 ,, 严青青, 张巨松,*, 李春艳*, 窦海涛*新疆农业大学农学院 / 教育部棉花工程研究中心, 新疆乌鲁木齐 830052
,, 严青青, 张巨松,*, 李春艳*, 窦海涛*新疆农业大学农学院 / 教育部棉花工程研究中心, 新疆乌鲁木齐 830052Compensation Effect of Nitrogen Fertilizer on Photosynthetic Characteristics and Yield during Cotton Flowering Boll-setting Stage under Non-sufficient Drip Irrigation
SHI Hong-Liang,, YAN Qing-Qing, ZHANG Ju-Song,*, LI Chun-Yan*, DOU Hai-Tao*Agriculture College, Xinjiang Agricultural University / Research Center of Cotton Engineering, Urumqi 830052, Xinjiang, China通讯作者:
第一联系人:
收稿日期:2017-11-20接受日期:2018-06-9网络出版日期:2018-06-11
| 基金资助: |
Received:2017-11-20Accepted:2018-06-9Online:2018-06-11
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (298KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
石洪亮, 严青青, 张巨松, 李春艳, 窦海涛. 氮肥对非充分灌溉下棉花花铃期光合特性及产量的补偿作用[J]. 作物学报, 2018, 44(8): 1196-1204. doi:10.3724/SP.J.1006.2018.01196
SHI Hong-Liang, YAN Qing-Qing, ZHANG Ju-Song, LI Chun-Yan, DOU Hai-Tao.
非充分灌溉是针对水资源紧缺与用水效率低而提出的, 粮食和经济等作物生产中先后出现了非充分灌溉, 又称为有限灌溉或亏缺灌溉, 不追求单位面积上最高产量, 允许一定限度的减产。水分亏缺是作物生长环境中普遍存在的一种逆境胁迫, 也是影响干旱半干旱地区作物生产的主要因素, 水分和养分具有强烈的交互作用, 干旱半干旱地区植物营养的基本问题是在水分不能完全满足作物需求的条件下合理的氮肥运筹, 达到“以肥调水”的目的, 提高作物的灌溉水生产力, 增强抗旱性, 促进作物对有限水资源的充分利用[1,2,3,4]。因此, 研究非充分灌溉下增施氮肥对棉花产量形成的影响, 对干旱区棉花水肥高效管理技术的完善具有重要意义。水分胁迫的补偿效应是指作物受到阈值内的水分亏缺后, 在具有恢复因子(复水)和过程(时间)条件下, 表现出在生理生化和农艺指标上有利于作物生长、产量提高和品质改善的能力[5], 但补偿效应的产生不仅发生在干旱复水条件下, 干旱后增施氮肥同样可以达到一定的补偿效应, 补偿因水分不足对作物造成的影响[6]。褚丽丽等[7]研究发现大豆产量的补偿效应是水分亏缺和氮素营养合力作用的结果; 张立新等[8]研究表明适当增施氮肥可有效地改善水分亏缺下玉米叶片的光合特性, 从而增强作物的抗旱性; 彭世彰等[9]研究显示水稻抽穗开花期水分亏缺造成了产量显著下降, 分蘖后期水分亏缺处理在复水后表现出明显的补偿效应, 水稻产量较对照持平略有增加; 同时, 蔡一霞等[10]、吴自明等[11]研究认为水稻灌浆期水分亏缺时适量增施氮肥, 对功能叶光合同化物生产与积累无显著影响, 降低干旱胁迫危害、增强叶片光合功能、延长叶片功能期, 产量在补偿效应作用下无显著降低。但也有研究指出在土壤水分有限的条件下, 增施氮肥会使作物的水分胁迫加重, 对产量造成不利影响[12]。总体上来看, 前人在不同生育时期调亏灌溉[13,14]、干湿交替[15,16,17,18]、干旱胁迫与复水[9,19-22]、亏缺灌溉与增施氮肥[7-8,11,23-24]方面对作物生长发育[14,23,25]、光合性能[8,11,24,26-27]、产量及补偿效应[7,9,23,28]影响的研究较为全面。关于全生育期非充分灌溉下氮肥对棉花光合特性、产量及灌溉水生产力方面的研究还鲜见报道。本试验研究氮肥对非充分灌溉下棉花花铃期光合特性、产量及灌溉水生产力的影响, 明确氮肥对棉花产量形成的补偿效应机制, 以期为干旱半干旱地区水肥高效管理提供理论依据。
1 材料与方法
1.1 试验区概况
新疆农业科学院经济作物研究所实验基地, 位于新疆阿克苏市阿瓦提县丰收二场一连, 属暖温带大陆性干旱气候, 无霜期183~227 d, 年均日照2750~3029 h, 全年≥10℃积温3802.9℃, 多年平均降水量46.7 mm, 多年平均蒸发量1890.7 mm, 试验区耕作层(0~40 cm)土壤为沙壤土, 土壤平均容重1.44 g cm-3, 播种前土壤质量含水率(0~60 cm)平均为16.24%。连续2年土壤基础理化性质见表1。Table 1
表1
表1土壤基础理化性质
Table 1
| 年份 Year | 土层深度 Soil depth (cm) | 全氮 Total N (g kg-1) | 有机质 Organic matter (g kg-1) | 水解性氮 Hydrolytic N (mg kg-1) | 有效磷 Available P (mg kg-1) | 速效钾 Available K (mg kg-1) |
|---|---|---|---|---|---|---|
| 2015 | 0-20 | 0.50 | 6.90 | 105.00 | 32.40 | 131.00 |
| 20-40 | 0.40 | 5.60 | 70.80 | 21.60 | 168.00 | |
| 40-60 | 0.20 | 2.20 | 45.00 | 2.60 | 240.00 | |
| 2016 | 0-20 | 0.62 | 8.00 | 32.60 | 24.80 | 111.00 |
| 20-40 | 0.44 | 3.00 | 23.20 | 6.00 | 126.00 | |
| 40-60 | 0.34 | 2.00 | 13.80 | 1.80 | 152.00 |
新窗口打开|下载CSV
1.2 试验方案
采用裂区试验设计, 主区为总灌溉量, 分别为非充分灌溉2800 m3 hm-2 (当地一般棉田平均灌溉水平)和常规灌溉3800 m3 hm-2 (当地高产棉田平均灌溉水平); 副区为4个施氮(N)水平(0、150、300、450 kg hm-2), 分别用N0、N150、N300、N450表示。供试棉花品种为新陆中54号。采用机采棉种植模式, 行距配置(66+10) cm, 膜间行距60 cm, 株距11 cm, 理论株数为24.3万株 hm-2。小区面积44.9 m2, 重复3次, 重复间距50 cm, 占地面积为1159.2 m2。根据棉花生育期需水情况, 本试验共10次滴灌, 自现蕾期开始每7 d滴灌1次; 每次滴灌量由水表控制, 氮肥经电子秤称量后, 对应各处理放入施氮罐中, 随水滴施, 按照一水一肥进行。施用的肥料为尿素(N 46%)、颗粒状过磷酸钙(P2O5 12%)和农用颗粒钾肥(K2O 40%)。基施总量的20%、颗粒状过磷酸钙200 kg hm-2和农用颗粒钾肥100 kg hm-2; 追施尿素总量的80% (表2)。
Table 2
表2
表2水、氮分配表
Table 2
| 处理 Treatment | 日期 Date (day/month) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10/04- 12/04 | 16/06- 20/06 | 23/06- 27/06 | 30/06- 04/07 | 07/07- 11/07 | 14/07- 18/07 | 21/07- 25/07 | 28/07- 01/08 | 04/08- 08/08 | 11/08- 15/08 | 18/08- 22/08 | ||
| 灌溉量 Drip irrigation amount | ||||||||||||
| 2800 m3 hm-2 | 168.0 | 224.0 | 280.0 | 336.0 | 392.0 | 392.0 | 336.0 | 280.0 | 224.0 | 168.0 | ||
| 3800 m3 hm-2 | 228.0 | 304.0 | 380.0 | 456.0 | 532.0 | 532.0 | 456.0 | 380.0 | 304.0 | 228.0 | ||
| 施氮量 Nitrogen application | ||||||||||||
| N0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| N150 | 30 | 7.2 | 9.6 | 12.0 | 14.4 | 16.8 | 16.8 | 14.4 | 12.0 | 9.6 | 7.2 | |
| N300 | 60 | 14.4 | 19.2 | 24.0 | 28.8 | 33.6 | 33.6 | 28.8 | 24.0 | 19.2 | 14.4 | |
| N450 | 90 | 21.6 | 28.8 | 36.0 | 43.2 | 50.4 | 50.4 | 43.2 | 36.0 | 28.8 | 21.6 | |
新窗口打开|下载CSV
1.3 测定项目与方法
1.3.1 土壤基础理化性质 2015—2016年播种前按五点对角线方法取0~20 cm、20~40 cm、40~60 cm土层土样, 送新疆农业科学院检测中心测定土壤全氮、速效N、速效P、速效K和有机质含量。1.3.2 叶面积指数(LAI) 2016年于棉花初花期、盛花期、盛铃期、吐絮期采用LAI-2000冠层仪测定棉花叶面积指数(LAI)。2015年采用打孔法测定的LAI, 采用的方法不一致, 故只用2016年数据。
1.3.3 光合参数 2015—2016年于棉花初花期、盛花期、盛铃期、吐絮期在11:00—13:00时间内的晴朗天气利用英国Hansatech公司生产的TPS-2测定叶片净光合速率(Pn)和蒸腾速率(Tr)(滴灌施肥后的第5天测定)。
1.3.4 植株地上部光合物质积累 2015—2016年初花期至吐絮期, 选取具有代表性的6株棉花, 将营养器官与生殖器官分开, 放入电热恒温鼓风干燥箱105℃杀青30 min, 80℃烘至恒重, 测定其质量。
1.3.5 产量 2015—2016年在棉花吐絮期, 记数每小区株数和铃数, 选取有代表性的棉株, 从上至下取棉花样品100朵, 测其铃重, 重复3次。
1.3.6 计算公式 灌溉水生产力(kg m-3) = 籽棉产量/总灌溉量; 籽棉增产率(%) = (施氮区籽棉重量-不施氮区籽棉重量)/施氮量区籽棉重量×100%
1.4 统计分析
采用Microsoft Excel 2013、SPSS 19.0进行统计分析, 采用General Linear Model-Univariate Proce- Dure进行方差分析, 采用Duncan’s新复极差法进行多重比较。2 结果与分析
2.1 非充分灌溉下氮肥对棉花花铃期叶面积指数(LAI)的影响
由图1可知, 非充分灌溉与常规灌溉下不同生育时期各氮肥处理棉花LAI存在显著性差异, 前者较后者下降, 初花期平均下降18.1%, 盛花期平均下降9.9%, 盛铃期平均下降7.7%, 吐絮期平均下降13.5%。非充分灌溉与常规灌溉下随着氮肥量的增加LAI呈先增加后下降的趋势, 表现为N300>N450> N150>N0, 在盛花期达到峰值(除非充分灌溉下N0处理在初花期达到峰值外)。盛花期至盛铃期调控效应最明显, 非充分灌溉下N150、N300、N450处理棉花LAI较N0处理分别增加20.3%、33.2%和38.0%; 常规灌溉下N150、N300、N450处理棉花LAI较N0处理分别增加11.3%、23.6%和31.3%。2种灌溉下均表现N450处理棉花LAI指数提高效果最显著, 非充分灌溉下在不同生育时期N150处理棉花LAI补偿效应最显著, N450处理补偿效应最低。图1
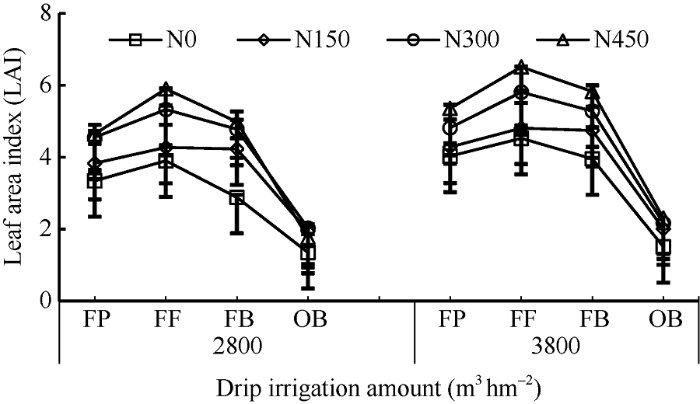 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1棉花花铃期LAI的比较
FP: 初花期; FF: 盛花期; FB: 盛铃期; OB: 吐絮期。
Fig. 1Comparison of LAI during cotton flowering and boll-setting stage
FP: flowering period; FF: full flower period; FB: full boll period; OB: opening boll period.
2.2 非充分灌溉下增施氮肥对棉花花铃期净光合速率(Pn)的影响
2015和2016两年数据趋势一致, 非充分灌溉与常规灌溉下不同生育时期各氮肥处理棉花Pn存在显著性差异(表3)。非充分灌溉较常规灌溉下各氮肥处理棉花Pn下降, 初花期、盛花期、盛铃期及吐絮期分别平均下降3.2%、2.7%、3.2%和10.9%, 可见盛铃期过后非充分灌溉下各氮肥处理棉花Pn下降快速。非充分灌溉与常规灌溉下, 不同生育时期随着施氮量的增加棉花Pn均表现为N300>N450> N150>N0。非充分灌溉下N150、N300、N450处理初花期棉花Pn较N0处理分别增加6.0%、20.4%和15.2%, 盛花期棉花Pn较N0处理分别增加9.6%、23.5%和18.6%, 盛铃期棉花Pn较N0处理分别增加9.7%、38.6%和32.5%; 常规灌溉下N150、N300、N450处理初花期棉花Pn较N0处理分别增加3.1%、18.3%和14.8%, 盛花期棉花Pn较N0处理分别增加6.8%、20.7%和16.0%, 盛铃期棉花Pn较N0处理分别增加9.1%、36.0%和29.5%。不同灌溉量下不同生育时期均表现为N300处理棉花Pn提高效果最显著, 非充分灌溉下N300处理棉花Pn补偿效应明显, 主要在盛花期, 较常规灌溉下N300处理提高12.9%。Table 3
表3
表3棉花花铃期功能叶(倒三叶) Pn的比较
Table 3
| 灌溉量 Drip irrigation amount | 氮肥处理 N treatment | 2015 | 2016 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 初花期 FP | 盛花期 FF | 盛铃期 FB | 吐絮期 OB | 初花期 FP | 盛花期 FF | 盛铃期 FB | 吐絮期 OB | ||
| 2800 m3 hm-2 | N0 | 32.5 e | 35.3 f | 28.8 d | 15.1 f | 31.9 f | 33.4 f | 25.1 d | 5.5 d |
| N150 | 34.7 d | 38.4 d | 34.8 c | 17.6 de | 33.8 de | 37.6 de | 25.7 d | 6.4 cd | |
| N300 | 40.5 ab | 45.9 b | 44.8 a | 22.7 a | 40.5 ab | 43.9 ab | 42.9 a | 9.5 a | |
| N450 | 38.2 c | 43.1 c | 41.8 b | 19.9 bc | 37.8 c | 41.3 c | 38.0 b | 7.3 bc | |
| 3800 m3 hm-2 | N0 | 34.1 d | 36.6 e | 30.1 d | 17.1 e | 33.5 e | 35.8 e | 27.1 cd | 7.3 bc |
| N150 | 35.0 d | 38.9 d | 35.1 c | 18.8 cd | 34.8 d | 38.8 d | 28.1 c | 7.7 bc | |
| N300 | 41.6 a | 46.8 a | 45.8 a | 23.4 a | 41.2 a | 44.6 a | 43.6 a | 10.3 a | |
| N450 | 39.8 b | 43.9 c | 42.6 b | 20.8 b | 39.7 b | 42.3 bc | 38.6 b | 8.8 ab | |
新窗口打开|下载CSV
2.3 非充分灌溉下增施氮肥对棉花花铃期蒸腾速率(Tr)的影响
非充分灌溉与常规灌溉下不同生育时期各氮肥处理的棉花Tr存在一定的显著性差异(表4)。前者较后者下降, 初花期、盛花期、盛铃期及吐絮期分别平均下降7.6%、6.3%、7.9%和11.6%, 说明随着生育进程的推进非充分灌溉下各氮肥处理不同程度限制了气孔开放, 导致光合性能降低。非充分灌溉与常规灌溉下, 不同生育时期均随施氮量的增加棉花Tr表现为N300>N450>N150>N0, 在盛花期达到峰值。不同灌溉下均表现为N300处理在盛花期提高效果显著, 非充分灌溉下N300处理盛花期棉花Tr较N0处理增加24.6%, 常规灌溉下N300处理盛花期棉花Tr较N0处理增加18.6%, 非充分灌溉下N300处理棉花Tr补偿效应明显, 主要在盛花期, 较常规灌溉下N300处理提高22.5%, 与Pn表现结果一致, 且两年数据趋势一致。Table 4
表4
表4棉花花铃期功能叶(倒三叶)Tr的比较
Table 4
| 灌溉量 Drip irrigation amount | 氮肥处理 N treatment | 2015 | 2016 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 初花期 FP | 盛花期 FF | 盛铃期 FB | 吐絮期 OB | 初花期 FP | 盛花期 FF | 盛铃期 FB | 吐絮期 OB | ||
| 2800 m3 hm-2 | N0 | 5.2 d | 5.7 d | 5.5 d | 3.1 d | 4.6 d | 5.1 c | 4.4 e | 1.8 d |
| N150 | 5.5 cd | 6.1 d | 5.9 cd | 3.4 cd | 5.4 c | 5.9 bc | 5.2 d | 2.2 de | |
| N300 | 6.2 ab | 7.3 ab | 7.1 ab | 4.8 ab | 6.2 ab | 7.0 a | 6.9 ab | 3.2 ab | |
| N450 | 5.9 abc | 6.8 bc | 6.8 b | 4.5 b | 5.6 bc | 6.5 ab | 6.1 c | 2.6 cd | |
| 3800 m3 hm-2 | N0 | 5.6 bcd | 6.2 d | 6.0 c | 3.5 cd | 5.8 abc | 5.9 bc | 5.9 cd | 2.5 cd |
| N150 | 5.9 abc | 6.3 cd | 6.2 c | 3.7 c | 6.1 ab | 6.3 ab | 6.1 c | 2.9 cd | |
| N300 | 6.4 a | 7.7 a | 7.2 a | 5.0 a | 6.4 a | 7.2 a | 7.1 a | 3.3 a | |
| N450 | 6.2 ab | 7.3 ab | 7.0 ab | 5.0 a | 5.9 abc | 6.9 a | 6.4 bc | 3.1 ab | |
新窗口打开|下载CSV
2.4 非充分灌溉下增施氮肥对单株棉花花铃期光合物质积累与分配的影响
从表5可以看出, 非充分灌溉较常规灌溉下单株棉花光合产物积累量和向生殖器官分配比例分别平均下降7.6%和3.2%; 单株光合产物积累向营养器官分配比例平均增加2.0%。非充分灌溉与常规灌溉下随着施氮量的增加单株棉花光合产物积累量表现为N450>N300>N150>N0; 单株光合产物积累向营养器官分配比例表现为N0>N150>N450>N300; 单株光合产物积累向生殖器官分配比例表现为N300>N450>N150>N0。非充分灌溉下N150、N300、N450处理单株棉花光合产物积累量较N0处理分别增加12.0%、41.4%和41.3%, 单株光合产物积累向生殖器官分配较N0处理分别提高10.3%、26.0%和20.2%; 常规灌溉下N150、N300、N450处理单株棉花光合产物积累量较N0处理分别增加9.9%、35.8% 和36.6%, 单株光合产物积累向生殖器官分配较N0处理分别提高8.4%、23.3%和16.6%。非充分灌溉下N300处理单株棉花光合产物积累向生殖器官分配补偿效应明显, 较常规灌溉下N300处理提高10.7%。Table 5
表5
表5单株棉花吐絮期地上部光合产物积累与分配的比较
Table 5
| 灌溉量 Drip irrigation amount | 氮肥处理 N treatment | 2015 | 2016 | ||||
|---|---|---|---|---|---|---|---|
| 光合物质积累 PA (g) | 营养器官分配 VOA (%) | 生殖器官分配 ROA (%) | 光合物质积累 PA (g) | 营养器官分配 VOA (%) | 生殖器官分配 ROA (%) | ||
| 2800 m3 hm-2 | N0 | 52.3 h | 72.1 a | 27.9 h | 42.2 h | 63.9 a | 36.1 h |
| N150 | 58.0 f | 68.2 c | 31.8 f | 49.1 g | 60.6 c | 39.4 f | |
| N300 | 84.7 d | 60.9 g | 39.1 b | 76.0 c | 52.7 g | 47.3 b | |
| N450 | 85.9 c | 64.6 e | 35.4 d | 74.7 d | 55.2 e | 44.8 d | |
| 3800 m3 hm-2 | N0 | 57.4 g | 70.2 b | 29.8 g | 50.4 f | 61.7 b | 38.3 g |
| N150 | 63.0 e | 66.8 d | 33.2 e | 56.6 e | 59.1 d | 40.9 e | |
| N300 | 87.9 b | 59.6 h | 40.4 a | 79.9 a | 51.9 h | 48.1 a | |
| N450 | 91.8 a | 64.3 f | 35.7 c | 78.4 b | 54.6 f | 45.4 c | |
新窗口打开|下载CSV
2.5 非充分灌溉下增施氮肥对棉花产量构成因素及灌溉水生产力的影响
方差分析(表6)显示, 年份对单株结铃数和单铃重的影响均达到极显著水平(P<0.01); 灌溉对灌溉水生产力的影响达到极显著水平(P<0.01); 氮肥对单株结铃数、单铃重、籽棉产量及灌溉水生产力的影响均达到极显著水平(P<0.01); 年份与氮肥对单株结铃数、单铃重、籽棉产量及灌溉水生产力的影响均达到极显著水平(P<0.01)。Table 6
表6
表6棉花产量构成因素及灌溉水生产力的方差分析
Table 6
| 变异来源 Source of variation | 单株结铃数BN | 单铃重BW | 籽棉产量SCY | 灌溉水生产力IWP | ||||
|---|---|---|---|---|---|---|---|---|
| MS | F | MS | F | MS | F | MS | F | |
| 年份 Year (Y) | 18.4 | 460.2** | 2.0 | 102.7** | 160534.9 | 2.8 | 0 | 2.6 |
| 灌溉 Drip irrigation (W) | 0.2 | 4.1 | 0 | 0.8 | 146835.9 | 2.6 | 2.5 | 408.4** |
| 氮肥 N fertilizer (N) | 24.2 | 201.5** | 6.2 | 106.7** | 32525954.1 | 190.2** | 3.2 | 176.6** |
| Y×W | 0 | 0 | 0 | 0.1 | 7971.4 | 0.1 | 0.0 | 0.0 |
| Y×N | 1.2 | 10.3** | 0.4 | 5.9** | 1984708.9 | 11.6** | 0.2 | 11.0** |
| W×N | 0.1 | 0.3 | 0 | 0.6 | 37744.6 | 0.2 | 0.1 | 5.8 |
| Y×W×N | 0 | 0.2 | 0 | 0.3 | 8937.6 | 0.1 | 0 | 0.4 |
| 误差 Error | 1.3 | 0.6 | 1824487.0 | 0.2 | ||||
新窗口打开|下载CSV
由表7可知, 非充分灌溉较常规灌溉下单株结铃数、单铃重及籽棉产量分别平均下降2.2%、0.6%和2.1%; 灌溉水生产力和籽棉增产率分别平均增加24.8%和4.9%。非充分灌溉与常规灌溉下随着施氮量的增加单株结铃数、单铃重、籽棉产量及灌溉水生产力均表现为N300>N450>N150>N0; 籽棉增产率表现为N300>N450>N150。非充分灌溉下N150、N300、N450处理单株结铃数较N0处理分别增加11.3%、28.7%和26.6%, 单铃重较N0处理分别提高11.1%、16.0%和14.8%, 灌溉水生产力较N0处理分别提高14.2%、34.3%和35.5%; 常规灌溉下N150、N300、N450处理单株结铃数较N0处理分别增加8.9%、27.3%和24.3%, 单铃重较N0处理分别提高8.5%、14.7%和13.1%, 灌溉水生产力较N0处理分别提高11.6%、31.8%和27.9%。非充分灌溉下N300处理补偿效应明显, 表现在单株结铃数、单铃重、灌溉水生产力及籽棉增产率较常规灌溉下N300处理分别提高5.0%、8.0%、7.5%和7.1%。两年数据趋势一致, 2016年数据表现更为显著。
Table 7
表7
表7棉花产量构成因素及灌溉水生产力的比较
Table 7
| 年份 Year | 灌溉量 Drip irrigation amount | 氮肥处理 N treatment | 单株结铃数 BN | 单铃重 BW (g) | 籽棉产量 SCY (kg hm-2) | 灌溉水生产力 IWP (kg m-3) | 籽棉增产率 SCYR (%) |
|---|---|---|---|---|---|---|---|
| 2015 | 2800 m3 hm-2 | N0 | 3.8 c | 5.2 b | 4147.4 c | 1.5 c | |
| N150 | 4.4 b | 6.1 a | 4796.1 b | 1.7 b | 13.4 c | ||
| N300 | 5.3 a | 6.2 a | 5797.5 a | 2.1 a | 28.2 a | ||
| N450 | 5.1 a | 6.2 a | 5543.3 a | 2.0 a | 18.8 abc | ||
| 3800 m3 hm-2 | N0 | 4.0 c | 5.3 b | 4321.1 c | 1.1 d | ||
| N150 | 4.5 b | 6.0 a | 4847.5 b | 1.3 d | 10.7 c | ||
| N300 | 5.4 a | 6.2 a | 5836.9 a | 1.5 c | 25.9 a | ||
| N450 | 5.1 a | 6.2 a | 5618.3 a | 1.5 c | 22.8 ab | ||
| 2016 | 2800 m3 hm-2 | N0 | 4.8 d | 5.0 d | 3808.4 d | 1.4 c | |
| N150 | 5.3 bc | 5.3 c | 4479.2 c | 1.6 b | 14.8 c | ||
| N300 | 6.7 a | 5.9 ab | 6337.9 ab | 2.3 s | 39.8 a | ||
| N450 | 6.7 a | 5.7 b | 6018.3 b | 2.2 a | 36.6 ab | ||
| 3800 m3 hm-2 | N0 | 5.0 cd | 5.0 d | 4041.9 d | 1.1 e | ||
| N150 | 5.3 b | 5.4 c | 4629.7 c | 1.2 d | 12.6 c | ||
| N300 | 7.0 a | 5.9 a | 6464.3 a | 1.7 b | 37.4 ab | ||
| N450 | 6.7 a | 5.7 b | 6053.6 b | 1.6 b | 33.2 b |
新窗口打开|下载CSV
3 讨论
研究表明, 拔节孕穗后期和抽穗开花期水分亏缺处理水稻光合产物积累量显著低于对照[9]。也有研究认为, 在全生育期水分亏缺处理条件下, 春青稞耗穗粒数、千粒质量和籽粒产量均小于充分灌溉处理[29]。本试验非充分灌溉下各氮肥处理在棉花花铃期叶面积指数(LAI)、蒸腾速率、净光合速率、光合产物积累、单株结铃数、单铃重及籽棉产量均低于常规灌溉处理, 但并无显著性降低。这表明非充分灌溉下增施氮肥可不同程度地缩小与常规灌溉的差异, 说明通过增施氮肥同样可以达到一定的补偿效应。研究发现, 返青期干旱胁迫后复水, 各施氮处理油菜的LAI、地上部净光合速率、光合产物积累、产量及产量构成均表现出一定程度的补偿效应, 补偿效果随施氮量的增加先增加后降低[23]。虽然本试验条件与干旱复水条件不一致, 但最终结果表现与其一致, 本试验结果表明, 非充分灌溉与常规灌溉下棉花净光合速率、光合产物积累、单铃重、单株结铃数、产量及灌溉水生产力补偿效果均随施氮量增加表现为先增加后下降, 均表现为N300处理补偿效应最为显著, 而LAI却随着施氮量增加而下降。说明无论在非充分灌溉还是常规灌溉下, 施用过多氮肥对棉花的影响均表现为负效应, 造成植株疯长, 导致贪青晚熟。研究认为, 施氮能不同程度降低水分亏缺下玉米[8]、烟草[30]、小麦[24]等作物叶片的蒸腾速率, 在一定程度上可延缓叶片衰老, 提高净光合速率, 从而减缓水分亏缺对光合作用的伤害[31]。本试验结果与前人研究结果趋于一致, 非充分灌溉下各氮肥处理通过降低蒸腾速率, 来减弱棉花叶片水分蒸腾, 从而提高了叶片的水分利用效率, 延长了叶片光合作用时间, 进而提高了棉花叶片的净光合速率, 通过生理代谢调节, 在一定程度上提高了叶片的光合性能。说明棉花通过自身机制的调节, 保证了光合作用对光能的吸收与利用, 同时也反映出增施氮肥对非充分灌溉下棉花光合作用具有较好补偿效果。干旱胁迫主要降低作物的光合产物积累量[22,28], 并减弱干物质由源到汇的转运能力[26], 降低籽粒灌浆速率[32], 最终影响其产量[33]。但也有研究表明, 适度水分亏缺会提高旱地小麦灌浆前期穗部的碳同化能力, 加速灌浆前期穗部光合产物向籽粒的转运, 以维持一定的产量水平[28]。本试验结果与前人研究结果不尽一致, 非充分灌溉虽然降低了光合产物积累量, 但向生殖器官分配及转运能力有所提高, 同时也表现出了不同程度的早衰, 尤其N0处理表现更为显著。通过增施氮肥可有效缓解这种情况, 同时光合产物向生殖器官分配及转运能力有较大的提高, 尤其N300处理表现最为显著。本试验补偿效应机制一方面是通过光合作用补偿, 另一方面主要是通过氮肥运筹的调节作用, 加强了光合产物积累和向生殖器官转运能力, 进而保证棉花产量无显著下降。另外针对无霜期较短的区域, 本试验条件下可能有效提高霜前花率。郝树荣等[34]发现高水高肥并不一定高产, 轻旱与低氮具有明显的协同互作效应, 在保产的同时达到节水的目的。本实验条件下, 非充分灌溉较常规灌溉节水26.3%, 籽棉产量及其构成因素无显著下降, 增产率高。
4 结论
非充分灌溉下N300处理籽棉产量下降了1.3%, 但产量补偿效果(籽棉增产率)与灌溉水生产力分别提高了7.1%和7.5%; 主要是光合作用的补偿能力提高, 进而增强单株光合产物积累和向生殖器官转运的补偿效果, 与常规灌溉相比, 棉花花铃期净光合速率平均提高10.9%, 棉花光合产物提高10.7%; 在南疆自然生态条件下, 以非充分灌溉下施氮300 kg hm-2时棉花花铃期LAI、净光合速率及光合物质积累量适宜, 向生殖器官转运补偿效果显著, 具有最大的产量补偿效应, 且节水26.3%。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3969/j.issn.1002-6819.2014.02.015URLMagsci [本文引用: 1]

地下滴灌技术是最为节水的灌溉技术,为了推进地下滴灌技术在紫花苜蓿生产中的应用,2012年在甘肃武威开展了大田试验,通过对建植当年第2茬苜蓿不同生育期设置不同的亏水处理,研究了地下滴灌条件下调亏灌溉对紫花苜蓿耗水、产量和品质的影响。结果表明:随亏水程度的加重,紫花苜蓿的产量降低,耗水量减少,水分利用效率(WUE)提高,干重茎叶比降低,粗蛋白含量提高,粗蛋白产量降低;全生育期调亏,生育期间的耗水强度呈现现蕾期>分枝期>分枝前期的规律,并且随着亏水程度的增加,规律愈加明显;分枝期调亏的紫花苜蓿,在现蕾期复水后耗水强度有明显的补偿效应;与现蕾期调亏相比,分枝前期和分枝期调亏有利于降低茎叶比,提高粗蛋白质量分数和粗蛋白产量。该研究揭示了地下滴灌条件下不同调亏处理对紫花苜蓿耗水、产量和品质的影响,为干旱地区优质、高产、节水苜蓿的生产提供了试验依据。
DOI:10.3969/j.issn.1002-6819.2014.02.015URLMagsci [本文引用: 1]
地下滴灌技术是最为节水的灌溉技术,为了推进地下滴灌技术在紫花苜蓿生产中的应用,2012年在甘肃武威开展了大田试验,通过对建植当年第2茬苜蓿不同生育期设置不同的亏水处理,研究了地下滴灌条件下调亏灌溉对紫花苜蓿耗水、产量和品质的影响。结果表明:随亏水程度的加重,紫花苜蓿的产量降低,耗水量减少,水分利用效率(WUE)提高,干重茎叶比降低,粗蛋白含量提高,粗蛋白产量降低;全生育期调亏,生育期间的耗水强度呈现现蕾期>分枝期>分枝前期的规律,并且随着亏水程度的增加,规律愈加明显;分枝期调亏的紫花苜蓿,在现蕾期复水后耗水强度有明显的补偿效应;与现蕾期调亏相比,分枝前期和分枝期调亏有利于降低茎叶比,提高粗蛋白质量分数和粗蛋白产量。该研究揭示了地下滴灌条件下不同调亏处理对紫花苜蓿耗水、产量和品质的影响,为干旱地区优质、高产、节水苜蓿的生产提供了试验依据。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1000-1980.2005.06.007URL [本文引用: 1]
根据国内外水分胁迫补偿效应的研究成果和笔者近年来的试验资料,对水分胁迫补偿效应的研究进展进行了分析和总结.分析结果表明,水分胁迫期间或旱后复水,作物在生长发育、干物质累积以及根系活力、叶绿素浓度、叶片光合作用等方面均可出现补偿效应;补偿程度与水分胁迫程度、胁迫时期和胁迫持续时间有关;营养生长阶段或生殖生长阶段的初期,水分胁迫往往具有较好的补偿效应.论述了光合作用、补偿生长之间的关系,对补偿效应产生的机制进行了初步探讨.提出了在补偿效应的产生条件、变化规律、生理生态机制以及在田间尺度上的效应等方面需要进一步研究的问题.
DOI:10.3321/j.issn:1000-1980.2005.06.007URL [本文引用: 1]
根据国内外水分胁迫补偿效应的研究成果和笔者近年来的试验资料,对水分胁迫补偿效应的研究进展进行了分析和总结.分析结果表明,水分胁迫期间或旱后复水,作物在生长发育、干物质累积以及根系活力、叶绿素浓度、叶片光合作用等方面均可出现补偿效应;补偿程度与水分胁迫程度、胁迫时期和胁迫持续时间有关;营养生长阶段或生殖生长阶段的初期,水分胁迫往往具有较好的补偿效应.论述了光合作用、补偿生长之间的关系,对补偿效应产生的机制进行了初步探讨.提出了在补偿效应的产生条件、变化规律、生理生态机制以及在田间尺度上的效应等方面需要进一步研究的问题.
URL [本文引用: 1]
总结了作物在水分亏缺下产生补偿效应的类型、机制及条件 .补偿效应类型可分为生长补偿、生理生化补偿、代谢及产量补偿等 ;从渗透调节、脱水保护等方面对补偿效应的生化及分子机制作了探讨 ,并阐明了作物产生补偿效应的生物学基础 ,指出了进一步研究的方向
URL [本文引用: 1]
总结了作物在水分亏缺下产生补偿效应的类型、机制及条件 .补偿效应类型可分为生长补偿、生理生化补偿、代谢及产量补偿等 ;从渗透调节、脱水保护等方面对补偿效应的生化及分子机制作了探讨 ,并阐明了作物产生补偿效应的生物学基础 ,指出了进一步研究的方向
[本文引用: 3]
[本文引用: 3]
DOI:10.11674/zwyf.2009.0112URL [本文引用: 4]
采用2种不同夏玉米基因型(陕单9号,抗旱品种;陕单911,不抗旱品种)的盆栽试验,研究了长期水分胁迫下氮、钾对各生育期叶片净光合速率、蒸腾速率、胞间二氧化碳浓度和叶绿素含量的影响,旨在从光合生理特性揭示这些因子的抗旱机理。结果表明,长期水分胁迫下叶片净光合速率、蒸腾速率、胞间二氧化碳浓度(除成熟期)和叶绿素含量显著降低,不抗旱品种降幅更甚。抗旱品种的净光合速率和叶绿素含量大于不抗旱品种,而蒸腾速率和胞间二氧化碳浓度则相反。两品种苗期光合作用较弱,净光合速率和叶绿素含量均较低,抽雄期达到高峰。施氮能不同程度降低水分胁迫下玉米叶片的蒸腾速率,增加叶绿素含量,提高净光合速率,从而减缓水分胁迫对光合作用的伤害。随氮肥用量增加,不抗旱品种净光合速率和叶绿素含量显著升高,蒸腾速率和胞间二氧化碳浓度明显降低,两种氮肥用量间有显著差异;抗旱品种存低氮用最时效果显著,但高低氮用量间无显著区别。钾对受水分胁迫的玉米表现出比氮肥更突出的效果。相反,存适量供水条件下,氮、钾肥的作用明显下降。以上结果表明,适当用量的氮、钾肥可以有效地改善水分胁迫下作物叶片的光合特性,从而增强作物的抗旱性。
DOI:10.11674/zwyf.2009.0112URL [本文引用: 4]
采用2种不同夏玉米基因型(陕单9号,抗旱品种;陕单911,不抗旱品种)的盆栽试验,研究了长期水分胁迫下氮、钾对各生育期叶片净光合速率、蒸腾速率、胞间二氧化碳浓度和叶绿素含量的影响,旨在从光合生理特性揭示这些因子的抗旱机理。结果表明,长期水分胁迫下叶片净光合速率、蒸腾速率、胞间二氧化碳浓度(除成熟期)和叶绿素含量显著降低,不抗旱品种降幅更甚。抗旱品种的净光合速率和叶绿素含量大于不抗旱品种,而蒸腾速率和胞间二氧化碳浓度则相反。两品种苗期光合作用较弱,净光合速率和叶绿素含量均较低,抽雄期达到高峰。施氮能不同程度降低水分胁迫下玉米叶片的蒸腾速率,增加叶绿素含量,提高净光合速率,从而减缓水分胁迫对光合作用的伤害。随氮肥用量增加,不抗旱品种净光合速率和叶绿素含量显著升高,蒸腾速率和胞间二氧化碳浓度明显降低,两种氮肥用量间有显著差异;抗旱品种存低氮用最时效果显著,但高低氮用量间无显著区别。钾对受水分胁迫的玉米表现出比氮肥更突出的效果。相反,存适量供水条件下,氮、钾肥的作用明显下降。以上结果表明,适当用量的氮、钾肥可以有效地改善水分胁迫下作物叶片的光合特性,从而增强作物的抗旱性。
[本文引用: 4]
[本文引用: 4]
DOI:10.3864/j.issn.0578-1752.2015.08.04URL [本文引用: 1]
【目的】探明抗旱性不同的单片段代换系(single segment substitution line,SSSL)水稻与其受体亲本在灌浆期亏缺灌溉条件下茎鞘非结构性碳水化合物的降解、运转及籽粒充实过程中相关酶活性的变化。【方法】以介入巴西 陆稻IAPAR9抗性基因片段的单片段代换系W27-3-5-2-4-3-3-5-7和受体亲本华粳籼74为材料,利用水平分根系统在水稻花后7 d设置全根水层(两个分隔室内均保持浅水层,W/W)、半根水分亏缺(一个分隔室内保持浅水层,另一个分隔室内土壤水分控制在-30—-40 kPa,W/D)、全根水分亏缺(两个分隔室内土壤水分均控制在-30—-40 kPa,D/D)3种土壤水分处理,研究花后7 d不同土壤水分亏缺对抗旱性不同的单片段代换系及其受体亲本的产量和构成的影响。【结果】与W/W相比,D/D处理下茎鞘和籽粒内源ABA含量显著增加, 灌浆中后期水稻剑叶的相对含水量、SPAD值及光合速率明显降低,水稻茎鞘中α-淀粉酶、β-淀粉酶活性有所增加,但是其蔗糖合酶(sucrose synthase,SS)的活性却显著降低。D/D处理下籽粒SS、酸性蔗糖转化酶(acid invertase,AINV)、可溶性淀粉合酶(soluble starch synthase,SSS)和Q酶活性在灌浆前中期也明显降低。然而,W/D处理下茎鞘和籽粒内源ABA有所增加,但与W/W差异不显著。W/D处理对水 稻剑叶的相对含水量、SPAD值、光合速率无显著影响,但对灌浆前中期籽粒SS、AINV、SSS和Q酶活性却有显著影响,使得灌浆前中期籽粒SS、 AINV、SSS和Q酶活性显著上升。分析灌浆期亏缺灌溉下单片段代换系与受体亲本间差异,发现亏缺灌溉下代换系的茎鞘非结构性碳水化合物的运转率明显地 高于华粳籼74,其籽粒灌浆充实过程中相关酶的活性、物质的动态变化及内源ABA含量等对水分亏缺响应较华粳籼74敏感,其产量在两种亏缺灌溉处理下降低 不明显,表现出较强的抗旱性。【结论】灌浆期适度土壤水分亏缺可提高稻株各器官ABA含量,利于茎鞘非结构性碳水化合物的降解和输出,但对灌浆期功能叶的 光合同化物的生产与积累无显著影响,因此产量在适度水分亏缺条件下不会显著降低。
DOI:10.3864/j.issn.0578-1752.2015.08.04URL [本文引用: 1]
【目的】探明抗旱性不同的单片段代换系(single segment substitution line,SSSL)水稻与其受体亲本在灌浆期亏缺灌溉条件下茎鞘非结构性碳水化合物的降解、运转及籽粒充实过程中相关酶活性的变化。【方法】以介入巴西 陆稻IAPAR9抗性基因片段的单片段代换系W27-3-5-2-4-3-3-5-7和受体亲本华粳籼74为材料,利用水平分根系统在水稻花后7 d设置全根水层(两个分隔室内均保持浅水层,W/W)、半根水分亏缺(一个分隔室内保持浅水层,另一个分隔室内土壤水分控制在-30—-40 kPa,W/D)、全根水分亏缺(两个分隔室内土壤水分均控制在-30—-40 kPa,D/D)3种土壤水分处理,研究花后7 d不同土壤水分亏缺对抗旱性不同的单片段代换系及其受体亲本的产量和构成的影响。【结果】与W/W相比,D/D处理下茎鞘和籽粒内源ABA含量显著增加, 灌浆中后期水稻剑叶的相对含水量、SPAD值及光合速率明显降低,水稻茎鞘中α-淀粉酶、β-淀粉酶活性有所增加,但是其蔗糖合酶(sucrose synthase,SS)的活性却显著降低。D/D处理下籽粒SS、酸性蔗糖转化酶(acid invertase,AINV)、可溶性淀粉合酶(soluble starch synthase,SSS)和Q酶活性在灌浆前中期也明显降低。然而,W/D处理下茎鞘和籽粒内源ABA有所增加,但与W/W差异不显著。W/D处理对水 稻剑叶的相对含水量、SPAD值、光合速率无显著影响,但对灌浆前中期籽粒SS、AINV、SSS和Q酶活性却有显著影响,使得灌浆前中期籽粒SS、 AINV、SSS和Q酶活性显著上升。分析灌浆期亏缺灌溉下单片段代换系与受体亲本间差异,发现亏缺灌溉下代换系的茎鞘非结构性碳水化合物的运转率明显地 高于华粳籼74,其籽粒灌浆充实过程中相关酶的活性、物质的动态变化及内源ABA含量等对水分亏缺响应较华粳籼74敏感,其产量在两种亏缺灌溉处理下降低 不明显,表现出较强的抗旱性。【结论】灌浆期适度土壤水分亏缺可提高稻株各器官ABA含量,利于茎鞘非结构性碳水化合物的降解和输出,但对灌浆期功能叶的 光合同化物的生产与积累无显著影响,因此产量在适度水分亏缺条件下不会显著降低。
[本文引用: 3]
[本文引用: 3]
DOI:10.3724/SP.J.1006.2008.01005URL [本文引用: 1]
以粳型陆稻中旱3号和粳型水稻扬粳9538为材料,设置裸地旱种和水种两种方式及低氮(LN,100kg hm^-2)、中氮(NN,200kg hm^-2)和高氮(HN,300kg hm^-2)3种N素水平,比较研究了氮素营养对陆稻和水稻产量形成的影响。结果表明,旱种HN处理下陆稻和水稻的产量以及水种HN处理下水稻的产量较NN有所下降;但水种HN处理下陆稻的产量较NN增加;随施N量增加,水、陆稻在两种种植方式下穗数均增加,每穗粒数表现不一,结实率均下降,但陆稻降幅小于水稻,陆稻千粒重差异不显著,而水稻则显著下降。与水种相比,旱种条件下陆稻的千粒重无显著变化,而水稻千粒重则显著下降,水、陆稻旱种的结实率均有所提高,但陆稻的提高幅度大于水稻。与水稻相比,陆稻不定根数少,吸N能力低,分蘖能力弱,成穗数少、穗型小,产量较低。拔节至抽穗期不定根数的增幅大,叶片含N率下降慢,花后叶片含N率和剑叶叶绿素(SPAD)值下降快。陆稻光合生产力对水分胁迫的负响应小,对增施N素正响应大。表明陆稻和水稻对种植方式和N素的响应有明显差异。对陆稻和水稻的产量增产途径进行了讨论。
DOI:10.3724/SP.J.1006.2008.01005URL [本文引用: 1]
以粳型陆稻中旱3号和粳型水稻扬粳9538为材料,设置裸地旱种和水种两种方式及低氮(LN,100kg hm^-2)、中氮(NN,200kg hm^-2)和高氮(HN,300kg hm^-2)3种N素水平,比较研究了氮素营养对陆稻和水稻产量形成的影响。结果表明,旱种HN处理下陆稻和水稻的产量以及水种HN处理下水稻的产量较NN有所下降;但水种HN处理下陆稻的产量较NN增加;随施N量增加,水、陆稻在两种种植方式下穗数均增加,每穗粒数表现不一,结实率均下降,但陆稻降幅小于水稻,陆稻千粒重差异不显著,而水稻则显著下降。与水种相比,旱种条件下陆稻的千粒重无显著变化,而水稻千粒重则显著下降,水、陆稻旱种的结实率均有所提高,但陆稻的提高幅度大于水稻。与水稻相比,陆稻不定根数少,吸N能力低,分蘖能力弱,成穗数少、穗型小,产量较低。拔节至抽穗期不定根数的增幅大,叶片含N率下降慢,花后叶片含N率和剑叶叶绿素(SPAD)值下降快。陆稻光合生产力对水分胁迫的负响应小,对增施N素正响应大。表明陆稻和水稻对种植方式和N素的响应有明显差异。对陆稻和水稻的产量增产途径进行了讨论。
[本文引用: 1]
[本文引用: 1]
DOI:10.6041/j.issn.1000-1298.2016.04.014URL [本文引用: 2]
以棉花(Gossypium hirsutum L.)为试验材料,采用防雨棚下桶栽土培方法,进行调亏灌溉(Regulated deficitirrigation,RDI)对棉花根、冠生长的影响研究.试验采用二因素随机区组设计,设置4个水分调亏阶段:苗期(Ⅰ)、蕾期(Ⅱ)、花铃期(Ⅲ)和吐絮期(Ⅳ);每个调亏阶段设置3个水分调亏度:轻度调亏(L)、中度调亏(M)和重度调亏(S),土壤相对含水率(占田间持水率的百分数)控制上、下限分别为60%~ 65% FC (Field capacity)、50%~ 55%FC和40%~45% FC;另设全生育期保持适宜土壤水分处理作为对照(CK),土壤相对含水率控制下限分别为60%FC(苗期)、60% FC(蕾期)、70% FC(花铃期)和60% FC(吐絮期).分别在水分调亏期间和复水后测定各处理根系参数和地上干物质质量.试验结果表明,RDI对植株根、冠生长发育的影响因不同水分调亏阶段和不同水分调亏度而异.水分调亏不改变棉花根系生长的原有总体趋势,但对根系生长速率具有促进作用.棉花各生育阶段的中度水分调亏(50% ~60% FC)在凋亏期间对根系生长有明显促进效应或维持较高的根质量,复水后有不同程度的根系补偿生长效应或延缓根系衰亡作用,后期仍保持较高的根冠比(R/S),因而是协调棉花根/冠关系的适宜水分调亏处理.RDI可以有效调控棉花根/冠生长关系.
DOI:10.6041/j.issn.1000-1298.2016.04.014URL [本文引用: 2]
以棉花(Gossypium hirsutum L.)为试验材料,采用防雨棚下桶栽土培方法,进行调亏灌溉(Regulated deficitirrigation,RDI)对棉花根、冠生长的影响研究.试验采用二因素随机区组设计,设置4个水分调亏阶段:苗期(Ⅰ)、蕾期(Ⅱ)、花铃期(Ⅲ)和吐絮期(Ⅳ);每个调亏阶段设置3个水分调亏度:轻度调亏(L)、中度调亏(M)和重度调亏(S),土壤相对含水率(占田间持水率的百分数)控制上、下限分别为60%~ 65% FC (Field capacity)、50%~ 55%FC和40%~45% FC;另设全生育期保持适宜土壤水分处理作为对照(CK),土壤相对含水率控制下限分别为60%FC(苗期)、60% FC(蕾期)、70% FC(花铃期)和60% FC(吐絮期).分别在水分调亏期间和复水后测定各处理根系参数和地上干物质质量.试验结果表明,RDI对植株根、冠生长发育的影响因不同水分调亏阶段和不同水分调亏度而异.水分调亏不改变棉花根系生长的原有总体趋势,但对根系生长速率具有促进作用.棉花各生育阶段的中度水分调亏(50% ~60% FC)在凋亏期间对根系生长有明显促进效应或维持较高的根质量,复水后有不同程度的根系补偿生长效应或延缓根系衰亡作用,后期仍保持较高的根冠比(R/S),因而是协调棉花根/冠关系的适宜水分调亏处理.RDI可以有效调控棉花根/冠生长关系.
URL [本文引用: 1]
干湿交替是水稻生产过程中常用的生产管理技术,通过调控稻田水分含量和土壤通气性,提高水稻根系活力,同时影响土壤中铁氧化物的存在形态。过氧化钙和过氧化尿素施用于水田土壤中可缓慢释放氧气,可能影响水田土壤中铁的形态和水稻根系活力。为了探明干湿交替和过氧化物对水稻根表铁膜及养分吸收的影响,以广东特色水稻(Oryza sativa L.)品种增城丝苗为材料,采用土壤盆栽试验研究了这两种管理方式对水稻分蘖期至拔节期的根际土壤无定形铁、结晶铁、根系活力、根系抗氧化酶活性、根表铁膜、干物质累积和养分吸收的影响。结果表明:与长期淹水处理相比,干湿交替3次分别提高水稻根际土壤中无定形铁浓度35.2%、根系活力11.3%和根表铁膜浓度55.2%,但对水稻氮、磷、钾、铁的吸收和干物质累积无明显影响;与未施用过氧化物且长期淹水相比,过氧化钙和过氧化尿素增加水稻根际土壤中无定形铁23.9%~54.2%、根系活力32.7%~58.0%和根表铁膜58.1%~169.7%,同时增加了水稻对氮、磷、钾、铁的吸收量和干物质的累积;干湿交替和过氧化物都可以增加土壤中无定形铁浓度,同时提高根系活力,从而诱导水稻根表形成更多的铁膜,其中过氧化物处理更有利于促进水稻对养分的吸收和干物质的累积。该研究为田间条件下调控水稻根表铁膜浓度提供了技术支撑。
URL [本文引用: 1]
干湿交替是水稻生产过程中常用的生产管理技术,通过调控稻田水分含量和土壤通气性,提高水稻根系活力,同时影响土壤中铁氧化物的存在形态。过氧化钙和过氧化尿素施用于水田土壤中可缓慢释放氧气,可能影响水田土壤中铁的形态和水稻根系活力。为了探明干湿交替和过氧化物对水稻根表铁膜及养分吸收的影响,以广东特色水稻(Oryza sativa L.)品种增城丝苗为材料,采用土壤盆栽试验研究了这两种管理方式对水稻分蘖期至拔节期的根际土壤无定形铁、结晶铁、根系活力、根系抗氧化酶活性、根表铁膜、干物质累积和养分吸收的影响。结果表明:与长期淹水处理相比,干湿交替3次分别提高水稻根际土壤中无定形铁浓度35.2%、根系活力11.3%和根表铁膜浓度55.2%,但对水稻氮、磷、钾、铁的吸收和干物质累积无明显影响;与未施用过氧化物且长期淹水相比,过氧化钙和过氧化尿素增加水稻根际土壤中无定形铁23.9%~54.2%、根系活力32.7%~58.0%和根表铁膜58.1%~169.7%,同时增加了水稻对氮、磷、钾、铁的吸收量和干物质的累积;干湿交替和过氧化物都可以增加土壤中无定形铁浓度,同时提高根系活力,从而诱导水稻根表形成更多的铁膜,其中过氧化物处理更有利于促进水稻对养分的吸收和干物质的累积。该研究为田间条件下调控水稻根表铁膜浓度提供了技术支撑。
DOI:10.5846/stxb201404090690URL [本文引用: 1]
由降水的不连续性引起的土壤干湿交替是荒漠生态系统土壤呼吸的重要影响因子。在恒温培养条件下,研究了固沙植被区土壤呼吸对不同降雨量(5、10、20 mm)条件下以10 d为降水周期的多重干湿交替过程的响应,结果表明:3个降雨量条件下的多重干湿交替过程中,土壤呼吸速率均在降雨后迅速增大,并在降雨0.5 h后达到最大值,尔后,随着土壤含水量的下降而逐渐减小并恢复到降雨前水平。随着干湿交替过程的依次进行,最大和平均土壤呼吸速率及累积碳释放量均呈现出逐渐减小的趋势。3个干湿循环周期平均呼吸速率和土壤碳释放量均随着降雨量的增加逐渐增大,土壤呼吸速率峰值表现为第1个干湿循环周期土壤呼吸速率峰值随着降雨量的增加而增大,而第2、3个周期各降雨处理下的土壤呼吸速率峰值显示出随着降雨前期土壤含水量的增加而减小的趋势。说明干湿交替过程对土壤呼吸具有显著的激发作用,且干湿交替程度(即降雨前后土壤含水量的变化)以及土壤前期所经历的干湿交替过程是影响荒漠生态系统土壤呼吸对干湿交替响应的重要因素。
DOI:10.5846/stxb201404090690URL [本文引用: 1]
由降水的不连续性引起的土壤干湿交替是荒漠生态系统土壤呼吸的重要影响因子。在恒温培养条件下,研究了固沙植被区土壤呼吸对不同降雨量(5、10、20 mm)条件下以10 d为降水周期的多重干湿交替过程的响应,结果表明:3个降雨量条件下的多重干湿交替过程中,土壤呼吸速率均在降雨后迅速增大,并在降雨0.5 h后达到最大值,尔后,随着土壤含水量的下降而逐渐减小并恢复到降雨前水平。随着干湿交替过程的依次进行,最大和平均土壤呼吸速率及累积碳释放量均呈现出逐渐减小的趋势。3个干湿循环周期平均呼吸速率和土壤碳释放量均随着降雨量的增加逐渐增大,土壤呼吸速率峰值表现为第1个干湿循环周期土壤呼吸速率峰值随着降雨量的增加而增大,而第2、3个周期各降雨处理下的土壤呼吸速率峰值显示出随着降雨前期土壤含水量的增加而减小的趋势。说明干湿交替过程对土壤呼吸具有显著的激发作用,且干湿交替程度(即降雨前后土壤含水量的变化)以及土壤前期所经历的干湿交替过程是影响荒漠生态系统土壤呼吸对干湿交替响应的重要因素。
DOI:10.3724/SP.J.1011.2014.31093URL [本文引用: 1]
在水稻高产栽培技术中,为合理利用水资源,改良土壤通气性,干湿交替灌溉技术已广泛应用.干湿交替灌溉技术的核心是协调土壤水分与氧气之间的平衡.本研究采用大田试验,在水稻生育后期进行干湿交替处理,同步监测稻田5 cm土层土壤氧气、水分及温度的变化,以探讨稻田土壤氧气和水分运移变化规律及其互作关系,为进一步揭示干湿交替灌溉技术的内在生理机制奠定基础.研究结果表明,干湿交替条件下,稻田土壤含水量处于饱和状态时,基本监测不到土壤含氧量;而稻田土壤在慢慢变干的过程中,5 cm土层土壤体积水分含量逐渐降低,土壤氧气含量逐渐升高.在花后19d、24d、29d,当5 cm土层土壤体积含水量分别下降为25.4%、25.1%、24.7%时,土壤含氧量则分别上升到17.5%、17.4%、17.4%.在水稻生育后期不同阶段,土壤氧气含量的日变化呈现先降低再升高的趋势,谷值一般出现在14:00-15:00之间;土壤含水量随时间呈波动式逐渐降低的变化趋势;土壤温度呈现先升高后降低的趋势,峰值一般出现在15:00-16:00之间.从上午8:00到下午17:59,当土壤温度升至峰值时,土壤水分含量较低,而土壤含氧量开始升高.水稻开花期、灌浆期和成熟期的土壤氧气含量与土壤含水量均表现出极显著负相关关系,土壤温度与土壤水分间呈显著负相关关系,而土壤氧气与土壤温度之间无显著相关关系.说明干湿交替条件下,水稻生育后期稻田土壤中的水分和氧气含量存在一定的此消彼长的关系.因此,通过适度的干湿交替管理措施,可在一定程度上调节水稻根系周围的土壤水分和氧气的平衡.
DOI:10.3724/SP.J.1011.2014.31093URL [本文引用: 1]
在水稻高产栽培技术中,为合理利用水资源,改良土壤通气性,干湿交替灌溉技术已广泛应用.干湿交替灌溉技术的核心是协调土壤水分与氧气之间的平衡.本研究采用大田试验,在水稻生育后期进行干湿交替处理,同步监测稻田5 cm土层土壤氧气、水分及温度的变化,以探讨稻田土壤氧气和水分运移变化规律及其互作关系,为进一步揭示干湿交替灌溉技术的内在生理机制奠定基础.研究结果表明,干湿交替条件下,稻田土壤含水量处于饱和状态时,基本监测不到土壤含氧量;而稻田土壤在慢慢变干的过程中,5 cm土层土壤体积水分含量逐渐降低,土壤氧气含量逐渐升高.在花后19d、24d、29d,当5 cm土层土壤体积含水量分别下降为25.4%、25.1%、24.7%时,土壤含氧量则分别上升到17.5%、17.4%、17.4%.在水稻生育后期不同阶段,土壤氧气含量的日变化呈现先降低再升高的趋势,谷值一般出现在14:00-15:00之间;土壤含水量随时间呈波动式逐渐降低的变化趋势;土壤温度呈现先升高后降低的趋势,峰值一般出现在15:00-16:00之间.从上午8:00到下午17:59,当土壤温度升至峰值时,土壤水分含量较低,而土壤含氧量开始升高.水稻开花期、灌浆期和成熟期的土壤氧气含量与土壤含水量均表现出极显著负相关关系,土壤温度与土壤水分间呈显著负相关关系,而土壤氧气与土壤温度之间无显著相关关系.说明干湿交替条件下,水稻生育后期稻田土壤中的水分和氧气含量存在一定的此消彼长的关系.因此,通过适度的干湿交替管理措施,可在一定程度上调节水稻根系周围的土壤水分和氧气的平衡.
DOI:10.3321/j.issn:1000-0933.2009.06.031URL [本文引用: 1]
全球气候变化的直接后果是气温 升高,同时还可能引起强降雨增多和干旱频发,形成干湿交替的格局。土壤呼吸在全球变化过程中发挥着重要作用。以黄土高原沟壑区小麦田土壤为研究对象,采用 3个全自动多通量箱以及相应的气象监测系统,对土壤呼吸和环境因子全天候连续测定,利用已有的单因子模型、双因子模型对测定的土壤呼吸与气温和湿度的关系 进行了拟合,通过优化,根据实际情况提出E-Q(exponential-quadratic)模型。结果表明:(1)干湿交替格局下,基于气温的单因子 模型(指数模型,幂函数模型和线性模型)不适合模拟土壤呼吸;(2)基于土壤湿度的单因子模型中,二次曲线模型最适合模拟干湿交替格局下土壤呼吸的响应情 况;(3)基于气温和土壤湿度的双因子模型中,E-Q模型SR=aebT(c+dW+fW2)g,既能反映土壤呼吸随气温的正向指数变化,又能表现土壤湿 度对土壤呼吸的双向调节作用,解释了土壤呼吸73.05%的变化情况,比其他双因子模型和单因子模型更能有效描述干湿交替情况下土壤呼吸对气温和土壤湿度 协同变化的响应特征。
DOI:10.3321/j.issn:1000-0933.2009.06.031URL [本文引用: 1]
全球气候变化的直接后果是气温 升高,同时还可能引起强降雨增多和干旱频发,形成干湿交替的格局。土壤呼吸在全球变化过程中发挥着重要作用。以黄土高原沟壑区小麦田土壤为研究对象,采用 3个全自动多通量箱以及相应的气象监测系统,对土壤呼吸和环境因子全天候连续测定,利用已有的单因子模型、双因子模型对测定的土壤呼吸与气温和湿度的关系 进行了拟合,通过优化,根据实际情况提出E-Q(exponential-quadratic)模型。结果表明:(1)干湿交替格局下,基于气温的单因子 模型(指数模型,幂函数模型和线性模型)不适合模拟土壤呼吸;(2)基于土壤湿度的单因子模型中,二次曲线模型最适合模拟干湿交替格局下土壤呼吸的响应情 况;(3)基于气温和土壤湿度的双因子模型中,E-Q模型SR=aebT(c+dW+fW2)g,既能反映土壤呼吸随气温的正向指数变化,又能表现土壤湿 度对土壤呼吸的双向调节作用,解释了土壤呼吸73.05%的变化情况,比其他双因子模型和单因子模型更能有效描述干湿交替情况下土壤呼吸对气温和土壤湿度 协同变化的响应特征。
DOI:10.3864/j.issn.0578-1752.2013.11.024URL [本文引用: 1]
【目的】明确持续干旱胁迫对棉花叶片光谱、光合和荧光特征的影响,为筛选水分敏感指标提供理论依据。【方法】以转基因抗虫棉银瑞361为试材,采用盆栽控水方式,测定持续干旱与复水条件下棉花叶片光谱、光合和荧光参数。【结果】两年试验结果表明,随着干旱胁迫时间的延长叶片相对含水量(RWC)、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ实际光化学量子产量(ΦPSⅡ)、光化学淬灭系数(qP)、光化学反射指数(PRI)呈下降趋势,气孔限制值(Ls)先上升后下降,非光化学淬灭系数(NPQ)则呈上升趋势,复水后除RWC外,其它指标都未恢复到初始状态。与初始状态相比,Pn、Gs、Tr、Fv/Fm、ΦPSⅡ、qP、PRI在干旱胁迫第4天时,两年平均下降了53.1%、79.9%、66.0%、3.8%、11.0%、5.7%、18.4%;Ls、NPQ在第4天时,两年平均上升了98.1%、28.6%。综合两年数据,只有Pn、Gs、Tr、Ls、Fv/Fm、PRI在第4天时与初始状态差异极显著,与RWC的R2分别为0.7640、0.7058、0.5625、0.3665、0.7319、0.4378。【结论】Pn、Gs、Tr、Ls、Fv/Fm、PRI均能在干旱胁迫初期灵敏地反映棉花叶片水分状况;在持续干旱胁迫中,Pn、Gs、Fv/Fm能更灵敏、准确地反映棉花叶片水分状况。
DOI:10.3864/j.issn.0578-1752.2013.11.024URL [本文引用: 1]
【目的】明确持续干旱胁迫对棉花叶片光谱、光合和荧光特征的影响,为筛选水分敏感指标提供理论依据。【方法】以转基因抗虫棉银瑞361为试材,采用盆栽控水方式,测定持续干旱与复水条件下棉花叶片光谱、光合和荧光参数。【结果】两年试验结果表明,随着干旱胁迫时间的延长叶片相对含水量(RWC)、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ实际光化学量子产量(ΦPSⅡ)、光化学淬灭系数(qP)、光化学反射指数(PRI)呈下降趋势,气孔限制值(Ls)先上升后下降,非光化学淬灭系数(NPQ)则呈上升趋势,复水后除RWC外,其它指标都未恢复到初始状态。与初始状态相比,Pn、Gs、Tr、Fv/Fm、ΦPSⅡ、qP、PRI在干旱胁迫第4天时,两年平均下降了53.1%、79.9%、66.0%、3.8%、11.0%、5.7%、18.4%;Ls、NPQ在第4天时,两年平均上升了98.1%、28.6%。综合两年数据,只有Pn、Gs、Tr、Ls、Fv/Fm、PRI在第4天时与初始状态差异极显著,与RWC的R2分别为0.7640、0.7058、0.5625、0.3665、0.7319、0.4378。【结论】Pn、Gs、Tr、Ls、Fv/Fm、PRI均能在干旱胁迫初期灵敏地反映棉花叶片水分状况;在持续干旱胁迫中,Pn、Gs、Fv/Fm能更灵敏、准确地反映棉花叶片水分状况。
DOI:10.3321/j.issn:1008-505X.2009.03.026URL
2005~2006年在南京农业大学卫岗试验站进行盆栽试验,设置正常灌水和棉花花铃期土壤 短期干旱处理,每个处理再设置3个氮素水平(N 0、240、480 kg/hm^2),研究氮素对花铃期短期干旱再复水后棉花纤维比强度形成的影响。结果表明,干旱处理结束时,干旱处理棉花的纤维可溶性蛋白含量较正常灌水 处理显著降低,而内源保护酶,即:超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性升高,丙二醛(MDA)含量增加;纤维加厚 发育相关酶,即:蔗糖合成酶、磷酸蔗糖合成酶、β-1,3-葡聚糖酶和IAA氧化酶的活性均显著降低。在复水后第10 d,干旱处理棉花的纤维内源保护酶活性迅速恢复到正常灌水处理水平,MDA含量降低,但纤维发育相关酶的活性仍低于相应正常灌水处理,纤维比强度亦显著低 于正常灌水处理。在棉花的纤维加厚发育期土壤短期干旱再复水条件下,以N 240 kg/hm^2最有利于形成高强纤维。干旱期间该处理棉花的纤维内源保护酶活性高,细胞膜脂过氧化程度最低,其纤维加厚发育相关酶活性均最高;复水后该处 理的纤维内源保护酶活性迅速恢复,MDA含量最低,棉花的纤维加厚发育相关酶活性仍处于最高值,有利于纤维素的合成与累积,最终纤维比强度亦最大。施氮不 足(N 0 kg/hm^2)或过量施氮(N 480 kg/hm^2)均表现出相反的趋势。
DOI:10.3321/j.issn:1008-505X.2009.03.026URL
2005~2006年在南京农业大学卫岗试验站进行盆栽试验,设置正常灌水和棉花花铃期土壤 短期干旱处理,每个处理再设置3个氮素水平(N 0、240、480 kg/hm^2),研究氮素对花铃期短期干旱再复水后棉花纤维比强度形成的影响。结果表明,干旱处理结束时,干旱处理棉花的纤维可溶性蛋白含量较正常灌水 处理显著降低,而内源保护酶,即:超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性升高,丙二醛(MDA)含量增加;纤维加厚 发育相关酶,即:蔗糖合成酶、磷酸蔗糖合成酶、β-1,3-葡聚糖酶和IAA氧化酶的活性均显著降低。在复水后第10 d,干旱处理棉花的纤维内源保护酶活性迅速恢复到正常灌水处理水平,MDA含量降低,但纤维发育相关酶的活性仍低于相应正常灌水处理,纤维比强度亦显著低 于正常灌水处理。在棉花的纤维加厚发育期土壤短期干旱再复水条件下,以N 240 kg/hm^2最有利于形成高强纤维。干旱期间该处理棉花的纤维内源保护酶活性高,细胞膜脂过氧化程度最低,其纤维加厚发育相关酶活性均最高;复水后该处 理的纤维内源保护酶活性迅速恢复,MDA含量最低,棉花的纤维加厚发育相关酶活性仍处于最高值,有利于纤维素的合成与累积,最终纤维比强度亦最大。施氮不 足(N 0 kg/hm^2)或过量施氮(N 480 kg/hm^2)均表现出相反的趋势。
URL
本试验以抗旱差异明显的中棉所45(CCRI-45)和中棉所60(CCRI-60)为材料,采用人工控水的方法,研究棉花花铃期不同干旱胁迫程度和干旱胁迫历时复水后棉花根系干物重、根系生理指标、皮棉产量和纤维品质的变化。主要结果如下:1、CCRI-45根系干物重和皮棉产量在SRWC((45±5)%)干旱胁迫6 d复水处理下最大,CCRI-60根系干物重和皮棉产量在SRWC((60±5)%)干旱胁迫6 d复水处理下最大,但CCRI-45和CCRI-60根系干物重和皮棉产量在SRWC((45±5)%)干旱胁迫9 d复水处理下显著低于对照。CCRI-45上半部平均长度、整齐度指数、伸长率和断裂比强度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6d复水处理下均高于对照,CCRI-60上半部平均长度、整齐度指数、伸长率和断裂比强度在SRWC((45±5)%)干旱胁迫3 d和SRWC((60±5)%)干旱胁迫6 d复水处理下均高于对照,CCRI-45和CCRI-60在SRWC((45±5)%)干旱胁迫9 d复水处理下纤维品质显著低与对照。CCRI-45根系干物重和棉花皮棉产量的增加幅度在SRWC((60±5)%)干旱胁迫6 d复水处理下高于CCRI-60,表明CCRI-45的补偿能力较强。同时,棉花根系干物重增加量与棉花皮棉增加量存在显著(p0.05)的正相关关系。2、随干旱胁迫程度的增加和干旱胁迫历时的延长,根系可溶性糖和脯氨酸含量增加幅度呈增大趋势,棉花根系可溶性蛋白含量在干旱胁迫3 d时降低,但干旱胁迫6 d和9 d时根系可溶性蛋白含量升高,复水后,根系可溶性糖和脯氨酸含量降低,可溶性蛋白含量在干旱胁迫3 d复水后升高,干旱胁迫6 d和9 d复水后降低,且根系可溶性糖、脯氨酸和溶性蛋白含量仍高于对照。CCRI-45根系可溶性糖、脯氨酸和可溶性蛋白含量增加幅度在SRWC((45±5)%)干旱胁迫6 d复复水处理下明显高于CCRI-60,表明,复水后CCRI-45根系中较高可溶性糖、脯氨酸和可溶性蛋白含量增强了细胞的渗透调节能力,进而提高了棉花根系的抗旱和补偿能力。3、随干旱胁迫程度的增加和干旱胁迫历时的延长,棉花根系赤霉素(GA)含量降低幅度呈增大趋势,根系油菜素内酯(BRs)含量增加幅度呈先升高后降低趋势,根系脱落酸(ABA)和乙烯(Eth)含量增加幅度呈增大趋势,复水后,根系GA含量升高,根系BRs、ABA和Eth含量降低。CCRI-45根系GA和BRs含量增加幅度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,CCRI-45根系ABA和Eth含量降低幅度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,表明,根系中较高GA和BRs含量促进了根系的恢复生长,弥补干旱期间造成的伤害,提高了棉花根系的补偿能力。4、随干旱胁迫程度的增加和干旱胁迫历时的延长,根系超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的增加幅度呈增大趋势,根系过氧化氢(CAT)活性降低幅度呈增大趋势,复水后,根系SOD和POD活性下降,根系CAT活性升高。CCRI-45根系SOD和POD活性增加幅度均在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫9 d复水处理下明显高于CCRI-60,CCRI-45根系CAT活性增加幅度在SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,表明,根系在干旱胁迫复水后保持较高的抗氧化酶活性通过降低活性氧含量,防止细胞损伤,保证根系生理过程正常进行,进而提高棉花根系的补偿能力。5、随干旱胁迫程度的增加和干旱历时的延长,CCRI-45根系干物重、总根长、根系表面积、根系总体积及根平均直径呈先升高后降低的趋势,CCRI-60根干物重和总根长呈呈先升高后降低趋势,而根系表面积、根系总体积和根平均直径均呈降低趋势。CCRI-45在10%PEG干旱胁迫24h、48h和20%PEG干旱胁迫24h处理下根系干物重、总根长、根系表面积、根系总体积和根平均直径均高于对照,CCRI-60在10%PEG干旱胁迫24h处理下根干物重和总根长高于对照,且CCRI-45根干物重和总根长增加幅度在10%PEG干旱胁迫24h处理下明显高于CCRI-60。随着干旱胁迫的增加和干旱历时的延长,细胞质壁分离严重,细胞核内容物降解,尤其线粒体内部结构降解。CCRI-45根系细胞中线粒体内部分解程度在20%PEG干旱胁迫48h处理下明显小于CCRI-60,表明抗旱性强的品种在轻度和中度干旱胁迫下,线粒体的结构比较稳定,可以保证胁迫过程中根系的能量供应,提高其抗旱性。
URL
本试验以抗旱差异明显的中棉所45(CCRI-45)和中棉所60(CCRI-60)为材料,采用人工控水的方法,研究棉花花铃期不同干旱胁迫程度和干旱胁迫历时复水后棉花根系干物重、根系生理指标、皮棉产量和纤维品质的变化。主要结果如下:1、CCRI-45根系干物重和皮棉产量在SRWC((45±5)%)干旱胁迫6 d复水处理下最大,CCRI-60根系干物重和皮棉产量在SRWC((60±5)%)干旱胁迫6 d复水处理下最大,但CCRI-45和CCRI-60根系干物重和皮棉产量在SRWC((45±5)%)干旱胁迫9 d复水处理下显著低于对照。CCRI-45上半部平均长度、整齐度指数、伸长率和断裂比强度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6d复水处理下均高于对照,CCRI-60上半部平均长度、整齐度指数、伸长率和断裂比强度在SRWC((45±5)%)干旱胁迫3 d和SRWC((60±5)%)干旱胁迫6 d复水处理下均高于对照,CCRI-45和CCRI-60在SRWC((45±5)%)干旱胁迫9 d复水处理下纤维品质显著低与对照。CCRI-45根系干物重和棉花皮棉产量的增加幅度在SRWC((60±5)%)干旱胁迫6 d复水处理下高于CCRI-60,表明CCRI-45的补偿能力较强。同时,棉花根系干物重增加量与棉花皮棉增加量存在显著(p0.05)的正相关关系。2、随干旱胁迫程度的增加和干旱胁迫历时的延长,根系可溶性糖和脯氨酸含量增加幅度呈增大趋势,棉花根系可溶性蛋白含量在干旱胁迫3 d时降低,但干旱胁迫6 d和9 d时根系可溶性蛋白含量升高,复水后,根系可溶性糖和脯氨酸含量降低,可溶性蛋白含量在干旱胁迫3 d复水后升高,干旱胁迫6 d和9 d复水后降低,且根系可溶性糖、脯氨酸和溶性蛋白含量仍高于对照。CCRI-45根系可溶性糖、脯氨酸和可溶性蛋白含量增加幅度在SRWC((45±5)%)干旱胁迫6 d复复水处理下明显高于CCRI-60,表明,复水后CCRI-45根系中较高可溶性糖、脯氨酸和可溶性蛋白含量增强了细胞的渗透调节能力,进而提高了棉花根系的抗旱和补偿能力。3、随干旱胁迫程度的增加和干旱胁迫历时的延长,棉花根系赤霉素(GA)含量降低幅度呈增大趋势,根系油菜素内酯(BRs)含量增加幅度呈先升高后降低趋势,根系脱落酸(ABA)和乙烯(Eth)含量增加幅度呈增大趋势,复水后,根系GA含量升高,根系BRs、ABA和Eth含量降低。CCRI-45根系GA和BRs含量增加幅度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,CCRI-45根系ABA和Eth含量降低幅度在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,表明,根系中较高GA和BRs含量促进了根系的恢复生长,弥补干旱期间造成的伤害,提高了棉花根系的补偿能力。4、随干旱胁迫程度的增加和干旱胁迫历时的延长,根系超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的增加幅度呈增大趋势,根系过氧化氢(CAT)活性降低幅度呈增大趋势,复水后,根系SOD和POD活性下降,根系CAT活性升高。CCRI-45根系SOD和POD活性增加幅度均在SRWC((60±5)%)和SRWC((45±5)%)干旱胁迫9 d复水处理下明显高于CCRI-60,CCRI-45根系CAT活性增加幅度在SRWC((45±5)%)干旱胁迫6 d复水处理下明显高于CCRI-60,表明,根系在干旱胁迫复水后保持较高的抗氧化酶活性通过降低活性氧含量,防止细胞损伤,保证根系生理过程正常进行,进而提高棉花根系的补偿能力。5、随干旱胁迫程度的增加和干旱历时的延长,CCRI-45根系干物重、总根长、根系表面积、根系总体积及根平均直径呈先升高后降低的趋势,CCRI-60根干物重和总根长呈呈先升高后降低趋势,而根系表面积、根系总体积和根平均直径均呈降低趋势。CCRI-45在10%PEG干旱胁迫24h、48h和20%PEG干旱胁迫24h处理下根系干物重、总根长、根系表面积、根系总体积和根平均直径均高于对照,CCRI-60在10%PEG干旱胁迫24h处理下根干物重和总根长高于对照,且CCRI-45根干物重和总根长增加幅度在10%PEG干旱胁迫24h处理下明显高于CCRI-60。随着干旱胁迫的增加和干旱历时的延长,细胞质壁分离严重,细胞核内容物降解,尤其线粒体内部结构降解。CCRI-45根系细胞中线粒体内部分解程度在20%PEG干旱胁迫48h处理下明显小于CCRI-60,表明抗旱性强的品种在轻度和中度干旱胁迫下,线粒体的结构比较稳定,可以保证胁迫过程中根系的能量供应,提高其抗旱性。
URL [本文引用: 2]
在玻璃日光温室条件下,利用盆栽人工控水的方法,研究不同程度的水分亏缺对玉米干物质累积、根系活力、水分利用效率(WUE)、脯氨酸(Pro)含量、超氧化物歧化酶(SOD)活性的影响及复水后的变化。结果表明:水分亏缺显著抑制玉米植株的生长、叶面积的扩展及干物质的累积,抑制程度与亏缺程度有关,复水后的株高和叶面积得到了较大幅度的补偿,随着亏缺程度的加重,根冠比、根系活力除了重度亏缺外均有所提高,WUE依次下降,Pro的累积量显著增加,而SOD活性先增大后减小。复水后各处理的WUE均高于对照,SOD活性除中度略高于对照外,其余均低于对照。
URL [本文引用: 2]
在玻璃日光温室条件下,利用盆栽人工控水的方法,研究不同程度的水分亏缺对玉米干物质累积、根系活力、水分利用效率(WUE)、脯氨酸(Pro)含量、超氧化物歧化酶(SOD)活性的影响及复水后的变化。结果表明:水分亏缺显著抑制玉米植株的生长、叶面积的扩展及干物质的累积,抑制程度与亏缺程度有关,复水后的株高和叶面积得到了较大幅度的补偿,随着亏缺程度的加重,根冠比、根系活力除了重度亏缺外均有所提高,WUE依次下降,Pro的累积量显著增加,而SOD活性先增大后减小。复水后各处理的WUE均高于对照,SOD活性除中度略高于对照外,其余均低于对照。
[本文引用: 4]
[本文引用: 4]
DOI:10.6048/j.issn.1001-4330.2013.11.002URL [本文引用: 3]
[目的]研究在水分胁迫下施氮量对抗旱春小麦品种花后旗叶生理特 性的影响.[方法]在大田试验条件下,以抗旱春小麦新品种新旱688为实验材料.以大田常规灌水为对照,以全生育期灌水两次为胁迫;采用0、150、 300和450 kg/hm2四种氮处理,研究水分胁迫下氮肥用量对新旱688主要生理特性的影响.[结果]水分胁迫下,在一定的施氮范围内叶片中Chl(a+b)、 Chla、Chlb、全N含量随着施氮量的增加而增加,Chla/Chlb值随着施氮量的增加逐渐减小.在0~300 kg/hm2施氮范围内净光合速率升高,胞间CO2浓度降低,当施氮量达到450 kg/hm2时,胞间CO2浓度开始升高,光合速率反而下降.干旱时适当施氮有利于可溶性糖的积累,在150 kg/hm2氮素水平时可溶性糖积累量高于其他氮素水平.[结论]水分胁迫下,适当施用氮肥有利于小麦花后旗叶生理特性的改善.
DOI:10.6048/j.issn.1001-4330.2013.11.002URL [本文引用: 3]
[目的]研究在水分胁迫下施氮量对抗旱春小麦品种花后旗叶生理特 性的影响.[方法]在大田试验条件下,以抗旱春小麦新品种新旱688为实验材料.以大田常规灌水为对照,以全生育期灌水两次为胁迫;采用0、150、 300和450 kg/hm2四种氮处理,研究水分胁迫下氮肥用量对新旱688主要生理特性的影响.[结果]水分胁迫下,在一定的施氮范围内叶片中Chl(a+b)、 Chla、Chlb、全N含量随着施氮量的增加而增加,Chla/Chlb值随着施氮量的增加逐渐减小.在0~300 kg/hm2施氮范围内净光合速率升高,胞间CO2浓度降低,当施氮量达到450 kg/hm2时,胞间CO2浓度开始升高,光合速率反而下降.干旱时适当施氮有利于可溶性糖的积累,在150 kg/hm2氮素水平时可溶性糖积累量高于其他氮素水平.[结论]水分胁迫下,适当施用氮肥有利于小麦花后旗叶生理特性的改善.
URL [本文引用: 1]
以“花育22号”花生为试验材料,在中度干旱胁迫和充足灌水两个水分条件下,分别设置不施氮肥(N0)、中氮(N1,90 kg·hm-2)、高氮(N2,180 kg·hm-2)3个施氮水平,研究不同土壤水分和氮肥条件对花生叶片生理活性及根系形态发育特征的影响.结果表明:与不施氮肥处理相比,两个水分条件下中氮处理均显著增加花生产量,但对收获指数无显著影响.干旱胁迫条件下,中氮处理对总根系生物量和总根长无显著影响,但显著增加花生总根系表面积;中氮和高氮处理均显著增加20 ~ 40 cm土层内根长和根系表面积,且高氮处理显著增加40 cm以下土层内根系生物量和根系表面积;施用氮肥显著提高叶片过氧化氢酶(CAT)和过氧化物酶(POD)活性,而丙二醛(MDA)含量随施氮量的增加而降低.正常供水条件下,施用氮肥显著降低了花生根系表面积和40 cm以下土层内根系生物量、根长和根系表面积,中氮处理可提高叶片保护酶活性.相关性分析表明,20 ~ 40 cm土层内根长和叶片超氧化物歧化酶(SOD)、CAT、POD活性与产量呈显著相关.
URL [本文引用: 1]
以“花育22号”花生为试验材料,在中度干旱胁迫和充足灌水两个水分条件下,分别设置不施氮肥(N0)、中氮(N1,90 kg·hm-2)、高氮(N2,180 kg·hm-2)3个施氮水平,研究不同土壤水分和氮肥条件对花生叶片生理活性及根系形态发育特征的影响.结果表明:与不施氮肥处理相比,两个水分条件下中氮处理均显著增加花生产量,但对收获指数无显著影响.干旱胁迫条件下,中氮处理对总根系生物量和总根长无显著影响,但显著增加花生总根系表面积;中氮和高氮处理均显著增加20 ~ 40 cm土层内根长和根系表面积,且高氮处理显著增加40 cm以下土层内根系生物量和根系表面积;施用氮肥显著提高叶片过氧化氢酶(CAT)和过氧化物酶(POD)活性,而丙二醛(MDA)含量随施氮量的增加而降低.正常供水条件下,施用氮肥显著降低了花生根系表面积和40 cm以下土层内根系生物量、根长和根系表面积,中氮处理可提高叶片保护酶活性.相关性分析表明,20 ~ 40 cm土层内根长和叶片超氧化物歧化酶(SOD)、CAT、POD活性与产量呈显著相关.
DOI:103969/jissn1000-6362201403002Magsci [本文引用: 2]
2011-2013年,在大田利用人工防雨棚开展了不同干旱水平下的冬小麦种植试验,设置土壤相对湿度分别为80%(CK)、60%(T2)和40%(T1)3个处理水平,对各处理不同生育期各器官干物质质量、可溶性糖含量和产量及其构成要素进行观测分析。结果表明,冬小麦不同器官干物质质量和可溶性糖含量因受干旱影响程度、胁迫时间以及发育期进程的不同而发生改变。与CK相比,T2处理(60%)冬小麦在受胁迫初期茎和鞘干物质质量出现不同程度的增加,扬花期及以后发育期表现为减少趋势;而叶和根干物质质量总体上在整个生育期均呈现不同程度的减少,前期减幅大后期减幅小;不同营养器官可溶性糖含量均有不同程度增加;产量构成要素主要表现为不孕小穗增加、穗粒重减小和千粒重下降。T1处理(40%)冬小麦根、茎、叶和鞘干物质质量均有大幅减少;冬小麦产量构成要素均受到明显影响(P<0.05)。干旱条件下(T2和T1处理),冬小麦茎秆干物质转运量和转运效率较其它营养器官显著提高(P<0.05),但与对照差异不显著。结论:干旱造成冬小麦不同器官干物资分配格局发生改变,可溶糖含量升高;促使茎秆干物质向穗部转移效率增加、对籽粒贡献率提高;导致冬小麦衰老提前和产量降低。
DOI:103969/jissn1000-6362201403002Magsci [本文引用: 2]
2011-2013年,在大田利用人工防雨棚开展了不同干旱水平下的冬小麦种植试验,设置土壤相对湿度分别为80%(CK)、60%(T2)和40%(T1)3个处理水平,对各处理不同生育期各器官干物质质量、可溶性糖含量和产量及其构成要素进行观测分析。结果表明,冬小麦不同器官干物质质量和可溶性糖含量因受干旱影响程度、胁迫时间以及发育期进程的不同而发生改变。与CK相比,T2处理(60%)冬小麦在受胁迫初期茎和鞘干物质质量出现不同程度的增加,扬花期及以后发育期表现为减少趋势;而叶和根干物质质量总体上在整个生育期均呈现不同程度的减少,前期减幅大后期减幅小;不同营养器官可溶性糖含量均有不同程度增加;产量构成要素主要表现为不孕小穗增加、穗粒重减小和千粒重下降。T1处理(40%)冬小麦根、茎、叶和鞘干物质质量均有大幅减少;冬小麦产量构成要素均受到明显影响(P<0.05)。干旱条件下(T2和T1处理),冬小麦茎秆干物质转运量和转运效率较其它营养器官显著提高(P<0.05),但与对照差异不显著。结论:干旱造成冬小麦不同器官干物资分配格局发生改变,可溶糖含量升高;促使茎秆干物质向穗部转移效率增加、对籽粒贡献率提高;导致冬小麦衰老提前和产量降低。
URLMagsci [本文引用: 1]
<p>通过真空表式土壤张力计监测土壤水势,以正常氮处理的烟株为对照,对生育后期由于施肥过多而严重影响烟叶品质的烤烟K326和NC89,进行有控制的水分亏缺处理,研究以水控氮对烤烟光合特性和氮代谢的影响,探讨以水调肥改善烟叶品质的生理原因。结果表明:两种氮肥营养下,土壤水分亏缺降低了叶片的净光合速率和蒸腾速率,而施高氮较好的抑制了这种下降;适度的土壤水分亏缺抑制了硝酸还原酶的活性,降低了中上部叶全氮的含量,从而有效抑制了高氮营养下烤烟成熟期的氮代谢。</p>
URLMagsci [本文引用: 1]
<p>通过真空表式土壤张力计监测土壤水势,以正常氮处理的烟株为对照,对生育后期由于施肥过多而严重影响烟叶品质的烤烟K326和NC89,进行有控制的水分亏缺处理,研究以水控氮对烤烟光合特性和氮代谢的影响,探讨以水调肥改善烟叶品质的生理原因。结果表明:两种氮肥营养下,土壤水分亏缺降低了叶片的净光合速率和蒸腾速率,而施高氮较好的抑制了这种下降;适度的土壤水分亏缺抑制了硝酸还原酶的活性,降低了中上部叶全氮的含量,从而有效抑制了高氮营养下烤烟成熟期的氮代谢。</p>
DOI:10.7606/j.issn.1009-1041.2012.04.015URL [本文引用: 3]
为了解水分亏缺下小麦穗部光合对产量的贡献,采用盆栽试验及14 C-同位素示踪方法,研究了水分亏缺对不同抗旱性小麦花后穗部光合及穗光合产物向籽粒转运及产量的影响。结果表明,与正常供水处理相比,水分亏缺后,旱地品种西农1043灌浆前期穗部净光合速率、颖壳中可溶性总糖含量、籽粒14 C-同化物分配率增加,而水地品种陕253均下降。收获后,水分亏缺显著降低了陕253颖壳及内外稃中光合同化物滞留率、籽粒产量和水分利用效率(P0.01),而对西农1043的影响不明显(P0.05)。说明适度水分亏缺会提高旱地小麦灌浆前期穗部的碳同化能力,加速灌浆前期穗部光合产物向籽粒的转运,以维持一定的产量水平。
DOI:10.7606/j.issn.1009-1041.2012.04.015URL [本文引用: 3]
为了解水分亏缺下小麦穗部光合对产量的贡献,采用盆栽试验及14 C-同位素示踪方法,研究了水分亏缺对不同抗旱性小麦花后穗部光合及穗光合产物向籽粒转运及产量的影响。结果表明,与正常供水处理相比,水分亏缺后,旱地品种西农1043灌浆前期穗部净光合速率、颖壳中可溶性总糖含量、籽粒14 C-同化物分配率增加,而水地品种陕253均下降。收获后,水分亏缺显著降低了陕253颖壳及内外稃中光合同化物滞留率、籽粒产量和水分利用效率(P0.01),而对西农1043的影响不明显(P0.05)。说明适度水分亏缺会提高旱地小麦灌浆前期穗部的碳同化能力,加速灌浆前期穗部光合产物向籽粒的转运,以维持一定的产量水平。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:0496-3490.2003.04.022URL [本文引用: 1]
以防雨池栽的方式研究了限量灌溉对冬小麦籽粒淀粉合成和积累的影响。结果表明 ,冬小麦籽粒中可溶性淀粉合成酶 (SSS)、淀粉粒结合态合成酶 (GBSS)和Q酶 (一种分支酶 )均是在灌浆中期 (花后 14d)活性最高。中度、严重干旱显著地降低了它们的活性。适宜的灌水处理 (处理C和处理D)使三种酶保持较高的活性 ,这是形成较高产量的生理基础。三种酶的活性均是鲁麦 2 1的较高。土壤水分亏缺严重 (处理A)显著降低籽粒中淀粉、支链淀粉和直链淀粉的积累 ,灌麦黄水最终减少了它们的含量。适宜的灌水处理 (处理C和处理D)使它们的含量增加。鲁麦 2 1的淀粉含量、支链淀粉含量高于济南 17,而直链淀粉含
DOI:10.3321/j.issn:0496-3490.2003.04.022URL [本文引用: 1]
以防雨池栽的方式研究了限量灌溉对冬小麦籽粒淀粉合成和积累的影响。结果表明 ,冬小麦籽粒中可溶性淀粉合成酶 (SSS)、淀粉粒结合态合成酶 (GBSS)和Q酶 (一种分支酶 )均是在灌浆中期 (花后 14d)活性最高。中度、严重干旱显著地降低了它们的活性。适宜的灌水处理 (处理C和处理D)使三种酶保持较高的活性 ,这是形成较高产量的生理基础。三种酶的活性均是鲁麦 2 1的较高。土壤水分亏缺严重 (处理A)显著降低籽粒中淀粉、支链淀粉和直链淀粉的积累 ,灌麦黄水最终减少了它们的含量。适宜的灌水处理 (处理C和处理D)使它们的含量增加。鲁麦 2 1的淀粉含量、支链淀粉含量高于济南 17,而直链淀粉含
DOI:10.1016/j.plantsci.2011.01.009URL [本文引用: 1]
DOI:10.6041/j.issn.1000-1298.2013.03.017URL [本文引用: 1]
通过盆栽试验研究了不同水氮互作条件下水稻各项生理指标短期和长期的后效变化。结果表明:适宜的水氮组合复水短期内对生理活性有一定的补偿效应,生育后期表现出延缓衰老的长期后效性;水、氮单因子复水短期内对叶片叶绿素含量、荧光参数、丙二醛含量等生理指标影响显著,水氮协同互作效应在复水15d后达显著水平;不同的水分胁迫后复水,水氮互作效应不同,重旱处理产量随复水后施氮量的增加而增加,但产量较其他处理明显降低。应避免水稻拔节期重旱,土壤含水率不能低于田间持水率的70%;充分灌溉和轻旱处理产量随施氮量的增加有所降低,拔节期高水高肥不一定高产,轻旱后复水结合低氮追肥,节水保产效果最佳。
DOI:10.6041/j.issn.1000-1298.2013.03.017URL [本文引用: 1]
通过盆栽试验研究了不同水氮互作条件下水稻各项生理指标短期和长期的后效变化。结果表明:适宜的水氮组合复水短期内对生理活性有一定的补偿效应,生育后期表现出延缓衰老的长期后效性;水、氮单因子复水短期内对叶片叶绿素含量、荧光参数、丙二醛含量等生理指标影响显著,水氮协同互作效应在复水15d后达显著水平;不同的水分胁迫后复水,水氮互作效应不同,重旱处理产量随复水后施氮量的增加而增加,但产量较其他处理明显降低。应避免水稻拔节期重旱,土壤含水率不能低于田间持水率的70%;充分灌溉和轻旱处理产量随施氮量的增加有所降低,拔节期高水高肥不一定高产,轻旱后复水结合低氮追肥,节水保产效果最佳。

{kind=link}
{kind=link}