2), 表明该基因除了能响应授粉刺激, 还可能响应多种信号。
Table 2
表2
表2BoC2H2基因上游调控区顺式作用元件
Table 2
| 相关功能预测Associated putative function | 启动子顺式作用元件 cis-elements in the promoter region |
|---|---|
| Light responsive element | CATT-motif, GAG-motif, Gap-box |
| Circadian control | Circadian |
| Auxin-responsive element | TGA-element |
| Gibberellin-responsive | GARE-motif |
| Zein metabolism regulation | O2-site |
| Meristem expression | CAT-box |
| Endosperm expression | GCN4_motif, Skn-1_motif |
| Defense and stress responsiveness | TC-rich repeats |
| Elicitor-responsive element | EIRE, ELI-box3 |
| Essential for the anaerobic induction | ARE |
| Promoter and enhancer regions | CAAT-box |
| MeJA-responsiveness | CGTCA-motif, TGACG-motif |
| Anoxic specific inducibility | GC-motif |
| Low-temperature responsiveness | LTR |
| MYB binding site involved in drought-inducibility MYB | MBS |
| Core promoter element around -30 of transcription start | TATA-box |
| Salicylic acid responsiveness | TCA-element |
新窗口打开|下载CSV
2.3 BoC2H2亚细胞定位
在线软件(http://www.csbio.sjtu.edu.cn/bioinf/ Cell-PLoc/)预测BoC2H2定位于细胞核和质膜, 利用农杆菌介导在本氏烟叶片细胞中和在拟南芥原生质体中表达BoC2H2-1300-GFP融合蛋白, 在激光共聚焦显微镜下观察GFP绿色荧光。对照为无定位功能的1300-GFP蛋白。结果显示, 与1300-GFP空载对照相比, BoC2H2-1300-GFP融合蛋白在烟草叶片细胞和拟南芥原生质体中的绿色荧光信号都分布在细胞核和细胞质中(图5和图6), 由此可见, 甘蓝BoC2H2蛋白位于细胞核和细胞质中。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5BoC2H2-1300-GFP融合蛋白在烟草表皮细胞中的亚细胞定位
A: 1300-GFP载体的亚细胞定位(对照); B: BoC2H2-1300-GFP的亚细胞定位; 箭头所指为细胞核位置。
Fig. 5Subcellular location of BoC2H2-1300-GFP in epidermal cells of tobacco
A: subcellular location of 1300-GFP vector; B: subcellular location of BoC2H2-1300-GFP; the arrows indicate the location of nucleus.
图6
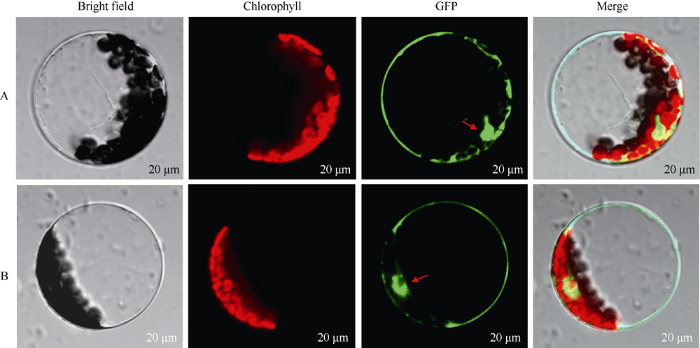 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6BoC2H2-1300-GFP融合蛋白在拟南芥原生质体中的亚细胞定位
A: 1300-GFP载体的亚细胞定位(对照); B: BoC2H2-1300-GFP的亚细胞定位; 箭头所指为细胞核位置。
Fig. 6Subcellular location of BoC2H2-1300-GFP in epidermal cells of Arabidopsis thaliana
A: subcellular location of 1300-GFP vector; B: subcellular location ofBoC2H2-1300-GFP; the arrows indicate the location of nucleus.
2.4 BoC2H2组织特异性分析
半定量PCR结果表明, BoC2H2基因在甘蓝花的不同组织中均有表达, 且表达量有差异(图7)。在花丝中的表达量最高, 柱头和花药中次之, 而萼片和花瓣中相对较低。图7
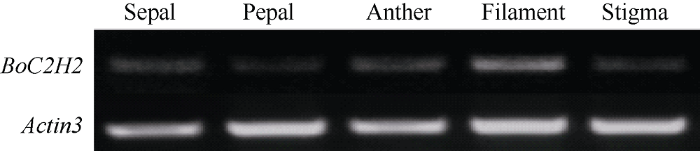 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7甘蓝BoC2H2基因在不同组织中的表达分析
Fig. 7Expression analysis of BoC2H2 in different organs of Brassica oleracea
2.5 BoC2H2启动子的特异性分析
在NCBI下载BoC2H2基因起始密码子上游2000 bp左右的核苷酸序列, 用同源重组的方法将载体pCAMBIA 1391和BoC2H2基因的cDNA序列连接, 获得BoC2H2-GUS融合表达载体。进一步将BoC2H2-GUS和pCAMBIA 1391载体分别通过遗传转化得到的T1代拟南芥种子, 用含有潮霉素的MS培养基筛选, 分别将其中生长正常的植株移栽, 3~4 d后取其叶片提取DNA, 利用GUS通用引物检测转化苗。由图8可知, 植株1和植株2为BoC2H2-GUS转基因植株, 并且都扩增得到了大小约为2000 bp的条带; 植株3为pCAMBIA 1391载体转基因植株, 并且扩增得到了大小约为1800 bp的条带, 而野生型拟南芥没有扩增到该片段, 说明得到的3棵拟南芥植株均为阳性苗。待3棵植株成熟以单个果荚收种, 并在含有潮霉素的平板上播种, 植株1和植株3的种子全部发芽, 而植株2的种子只有部分发芽, 说明植株1为纯合BoC2H2-GUS转基因植株, 植株3为纯合pCAMBIA 1391转基因植株。图8
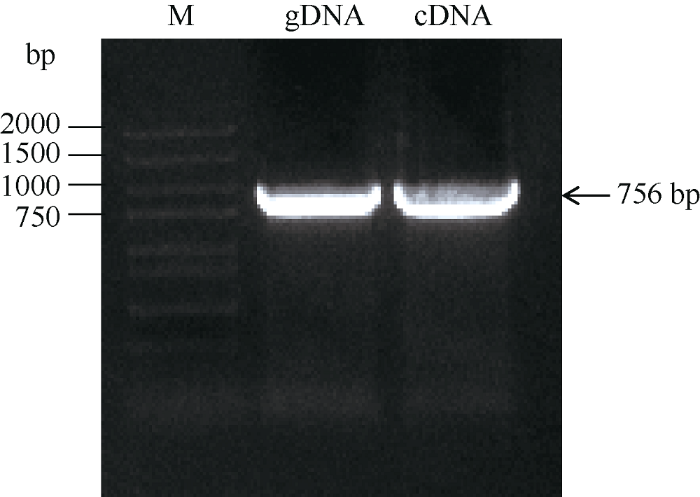 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8转基因植株PCR检测
1, 2条带为BoC2H2-GUS转基因植株, 3条带为pCAMBIA 1391转基因植株, WT为野生型拟南芥。
Fig. 8PCR analysis of transgenic plants
1, 2, and 3 bands were BoC2H2-GUS and pCAMBIA 1391 transgenic plants, respectively. WT was wild-type Arabidopsis thaliana.
选取上述已纯合的转基因植株1和植株3的果荚, 通过干燥、春化后, 一部分在MS培养基上播种, 一部分在土里播种, 分别对不同发育时期的不同组织进行GUS染色分析。结果发现, pCAMBIA 1391转基因植株在不同发育时期均未检测到GUS信号。BoC2H2-GUS转基因植株GUS染色结果如图9所示, 在幼苗期, BoC2H2主要在子叶和下胚轴表达, 而在下胚轴的维管组织中的表达量最高(图9-a, b); 在叶片中主要在成熟叶的叶脉中表达(图9-c~f); 在花中表达主要集中在柱头、花丝和萼片, 在整个花的发育过程中, 在花丝和萼片中都是高表达, 在柱头中的表达量随着发育时间而变化, 于开花当天后表达量降低, 并且主要集中在柱头乳突细胞以下的基本组织和维管组织中(图9-g~k); 在花粉中并没有表达(图9-i~k), 这与BoC2H2基因在甘蓝的花的组织中的表达情况不一致, 可能是在取甘蓝花药过程中将一部分花丝带入的原因, 因此BoC2H2基因在花药中并没有表达。在果荚中表达主要集中在离层区, 并且在果荚的发育过程中都是高表达(图9-l, m)。
图9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9GUS染色分析
a~b: 幼苗不同发育时期; c~f: 叶子不同发育时期; g~k: 花的不同发育时期; l~m: 果荚不同发育时期。
Fig. 9GUS staining analysis
a-b: seedlings at different developmental stages; c-f: leaves at different developmental stages; g-k: flowers at different developmental stages; l-m: pods at different developmental stages.
2.6 BoC2H2基因在自花和异花授粉后的表达分析
利用荧光定量PCR分析BoC2H2基因在自花和异花授粉后不同时间柱头内的表达情况表明, BoC2H2基因在自花授粉过程中, 0~60 min持续显著上调表达; 在异花授粉过程中, 0~15 min BoC2H2基因上调表达, 在15~60 min其表达量有变化, 但是变化并不明显(图10)。该结果与转录组分析结果基本一致。甘蓝自交不亲和过程主要是在柱头中发生, 这进一步验证自花授粉能强烈诱导该基因在甘蓝柱头中的表达, 表明BoC2H2基因响应甘蓝自花授粉后的反应。图10
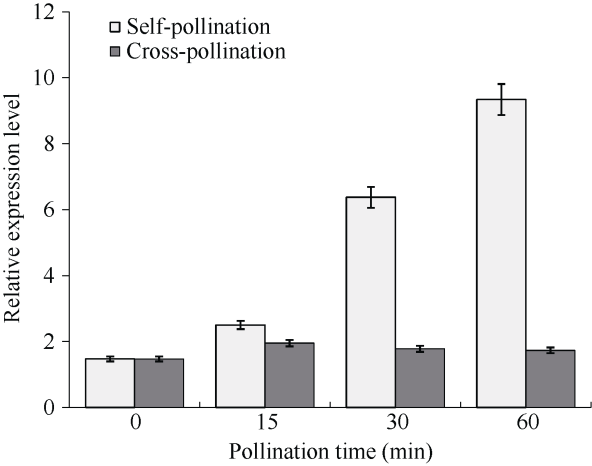 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10甘蓝柱头内BoC2H2响应自花和异花授粉后表达模式
Fig. 10Expression pattern of BoC2H2 in stigmas in response to self-pollination and cross-pollination
3 讨论
本研究基于以自交不亲和甘蓝不同授粉处理后的转录组文库, 筛选出一个受自花授粉诱导显著上调表达的C2H2型锌指蛋白基因BoC2H2。授粉诱导表达分析表明, BoC2H2基因在自花授粉0~60 min持续显著上调表达, 而在异花授粉0~60 min表达量变化并不明显, 说明自花授粉能显著诱导该基因的表达。BoC2H2蛋白质含有一个C2H2型锌指蛋白家族高度保守的ZnF_C2H2结构域, 表明该基因可能参与甘蓝的生长发育及环境胁迫下基因的表达调控[16]。BoC2H2-1300-GFP融合蛋白绿色荧光主要出现在细胞核和细胞质区域, 与相关基因的研究结果一致[41,42,43,44]。BoC2H2基因的启动子区域含有光响应、昼夜节律、茉莉酸响应、蛋白代谢、胚乳表达、防御和应激反应等多种顺式作用元件, 表明BoC2H2基因可能参与调控复杂的生物反应过程。BoC2H2在幼苗下胚轴的维管组织中高表达, 并且在子叶和成熟叶片的叶脉中也有表达, 表明BoC2H2可能参与植物水分和营养物质的运输和吸收。Ishigoro等[45]提出花丝可能是花中茉莉酸的主要来源, 茉莉酸可以调控雄蕊和花瓣的水分运输。BoC2H2不仅在花丝中高表达, 并且其启动子区域含有茉莉酸响应元件, 说明BoC2H2可能通过花丝中合成的茉莉酸调控雄蕊和花瓣的发育。在柱头中的表达量随着发育时间而变化, 于开花当天后表达量降低, 甘蓝自交不亲和反应主要在自花授粉后的柱头中发生, 且在开花后0.5 h~1.0 h之内完成[46], 进一步表明该基因可能参与自花授粉后花粉与柱头相互作用的复杂反应过程。
Jiang等[46]对授粉后不同时间段的雌蕊用SEM观察, 得到白菜花粉的粘附和水合分别在授粉后0.5 h和1.0 h。甘蓝等芸薹属作物属于典型孢子体型自交不亲和, 通常有一个干型柱头, 只有亲和花粉落到柱头后能诱导柱头释放花粉水化、萌发所需水分和其他因子, 不亲和花粉落到柱头后则在水化前或者穿过柱头前就被抑制[47]。本试验通过转录组测序和荧光定量分析结果显示, BoC2H2基因自花授粉后显著上调表达, 而异花授粉表达量较低且趋于稳定。BoC2H2基因可能在自花授粉0~30 min之间调控花粉黏附, 在30~60 min抑制花粉的水合, 所以表达量趋于上调。这些结果进一步表明, BoC2H2在自花花粉黏附、水合、萌发和花粉管伸长的过程中发挥作用, 进而参与调控甘蓝的自交不亲和过程。在今后的研究中, 我们将利用超表达和基因突变等技术进一步验证该基因的功能及其作用机制。
4 结论
从高代自交不亲和甘蓝‘A4’材料自花和异花授粉处理的柱头转录组数据中筛选到一个差异表达的基因BoC2H2, 该基因的开放阅读框为756 bp, 编码251个氨基酸, 含有C2H2型锌指蛋白家族高度保守的ZnF_C2H2结构域, 其编码蛋白大小为26.7 kDa。BoC2H2蛋白分布于细胞核和细胞质。BoC2H2基因在下胚轴、叶片和花中都有表达, 且柱头中的表达量随发育时间而变化, 于开花当天后表达量降低。该基因在自花授粉过程中, 0~60 min持续显著上调表达; 在异花授粉过程中表达量变化并不明显。初步推断BoC2H2参与了柱头响应花粉刺激的分子过程, 这有利于揭示BoC2H2在甘蓝自交不亲和过程中的作用机制, 为研究C2H2型转录因子在甘蓝自交不亲和过程中的调控机制提供参考依据。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/s00018-007-7473-4URL [本文引用: 1]
DOI:10.1111/j.1399-3054.2008.01090.xURLPMID:18346071 [本文引用: 1]

Reactive oxygen species (ROS) play a dual role in plant biology acting on the one hand as important signal transduction molecules and on the other as toxic by-products of aerobic metabolism that accumulate in cells during different stress conditions. Because of their toxicity as well as their important signaling role, the level of ROS in cells is tightly controlled by a vast network of genes termed the 'ROS gene network'. Using mutants deficient in key ROS-scavenging enzymes, we have defined a signaling pathway that is activated in cells in response to ROS accumulation. Interestingly, many of the key players in this pathway, including different zinc finger proteins and WRKY transcription factors, are also central regulators of abiotic stress responses involved in temperature, salinity and osmotic stresses. Here, we describe our recent findings and discuss how ROS integrate different signals originating from different cellular compartments during abiotic stress.
[本文引用: 1]
DOI:10.1111/jipb.2012.54.issue-7URL [本文引用: 1]
.
[本文引用: 1]
DOI:10.3321/j.issn:0253-9772.2004.03.029URL [本文引用: 1]
锌指蛋白是一类具有指状结构域的转录因子。根据半胱氨酸(C)和组氨酸(H)残基的数目和位置可将锌指蛋白分为C2H2、C2HC、C2C2、C2HCC2C2、C2C2C2C2等亚类。C2H2型锌指蛋白是最多也是研究最为清楚的一类锌指蛋白,在植物中已经克隆了50多个,主要涉及植物的生长发育和对环境胁迫的应答反应。该类锌指蛋白大部分在锌指区具有植物中特有的QALGGH保守结构,可能涉及调控植物特有的生物学功能。文章主要讨论了植物C2H2型锌指转录因子的结构、对靶DNA的识别及在生长发育和环境胁迫反应中可能的调控功能。
DOI:10.3321/j.issn:0253-9772.2004.03.029URL [本文引用: 1]
锌指蛋白是一类具有指状结构域的转录因子。根据半胱氨酸(C)和组氨酸(H)残基的数目和位置可将锌指蛋白分为C2H2、C2HC、C2C2、C2HCC2C2、C2C2C2C2等亚类。C2H2型锌指蛋白是最多也是研究最为清楚的一类锌指蛋白,在植物中已经克隆了50多个,主要涉及植物的生长发育和对环境胁迫的应答反应。该类锌指蛋白大部分在锌指区具有植物中特有的QALGGH保守结构,可能涉及调控植物特有的生物学功能。文章主要讨论了植物C2H2型锌指转录因子的结构、对靶DNA的识别及在生长发育和环境胁迫反应中可能的调控功能。
DOI:10.1126/science.271.5252.1081URLPMID:8599083 [本文引用: 1]
Zinc ions are key structural components of a large number of proteins. The binding of zinc stabilizes the folded conformations of domains so that they may facilitate interactions between the proteins and other macromolecules such as DNA. The modular nature of some of these zinc-containing proteins has allowed the rational design of site-specific DNA binding proteins. The ability of zinc to be bound specifically within a range of tetrahedral sites appears to be responsible for the evolution of the wide range of zinc-stabilized structural domains now known to exist. The lack of redox activity for the zinc ion and its binding and exchange kinetics also may be important in the use of zinc for specific functional roles.
DOI:10.1093/bfgp/1.4.342URLPMID:15239882 [本文引用: 1]
Within the last 20 years, the understanding of the biology of the 'classical' or Cys(2)His(2) zinc finger domain has progressed rapidly from the initial identification of the zinc finger as a repetitive zinc-binding motif in transcription factors to its use in biotechnology. The domain is the most abundant DNA-binding motif in the human genome and is a component of many key eukaryotic transcription factors involved in growth and development. Numerous structures now exist for this domain and its mode of action is known in a variety of zinc finger-DNA complexes. Application of this knowledge has led to the development of 'designer' transcription factors where zinc fingers have been engineered to bind desired DNA sequences. Recently, advances have been made in this field that potentially allow the targeting of any DNA site. Consideration of chromatin structure and the use of effector domains in these 'designer' transcription factors have made possible the regulation of a number of endogenous genes. These advances in the customised regulation of genes will be discussed in detail, as well as the potential to use these proteins in functional genomics and gene therapy applications.
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
.
[本文引用: 1]
DOI:10.1007/PL00000885URLPMID:11361095 [本文引用: 1]
C 2 H 2 zinc finger proteins probably comprise the largest family of regulatory proteins in mammals. Most zinc fingers bind to a cognate DNA. In addition to DNA, many of the proteins also bind to RNA or protein, and some bind to RNA only. The binding properties depend on the amino acid sequence of the finger domains and of the linker between fingers, as well as on the higher-order structures and the number of fingers. C 2 H 2 zinc finger proteins contain from 1 to more than 30 figures. Based on the number and the pattern of the fingers, most of the proteins can be classified into one of three groups: triple-C 2 H 2 , multiple-adjacent-C 2 H 2 , and separated-paired-C 2 H 2 finger proteins. In contrast to proteins with triple-C 2 H 2 fingers, proteins with multiple-adjacent-C 2 H 2 fingers can bind multiple, different ligands. Proteins with a number of separated-paired fingers bind to the target by means of only a single pair.
DOI:10.1007/s11103-007-9199-yURL [本文引用: 1]
DOI:10.1073/pnas.0806019105URL [本文引用: 1]
[本文引用: 2]
.
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
DOI:10.7666/d.Y2360507URL [本文引用: 1]
芸薹属植物在我国种植面积大,分布范围广,其中包括白菜、甘蓝、芥菜等重要的蔬菜作物。蔬菜作物在整个生长过程中不可避免的要受到各种环境条件的影响,如高温、干旱、盐胁迫等。本文通过对大白菜、不结球白菜、甘蓝等供试材料的苗期进行耐盐性筛选、苗期生理指标的测定、目的基因的克隆及其序列分析。获得如下研究结果: 1芸薹属植物耐盐相关生理性状的分析 水培盐处理结果表明,盐胁迫下的大白菜、不结球白菜、甘蓝的生物量与对照相比都有不同程度的降低;叶片相对电导率,随着处理时间的延长有明显的下降趋势;也有个别品种随处理时问的延长,其相对电导率无明显变化;净光合速率受抑制程度不明显,与对照相比,个别品种表现出盐处理促进了光合作用;胞间CO2浓度除个别品种与对照变化趋势一致外,多数品种都有不同程度的降低;气孔导度,蒸腾速率均表现出不同程度的降低趋势。 2芸薹属植物耐盐相关基因的克隆 以大白菜、不结球白菜、甘蓝为试材,通过拟南芥耐盐基因(AtSTZ)全长序列和其近缘物种同源序列设计引物TNS1,利用同源克隆方法分离出耐盐相关基因BcSTZ、 BSTZ,、QSTZ各4条序列。对其序列进行比对分析,与已发现的大量植物中的C2H2型锌指蛋白保守基序具有很高的同源性。含有两个由21个氨基酸组成的C2H2保守基序,每个保守基序都具有植物C2H2型锌指蛋白特有的QALGGH序列,是一个典型的C2H2型锌指蛋白。同时,克隆的序列里都含有L-Box和DLN-Box等结构序列。将克隆的序列与已知功能的锌指蛋白基因进行聚类分析,结果表明,克隆的BcSTZ、 BSTZ、 QSTZ基因与已知的胁迫应答相关基因聚为一类。由此可以推测所克隆的基因序列与耐盐胁迫相关,但具体功能还需进一步验证。
DOI:10.7666/d.Y2360507URL [本文引用: 1]
芸薹属植物在我国种植面积大,分布范围广,其中包括白菜、甘蓝、芥菜等重要的蔬菜作物。蔬菜作物在整个生长过程中不可避免的要受到各种环境条件的影响,如高温、干旱、盐胁迫等。本文通过对大白菜、不结球白菜、甘蓝等供试材料的苗期进行耐盐性筛选、苗期生理指标的测定、目的基因的克隆及其序列分析。获得如下研究结果: 1芸薹属植物耐盐相关生理性状的分析 水培盐处理结果表明,盐胁迫下的大白菜、不结球白菜、甘蓝的生物量与对照相比都有不同程度的降低;叶片相对电导率,随着处理时间的延长有明显的下降趋势;也有个别品种随处理时问的延长,其相对电导率无明显变化;净光合速率受抑制程度不明显,与对照相比,个别品种表现出盐处理促进了光合作用;胞间CO2浓度除个别品种与对照变化趋势一致外,多数品种都有不同程度的降低;气孔导度,蒸腾速率均表现出不同程度的降低趋势。 2芸薹属植物耐盐相关基因的克隆 以大白菜、不结球白菜、甘蓝为试材,通过拟南芥耐盐基因(AtSTZ)全长序列和其近缘物种同源序列设计引物TNS1,利用同源克隆方法分离出耐盐相关基因BcSTZ、 BSTZ,、QSTZ各4条序列。对其序列进行比对分析,与已发现的大量植物中的C2H2型锌指蛋白保守基序具有很高的同源性。含有两个由21个氨基酸组成的C2H2保守基序,每个保守基序都具有植物C2H2型锌指蛋白特有的QALGGH序列,是一个典型的C2H2型锌指蛋白。同时,克隆的序列里都含有L-Box和DLN-Box等结构序列。将克隆的序列与已知功能的锌指蛋白基因进行聚类分析,结果表明,克隆的BcSTZ、 BSTZ、 QSTZ基因与已知的胁迫应答相关基因聚为一类。由此可以推测所克隆的基因序列与耐盐胁迫相关,但具体功能还需进一步验证。
.
[本文引用: 1]
.
DOI:10.1111/j.1467-7652.2010.00578.xURLPMID:21114612 [本文引用: 1]
We show here that transgenic Arabidopsis plants that expressed chimeric repressors derived from the AtMYB102, ANAC047, HRS1, ZAT6 and AtERF5 transcription factors were tolerant to treatment with 400 mm NaCl, which was lethal to wild-type plants. The transgenic plants grew well, without any apparent differences from the wild-type plants under normal growth condition. The transgenic lines expressing the AtMYB102, ANAC047 and HRS1 chimeric repressors germinated in the presence of 225 mm NaCl, while those expressing the ZAT6 and AtERF5 did not. However, the latter lines were tolerant to osmotic stress and germinated in the presence of 600 mm mannitol, suggesting a link between responses to salt and osmotic stress. Expression of the AtMYB102, ANAC047, ZAT6 and AtERF5 genes was induced by salt treatment, while that of HRS1 was repressed. HRS1 has transcriptional repressive activity and appears to suppress the expression of factors that negatively regulate salt tolerance. Microarray analysis revealed that the levels of expression of DREB1A, DREB2B and several genes for ZAT transcription factors rose 10- to 100-fold in the AtMYB102 chimeric repressor line under both normal and stress conditions. Elevated expression of DREB- and ZAT- related genes might be involved in the salt tolerance of the AtMYB102 chimeric repressor line. Transgenic rice plants expressing chimeric repressors derived from Os02g0325600 and Os03g0327800, rice homologues of HRS1 and ANAC047, were tolerant to salinity stress demonstrated by suppression of growth inhibition and ion leakages. Expression of a chimeric repressor provides an effective strategy for enhancing tolerance of plants to abiotic stress.
DOI:10.1093/jxb/eru313URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
DOI:10.1186/s12864-017-4082-yURL [本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
DOI:10.1016/1043-4666(92)90051-RURLPMID:1352237 [本文引用: 1]
Abstract We describe a locus, SUPERMAN, mutations in which result in extra stamens developing at the expense of the central carpels in the Arabidopsis thaliana flower. The development of superman flowers, from initial primordium to mature flower, is described by scanning electron microscopy. The development of doubly and triply mutant strains, constructed with superman alleles and previously identified homeotic mutations that cause alterations in floral organ identity, is also described. Essentially additive phenotypes are observed in superman agamous and superman apetala2 double mutants. The epistatic relationships observed between either apetala3 or pistillata and superman alleles suggest that the SUPERMAN gene product could be a regulator of these floral homeotic genes. To test this, the expression patterns of AGAMOUS and APETALA3 were examined in superman flowers. In wild-type flowers, APETALA3 expression is restricted to the second and third whorls where it is required for the specification of petals and stamens. In contrast, in superman flowers, APETALA3 expression expands to include most of the cells that would normally constitute the fourth whorl. This ectopic APETALA3 expression is proposed to be one of the causes of the development of the extra stamens in superman flowers. The spatial pattern of AGAMOUS expression remains unaltered in superman flowers as compared to wild-type flowers. Taken together these data indicate that one of the functions of the wild-type SUPERMAN gene product is to negatively regulate APETALA3 in the fourth whorl of the flower. In addition, superman mutants exhibit a loss of determinacy of the floral meristem, an effect that appears to be mediated by the APETALA3 and PISTILLATA gene products.
DOI:10.1046/j.1365-313X.1998.00043.xURL [本文引用: 1]
URL [本文引用: 1]
In order to elucidate categorization evolvement of C2H2 zinc finger protein, and clarify mechanism on C2H2 zinc finger protein in pollen development among plants at molecular level, the genes encoding C2H2 zinc finger protein analogues from 18 species of genera Brassica and Raphanus in Cuciferae were obtained by PCR strategy using specific primers designed from the full length of BcMF20 , a putative gene encoding C2H2 zinc finger protein which was related to the male sterility. The phylogenetic relationships of these species belonging to the family Cruciferae were investigated through comparison of the sequences. Homologous sequences of BcMF20 comparison indicated that the similarities among the genes at nucleotide and amino acid levels were 86.9% ~ 100% and 77.6%~100%, respectively. In the zinc finger regions of homologous sequences, the amino acid sequences were identical. These results showed that the BcMF20 was relative conservation in evolution in Cruciferae, and BcMF20 may play an important role in pollen development.
URL [本文引用: 1]
In order to elucidate categorization evolvement of C2H2 zinc finger protein, and clarify mechanism on C2H2 zinc finger protein in pollen development among plants at molecular level, the genes encoding C2H2 zinc finger protein analogues from 18 species of genera Brassica and Raphanus in Cuciferae were obtained by PCR strategy using specific primers designed from the full length of BcMF20 , a putative gene encoding C2H2 zinc finger protein which was related to the male sterility. The phylogenetic relationships of these species belonging to the family Cruciferae were investigated through comparison of the sequences. Homologous sequences of BcMF20 comparison indicated that the similarities among the genes at nucleotide and amino acid levels were 86.9% ~ 100% and 77.6%~100%, respectively. In the zinc finger regions of homologous sequences, the amino acid sequences were identical. These results showed that the BcMF20 was relative conservation in evolution in Cruciferae, and BcMF20 may play an important role in pollen development.
.
DOI:10.1038/378199a0URLPMID:7477325 [本文引用: 1]
THE Arabidopsis gene SUPERMAN ( SUP ) is necessary for the proper spatial development of reproductive floral tissues 1–3 . Recessive mutations cause extra stamens to form interior to the normal third whorl stamens, at the expense of fourth whorl carpel development 1–3 . The mutant phenotype is associated with the ectopic expression of the B function genes, AP3 and PI , in the altered floral region, closer to the centre of the flower than in the wild type 3 and ap3 sup and pi sup double mutants exhibit a phenotype similar to ap3 and pi single mutants. These findings led to SUP being interpreted as an upstream negative regulator of the B function organ-identity genes, acting in the fourth whorl 2,3 , to establish a boundary between stamen and carpel whorls. Here we show, using molecular cloning and analysis, that it is expressed in the third whorl and acts to maintain this boundary in developing flowers. The putative SUPERMAN protein contains one zinc-finger and a region resembling a basic leucine zipper motif, suggesting a function in transcriptional regulation.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
URL [本文引用: 1]
自交不亲和性是甘蓝在长期进化过程中形成的防止自交衰败、促进杂交优势的一种复杂而完善的遗传机制。克隆自交不亲和性相关基因对甘蓝自交不亲和性的深入研究和利用有重要意义。本研究通过挖掘0~60 min自花和异花授粉的甘蓝柱头转录组数据,筛选到一个受自花授粉诱导上调表达的基因,命名为BoSPI。BoSPI开放阅读框534bp,编码177个氨基酸,理论等电点为4.21,不包含信号肽和跨膜区,含有4个保守的EF-hand结构域。BoSPI基因起始密码子上游2000 bp的核苷酸序列中含有真菌诱导响应、代谢调节以及器官形成等应答元件。BoSPI基因在大肠杆菌中可诱导表达为17 k D的蛋白。BoSPI在柱头中表达量最高,在花瓣、萼片、叶片、雄蕊表达量较低。BoSPI蛋白被定位在细胞膜和细胞质。自花授粉30 min后对BoSPI基因的诱导表达显著增强。表明BoSPI参与了柱头响应自花花粉刺激的分子过程,可能是实现甘蓝自交不亲和性相关的某种新功能基因。
URL [本文引用: 1]
自交不亲和性是甘蓝在长期进化过程中形成的防止自交衰败、促进杂交优势的一种复杂而完善的遗传机制。克隆自交不亲和性相关基因对甘蓝自交不亲和性的深入研究和利用有重要意义。本研究通过挖掘0~60 min自花和异花授粉的甘蓝柱头转录组数据,筛选到一个受自花授粉诱导上调表达的基因,命名为BoSPI。BoSPI开放阅读框534bp,编码177个氨基酸,理论等电点为4.21,不包含信号肽和跨膜区,含有4个保守的EF-hand结构域。BoSPI基因起始密码子上游2000 bp的核苷酸序列中含有真菌诱导响应、代谢调节以及器官形成等应答元件。BoSPI基因在大肠杆菌中可诱导表达为17 k D的蛋白。BoSPI在柱头中表达量最高,在花瓣、萼片、叶片、雄蕊表达量较低。BoSPI蛋白被定位在细胞膜和细胞质。自花授粉30 min后对BoSPI基因的诱导表达显著增强。表明BoSPI参与了柱头响应自花花粉刺激的分子过程,可能是实现甘蓝自交不亲和性相关的某种新功能基因。
DOI:10.3321/j.issn:1000-3061.2002.04.003URL [本文引用: 1]
农杆菌渗入法 (Agroinfiltration)是近几年发展起来的一项快速、高效、重复性好的植物瞬间基因表达系统 ,已应用于外源基因表达分析、防卫反应、基因沉默、启动子分析及分离新的防御基因等领域。介绍了Agroinfiltration的原理、技术及其在植物分子生物学研究中的应用 ,并结合我们的经验介绍了对该项技术的改进。
DOI:10.3321/j.issn:1000-3061.2002.04.003URL [本文引用: 1]
农杆菌渗入法 (Agroinfiltration)是近几年发展起来的一项快速、高效、重复性好的植物瞬间基因表达系统 ,已应用于外源基因表达分析、防卫反应、基因沉默、启动子分析及分离新的防御基因等领域。介绍了Agroinfiltration的原理、技术及其在植物分子生物学研究中的应用 ,并结合我们的经验介绍了对该项技术的改进。
.
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2014.01174URL [本文引用: 1]
). was screened out from the contains open reading frame of 735 bp that encodes a new C2H2-type zinc finger protein with an isoelectric point (pI) of 9.2 and a molecular weight of 26.6 kD. The protein consists of two typical zinc-finger domains and contains two conserved QALGGH amino acid sequences. Subcellular localization showed that Bj26 is located in the nuclear. The histochemical and quantitative GUS assays, through transient gene expression in expression obviously increased under the induction of the fungal elicitor (Hexa-N-Acetyl-Chitohexaose). CDS alignments and phylogenetic analysis of
DOI:10.3724/SP.J.1006.2014.01174URL [本文引用: 1]
). was screened out from the contains open reading frame of 735 bp that encodes a new C2H2-type zinc finger protein with an isoelectric point (pI) of 9.2 and a molecular weight of 26.6 kD. The protein consists of two typical zinc-finger domains and contains two conserved QALGGH amino acid sequences. Subcellular localization showed that Bj26 is located in the nuclear. The histochemical and quantitative GUS assays, through transient gene expression in expression obviously increased under the induction of the fungal elicitor (Hexa-N-Acetyl-Chitohexaose). CDS alignments and phylogenetic analysis of
DOI:10.1016/j.plaphy.2016.04.033URLPMID:27156137 [本文引用: 1]
61TaZNF belongs to C2H2 zinc finger transcription factor.61TaZNFexpression was induced by salt, polyethylene glycol and abscisic acid.61TaZNFimproves salt tolerance of plants through Na+excretion.61TaZNFimproves salt tolerance of plants through narrowing of stomatal aperture.61TaZNFpromoter contained MeJA and salt response elements.
[本文引用: 1]
DOI:10.1016/j.postharvbio.2015.12.029URL [本文引用: 1]
.
[本文引用: 1]
.
[本文引用: 2]
DOI:10.1093/oxfordjournals.aob.a085414URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}