关键词:甘薯; 单株有效薯块数; 根系特性; 糖组分; 产量 Comparison of Root Characteristics and Sugar Components in Root and Leaf at Early Growth Phase of Sweet Potato Varieties with Significant Difference in Valid Storage Root Number WANG Cui-Juan1, SHI Chun-Yu1,*, LIU Na2, LIU Shuang-Rong1, YU Xin-Di1 1College of Agronomic Sciences, Shandong Agricultural University / State Key Laboratory of Crop Biology, Tai’an 271018, China
2 Resources of Horticulture Science and Engineering, Shandong Agricultural University / State Key Laboratory of Crop Biology, Tai’an 271018, China
Fund:This study was supported by the National Natural Science Foundation of China (31371577) and the Potato Innovation Program for Chief Expert of Shandong Province (SDAIT-10-011-01) AbstractStarch sweet potato varieties Shangshu 19 and Jixu 23 differing in valid storage root number per plant significantly were used to investigate root characteristics, sugar components in root and leaf at early growth phase and their relationship with the formation of storage root per plant at top cover stage. The results showed that, Shangshu 19 with higher valid storage root number regrew quickly with more new roots after seedling transplanting, developed fiberous roots mainly in the whole early growth phase, and formed a stable rate of top biomass/total root system biomass (T/TR) at 15 and 30 days after planting. Jixu 23 achieved the steady number of adventitious root and valid storage root at 15 and 30 days after planting, respectively. Meanwhile, Jixu 23 showed the lower rate of top biomass/storage root biomass (T/SR) at top cover stage (45 days after planting). On the other hand, in point of metabolism of sucrose and hexoses, and the formation of stored polysaccharide polymers, Shangshu 19 had significantly lower rate of sucrose/hexoses in the whole early growth phase, formed the greater sucrose concentration gradient between leaves and roots, and had 1-kestose and nystose in roots at 15 and 30 days after planting. Its starch content in leaf was significantly lower than that of Jixu 23 at 30 and 45 days after planting. Meanwhile, Jixu 23 only had Nystose in roots before storage root formation, with significantly lowest rate of sucrose/starch when the rate of sucrose/total soluble sugar was similar to that of Shangshu 19 at 45 days after planting. A two-year field trials (2013-2014) were performed to investigate yield-contributing traits and the fresh storage root yield at top cover stage and harvest period, in which, Shangshu19 showed the more valid storage roots per plant, higher valid storage root fresh weight per plant or storage root at top cover stage and harvest period, meanwhile, Jixu23 had significantly higher average fresh weight per storage root.
Keyword:Sweet potato; Valid storage root per plant; Root characteristics; Sugar components; Storage root yield Show Figures Show Figures
表1 封垄期和收获期与产量有关的性状(大田, 2013-2014) Table 1 Yield traits and fresh yield of storage root in field trials at top cover stage and harvest period in 2013-2014
年份 Year
生育时期 Growth stage
品种 Variety
单株有效薯块数 VSRNP
平均单薯鲜重 AFWP (g)
单株有效薯块鲜重 VSRFWP (g)
块根产量 FYSR (kg hm-2)
2013
45 d
商薯19 Shangshu 19
5.67 a
10.46 b
38.58 a
—
济徐23 Jixu 23
3.00 b
16.02 a
31.29 b
—
收获Harvest
商薯19 Shangshu 19
5.00 a
168.13 b
840.63 a
42031.25 a
济徐23 Jixu 23
3.00 b
206.77 a
620.31 b
31015.63 b
2014
45 d
商薯19 Shangshu 19
7.33 a
3.20 b
23.46 a
—
济徐23 Jixu 23
5.00 b
3.97 a
19.83 b
—
收获Harvest
商薯19 Shangshu 19
6.67 a
111.04 b
740.63 a
37031.25 a
济徐23 Jixu 23
4.33 b
127.02 a
550.00 b
27500.00 b
Values in each column followed by different letters are significantly different at the 0.05 probability level. VSRNP: valid storage root number per plant; AFWP: average fresh weight per tube; VSRFWP: valid storage root fresh weight per plant; FYSR: fresh yield of storage root. 同一列中不同小写字母表示数值在0.05的水平上差异显著。
表1 封垄期和收获期与产量有关的性状(大田, 2013-2014) Table 1 Yield traits and fresh yield of storage root in field trials at top cover stage and harvest period in 2013-2014
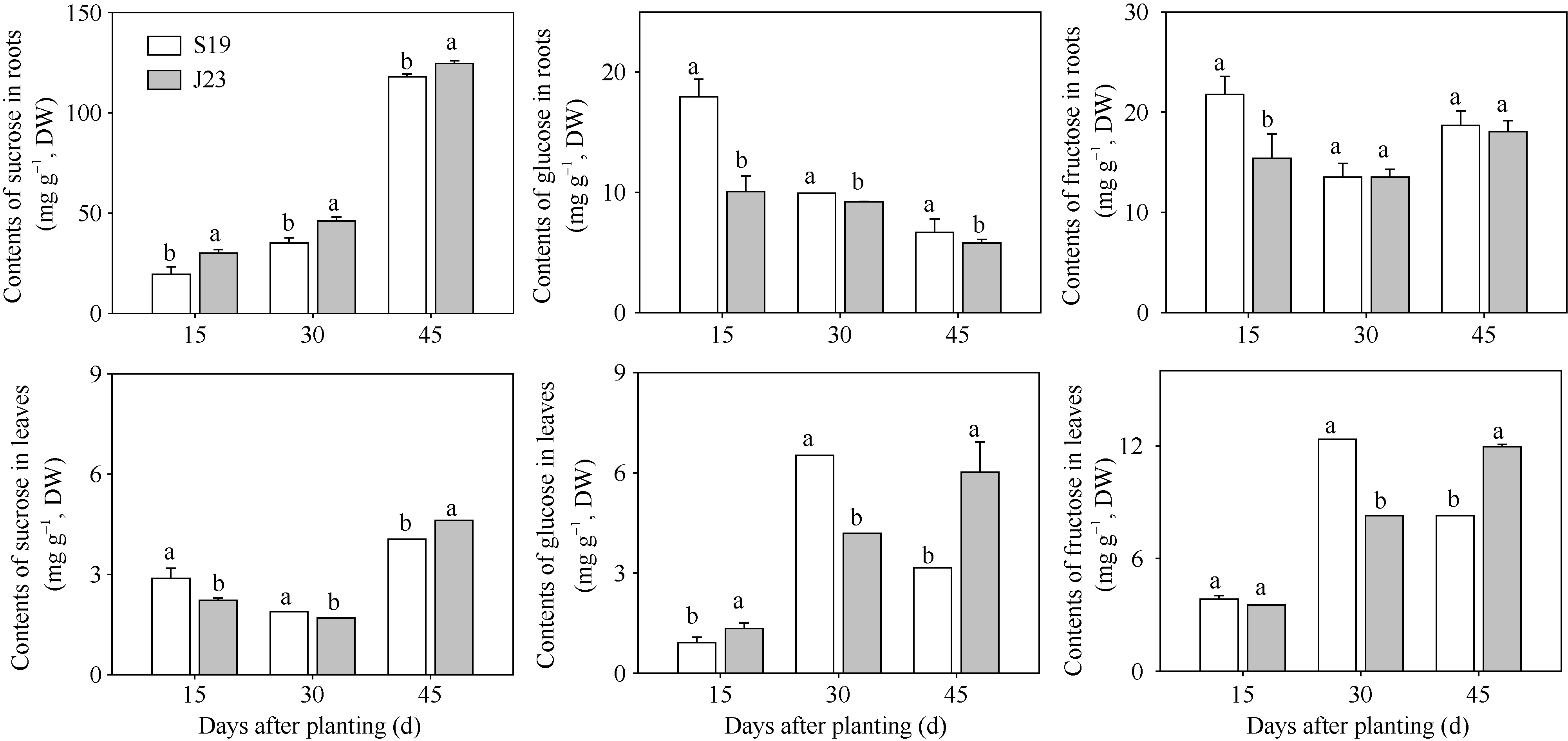
图1 秧苗栽后15、30和45 d甘薯根系和叶片蔗糖、葡萄糖、果糖的含量变化(盆栽, 2014) S19: 商薯19; J23: 济徐23。图柱上标以不同小写字母表示糖含量在0.05水平上差异显著。Fig. 1 Changes of contents of sucrose, glucose and fructose at 15, 30, and 45 days after planting in root and leaf in pot trials in 2014 S19: Shangshu 19; J23: Jixu 23. Bars superscripted by different letters are significantly different at the 0.05 probability level among sugar contents.
图2 秧苗栽后15、30和45 d甘薯根系、叶片淀粉和可溶性总糖的含量变化(盆栽, 2014) S19: 商薯19; J23: 济徐23。图柱上标以不同小写字母表示糖含量在0.05水平上差异显著。Fig. 2 Changes of contents of starch and total soluble sugar in roots and leaves at 15, 30, and 45 days after planting in pot trials in 2014 S19: Shangshu 19; J23: Jixu 23. Bars superscripted by different letters are significantly different at the 0.05 probability level among sugar contents.
表5 秧苗栽后15、30和45 d根系的糖组分比率(盆栽, 2014) Table 5 Changes of content ratio of carbohydrate composition in root at 15, 30, and 45 days after planting in pot trials (2014)
生育时期 Growth stage
品种 Variety
蔗糖/己糖 Sucrose/hexoses
蔗糖/可溶性总糖 Sucrose/total soluble sugar
蔗糖/淀粉 Sucrose/starch
15 d
商薯19 Shangshu 19
0.488± 0.036 b
0.205± 0.033 b
0.201± 0.031 b
济徐23 Jixu 23
1.190± 0.064 a
0.355± 0.024 a
0.249± 0.006 a
30 d
商薯19 Shangshu 19
1.516± 0.104 b
0.434± 0.028 b
0.194± 0.015 b
济徐23 Jixu 23
2.030± 0.066 a
0.527± 0.029 a
0.241± 0.031 a
45 d
商薯19 Shangshu 19
4.539± 0.355 b
0.767± 0.042 a
0.322± 0.016 a
济徐23 Jixu 23
5.231± 0.175 a
0.827± 0.005 a
0.270± 0.009 b
己糖=葡萄糖+果糖。Hexose = glucose +fructose.
表5 秧苗栽后15、30和45 d根系的糖组分比率(盆栽, 2014) Table 5 Changes of content ratio of carbohydrate composition in root at 15, 30, and 45 days after planting in pot trials (2014)
梁康迳, 王雪仁, 林文雄, 陈志雄, 李亚娟. 水稻产量形成的生理生态研究进展. 中国生态农业学报, 2002, 10(3): 63-65Liang KJ, Wang XR, Lin WX, Chen ZX, Li YJ. Advancement in physiological studies on yield formation in rice. Chin J Eco-Agric, 2002, 10(3): 63-65 (in Chinese with English abstract)[本文引用:1]
[2]
李朝霞, 赵世杰, 孟庆伟, 邹琦. 高粒叶比小麦群体生理基础研究进展. 麦类作物学报, 2002, 22(4): 79-83Li ZX, Zhao SJ, Meng QW, ZouQ. Advances in the study on physiological base of wheat population with high grain leaf area ratio. J Triticeae Crops, 2002, 22(4): 79-83 (in Chinese with English abstract)[本文引用:1]
[3]
史春余, 王振林, 赵秉强, 郭风法, 余松烈. 钾营养对甘薯某些生理特性和产量形成的影响. 植物营养与肥料学报, 2002, 8: 81-85Shi CY, Wang ZZ, Zhao BQ, Guo FF, Yu SL. Effect of potassium nutrition on some physiological characteristics and yield formation of sweet potato. Plant Nutr Fert Sci, 2002, 8: 81-85 (in Chinese with English abstract)[本文引用:1]
[4]
陈晓光, 李洪民, 张爱君, 史新敏, 唐忠厚, 魏猛, 史春余. 不同氮水平下多效唑对食用型甘薯光合和淀粉积累的影响. 作物学报, 2012, 38: 1728-1733Chen XG, Li HM, Zhang AJ, Shi XM, Tang ZH, WeiM, Shi CY. Effect of paclobutrazol under different N-application rates on photosynthesis and starch accumulation in edible sweet potato. Acta Agron Sin, 2012, 38: 1728-1733 (in Chinese with English abstract)[本文引用:1]
[5]
马代夫, 朱崇文. 甘薯壮苗增产的生理特点分析. 作物杂志, 1989, (4): 22-23Ma DF, Zhu CW. Physiological basis of sweet potato strong seedling on high tuberous root yield. Crops, 1989, 4: 22-23 (in Chinese)[本文引用:1]
[6]
周全卢. 秋甘薯不同类型品种干物质积累特性研究. 西南大学硕士学位论文, 重庆, 2007. p 52Zhou QL. Research of Dry Matter Accumulating Characters on Autumn Sweet Potato Varieties of Different Types. MS Thesis of Southwest University, Chongqing, China, 2007. p 52 (in Chinese with English abstract)[本文引用:3]
[7]
王翠娟, 史春余, 王振振, 柴沙沙, 史衍玺. 覆膜栽培对甘薯幼根生长发育、块根形成及产量的影响, 作物学报, 2014, 40: 1677-1685Wang CJ, Shi CY, Wang ZZ, Chai SS, Shi YX. Effects of plastic film mulching cultivation on young roots growth development, tuber formation and tuber yield of sweet potato. Acta Agron Sin, 2014, 40: 1677-1685 (in Chinese with English abstract)[本文引用:1]
[8]
陆漱韵, 刘庆昌, 李惟基. 甘薯育种学. 北京: 中国农业出版社, 1998. p 211Lu SY, Liu QC, Li W J. Sweet Potato Breeding. Beijing: China Agriculture Press, 1998. p 211 (in Chinese)[本文引用:1]
[9]
Villordon AQ, La Bonte D R, Firon N, Kfir Y, Pressman E, Schwartz A. Characterization of adventitious root development in sweet potato. Hort Sci, 2009, 44: 651-655[本文引用:4]
[10]
Dubrovsky JG, Forde BG. Quantitative analysis of lateral root development: pitfalls and how to avoid them. Plant Cell, 2012, 24: 4-14[本文引用:2]
[11]
VillordonA, LaBonte D, Solis J, Firon N. Characterization of lateral root development at the onset of storage root initiation in ‘Beauregard’ sweet potato adventitious roots. Hort Sci, 2012, 47: 961-968[本文引用:4]
[12]
Villordon AQ, Clark CA. Variation in virus symptom development and root architecture attributes at the onset of storage root initiation in ‘Beauregard’ sweet potato plants grown with or without nitrogen. PloS One, 2014, 9: e107384[本文引用:3]
[13]
潘庆民, 韩兴国, 白永飞, 杨景成. 植物非结构性贮藏碳水化合物的生理生态学研究进展, 植物学通报, 2002, (1): 30-38Pan QM, Han XG, Bai YF, Yang JC. Advances in physiology and ecology studies on stored non-structure carbohydrates in plants. Chin Bull Bot, 2002, (1): 30-38 (in Chinese with English abstract)[本文引用:1]
[14]
KageH, KochlerM, StutzelH. Root growth and dry matter partitioning of cauliflower under drought stress conditions: measurement and simulation. Eur J Agron, 2004, 20: 379-394[本文引用:1]
刘颖慧, 贾海坤, 高琼. 植物同化物分配及其模型研究综述, 生态学报, 2006, 26: 1981-1992Liu YH, Jia HK, GaoQ. Review on researches of photoassimilates partitioning and its models. Acta Ecol Sin, 2006, 26: 1981-1992 (in Chinese with English abstract)[本文引用:1]
[17]
Noh SA, Lee HS, Kim YS, Paek KH, Shin JS, Bae JM. Down-regulation of the IbEXP1 gene enhanced storage root development in sweet potato. J Exp Bot, 2013, 64: 129-142[本文引用:1]
[18]
TanakaM, KatoN, NakayamaH, NakataniM, TakahataY. Expression of class 1Knotted1-like homeobox genes in the storage roots of sweet potato (Ipomoea batatas). J Plant Physiol, 2008, 165: 1726-1735[本文引用:1]
[19]
RaviV, IndiraP. Crop physiology of sweet potato. In: Janick J ed. Horticultural reviews. New York: John Wiley & Sons, Inc, 1999, Vol. 23, pp 277-339[本文引用:1]
[20]
Wilson LA, Low SB. The anatomy of the root system in West Indian sweet potato cultivars. Ann Bot, 1973, 37: 633-643[本文引用:1]
[21]
BelehuT, Hammes P S, Robbertse P J. The origin and structure of adventitious roots in sweet potato (Ipomoea batatas). Aust J Bot, 2004, 52: 551-558[本文引用:1]
Kays SJ. The physiology of yield in the sweet potato. A Natu Reso Trop, 1985, 1: 79-132[本文引用:1]
[24]
陆漱韵, 刘庆昌, 李惟基. 甘薯育种学. 北京: 中国农业出版社, 1998. p 58Lu SY, Liu QC, Li W J. Sweet Potato Breeding. Beijing: China Agriculture Press, 1998. p 58 (in Chinese)[本文引用:1]
[25]
GaltierN, Foyer CH, HuberJ, Voelker TA, Huber SC. Effects of elevated sucrose-phosphate synthase activity on photosynthesis, assimilate partitioning, and growth in tomato. Plant Physiol, 1993, 101: 535-543[本文引用:1]
[26]
FarrarJ, PollockC, GallagherJ. Sucrose and the integration of metabolism in vascular plants. Plant Sci, 2000, 154: 1-11[本文引用:1]
[27]
RoitschT, González MC. Function and regulation of plant invertases: sweet sensations. Trends Plant Sci, 2004, 9: 606-610[本文引用:1]
[28]
Hendriks JH, KolbeA, GibonY, StittM, GeigenbergerP. ADP-glucose pyrophosphorylase is activated by posttranslational redoxmodification in response to light and to sugars in leaves of Arabidopsis and other plant species. Plant Physiol, 2003, 133: 838-849[本文引用:1]
[29]
RonteinD, Dieuaide-NoubhaniM, Dufourc EJ, RaymondP, RolinD. The metabolic architecture of plant cells. Stability of central metabolism and flexibility of anabolic pathways during the growth cycle of tomato cells. J Biol Chem, 2002, 277: 43948-43960[本文引用:1]
[30]
LalondeS, WipfD, Frommer WB. Transport mechanisms for organic forms of carbon and nitrogen between source and sink. Annu Rev Plant Biol, 2004, 55: 341-372[本文引用:1]
[31]
TurgeonR. The role of phloem loading reconsidered. Plant Physiol, 2010, 152: 1817-1823[本文引用:1]
[32]
潘秋红, 张大鹏. 植物转化酶的种类﹑特性与功能. 植物生理学通讯, 2004, 40: 275-280Pan QH, Zhang DP. Isoforms, characteristics and roles of plant invertases. Plant Physiol J, 2004, 40: 275-280 (in Chinese)[本文引用:2]
[33]
Schiefelbein JW, Benfey PN. The development of plant roots: new approaches to underground problems. Plant Cell, 1991, 3: 1147[本文引用:1]
[34]
Tang GQ, LusvherM, SturmA. Antisense repression of vacular and cell wall invertase in transgenic carrot alters early plant development and partitioning. Plant Cell, 1999, 11: 177-189[本文引用:1]
[35]
王玮, 龚义勤, 柳李旺, 王燕, 荆赞革, 黄丹琼, 汪隆植. 萝卜肉质根膨大过程中糖含量及蔗糖代谢相关酶活性分析. 园艺学报, 2007, 34: 1313-1316Wang W, Gong Y Q, Liu L W, Wang Y, Jing Z G, Huang D Q; Wang LZ. Changes of sugar content and sucrose metabolizing enzyme activities during fleshy tap root development in radish. Acta Hort Sin, 2007, 34: 1313-1316 (in Chinese with English abstract)[本文引用:1]
[36]
姜立娜. 萝卜肉质根形成性状的分子生物学基础. 南京农业大学博士学位论文, 江苏南京, 2012. p 24Jiang LN. Molecular Characterization of Taproot Formation Traits in Radish. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2012. p 24 (in Chinese with English abstract)[本文引用:1]
[37]
WeberH, BorisjukL, HeimU, SauerN, WobusU. A role for sugar transporters during seed development molecular characterization of a hexose and a sucrose carrier in fava bean seeds. Plant Cell, 1997, 9: 895-908[本文引用:1]
[38]
谢鸣, 陈俊伟, 程建徽, 秦巧平, 蒋桂华, 王力宏, 王允镔, 戚行江. 杨梅果实发育与糖的积累及其关系研究. 果树学报, 2005, 6: 38-42XieM, Chen JW, Cheng JH, Qin QP, Jiang GZ, Wang LH, Wang YB, Qi XJ. Studies on the fruit development and its relationship with sugar accumulation in bayberry fruit. J Fruit Sci, 2005, 6: 38-42 (in Chinese with English abstract)[本文引用:1]
[39]
KatoT. Change of sucrose synthase activity in developing endosperm of rice cultivars. Crop Sci, 1995, 35: 827-831[本文引用:1]
[40]
HeinekeD, SonnewaldU, BüssisD, GünterG, LeidreiterK, WilkeI, Heldt HW. Apoplastic expression of yeast-derived invertase in potato. Plant Physiol, 1992, 100: 301-308[本文引用:1]
[41]
WeschkeW, PanitzR, GubatzS, WangQ, RadchukR, WeberH, WobusU. The role of invertases and hexose transporters incontrolling sugar ratios in maternal and filial tissues of barley caryopses during early development. Plant J, 2003, 33: 395-411[本文引用:1]
[42]
WeberH, BorisjukL, HeimU, BuchnerP, WobusU. Seed coat-associated invertases of fava bean control both unloading and storage functions: cloning of cDNAs and cell type-specific expression. Plant Cell, 1995, 7: 1835-1846[本文引用:1]
[43]
Wang SJ, Chen MH, Yeh KW, Tsai CY. Changes in carbohydrate content and gene expression during tuberous root development of sweet potato. J Plant Biochem Biotech, 2006, 15: 21-25[本文引用:1]
[44]
许欢欢, 康健, 梁明祥. 植物果聚糖的代谢途径及其在植物抗逆中的功能研究进展. 植物学报, 2014, 2: 209-220Xu HH, KangJ, Liang MX. Research advances in the metabolism of fructan in plant stress resistance, Acta Bot Sin, 2014, 2: 209-220 (in Chinese with English abstract)[本文引用:1]
[45]
KǜhbauchW, ThomeU. Nonstructural carbohydrates of wheat stems as influenced by sink-source manipulation. J Plant Physiol, 1989, 134: 243-250[本文引用:2]
[46]
SuzukiM. Fructans in crop production and preservation. Sci Tech Fruc, 1993: 227-255[本文引用:3]
[47]
Escalada JA, Moss DN. Changes in nonstructural carbohydrate fractions of developing spring wheat kernels. Crop Sci, 1976, 16: 627-631[本文引用:2]
Pollock CJ. Fructans and the metabolism of sucrose in vascular plants. New Phytol, 1986, 104: 1-24[本文引用:1]
[50]
Van den Ende W, Michiels A, De Roover J, Verhaert P, Van Laere A. Cloning and functional analysis of chicory root fructan1_ exohydrolase I(1-FEHI): a vacuolar enzyme derived from a cell wall invertase ancestor Mass fingerprint of the 1-FEHI enzyme. Plant J, 2000, 24: 447-456[本文引用:1]
[51]
Stoop JM, Van ArkelJ, Hakkert JC, TyreeC, Caimi PG, Koops AJ. Developmental modulation of inulin accumulation in storage organs of transgenic maize and transgenic potato. Plant Sci, 2007, 173: 172-181[本文引用:1]
[52]
Caimi PG, McCole L M, Klein T M, Hershey H P. Cytosolic expression of the Bacillus amyloliquefaciens SacB protein inhibits tissue development in transgenic tobacco and potato. New Phytol, 1997, 136: 19-28[本文引用:1]
[53]
Pollock CJ, JonesT. Seasonal patterns of fructan metabolism in forage grasses. New Phytol, 1979, 83: 9-15[本文引用:1]
 , 史春余
, 史春余


{kind=link}
{kind=link}