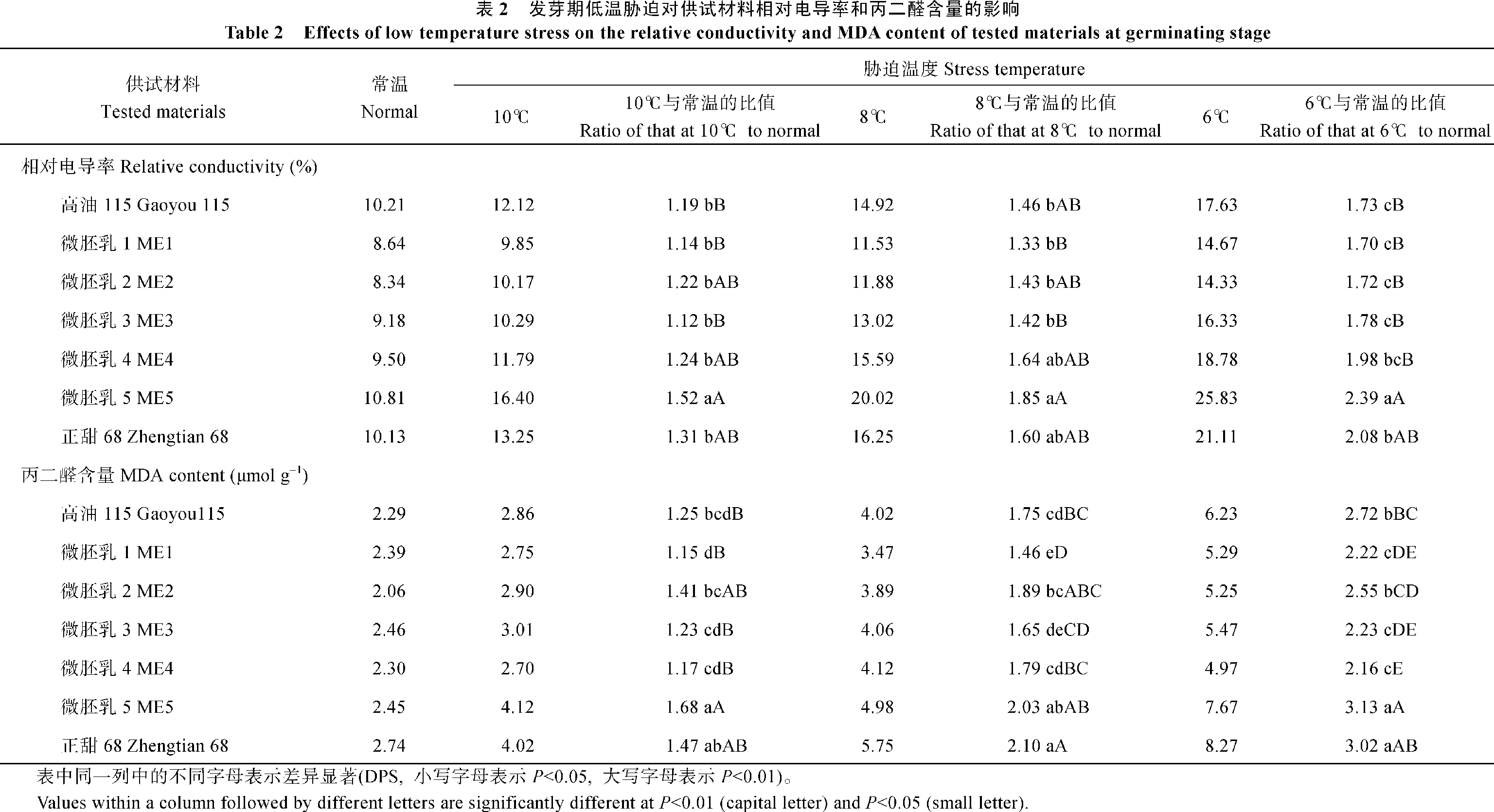
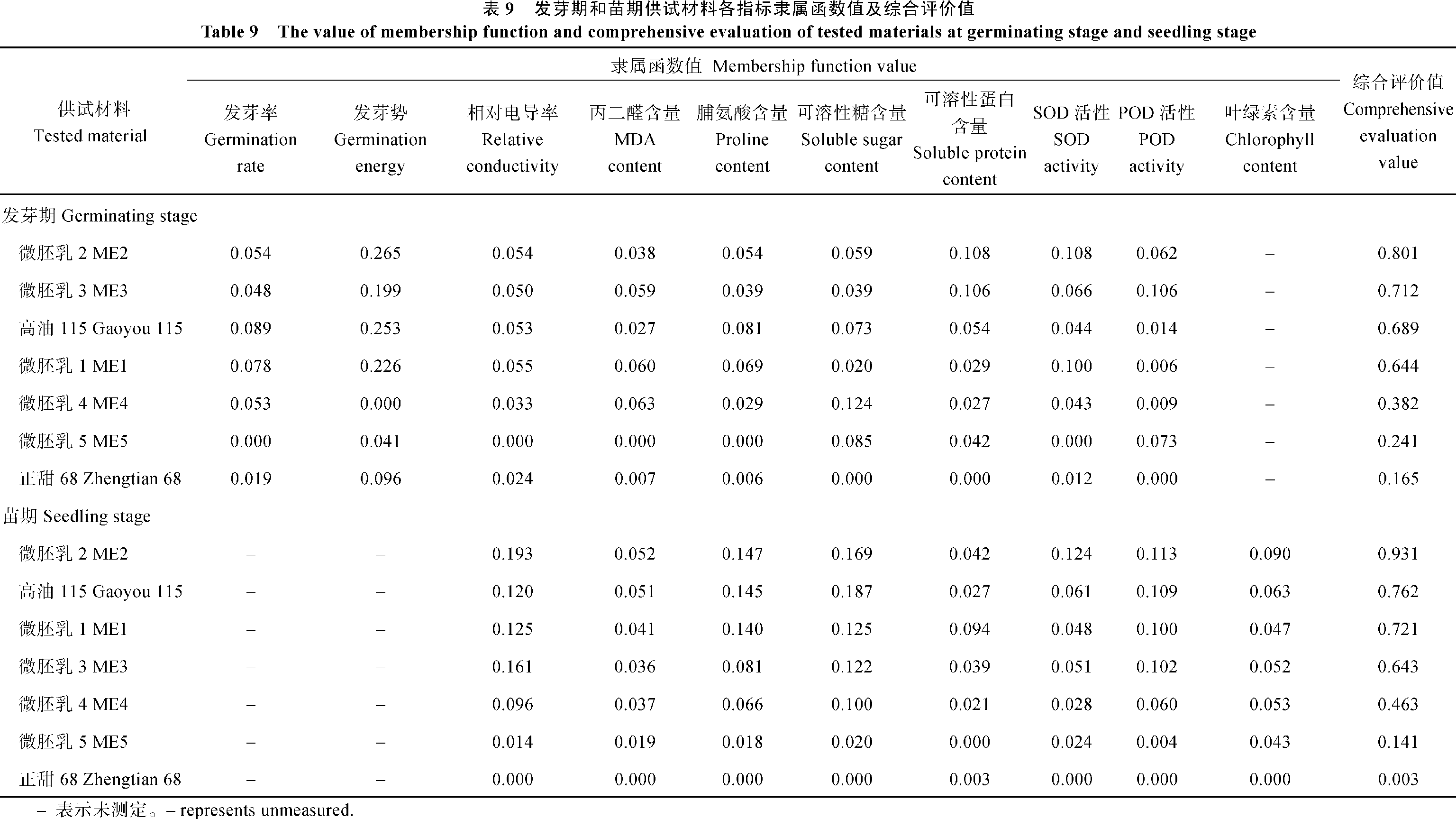
关键词:微胚乳超甜超高油玉米; 低温胁迫; 耐寒性; 生理生化特性 Effects of Low Temperature Stress on the Physiological and Biochemical Characteristics of Cold Tolerance in Micro-endosperm Super Sweet and Super High Oil Maize HAO Xiao-Qin, YAO Peng-He, GAO Zheng-Rong, WU Zi-Kai Agricultural College of Guangxi University, Nanning 530005, China Fund: AbstractIn the germination and seedling stages of the maize, five micro-endosperm super sweet and super high oil maize (abbreviation as ME) hybrid combinations, two control materials Gaoyou 115 and Zhengtian 68 were used to study the changes of physiological and biochemical characteristics, such as germination rate, germination energy, relative conductivity, contents of MDA, proline, soluble sugar, soluble protein, chlorophyll, and SOD, POD activities under low temperature stress. At the same time, the identification of cold tolerance of the materials were conducted. Based on the multiple physiological and biochemical parameters, we employed the fuzzy membership function to evaluate the cold tolerance of the experiment materials comprehensively. The results showed that the relative conductivity had a little increase, but the proline content and SOD activity increased largely in ME1, ME2, ME3, and Gaoyou 115, as compared with ME5, ME4 and Zhengtian 68 in the germination stage. It was advised that relative conductivity, proline content and SOD activity could be used to predict cold tolerance of the materials more effectively in germination stage. In the seedling stage, compared with ME5 and Zhengtian 68, the MDA content increased a little and activities of SOD, POD and proline content increased largely in ME2, ME1, and Gaoyou 115. Therefore, we thought that MDA content, proline content, SOD and POD activities can be used to predict the cold tolerance of the experiment materials in seedling stage. Meanwhile, membership function value showed that the cold tolerance of the five ME materials was stronger than that of Zhengtian 68. Among the five ME materials, ME2 showed a strong ability to tolerate low temperature stress in germination and seedling stages, ME1 and ME3 showed moderate tolerance, and ME4 and ME5 showed weak tolerance.
Keyword:Micro-endosperm super sweet and super high oil maize; Low temperature stress; Cold tolerance; Physiological and biochemical characteristics Show Figures Show Figures
表4 发芽期低温胁迫对供试材料SOD和POD活性的影响 Table 4 Effects of low temperature stress on SOD and POD activities of tested materials at germinating stage
供试材料 Tested materials
常温 Normal
胁迫温度Stress temperature
10℃
10℃与常温的比值 Ratio of that at 10℃ to normal
8℃
8℃与常温的比值 Ratio of that at 8℃ to normal
6℃
6℃与常温的比值 Ratio of that at 6℃ to normal
SOD酶活性 SOD activity (U g-1)
高油115 Gaoyou 115
164.20
198.01
1.21 bB
135.68
0.83 abcdAB
87.56
0.53 bBC
微胚乳1 ME1
205.69
274.23
1.33 abAB
180.75
0.88 abAB
151.99
0.74 aA
微胚乳2 ME2
181.61
246.76
1.36 aAB
174.90
0.96 aA
139.09
0.77 aA
微胚乳3 ME3
163.91
229.59
1.40 aAB
140.52
0.86 abcAB
100.67
0.61 bB
微胚乳4 ME4
147.83
209.52
1.42 aA
107.41
0.73 bcdB
78.57
0.53 bBC
微胚乳5 ME5
188.70
250.37
1.33 abAB
134.83
0.72 cdB
70.73
0.42 cCD
正甜68 Zhengtian 68
195.34
251.92
1.29 abAB
131.31
0.67 dB
81.99
0.37 cD
POD酶活性 POD activity (A470 g-1 FW min-1)
高油115 Gaoyou 115
0.47
0.14
0.29 cdCD
0.18
0.39 bBC
0.14
0.30 bcB
微胚乳1 ME1
1.22
0.31
0.25 dD
0.40
0.33 bC
0.34
0.28 bcB
微胚乳2 ME2
0.84
0.30
0.35 bcBCD
0.56
0.66 aA
0.34
0.40 abcAB
微胚乳3 ME3
0.77
0.24
0.31 cdCD
0.24
0.32 bC
0.38
0.50 aA
微胚乳4 ME4
0.63
0.31
0.49 aA
0.35
0.55 aAB
0.18
0.29 bcB
微胚乳5 ME5
1.06
0.46
0.44 abAB
0.64
0.61 aA
0.45
0.42 abAB
正甜68 Zhengtian 68
0.63
0.25
0.40 abABC
0.25
0.39 bBC
0.17
0.27 cB
表中同一列中的不同字母表示差异显著(DPS, 小写字母表示 P<0.05, 大写字母表示 P<0.01)。 Values within a column followed by different letters are significantly different at P<0.01 (capital letter) and P<0.05 (small letter).
表4 发芽期低温胁迫对供试材料SOD和POD活性的影响 Table 4 Effects of low temperature stress on SOD and POD activities of tested materials at germinating stage
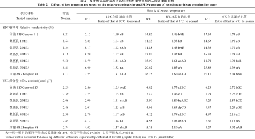
表5 苗期低温胁迫对供试材料相对电导率和丙二醛含量的影响 Table 5 Effects of low temperature stress on the relative conductivity and MDA content of tested materials at seedling stage
供试材料 Tested materials
低温胁迫时间Time of low temperature stress
0 d
1 d
胁迫1 d与0 d 的比值 Ratio of stress 1 d to 0 d
3 d
胁迫3 d与0 d的比值 Ratio of stress 3 d to 0 d
5 d
胁迫5 d与0 d 的比值 Ratio of stress 5 d to 0 d
相对电导率 Relative conductivity (%)
高油115 Gaoyou 115
6.51
9.36
1.44 abAB
11.69
1.80 cdB
20.17
3.10 bcBC
微胚乳1 ME1
6.68
9.05
1.35 bAB
16.17
2.42 bA
20.29
3.04 bcBC
微胚乳2 ME2
8.44
9.96
1.18 bB
12.32
1.46 cdB
18.98
2.25 cC
微胚乳3 ME3
9.44
13.12
1.39 bAB
22.32
2.36 bA
24.78
2.63 bcC
微胚乳4 ME4
10.25
11.94
1.17 bB
14.66
1.43 dB
34.62
3.38 bABC
微胚乳5 ME5
13.77
16.74
1.22 bB
25.21
1.83 cB
59.69
4.34 aAB
正甜68 Zhengtian 68
6.12
10.59
1.73 aA
17.64
2.88 aA
27.52
4.49 aA
丙二醛含量 MDA content (μmol g-1)
高油115 Gaoyou115
3.84
5.10
1.33 bA
7.14
1.86 cdBC
11.97
3.11 dC
微胚乳1 ME1
3.38
4.90
1.45 abA
6.91
2.05 abAB
10.92
3.24 cdC
微胚乳2 ME2
3.78
4.96
1.31 bA
6.75
1.79 dC
11.70
3.10 dC
微胚乳3 ME3
3.73
5.00
1.34 bA
7.35
1.97 bcABC
12.30
3.30 cC
微胚乳4 ME4
3.56
4.69
1.32 bA
7.07
1.99 bcABC
11.69
3.28 cC
微胚乳5 ME5
3.98
5.73
1.44 abA
8.14
2.05 abAB
13.93
3.50 bB
正甜68 Zhengtian 68
3.75
5.76
1.53 aA
8.02
2.14 aA
14.03
3.74 aA
表中同一列中的不同字母表示差异显著(DPS, 小写字母表示 P<0.05, 大写字母表示 P<0.01)。 Values within a column followed by different letters are significantly different at P<0.01 (capital letter) and P<0.05 (small letter).
表5 苗期低温胁迫对供试材料相对电导率和丙二醛含量的影响 Table 5 Effects of low temperature stress on the relative conductivity and MDA content of tested materials at seedling stage
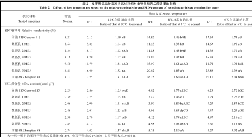
表6 苗期低温胁迫对供试材料脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 Table 6 Effects of low temperature stress on proline content, soluble sugar content and soluble protein content of tested materials at seedling stage
供试材料 Tested materials
低温胁迫时间Time of low temperature stress
0 d
1 d
胁迫1 d与0 d的比值 Ratio of stress 1 d to 0 d
3 d
胁迫3 d与0 d的比值 Ratio of stress 3 d to 0 d
5 d
胁迫5 d与0 d的比值 Ratio of stress 5 d to 0 d
脯氨酸含量 Proline content (µg g-1)
高油115 Gaoyou 115
11.44
11.89
1.04 aA
18.51
1.62 abAB
35.96
3.14 aA
微胚乳1 ME1
12.05
15.28
1.27 aA
23.61
1.96 aA
37.31
3.10 abA
微胚乳2 ME2
15.64
20.33
1.30 aA
24.45
1.56 abAB
49.35
3.16 aA
微胚乳3 ME3
17.52
19.25
1.10 aA
28.35
1.62 abAB
45.81
2.62 bcAB
微胚乳4 ME4
16.74
19.80
1.18 aA
21.42
1.28 bB
41.63
2.49 cdAB
微胚乳5 ME5
15.89
16.50
1.04 aA
24.39
1.54 abAB
33.26
2.09 deB
正甜68 Zhengtian 68
16.79
17.87
1.07 aA
21.16
1.26 bB
32.63
1.94 eB
可溶性糖含量 Soluble sugar content (mg g-1)
高油115 Gaoyou 115
9.04
8.44
0.93 aA
11.48
1.27 aA
12.01
1.33 aA
微胚乳1 ME1
8.34
5.68
0.68 bA
6.28
0.75 dC
9.24
1.11 abA
微胚乳2 ME2
6.28
5.71
0.91 aA
7.88
1.26 aA
7.94
1.27 aA
微胚乳3 ME3
6.64
5.28
0.79 abA
5.61
0.84 cdBC
7.28
1.10 abA
微胚乳4 ME4
6.84
5.38
0.79 abA
6.68
0.98 bB
6.98
1.02 bABC
微胚乳5 ME5
6.51
4.84
0.73 abA
5.88
0.88 bcBC
4.88
0.73 cBC
正甜68 Zhengtian 68
10.28
6.94
0.68 bA
8.28
0.81 cdC
6.81
0.66 cC
可溶性蛋白含量 Soluble protein content (mg g-1)
高油115 Gaoyou 115
1.17
7.69
6.60 cB
4.02
3.45 cBC
3.32
2.85 aA
微胚乳1 ME1
0.79
6.93
8.75 aA
4.62
5.84 aA
2.86
3.61 aA
微胚乳2 ME2
1.12
7.17
6.41 cBC
3.98
3.55 cBC
3.40
3.03 aA
微胚乳3 ME3
1.36
6.21
4.57 dD
3.93
2.89 cC
4.07
2.99 aA
微胚乳4 ME4
1.16
6.14
5.31 dCD
4.04
3.50 cBC
3.22
2.79 aA
微胚乳5 ME5
0.86
6.40
7.48 bB
4.00
4.67 bAB
2.19
2.55 aA
正甜68 Zhengtian 68
0.78
6.91
8.81 aA
4.21
5.37 abA
2.03
2.59 aA
表中同一列中的不同字母表示差异显著(DPS, 小写字母表示 P<0.05, 大写字母表示 P<0.01)。 Values within a column followed by different letters are significantly different at P<0.01 (capital letter) and P<0.05 (small letter).
表6 苗期低温胁迫对供试材料脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 Table 6 Effects of low temperature stress on proline content, soluble sugar content and soluble protein content of tested materials at seedling stage
表7 苗期低温胁迫对供试材料SOD、POD酶活性和叶绿素含量的影响 Table 7 Effects of low temperature stress on SOD, POD activities and chlorophyll content of tested materials at seedling stage
供试材料 Tested materials
低温胁迫时间Time of low temperature stress
0 d
1 d
胁迫1 d与0 d的比值 Ratio of stress 1 d to 0 d
3 d
胁迫3 d与0 d的比值 Ratio of stress 3 d to 0 d
5 d
胁迫5 d与0 d的比值 Ratio of stress 5 d to 0 d
SOD酶活性SOD activity (U g-1)
高油115 Gaoyou 115
185.18
266.08
1.44 aA
204.81
1.11 abcABC
215.31
1.16 abAB
微胚乳1 ME1
273.68
330.09
1.21 aA
233.95
0.86 dBC
302.86
1.11 bAB
微胚乳2 ME2
209.81
253.12
1.21 aA
175.95
0.84 dC
304.47
1.45 aA
微胚乳3 ME3
227.57
287.29
1.26 aA
265.84
1.17 abAB
254.55
1.12 bAB
微胚乳4 ME4
224.99
295.93
1.32 aA
270.52
1.20 aA
228.71
1.02 bAB
微胚乳5 ME5
198.64
264.51
1.33 aA
193.81
0.98 bcdABC
197.77
0.99 bAB
正甜68 Zhengtian 68
264.76
305.35
1.15 aA
241.92
0.91 cdABC
234.77
0.89 bB
POD酶活性 POD activity (A470 g-1 FW min-1)
高油115 Gaoyou 115
0.31
0.42
1.36 abA
0.26
0.85 bAB
0.40
1.29 aA
微胚乳1 ME1
0.39
0.62
1.58 aA
0.46
1.16 aA
0.49
1.26 aA
微胚乳2 ME2
0.30
0.38
1.28 abA
0.22
0.73 bB
0.39
1.30 aA
微胚乳3 ME3
0.31
0.41
1.34 abA
0.36
1.17 aA
0.39
1.27 aA
微胚乳4 ME4
0.42
0.44
1.06 bA
0.32
0.78 bB
0.47
1.13 abAB
微胚乳5 ME5
0.40
0.44
1.10 bA
0.34
0.86 bAB
0.37
0.93 bB
正甜68 Zhengtian 68
0.39
0.44
1.14 bA
0.31
0.79 bB
0.35
0.92 bB
叶绿素含量 Chlorophyll content (mg g-1)
高油115 Gaoyou 115
3.86
3.78
0.98 aA
2.96
0.77 aA
2.74
0.71 abA
微胚乳1 ME1
3.44
3.12
0.91 abA
2.66
0.77 aA
2.29
0.66 bAB
微胚乳2 ME2
3.28
3.03
0.92 abA
2.53
0.77 aA
2.59
0.79 aA
微胚乳3 ME3
4.12
3.82
0.93 abA
3.02
0.73 aA
2.79
0.68 abAB
微胚乳4 ME4
3.69
2.93
0.80 bA
2.79
0.76 aA
2.51
0.68 abAB
微胚乳5 ME5
3.22
2.74
0.85 abA
2.48
0.77 aA
2.10
0.65 bAB
正甜68 Zhengtian 68
3.33
2.89
0.87 abA
2.16
0.65 aA
1.75
0.53 cB
表中同一列中的不同字母表示差异显著(DPS, 小写字母表示 P<0.05, 大写字母表示 P<0.01)。 Values within a column followed by different letters are significantly different at P<0.01 (capital letter) and P<0.05 (small letter).
表7 苗期低温胁迫对供试材料SOD、POD酶活性和叶绿素含量的影响 Table 7 Effects of low temperature stress on SOD, POD activities and chlorophyll content of tested materials at seedling stage
扈光辉. 玉米抗冷性研究进展. 黑龙江农业科学, 2009, (2): 150-152HuG H. Research advancement on chilling tolerance in maize. Heilongjiang Agric Sci, 2009, (2): 150-152 (in Chinese with English abstract)[本文引用:1][CJCR: 0.3268]
[2]
谭振波, 刘昕, 曹鸣庆. 玉米抗寒性的研究进展. 玉米科学, 2002, 10(2): 56-60TanZ B, LiuX, CaoM Q. Recent advances on mechanisms of maize chilling tolerance. J Maize Sci, 2002, 10(2): 56-60 (in Chinese with English abstract)[本文引用:1][CJCR: 0.965]
[3]
LyonsJ M, RaisonJ K. Oxidative activity of mitochondria isolated from plant tissues sensitive and resistant to chilling injury. Plant Physiol, 1970, 45: 386-389[本文引用:1][JCR: 6.555]
[4]
LyonsJ M. Relationship between the physical nature of mitochondrial membranes and chilling sensitivity in plants. Am Oil Chem Soc, 1973, 42: 1056-1058[本文引用:1][JCR: 1.592]
[5]
王华, 王飞, 陈登文, 丁勤. 低温胁迫对杏花SOD活性和膜脂过氧化的影响. 果树科学, 2000, 17: 197-201WangH, WangF, ChenD W, DingQ. Effects of low temperature stress on SOD activity and membrane deroxidization of apricot flowers. J Fruit Sci, 2000, 17: 197-201 (in Chinese with English abstract)[本文引用:1][CJCR: 1.045]
[6]
冯献宾, 董倩, 王洁, 庞曼, 路丙社. 低温胁迫对黄连木抗寒生理指标的影响. 中国农学通报, 2011, 27(8): 23-26FengX B, DongQ, WangJ, PangM, LuB S. Effects of low temperature on cold resistance of Pistacia chinensis. Chin Agric Sci Bull, 2011, 27(8): 23-26 (in Chinese with English abstract)[本文引用:1]
[7]
马凤鸣, 王瑞, 石振. 低温胁迫对玉米幼苗某些生理指标的影响. 作物杂志, 2007, (5): 41-45MaF M, WangR, ShiZ. Effect of low temperature stress on some physiology parameter of maize seedling. Crops, 2007, (5): 41-45 (in Chinese)[本文引用:2][CJCR: 0.6276]
[8]
高素华. 玉米低温冷害机理研究综述. 气象科技, 1997, (4): 37-43GaoS H. Summarize of research on mechanism of low temperature cold harm in maize. Sci Technol Meteorol, 1997, (4): 37-43 (in Chinese)[本文引用:2]
[9]
杨亚军, 郑雷英, 王新超. 冷驯化和ABA对茶树抗寒力及其体内脯氨酸含量的影响. 茶叶科学, 2004, 24(3): 177-182YangY J, ZhengL Y, WangX C. Effect of cold acclimation and ABA on cold hardiness contents of proline in tea plants. J Tea Sci, 2004, 24(3): 177-182 (in Chinese with English abstract)[本文引用:1][CJCR: 1.225]
[10]
杨光, 刘宏魁, 李世鹏, 吴颖, 苏胜忠, 单晓辉, 原亚萍. 玉米抗冷种质资源的筛选与鉴定. 玉米科学, 2012, 20(1): 57-60YangG, LiuH K, LiS P, WuY, SuS Z, ShanX H, YuanY P. Screening and identification of chilling resistant germplasm resources in maize. J Maize Sci, 2012, 20(1): 57-60 (in Chinese with English abstract)[本文引用:1][CJCR: 0.965]
[11]
王淑杰, 王连君, 王家民, 杨晓光, 马秀华. 果树抗寒生理研究进展. 北方园艺, 1998, (5): 28-29WangS J, WangL J, WangJ M, YangX G, MaX H. Research advancement on physiology of cold resistance in fruit. Northern Hortic, 1998, (5): 28-29 (in Chinese)[本文引用:2]
[12]
马尧, 杨国会, 王春兰, 田永清. 在干旱和低温胁迫下薏苡幼苗抗逆性研究. 农业与技术, 2000, 20(5): 41-42MaY, YangG H, WangC L, TianY Q. Research of jobstears seedling in adversity resistance on cold treatment and drought condition. Agric & Technol, 2000, 20(5): 41-42 (in Chinese with English abstract)[本文引用:1][JCR: 0.114]
[13]
高志红, 章镇, 韩振海. 果梅种质枝条抗寒性鉴定. 果树学报, 2005, 22: 709-711GaoZ H, ZhangZ, HanZ H. Identification of freezing resistance in Japanese apricot germplasm. J Fruit Sci, 2005, 22: 709-711 (in Chinese with English abstract)[本文引用:1][CJCR: 1.045]
[14]
LevittJ. Responses of Plant to Environmental Stress. New York: Academic Press, 1980. pp365-488[本文引用:1]
[15]
UemuraS, GilmourS J, ThomashowM F. Effects of COR 6. 6 and COR 15am polypeptides encoded by COR gene of Arabidopsis thaliana. Plant Physiol, 1996, 111: 313-327[本文引用:1][JCR: 6.555]
[16]
武兰芳. 低温下玉米幼苗叶片某些细胞器可溶性蛋白质的变化. 华北农学报, 1990, 5(2): 38-43WuL F. Changes of soluble protein in some organelles of maize seedling leaf at low temperature. Acta Agric Boreali-Sin, 1990, 5(2): 38-43 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[17]
张敬贤, 李俊明, 崔四平, 魏建昆, 张海明, 耿庆汉. 低温对玉米幼苗细胞保护酶活性及胞质质量参数的影响. 华北农学报, 1993, 8(3): 9-12ZhangJ X, LiJ M, CuiS P, WeiJ K, ZhangH M, GengQ H. The effect of low temperature on activities of cell protective enzymes and protoplasmic parameters in leaves of Zea mays. Acta Agric Boreali-Sin, 1993, 8(3): 9-12 (in Chinese with English abstract)[本文引用:2][CJCR: 0.951]
[18]
王金胜, 郭春绒, 张述义. 低温对不同抗冷性玉米幼苗H2O2及其清除酶类的影响. 山西农业大学学报, 1993, 13: 240-243WangJ S, GuoC R, ZhangS Y. The effect of low temperature on H2O2 and its clearance enzymes in maize seeklings with different chilling resistance. J Shanxi Agric Univ, 1993, 13: 240-243 (in Chinese with English abstract)[本文引用:2]
[19]
王兰, 蔡千蕙. 低温胁迫对水稻苗期SOD、POD活性的影响. 湖南农业科学, 2011, (11): 56-58WangL, CaiQ H. Impacts of cold stress on activities of SOD and POD of rice at seedling stage. Hunan Agric Sci, 2011, (11): 56-58 (in Chinese with English abstract)[本文引用:2][CJCR: 0.451]
[20]
陈卫国, 刘勇, 周冀衡. 低温胁迫对烟草膜保护酶系统的影响. 安徽农业科学, 2011, 39: 3978-3980ChenW G, LiuY, ZhouJ H. Influence of low temperature stress on the membrane protected enzyme system in tobacco. J Anhui Agric Sci, 2011, 39: 3978-3980 (in Chinese with English abstract)[本文引用:2][CJCR: 0.687]
[21]
王晨光, 王希, 苍晶, 杨丽娟. 低温胁迫对水稻幼苗抗冷性的影响. 东北农业大学学报, 2004, 35(2): 129-134WangC G, WangX, CangJ, YangL J. Effect of low-tempera-ture stress on cold-resistance ability of rice seedlings. J Northeast Agric Univ, 2004, 35(2): 129-134 (in Chinese with English abstract)[本文引用:1]
[22]
朱祥春, 矫洪涛, 史芝文, 袁海峰. 低温胁迫对玉米幼苗抗冷性鉴定. 农业与技术, 2004, 24(1): 44-49ZhuX C, JiaoH T, ShiZ W, YuanH F. The low temperature is forced combating cooling quality of the appraisal to the corn seedling. Agric & Technol, 2004, 24(1): 44-49 (in Chinese with English abstract)[本文引用:1][JCR: 0.114]
[23]
杨静, 王华田, 宋承东, 张培法, 王迎. 持续低温胁迫对红叶石楠抗寒生理生化特性的影响. 江西农业大学学报, 2007, 29: 988-992YangJ, WangH T, SongC D, ZhangP F, WangY. Effect of durative low temperature on physiological and biochemical characteristics of Photinia glabra var Rubens. Acta Agric Univ Jiang-xiensis, 2007, 29: 988-992 (in Chinese with English abstract)[本文引用:1]
[24]
王宁, 吴军, 夏鹏云, 苏金乐. 低温胁迫对大叶冬青生理特性的影响. 河南农业大学学报, 2011, 45: 407-410WangN, WuJ, XiaP Y, SuJ L. Effects of low temperature stress on physiological characteristics of Ilex latifolia Thunb. J Henan Agric Univ, 2011, 45: 407-410 (in Chinese with English abstract)[本文引用:1][CJCR: 0.692]
[25]
吴子恺. 新型超高油玉米种质的选育. 作物学报, 2004, 30: 739-744WuZ K. Breeding for new germplasm of super-high oil in corn. Acta Agron Sin, 2004, 30: 739-744 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
张宪政. 作物生理研究方法. 北京: 农业出版社, 1992. pp144-216ZhangX Z. Study Method of Physiology in Crops. Beijing: Agriculture Press, 1992. pp144-216(in Chinese)[本文引用:1]
[27]
许桂芳, 张朝阳, 向佐湘. 利用隶属函数法对4种珍珠菜属植物的抗寒性综合评价. 西北林学院学报, 2009, 24(3): 24-26XuG F, ZhangZ Y, XiangZ X. Comprehensive evaluation of cold resistance on four lysimachia plants by subordinate function values analysis. J Northwest For Univ, 2009, 24(3): 24-26 (in Chinese with English abstract)[本文引用:1]
[28]
孙清鹏, 许煌灿, 张方秋, 尹光天, 张玉福. 低温胁迫对大叶相思和马占相思某些生理特性的影响. 林业科学研究, 2002, 15: 34-40SunQ P, XuH C, ZhangF Q, YinG T, ZhangY F. Effects of low temperature stress on some physiological changes of Acacia auriculaeformis and A. mangium. For Res, 2002, 15: 34-40 (in Chinese with English abstract)[本文引用:1][JCR: 1.847]
 , 姚鹏鹤, 高峥荣, 吴子恺
, 姚鹏鹤, 高峥荣, 吴子恺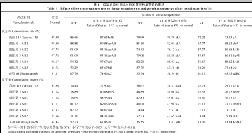


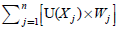 , j= 1, 2, 3,
, j= 1, 2, 3, 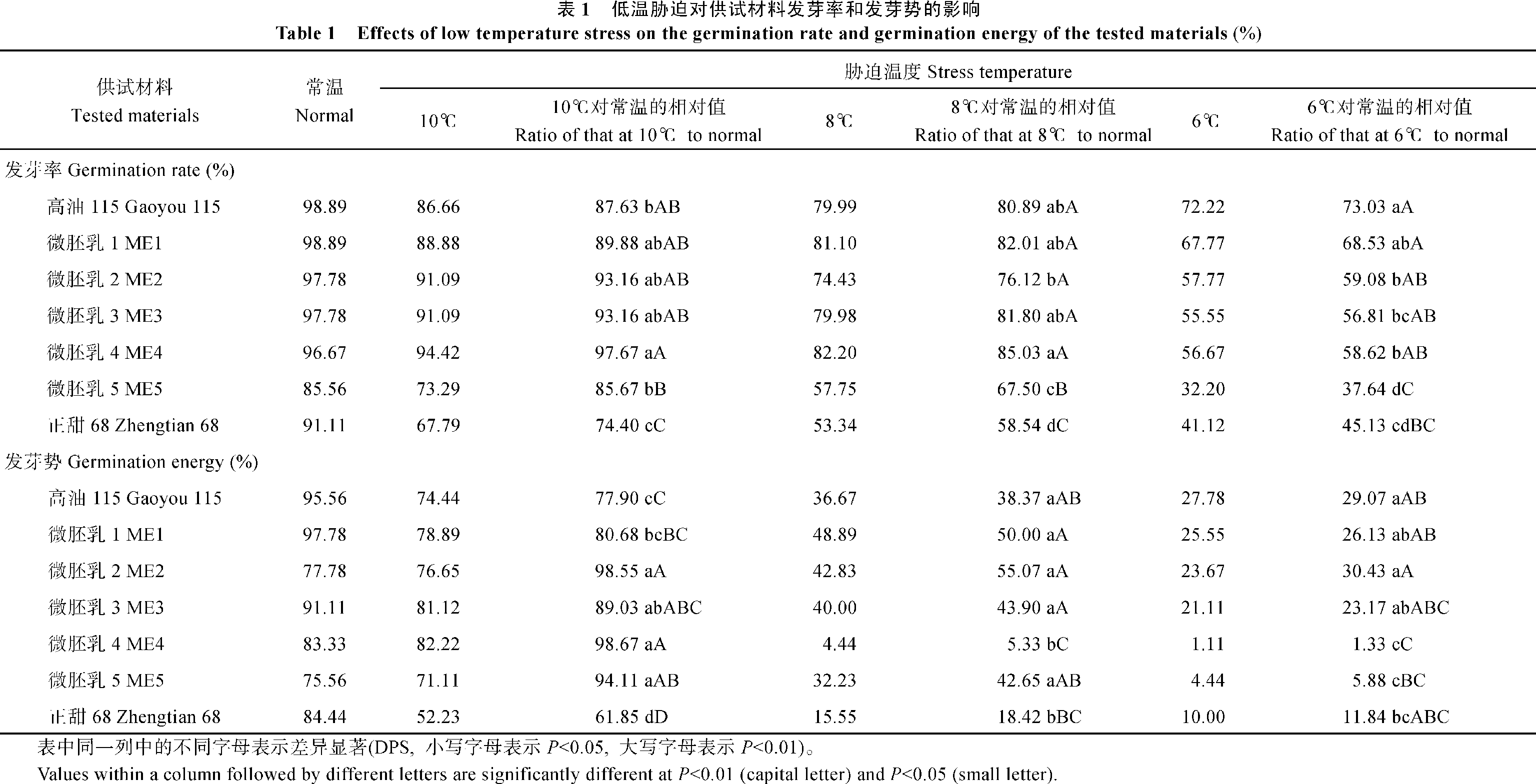



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}