关键词:普通小麦; 芒长; QTL; SSR标记; DArT标记 Mapping QTLs for Awn Length in Recombinant Inbred Line Population Derived from the Cross between Common Wheat and Tibetan Semi-wild Wheat GONG Xi1, JIANG Yun-Feng2, XU Bin-Jie2, QIAO Yuan-Yuan2, HUA Shi-Yu1, WU Wang1, MA Jian2, ZHOU Xiao-Hong2, QI Peng-Fei2,*, LAN Xiu-Jin2 1College of Agronomy, Sichuan Agricultural University, Chengdu 611130, China
2Triticeae Research Institute, Sichuan Agricultural University, Chengdu, 611130, China
Fund:This study was supported by the National Natural Science Foundation of China (31570335). AbstractAwn length is an important agronomic trait in hexaploid wheat and controlled by multiple genes. A recombinant inbred line (RIL) population containing 186 lines were developed by crossing Tibetan semi-wild wheat accession Q1028 (female) and common wheat variety Zhengmai 9023, and a genome-wide genetic map (2597 cM) was constructed using SSR and DArT markers. QTLs controlling awn length were identified with two-year phenotypic data and the genetic map by using the Inclusive Composite Interval Mapping (ICIM) method. Two QTLs associated with awn length, designated Qwa.sau-4AS and Qwa.sau-5AL, were detected on chromosomes 4A and 5A, which explained 7.4% and 27.3% of phenotypic variations, respectively. According to genetic locations, effects of the two QTLs might be from Hdand B1 genes that could reduce awn length, respectively. Genetic analysis showed that Qwa.sau-4ASand Qwa.sau-5ALhad cumulative effects, with stronger inhibiting effect in Qwa.sau-5AL than in Qwa.sau-4AS. These results are valuable for fine mapping and cloning target genes controlling awn length in the future.
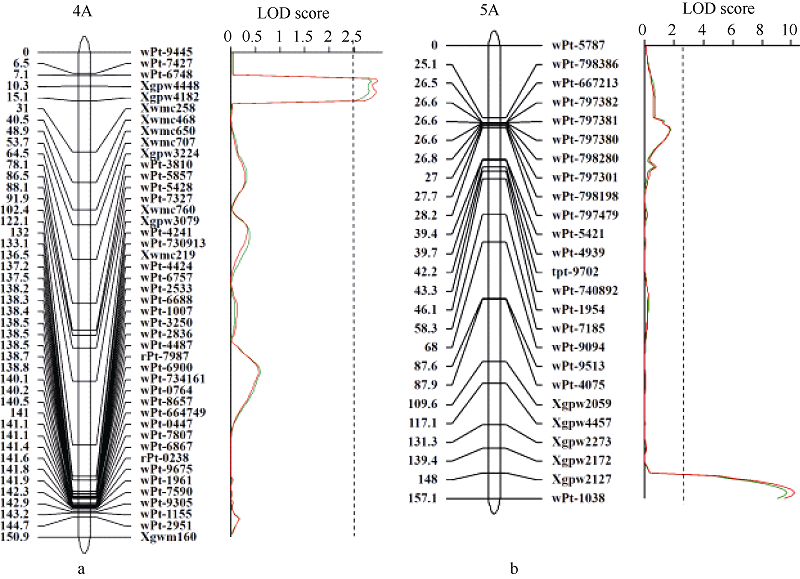
图3 4A染色体(a)和5A染色体(b)遗传图谱及芒长性状分析的LOD曲线图 连锁图谱右边是标记名称, 左边是标记位置(cM)。红色和绿色曲线分别为2012和2013年数据; 虚线指示LOD阈值(LOD = 2.5)。Fig. 3 Genetic maps of chromosomes 4A (a) and 5A (b) and the LOD plots of awn length In the linkage map, marker names and positions (cM) are on the right and left, respectively. LOD scores in 2012 and 2013 are shown with the red and green curves, respectivley. The vertical dashed line is the threshold of significance (LOD = 2.5).
表1 Table 1 表1(Table 1)
表1 郑麦9023 × Q1028群体的芒长QTL Table 1 QTLs for awn length identified in the population of ZM9023 × Q1028
QTL
染色体 Chromosome
标记区间 Marker interval
位置 Position (cM)
年份 Year
LOD
表型贡献率 PVE (%)
加性效应 Additive effect
Qawn.sau-4A
4AS
wPt6748-Xgpw4448
9
2012
2.98
7.5220
-0.5287
9
2013
2.85
7.2147
-0.5289
Qawn.sau-5A
5AL
Xgpw2127-wPt_1038
155
2012
10.23
27.9576
-1.0011
155
2013
9.67
26.6541
-0.9985
A negative additive effect indicates that the QTL shortens awn length. PVE: phenotypic variation explained. 加性效应负数表示该QTL缩短芒长。
表1 郑麦9023 × Q1028群体的芒长QTL Table 1 QTLs for awn length identified in the population of ZM9023 × Q1028
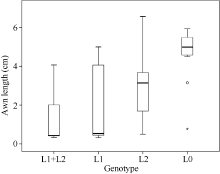
图4 RIL群体不同基因型的芒长箱型分布图Fig. 4 Box plot distribution of awn length in different genotypes of the RIL population L1: Qwa.sau-5AL; L2: Qwa.sau-4AS ; L0: without QTL.
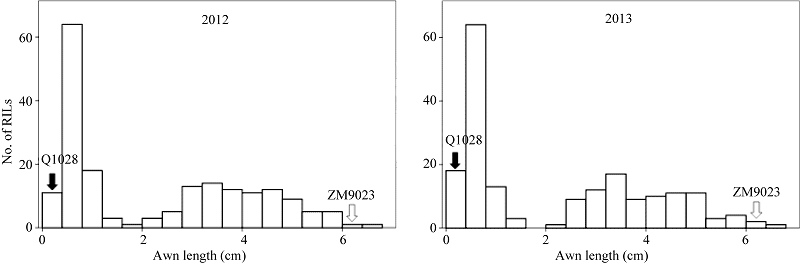
3 讨论本研究基于郑麦9023和Q1028构建的RIL群体及其全基因组遗传图谱, 共检测到2个与芒长相关的QTL。其中Qwa.sau-4AS被定位于染色体4AS上的Xgpw4448附近, 与Sourdille等[15]的定位结果相比较, Xgpw4448与Xfba78之间的遗传距离接近, Qwa.sau-4AS应与之前报道的Hd基因位于同一区域, 因此推测该QTL效应来源于钩芒基因Hd。Qwa.sau-5AL与wpt-1038连锁, 与杜斌等[9]的定位结果相比较, wpt-1038与Xgwm291都位于5AL的末端区域, 与B1基因在同一区域, 并表现出同样强烈的芒抑制作用, 因此该QTL的效应应该来源于强抑芒基因B1。 2个抑芒位点均来源于无芒亲本Q1028, 对芒长具有显著抑制, 并存在累加效应。通过比较不同基因型株系的芒长数据发现, 只携带Hd基因的株系平均芒长为2.97 cm, 而只携带B1基因的株系平均芒长为1.71 cm, 说明B1对芒长的抑制效果强于Hd基因。而当同一株系上同时存在这2个基因时, 平均芒长最小。由此可见, B1基因与Hd基因对芒长的抑制效果具有累加效应。这2个位点累计可解释33.9%~35.5%的表型变异, 推测存在未检测出的QTL。 无芒小麦中国春作为一个普通小麦的模式植物, 对其芒长性状已经有较深入研究。普遍认为其无芒特性是由Hd、B1和B2联合作用所致[15]。本试验所采用的无芒亲本为西藏半野生小麦, 许多系统分类及聚类研究已经证明其与中国春具有密切的亲缘关系[22, 23, 24]。未被检测的QTL效应可能源于6BL染色体的B2, 我们采用连锁图谱的6B染色体包含着与B2基因连锁的标记Xwmc182, 但该位点及其所在区域与芒长无显著连锁, 因此未检测到预期的QTL。其原因可能是所采用的2个亲本之间B2基因型相同。 本研究结果为Hd与B1基因后续精细定位与克隆提供了分子标记基础。我们已经利用与钩芒基因Hd连锁的分子标记, 从F8的自交系群体中筛选到杂合单株构建近等基因系群体, 正在进行精细定位相关工作。 4 结论共定位2个与芒长性状相关的QTL, 分别位于4AS和5AL染色体, 共解释34.7%的表型变异。这2个QTL应是钩芒基因Hd和抑芒基因B1。2个基因对芒长的抑制作用具有累加效应, 且B1的抑制效果强于Hd。本研究为后续筛选近等基因系及基因精细定位奠定了基础。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Biscoe PV, Littleton EJ, Scott RK. Stomatal control of gas exchange in barley awns. Ann Appl Biol, 1973, 75: 285-297[本文引用:1]
[2]
Blum A. Photosynthesis and transpiration in leaves and ears of wheat and barley varieties. J Exp Bot, 1985, 36: 432-440[本文引用:1]
[3]
黄瑾, 骆惠生, 张勃, 贾秋珍, 金明安, 曹世勤, 金社林. 普通小麦芒的遗传分析. 甘肃农业科技, 2011, (2): 11-12HuangJ, Luo HS, ZhangB, Jia QZ, Jin MA, Cao SQ, Jin SL. Genetic analysis of mount of common wheat. Gansu Agric Sci Tech, 2011, (2): 11-12 (in Chinese with English abstract)[本文引用:1]
[4]
巴青松, 傅兆麟, 白凡杰. 小麦芒的研究. 淮北煤炭师范学院学报(自然科学版), 2010, 31(1): 29-33Ba QS, Fu ZL, Bai FJ. The research of wheat awns. J Huaibei Coal Ind Teachers Coll (Nat Sci Edn), 2010, 31(1): 29-33 (in Chinese with English abstract)[本文引用:1]
[5]
Maydup ML, AntoniettaM, GracianoC, Guiamet JJ, Tambussi EA. The contribution of the awns of bread wheat (Triticum aestivum L. ) to grain filling: responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crops Res, 2014, 167: 102-111[本文引用:1]
[6]
王忠, 顾蕴洁, 高煜珠. 麦芒的结构及其光合特性. 植物学报, 1993, 35: 921-928WangZ, Gu YJ, Gao YZ. Structure and photosynthetic characteristics of awns of wheat and barley. Acta Bot Sin, 1993, 35: 921-928 (in Chinese with English abstract)[本文引用:1]
[7]
Li XJ, Wang HG, Li HB, Zhang LY, Teng NJ, Lin QQ, WangJ, Kuang TY, Li ZS, LiB, Zhang AM, Lin JX. Awns play a dominant role in carbohydrate production during the grain-filling stages in wheat (Triticum aestivum). Physiol Plant, 2006, 127: 701-709[本文引用:1]
[8]
Rebetzke GJ, Bonnett DG, Reynolds MP. Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J Exp Bot, 2016, 67: 2573-2586[本文引用:1]
[9]
杜斌, 崔法, 王洪刚, 李兴锋. 小麦芒长抑制基因B1近等基因系的鉴定及遗传分析. 分子植物育种, 2010, 8: 259-264DuB, CuiF, Wang HG, Li XF. Characterization and genetic analysis of near-isogenic lines for awn-inhibitor gene B1 of common wheat. Mol Plant Breed, 2010, 8: 259-264 (in Chinese with English abstract)[本文引用:4]
[10]
ElbaumR, ZaltzmanL, BurgertI, FratzlP. The Role of wheat awns in the seed dispersal unit. Science, 2007, 316: 884-886[本文引用:1]
[11]
姚国新, 张强, 吴建涛, 胡广隆, 李自超. 利用近等基因系对水稻芒基因AWN3-1的遗传定位. 中国农业大学学报, 2010, 15(5): 1-5Yao GX, ZhangQ, Wu JT, Hu GL, Li ZC. Mapping awn gene AWN3-1 with near-isogenic line of rice. J China Agric Univ, 2010, 15(5): 1-5 (in Chinese with English abstract)[本文引用:1]
[12]
KosugeK, WatanabeN, KuboyamaT, Melnik VM, Yanchenko VI, Rosova MA, Goncharov NP. Cytological and microsatellite mapping of mutant genes for spherical grain and compact spikes in durum wheat. Euphytica, 2008, 159: 289-296[本文引用:1]
[13]
LiH, Han YC, Guo XX, XueF, Wang CY, Ji WQ. Genetic effect of locus B2 inhibiting awning in double-ditelosomic 6B of Triticum durum DR147. Genet Resour Crop Evol, 2015, 62: 407-418[本文引用:1]
[14]
金善宝. 中国小麦学. 北京: 中国农业出版社, 1996. p 13Jin SB. Wheat Science in China. Beijing: China Agriculture Press, 1996. p 13 (in Chinese)[本文引用:1]
[15]
SourdilleP, CadalenT, GayG, GillB, BernardM. Molecular and physical mapping of genes affecting awning in wheat. Plant Breed, 2002, 121: 320-324[本文引用:3]
[16]
Röder MS, KorzunV, WendehakeK, PlaschkeJ, Tixier MH, LeroyP, Ganal MW. A microsatellite map of wheat. Genetics, 1998, 149: 2007-2023[本文引用:1]
[17]
Somers DJ, IsaacP, EdwardsK. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 1105-1114[本文引用:]
[18]
WenzlP, CarlingJ, KudrnaD, JaccoudD, HuttnerE, KleinhofsA, KilianA. Diversity arrays technology (DArT) for whole-genome profiling of barley. Proc Natl Acad Sci USA, 2004, 101: 9915-9920[本文引用:1]
[19]
Jiang YF, Lan XJ, LuoW, Kong XC, Qi PF, Wang JR, Wei YM, Jiang QT, Liu YX, Peng YY, Chen GY, Dai SF, Zheng YL. Genome-wide quantitative trait locus mapping identifies multiple major loci for brittle rachis and threshability in Tibetan semi-wild wheat (Triticum aestivum ssp. tibetanum Shao). PloS One, 2014, 9: e114066[本文引用:1]
[20]
MengL, LiH, ZhangL, WangJ. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J, 2015, 3: 269-283[本文引用:1]
[21]
Li HH, Ye GY, Wang JK. A modified algorithm for the Improvement of Composite Interval Mapping. Genetics, 2007, 175: 361-374[本文引用:1]
[22]
Wang YJ, Wang CY, ZhangH, Yue ZN, Liu XL, Ji WQ. Genetic analysis of wheat (Triticum aestivum L. ) and related species with SSR markers. Genet Resour Crop Evol, 2013, 60: 1105-1117[本文引用:1]
[23]
Wang HY, Wang XE, Liu DJ. Assessment of genetic diversity of Yunnan, Tibetan, and Xinjiang wheat using SSR markers. J Genet Genomics, 2007, 34: 623-633[本文引用:1]
[24]
KosugeK, WatanabeN, KuboyamaT, Melnik VM, Yanchenko VI, Rosova MA, Goncharov NP. Comparison of the genetic diversity between Triticum aestivum ssp. tibetanum Shao and Tibetan wheat land races (Triticum aestivum L. ) by using intron-splice junction primers. Euphytica, 2008, 159: 289-296[本文引用:1]
 , 蒋云峰
, 蒋云峰





{kind=link}
{kind=link}
{kind=link}
{kind=link}