关键词:玉米; 密度; 一般配合力; 特殊配合力; AMMI模型 Combining Ability of Maize Inbred Lines from Shaan A Group and Shaan B Group under Different Density Conditions WANG Bo-Xin, WANG Ya-Hui, CHEN Peng-Fei, LIU Xu-Dong-Yu, FENG Zhi-Qian, HAO Yin-Chuan, ZHANG Ren-He, ZHANG Xing-Hua, XUE Ji-Quan* College of Agronomy, Northwest A&F University / Key Laboratory of Biology and Genetic Improvement of Maize in Arid Area of Northwest Region, Ministry of Agriculture, Yangling 712100, China Fund:The study was supported by the China Agriculture Research System (CARS-02-64) and Shaanxi Key Research and Development Project about Innovation and Utilization of Maize Elite Germplasm (2016TZC-N-2-1) AbstractThe objective of this study was to investigate the effectiveness of high-density breeding strategy through analyzing genetic components and general combining ability (GCA) effects under different planting densities. Seventeen maize inbred lines selected under high density were test-crossed with two testers (Zheng 58, Chang 7-2) in the North Carolina Design II. Thirty-four crosses were evaluated under different density conditions (45 000, 67 500, and 90 000 plants ha-1) in 2014 and 2015 at Yangling, Changwu and Yulin in Shaanxi province. The genetic variance components for grain yield and other traits were estimated using PROC VARCOMP method. Frequency distribution was used to analyze the response of GCA for grain yield and other traits under different densities. The stability of maize inbred lines and crosses was evaluated based on AMMI model. The grain yield, stalk lodging rate and rind penetration strength were mostly controlled by additive gene action, while the inheritance of sterile plant rate controlled by non-additive gene action. Also, the contribution of additive gene action to total variance increased with increasing planting density. The average of general combining ability for grain yield, sterile plant rate, stalk lodging rate and rind penetration strength increased by 0.28, -0.21, -0.03, and 0.02, respectively, from low density to high density. The correlations between GCA of the inbred lines and their crosses were significant ( r=0.877**, r=0.811**, r=0.672**) under different density. As the density increased, the number of stable maize inbred lines and crosses increased. These results indicate that increasing plant density to improve GCA for grain yield and other traits of inbred lines is an effective strategy to enhance tolerance to high plant density and yield stability.
Keyword:Maize; Density; General combining ability; Specific combining ability; AMMI model Show Figures Show Figures
表2 不同密度条件下玉米自交系产量及耐密性相关性状联合方差分析 Table 2 Mean squares from combined analysis of variance for grain yield and other traits of maize inbred lines under different densities
变异来源 Source
自由度 df
产量 Grain yield
空秆率 Sterile plant rate
倒伏率 Stalk lodging rate
茎秆强度 Rind penetration strength
45 000 plants hm-2
重复Rep
2
17.40
30.63
6777.00
1299.35
环境Env
5
1037.51* *
24.13
6871.90
4485.71*
基因型Gen
33
13.03* *
14.72* *
1055.00* *
102.56* *
一般配合力GCA
16
13.37* *
10.42
496.50* *
114.88* *
特殊配合力SCA
16
13.21* *
5.38* *
431.30*
27.68
环境× 基因型E× Gen
165
6.44* *
5.07
332.50*
30.01
环境× 一般配合力E× GCA
80
7.14* * '
14.20
174.30
73.02*
环境× 特殊配合力E× SCA
80
5.13* *
4.45
173.20
32.81
误差Error
338
1.45
6.43
227.70
37.24
67 500 plants hm-2
重复Rep
2
0.27
0.25
3565.40
595.20
环境Env
5
1272.93* *
9.54
6469.80
6268.23* *
基因型Gen
33
25.00* *
17.01* *
1150.20* *
84.45* *
一般配合力GCA
16
19.02* *
18.38* *
565.20* *
101.74* *
特殊配合力SCA
16
25.70* *
15.46* *
494.00* *
30.08
环境× 基因型E× Gen
165
10.11* *
1.59
397.20* *
31.07
环境× 一般配合力E× GCA
80
9.83* *
1.03
246.70
34.72
环境× 特殊配合力E× SCA
80
8.78* *
2.01
236.10
18.74
误差Error
338
2.93
3.91
175.70
32.32
90 000 plants hm-2
重复Rep
2
19.77
39.09
4097.60
974.81
环境Env
5
1195.23* *
15.82
3391.80
4801.34*
基因型Gen
33
19.54* *
14.88* *
1031.00* *
68.89*
一般配合力GCA
16
20.78* *
11.61* *
580.30* *
70.36* *
特殊配合力SCA
16
16.00* *
15.07* *
578.10* *
71.65
环境× 基因型E× Gen
165
11.13* *
3.47
254.50
47.37
环境× 一般配合力E× GCA
80
13.89* *
4.44
171.10
45.34
环境× 特殊配合力E× SCA
80
7.95* *
2.59
159.60
43.85
误差Error
338
4.99
6.98
203.50
45.69
* , * * Significant at the 0.05 and 0.01 probability levels, respectively. Rep: repeat; Env: environment; Gen: genotype; GCA: general combining ability; SCA: special combining ability; E× Gen: environment× genotype; E× GCA: environment× general combining ability; E× SCA: environment× special combining ability. * , * * 分别表示在0.05, 0.01水平上存在显著差异. Rep: 重复; Env: 环境; Gen: 基因型; GCA: 一般配合力; SCA: 特殊配合力; E× Gen: 环境× 基因型; E× GCA: 环境× 一般配合力; E× SCA: 环境× 特殊配合力。
表2 不同密度条件下玉米自交系产量及耐密性相关性状联合方差分析 Table 2 Mean squares from combined analysis of variance for grain yield and other traits of maize inbred lines under different densities
表3 不同密度条件下产量及耐密相关性状的一般配合力、特殊配合力方差分量及遗传力 Table 3 The variance component of general combining ability and special combining ability and heritability for grain yield and other traits under different density
方差分量 Variance component
产量 Grain yield
空秆率 Sterile plant rate
倒伏率 Stalk lodging rate
茎秆强度 Rind penetration strength
45 000 plants hm-2
一般配合力方差Vg
0.25
0.21
97.03
4.67
特殊配合力方差Vs
0.40
1.08
28.68
4.47
加性方差比值Va/Vt
55.23
28.05
87.12
67.66
遗传力H2(%)
72.48
65.56
68.49
70.73
67 500 plants hm-2
一般配合力方差Vg
0.35
0.23
103.80
4.13
特殊配合力方差Vs
0.43
1.49
28.66
2.26
加性方差比值Va/Vt
61.81
23.86
87.87
87.30
遗传力H2(%)
62.83
71.57
65.46
59.81
90 000 plants hm-2
一般配合力方差Vg
0.35
0.62
88.69
3.22
特殊配合力方差Vs
0.36
1.39
30.50
2.09
加性方差比值Va/Vt
65.77
47.20
85.33
75.52
遗传力H2(%)
58.39
76.69
75.31
31.22
Vg: general combining ability variance; Vs: special combining ability variance; Va: additive variance; Vt: total genotype variance; H2: heritability. Vg: 一般配合力方差; Vs: 特殊配合力方差; Va: 加性遗传方差; Vt: 总基因型方差; H2: 遗传力。
表3 不同密度条件下产量及耐密相关性状的一般配合力、特殊配合力方差分量及遗传力 Table 3 The variance component of general combining ability and special combining ability and heritability for grain yield and other traits under different density
图1 不同密度条件下玉米自交系产量(A)、空秆率(B)、倒伏率(C)和茎秆强度(D)一般配合力的频率分布直方图Fig. 1 Frequency distribution of general combining ability effects of grain yield (A), sterile plant rate (B), stalk lodging rate (C), and rind penetration strength (D) of maize inbred lines under different densities
图3 不同密度下玉米自交系的产量GCA与杂交组合产量的相关性Fig. 3 Correlation between general combining ability effects for grain yield of inbred lines and grain yield of their crosses under different densities
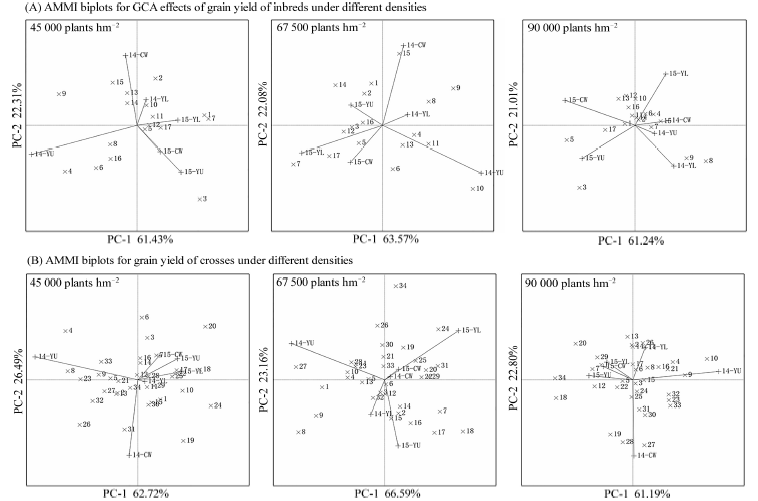
图4 不同密度条件下(A)玉米自交系的产量GCA, (B)杂交组合的产量AMMI交互作用双标图14-YL: 2014年杨凌, 14-CW: 2014年长武, 14-YU: 2014年榆林, 15-YL: 2015年杨凌, 15-CW: 2015年长武, 15-YU: 2015年榆林。Fig. 4 AMMI biplots for GCA effects for grain yield of inbred lines (A), grain yield of crosses (B) under different densities14-YL: at Yangling in 2014, 14-CW: at Changwu in 2014, 14-YU: at Yulin in 2014, 15-YL: at Yangling in 2015, 15-CW: at Changwu in 2015, 15-YU: at Yulin in 2015.
4 结论玉米杂交组合的产量与玉米自交系的一般配合力密切相关, 实施高密度选择策略对群体内基因加性效应的累积是有效的。随种植密度的增加, 加性遗传效应对产量及耐密性相关性状的贡献率呈上升趋势, 产量及耐密性相关性状的一般配合力平均数逐步朝着有利方向移动, 表现稳定的玉米自交系及测配杂交组合的数量呈上升趋势。因此, 强化逆境选择压力, 实施高密度选择策略, 是增强玉米自交系耐密性和抗倒性, 提升一般配合力, 实现产量增益的有效措施。 The authors have declared that no competing interests exist.
Duvick DN. The contribution of breeding to yield advances in maize. Adv Agron, 2005, 86: 83-145[本文引用:1]
[2]
Duvick DN. Genetic progress in yield of United States maize (Zea mays L. ). Maydica, 2005, 50: 193-200[本文引用:1]
[3]
Luque SF, Cirilo AG, Otegui ME. Genetic gains in grain yield and related physiological attributes in Argentine maize hybrids. Field Crops Res, 2006, 95: 383-397[本文引用:1]
[4]
CiX, LiM, LiangZ, XieZ, ZhangD, LiX, LuZ, RuG, BaiL, XieC, HaoZ, ZhangS. Genetic contribution to advanced yield for maize hybrids released from 1970 to 2000 in China. Crop Sci, 2011, 51: 13-20[本文引用:1]
[5]
Bruulsema T WM, TollenaarM, Heckman JR. Boosting crop yields in the next century. Better Crops Plant Food, 2000, 84: 9-13[本文引用:1]
[6]
Duvick DN. Genetic contributions to yield gains of U. S. hybrid maize, 1930-1980. In: Genetic Contributions to Yield Gains of Five Major Crop Plants. Proceedings of a Symposium. Madison, Wis. USA, 1984. pp 15-47[本文引用:1]
[7]
Duvick DN. What is yield? In: Developing Drought and LowN-tolerant Maize. Proceedings of Symposium. El Batan, Mexico: CIMMYT, 1996. pp 332-335[本文引用:1]
[8]
Troyer AF. Adaptation and heterosis in corn and mule hybrids. Crop Sci, 2006, 46: 528-543[本文引用:2]
[9]
TollenaarM, WuJ. Yield improvement in temperate maize is attributable to greater stress tolerance. Crop Sci, 1999, 39: 1597-1604[本文引用:2]
[10]
张世煌. 郑单958带给我们的创新思路和发展机遇. 玉米科学, 2006, 14(6): 4-6Zhang SH. Revelation and challenges on maize breeding strategy revealed by hybrid Zhengdan 958. J Maize Sci, 2006, 14(6): 4-6 (in Chinese with English abstract)[本文引用:2]
[11]
薛吉全, 张仁和, 马国胜, 路海东, 张兴华, 李凤艳, 郝引川, 邰书静. 种植密度、氮肥和水分胁迫对玉米产量形成的影响. 作物学报, 2010, 36: 1022-1029Xue JQ, Zhang RH, Ma GS, Lu HD, Zhang XH, Li FY, Hao YC, Tai SJ. Effects of plant density, nitrogen application, and water stress on yield formation of maize. Acta Agron Sin, 2010, 36: 1022-1029 (in Chinese with English abstract)[本文引用:1]
[12]
Reif JC, Hallauer AR, Melchinger AE. Heterosis and heterotic patterns in maize. Maydica, 2005, 50: 215-223[本文引用:1]
[13]
杨晓钦, 张仁和, 薛吉全, 邰书静, 张兴华, 郭艳萍, 郭德林. 非生物胁迫对玉米杂交种及其亲本自交系产量性状的影响. 作物学报, 2013, 39: 1325-1329Yang XQ, Zhang RH, Xue JQ, Tai SJ, Zhang XH, Guo YP, Guo DL. Effects of abiotic stress on yield traits of maize hybrids and their parental inbred lines. Acta Agron Sin, 2013, 39: 1325-1329 (in Chinese with English abstract)[本文引用:1]
[14]
Nyquist WE, Baker RJ. Estimation of heritability and prediction of selection response in plant populations. Crit Rev Plant Sci, 1991, 10: 235-322[本文引用:2]
CoxJ, Frey KJ. Combining ability and the selection of parents for specific oat mating. Crop Sci, 1984, 24: 963-967[本文引用:1]
[18]
Gauchjr HG, Zobel RW. Predictive and postdictive success of statistical analyses of yield trials. Theor Appl Genet, 1988, 76: 1-10[本文引用:1]
[19]
Goodnight CJ. Epitasis and the effect of founder events on the additive genetic variance. Evolution, 1988, 42: 441-454[本文引用:1]
[20]
Betrán FJ, BeckD, BänzigerM, Edmeades GO. Genetic analysis of inbred and hybrid grain yield under stress and non-stress environments in tropical maize. Crop Sci, 2003, 43: 807-817[本文引用:1]
[21]
Badu-AprakuB, OyekunleM, Akinwale RO. Combining ability and heterotic groups of early-maturing tropical white maize inbred lines under stress and non-stress environments. Agron J, 2011, 103: 544-557[本文引用:1]
[22]
梁文科, 张世煌, 戚廷香, 庹洪章, 刘永忠, 郑用琏, 徐尚忠. 热带温带玉米群体产量性状遗传力及遗传方差分量的剖析. 中国农业科学, 2006, 39: 2178-2185Liang WK, Zhang SH, Qi TX, Tuo HZ, Liu YZ, Zheng YL, Xu SZ. Dissection of heritability and genetic variance components for yield traits in tropical and temperate maize populations. Sci Agric Sin, 2006, 39: 2178-2185 (in Chinese with English abstract)[本文引用:1]
[23]
王文斌, 徐淑兔, 高杰, 张兴华, 郭东伟, 李向阳, 薛吉全. 基于SNP标记的玉米自交系遗传多样性分析. 玉米科学, 2015, 23(2): 41-45Wang WB, Xu ST, GaoJ, Zhang XH, Guo DW, Li XY, Xue JQ. Analysis of genetic diversity of maize inbred lines based on SNP markers. J Maize Sci, 2015, 23(2): 41-45 (in Chinese with English abstract)[本文引用:1]
[24]
杨泽勇, 张兴华, 薛吉全. 3种密度玉米自交系选择的配合力分析. 湖北农业科学, 2009, 48: 576-579Yang ZY, Zhang XH, Xue JQ. Study on combining ability of maize inbred lines selected in three densities. Hubei Agric Sci, 2009, 48: 576-579 (in Chinese with English abstract)[本文引用:1]
[25]
高瑞景, 李泾孝, 张仁和, 毛建昌. 两种密度系谱法选择玉米自交系的一般配合力分析. 西北农林科技大学学报(自然科学版), 2008, 36(6): 56-60Gao RJ, Li JX, Zhang RH, Mao JC. Selection of maize inbred lines with pedigree under two densities-based on the general combination ability analysis. J Southwest A&F Univ (Nat Sci Edn), 2008, 36(6): 56-60 (in Chinese with English abstract)[本文引用:1]
[26]
梁雨娟, 刘振库, 李继竹, 尹日成, 王远路, 姜龙, 杨伟光. 不同密度条件下玉米杂种优势及配合力研究. 玉米科学, 2012, 20(6): 25-29Liang YJ, Liu ZK, Li JZ, Yin RC, Wang YL, JiangL, Yang WG. Heterosis and combining ability of maize under different density. J Maize Sci, 2012, 20(6): 25-29 (in Chinese with English abstract)[本文引用:1]
[27]
张铭堂, 张国良, 才卓. 玉米自交系选育的理论基础与实践经验. 玉米科学, 2010, 18(2): 1-4Zhang MT, Zhang GL, CaiZ. Theoretical foundation and practice experience of breeding for maize inbred lines. J Maize Sci, 2010, 18(2): 1-4 (in Chinese with English abstract)[本文引用:1]
[28]
Francis TR, Kannenberg LW. Yield stability studies in short-season maize: I. A descriptive method for grouping genotypes. Can J Plant Sci, 1978, 58: 1029-1034[本文引用:1]
[29]
Matteo JA, Ferreyra JM, Cerrudo AA, EcharteL, Andrade FH. Yield potential and yield stability of Argentine maize hybrids over 45 years of breeding. Field Crops Res, 2016, 197: 107-116[本文引用:1]
[30]
Dow EW, Daynard TB, Muldoon JF, Major DJ, Thurtell GW. Resistance to drought and density stress in Canadian and European maize (Zea mays L. ) hybrids. Can J Plant Sci, 1984, 64: 575-585[本文引用:1]
 , 王亚辉, 陈朋飞, 刘徐冬雨, 冯志前, 郝引川, 张仁和, 张兴华, 薛吉全
, 王亚辉, 陈朋飞, 刘徐冬雨, 冯志前, 郝引川, 张仁和, 张兴华, 薛吉全




{kind=link}
{kind=link}
{kind=link}
{kind=link}