关键词:水稻; 化感作用; 转录因子; 酚酸; 基因表达调控 Phenolic Acid Synthesis of Allelopathic Rice Regulated by OsMYB and Its Weed Inhibition SHEN Li-Hua1,2, LI Bi-Liang1, REN Yong-Jie1, LI Cheng-Xun1, ZHONG Yong-Jia2, FANG Chang-Xun1,2, LIN Wen-Xiong1,2,* 1College of Life Sciences, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2Institute of Agro-ecology, Fujian Agriculture and Forestry University, Fuzhou 350002, China
AbstractRegulation of gene expression is a vital process in the formation of rice allelopathy. In this study, allelopathic rice PI312777 ( Oryza sativa L.) was modified by RNA interference (RNAi) and overexpression(OE) technologies to inhibit or enhance gene expression of OsMYB ( CT829537) in PI312777 respectively. The results showed that up-regulation of phenolic synthesis related genes was found in CT829537-OE transgenic PI312777 co-cultured with barnyardgrass (BYG), compared with that in wild type. However, the reverse was true in the CT829537-RNAi transgenic PI312777. Up-regulation of the gene expression in CT829537-OE transgenic PI312777 increased phenolic acids contents in rice root and root exudates, which led to enhance allelopathic inhibition on barnyardgrass. In contrast, decreases of phenolic acids contents and weed inhibition were found in CT829537-RNAi transgenic PI312777. These results implied that CT829537 is responsible for regulating phenolic synthesis in allelopathic rice PI312777 and then enhances allelopathic inhibition on weeds.
Keyword:Rice; Allelopathy; Transcription factor; Phenolic acid; Gene expression regulation Show Figures Show Figures
图3 半定量PCR检测CT829537基因在转基因水稻与野生型水稻中的表达量 缩写同图1。Fig. 3 Gene expression level of CT829537 in transgenic rice and WT detected by semi-quantitative RT-PCR Abbreviations are the same as those given in Fig. 1.
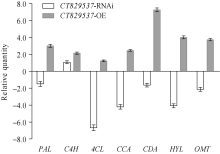
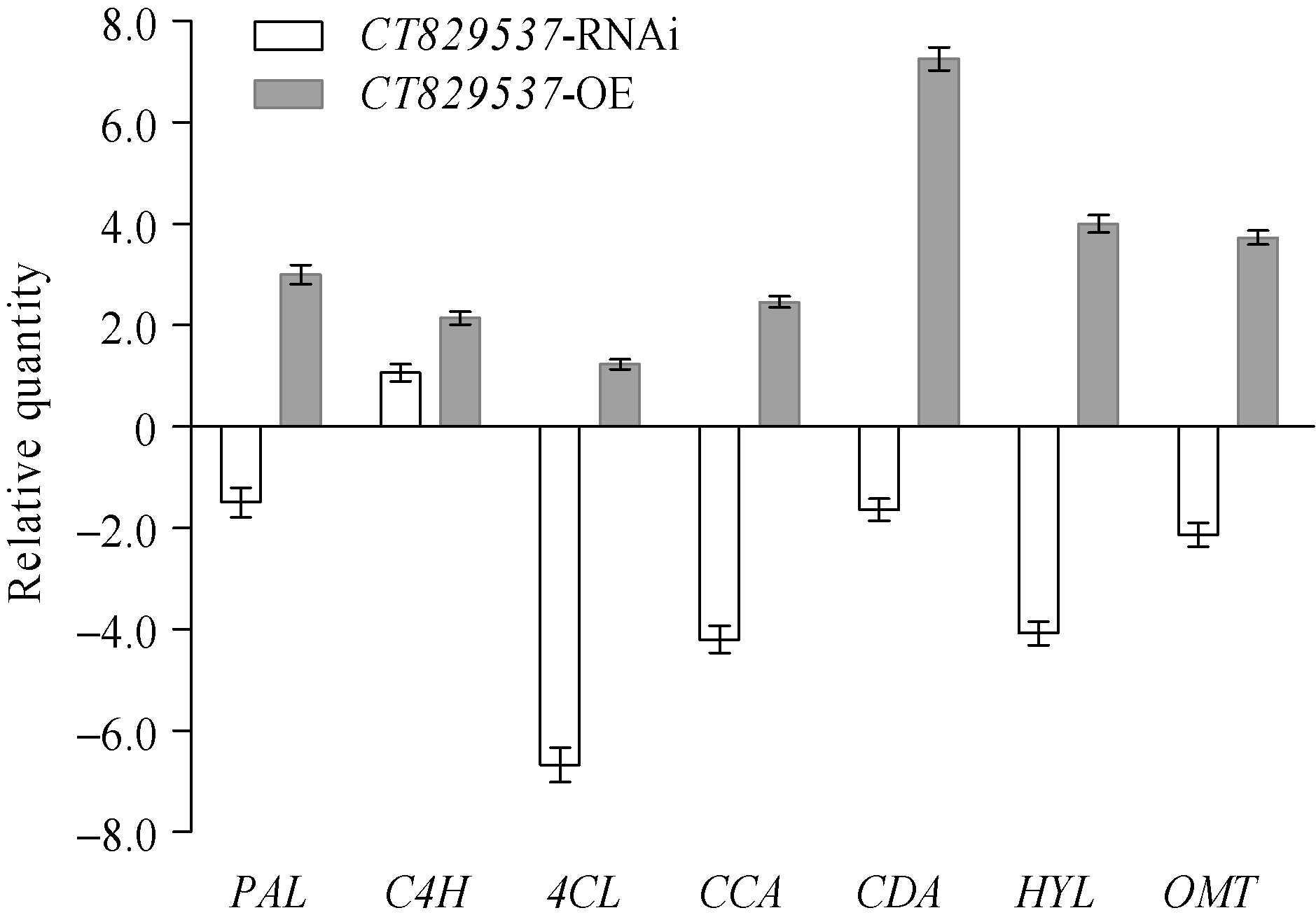
图4 与稗草共培养下, 转基因PI312777与其野生型植株的酚酸代谢关键基因的表达变化 缩写同图1。Fig. 4 Relative expression analysis of the genes relative to phenolic acid synthesis in transgenic PI312777 and WT under rice/BYG co-cultured condition Abbreviations are the same as those given in Fig. 1
表3 稗草共培养处理对转基因水稻和野生型水稻的总酚含量影响 Table 3 Comparison of total phenol content between transgenic PI312777 and WT under rice/BYG co-cultured condition
处理 Treatment
供试材料 Experimental materials
根系总酚含量 Total phenol content in root (μ mol g-1)
水培液总酚含量 Total phenol content in culture solution (μ mol L-1)
与稗草共培 Co-cultured with BYG
WT
19.69 a
24.17 b
CT829537-RNAi
15.20 b
13.74 d
CT829537-OE
20.28 a
26.90 a
无稗草共培 Without BYG
WT
15.21 b
14.20 d
CT829537-RNAi
13.52 c
13.18 d
CT829537-OE
16.92 b
20.88 c
Values followed by different small letters within a column are significantly different at the 0.05 level. Abbreviations are the same as those given in Fig. 1 同列不同小写字母表示差异显著(P< 0.05)。缩写同图1。
表3 稗草共培养处理对转基因水稻和野生型水稻的总酚含量影响 Table 3 Comparison of total phenol content between transgenic PI312777 and WT under rice/BYG co-cultured condition
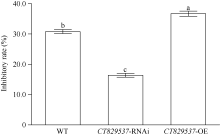
图5 转基因PI312777及野生型PI312777的根系分泌物对稗草干重的抑制率 缩写同图1。a、b、c表示不同处理之间存在显著差异(P< 0.05)。Fig. 5 Inhibitory rates of root exudates from transgenic PI312777and WT on dry weight of BYG Abbreviations are the same as those given in Fig. 1. Superscript letters indicate statistical groups that are significantly different (P< 0.05).
OlofsdotterM, NavarezD, RebulananM, Streibig JC. Weed-suppressing rice cultivars: does allelopathy play a role. Weed Res, 1999, 39: 441-454[本文引用:1]
[2]
Chou CH, Chang FJ, Oka HI. Allelopathic potentials of wild rice, Oryza perennis. Taiwania, 1991, 36: 201-210[本文引用:1]
[3]
Chou CH. Adaptive autointoxication mechanisms of Oryza sativa. In: Olofsdotter M, ed. Allelopathy in Rice. Los Banzos, Philippines: IRRI, 1996. pp 25-27[本文引用:1]
[4]
FujiiY. The allelopathic effect of some rice varities. In: Proceedings of the International Seminar Biological Control and Integrated Management of Paddy and Aquatic Weeds in Asia. Tsukuba, Japan, 1992. pp 160-165[本文引用:1]
[5]
Dilday RH, LinJ, YanW. Identification of allelopathy in the USDA-ARS rice germplasm collection. Aust J Exp Agric, 1994, 34: 907-910[本文引用:1][JCR: 1.621]
[6]
Dilday RH, Yan WG, Moldenhauer K A K, Gravois K A. Allelopathic activity in rice for controlling major aquatic weeds. In: Olofsdotter M, ed. Proceedings of Workshop on Allelopathy in Rice. Manila, Philippines: IRRI, 1998. pp 7-26[本文引用:3]
[7]
Dilday RH, NastasiP, Smith R J Jr. Allelopathic observation in rice (Oryza sativa L. ) to ducksalad (Heteranthera limosa). Proc Arkansas Acad Sci, 1989, 43: 21-22[本文引用:2]
[8]
Dilday RH, LinJ, YanW. Identification of allelopathy in the USDA-ARS rice germplasm collection. Aust J Exp Agric, 1994, 34: 907-910[本文引用:2][JCR: 1.621]
[9]
Dilday RH, Mattice JD, Moldenhauer KA. An overview of rice allelopathy in the USA. In: Kim K U, Shin D H, eds. Rice Allelopathy. Taegu (Korea): Kyungpook National University, 2000. pp 15-26[本文引用:3]
[10]
Jensen LB, CourtoisB, Shin LS, Li ZK, OlofsdotterM, Mauleon RP. Locating genes controlling allelopathic effects against barnyardgrass in upland rice. Agron J, 2001, 93: 21-26[本文引用:1][JCR: 1.518]
[11]
EbanaK, Yan WG, DildayR, NamaiH, OkunoK. Analysis of QTL associated with the allelopathic effect of rice using water soluble extracts. Breed Sci, 2001, 51: 47-51[本文引用:1][JCR: 1.04]
[12]
XiongJ, Jia XL, Deng JY. Analysis of epistatic effect and QTL interactions with environment for allelopathy in rice (Oryza sativa L. ). Allelopathy J, 2007, 20: 259-268[本文引用:1][JCR: 0.685]
[13]
Lee SB, Seo KI, Koo JH, Hur HS, Shin JC. QTLs and molecular markers associated with rice allelopathy. In: Haper J D I, An M, Kent J H, eds. Proceedings of the Fourth World Congresson Allelopathy “Establishing the scientific base”. Australia: Charles Sturt Universit, Wagga Wagga, NSW, 2005. pp 505-507[本文引用:1]
王海斌, 何海斌, 熊君, 邱龙, 方长旬, 曾聪明, 严琳, 林文雄. 低钾胁迫对水稻(Oryza sativa L. )化感潜力变化的影响. 生态学报, 2008, 28: 6219-6227Wang HB, He HB, XiongJ, QiuL, Fang CX, Zeng CM, YanL, Lin WX. Effects of potassium stress on allelopathic of rice (Oryza sativa L. ). Acta Ecol Sin, 2008, 28: 6219-6227 (in Chinese with English abstract)[本文引用:1]
[16]
王海斌, 何海斌, 叶陈英, 邱龙, 方长旬, 林文雄. 不同化感潜力水稻秧苗响应低钾的光合生理特性. 中国生态农业学报, 2008, 16: 1474-1477Wang HB, He HB, Ye CY, QiuL, Fang CX, Lin WX. Photosynthetic physiology of different allelopathic rice accessions at seedling stage under phtossium stress. Chin J Eco-Agric, 2008, 16: 1474-1477 (in Chinese with English abstract)[本文引用:1][CJCR: 0.795]
[17]
王海斌, 何海斌, 曾聪明, 吴良展, 沈荔花, 熊君, 林瑞余, 林文雄. 低磷胁迫下不同化感潜力水稻秧苗生长的分子生理特性. 应用与环境生物学报, 2008, 14: 593-598Wang HB, He HB, Zeng CM, Wu LZ, Shen LH, XiongJ, Lin RY, Lin WX. Molecular physiological properties of different rice accessions mediated by different phosphorus supplies at seedling stage. Chin J Appl Environ Biol, 2008, 14: 593-598 (in Chinese with English abstract)[本文引用:1][CJCR: 0.867]
[18]
MatticeJ, LavyT, SdulmanB, Dilday RH. Searching for allelochemicals in rice that control ducksalad. In: Olofsdotter M ed. Proceedings of Workshop on Allelopathy in Rice. Manila, Philippines: IRRI, 1998. pp81-98[本文引用:1]
[19]
Seal AN, Pratley JE, HaigT, AnM. Identification and quantitation of compounds in a series of allelopathic and nonallelopathic rice root exudates. J Chem Ecol, 2004, 30: 1647-1662[本文引用:1][JCR: 2.462]
[20]
Macias FA, ChinchillaN, Varela RM, Molinllo J M G. Bioactive steroids from Oryza sativa L. Steroids, 2006, 71: 603-608[本文引用:1][JCR: 2.803]
[21]
He HB, Wang HB, Fang, CX, Lin Z H, Yu Z M, Lin W X. Separation of allelopathy from resource competition using rice/barnyardgrass mixed-cultures. PLoS One, 2012, 7(5): e37201[本文引用:1][JCR: 3.73]
[22]
Kato NH. Barnyard grass-induced rice allelopathy and momilactone B. Plant Physiol, 2011, 168: 1016-1020[本文引用:1][JCR: 6.555]
[23]
Fang CX, He HB, Wang QS, QiuL, Wang HB, Zhuang YE, XiongJ, Lin WX. Genomic analysis of allelopathic response to low nitrogen and barnyardgrass competition in rice (Oryza sativa L. ). Plant Growth Regul, 2010, 61: 277-286[本文引用:2][JCR: 1.67]
[24]
Fang CX, Zhuang YE, Xu TC, Li YZ, LiY, Lin WX. Changes in rice allelopathy and rhizosphere microflora by inhibiting rice henylalanine ammonia-lyase gene expression. J Chem Ecol, 2013, 39: 204-212[本文引用:2][JCR: 2.462]
[25]
PutuS, TsutomuS, HidenariS, MasahiroN, MasayukiN, MincoK. Development of simple and efficient in planta transformation method for rice (Oryza sativa L. ) using Agrobacterium tumefaciens. J Biosci Bioeng, 2005, 100: 391-397[本文引用:1][JCR: 1.737]
[26]
Fang CX, Wang QS, YuY, Lin QM, Zhang HL, Wu XC, ChenT, Lin WX. Suppression and overexpression of Lsi1 induce differential gene expression in rice under ultraviolet radiation. Plant Growth Regul, 2011, 65: 1-10[本文引用:1]
[27]
熊君, 王海斌, 方长旬, 邱龙, 吴文祥, 何海斌, 林文雄. 不同氮素供应下水稻酚类物质代谢关键酶基因差异表达. 植物生理与分子生物学学报, 2007, 33: 387-394XiongJ, Wang HB, Fang CX, QiuL, Wu WX, He HB, Lin WX. The differential expression of the genes of the key enzymes involved in phenolic compound metabolism in rice (Oryza sativa L. ) under different nitrogen supply. J Plant Physiol Mol Biol, 2007, 33: 387-394 (in Chinese with English abstract)[本文引用:1][CJCR: 1.344]
[28]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
[29]
庄月娥. 水稻抑草作用关键基因的分子生态机制研究. 福建农林大学硕士学位论文, 福建福州, 2011Zhuang YE. Molecular Ecological Mechanism of the Key Gene for Weed-suppression in Allelopathic Rice. MS Thesis of Fujian Agriculture and Forestry University, Fuzhou, China, 2011 (in Chinese with English abstract)[本文引用:1]
[30]
Song BQ, XiongJ, Fang CX, QiuL, Lin RY, Liang YY, Lin WX. Allelopathic enhancement and differential gene expression in rice under low nitrogen treatment. J Chem Ecol, 2008, 34: 688-695[本文引用:1][JCR: 2.462]
[31]
XiongJ, WangH, QiuL, Wu HW, Chen RS, He HB, Lin RY, Lin W X. qRT-PCR analysis of key enzymatic genes related to phenolic acid metabolism in rice accessions (Oryza sativa L. ) exposed to low nitrogen treatment. Allelopathy J, 2010, 25: 345-356[本文引用:1][JCR: 0.685]
[32]
王海斌, 熊君, 方长旬, 邱龙, 吴文祥, 何海斌, 林文雄. 氮素胁迫下强、弱化感水稻萜类代谢途径中关键酶基因差异表达的FQ-PCR分析. 作物学报, 2007, 33(8): 1329-1334Wang HB, XiongJ, Fang CX, QiuL, Wu WX, He HB, Lin WX. FQ-PCR analysis on the differential expression of the key enzyme genes involved in isoprenoid metabolic pathway in allelopathic and weak allelopathic rice accessions (Oryza sativa L. ) under nitrogen stress condition. Acta Agron Sin, 2007, 33(8): 1329-1334 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[33]
孙小霞, 王海斌, 林辉锋, 何海斌, 陆锦池, 曾聪明, 熊君, 林文雄. 田间旱育条件下不同化感潜力水稻的抑草效应分析. 中国生态农业学报, 2009, 17: 842-846Sun XX, Wang HB, Lin HF, He HB, Lu JC, Zeng CM, XiongJ, Lin WX. Effects of weed suppression by different allelopathic rice varieties under dry-raising condition. Chin J Eco-Agric, 2009, 17: 842-846 (in Chinese with English abstract)[本文引用:1][CJCR: 0.795]
[34]
孙小霞, 王海斌, 何海斌, 陆锦池, 林文雄. 田间旱育条件下不同化感潜力水稻根际土壤酚酸类和萜类物质分析. 中国生态农业学报, 2014, 22: 806-812Sun XX, Wang HB, He HB, Lu JC, Lin WX. Analysis of phenolic acids and terpenoids in rhizosphere soils of different allelopathic rice varieties under dry field conditions. Chin J Eco-Agric, 2014, 22: 806-812 (in Chinese with English abstract)[本文引用:1][CJCR: 0.795]
[35]
JunaediA, Jung WS, Chung IM, Kim KH. Differentially expressed genes of potentially allelopathic rice in response against barnyardgrass. J Crop Sci Biotechnol, 2007, 10: 231-236[本文引用:1]
[36]
PazaresJ, GhosalD, Wienand U, Peterson PA, SaedlerH. There gulatory C1 locus of Zea mays encodes a proterin with homology to myb-oncogene products and with structural similarities to transcriptional activators. EMBO J, 1987, 6: 3553-3558[本文引用:1][JCR: 9.822]
[37]
Cone KC, Burr FA, BurrB. Molecular analysis of the maize anthocyanin regulatory locus C1. Proc Natl Acad Sci USA, 1986, 83: 9631-9635[本文引用:1][JCR: 9.737]
[38]
MoyanoE, Martinez-Garcia J F, Martin C. Apparent redundancy in myb gene function provides gearing for the control of flavonoid biosynthesis in Antirrhinum flowers. Plant Cell, 1996, 8: 1519-1532[本文引用:1][JCR: 9.251]
[39]
LegayS, LacombeE, GoicoecheaM, BrièreC, SéguinA, MackayJ, Grima-PettenatiJ. Molecular characterization of EgMYB1, a putative transcriptional repressor of the lignin biosynthetic pathway. Plant Sci, 2007, 173: 542-549[本文引用:1][JCR: 2.922]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}