关键词:水稻; 分蘖盛期; 1; 2; 4-三氯苯; 毒性; 生长发育; 抗氧化酶 Effects of 1,2,4-trichlorobenzene on Growth and Physiological Characteristics of Rice at Maximum Tillering Stage DING Xiu-Wen1,2,4, ZHANG Guo-Liang2,3,*, DAI Qi-Gen1,*, ZHU Qing2 1Key Laboratory of Crop Genetics and Physiology of Jiangsu Province / Key Laboratory of Crop Cultivation and Physiology of Jiangsu Province / Yangzhou University, Yangzhou 225009, China
2College of Life Science and Chemistry Engineering / Jiangsu Province Engineering Laboratory for Biomass Conversion and Process Integration, Huaiyin Institute of Technology, Huai#cod#x02019;an 223003, China
3Institute of Soil Science / State Key Laboratory of Soil and Sustainable Agriculture, Chinese Academy of Sciences, Nanjing 210008, China
4Daoshu Agricultural Service Center of Danyang City, Jiangsu Province, Danyang 212300, China
Fund: AbstractA soil culture experiment was conducted to study the effects of 1,2,4-trichlorobenzene (TCB) on morphological and physiological indexes of rice at maximum tillering stage, using rice cultivars of Ningjing 1 (TCB sensitive genotype) and Yangfujing 8 (TCB tolerant genotype), with four treatments of TCB concentrations including 0 (CK), 20, 40, and 60 mg kg-1. The results indicated that the maximum root length, plant height, tillers per hill, shoot and root dry weight of Ningjing 1 were all decreased significantly (P<0.05) under different concentrations of TCB. Low TCB stress (20 mg kg-1) significantly increased Yangfujing 8’s maximum root length, shoot and root dry weight, root activity (P<0.01), while medium and high TCB stresses (40 and 60 mg kg-1) decreased maximum root length, plant height, tillers per hill, shoot and root dry weight significantly (P<0.05). Ningjing 1’s root activity, chlorophyll content, soluble protein content in leaves and roots were relatively low under different concentrations of TCB, at the same time, its activities of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) in leaves and roots decreased, while O2- producing velocity and malnodialdehyde (MDA) content increased significantly (P<0.05). Low TCB stress (20 mg kg-1) significantly increased Yangfujing 8’s soluble protein content and activities of SOD, POD, CAT in leaves and roots, and significantly (P<0.05) decreased its O2- producing velocity and MDA content. Under medium TCB stress (40 mg kg-1), the activities of SOD, POD, CAT in leaves and roots were enhanced, however, its O2? producing velocity and MDA content increased significantly. High TCB stress (60 mg kg-1) decreased its root activity, chlorophyll content, the activities of SOD, POD, CAT in leaves and roots, and increased O2- producing velocity and MDA content significantly (P<0.05). In a word, compared with sensitive genotype, well growth vigor, high protein content in leaves and roots, better ability of antioxidant system in scavenging reactive oxygen speciesand lower membrane lipid peroxidation under low concentration TCB stress (20 mg kg-1) are regarded as the main features of TCB tolerant genotype in rice.
Keyword:Rice; Maximum tillering stage; 1; 2; 4-trichlorobenzene; Toxicity; Growth and development; Antioxidant enzymes Show Figures Show Figures
表1 TCB胁迫对2种基因型水稻分蘖盛期各形态指标的影响 Table 1 Effect of TCB on morphological index in the two rice genotypes at maximum tillering stage
基因型 Genotype
TCB浓度 TCB concentration (mg kg-1)
不定根 最长根长 The longest adventitious root length (cm)
不定根 平均根粗 Average adventitious root diameter (mm)
株高 Plant height (cm)
单穴分蘖数 Tillers per hill
单穴地上部 干物重 Shoot dry weight (g per hill)
单穴地下部 干物重 Root dry weight (g per hill)
宁粳1号 Ningjing 1
0 (CK)
16.9 cC
0.83 fE
42.07 bB
10.7 cBC
3.68 cC
0.63 cC
20
15.4 eE
0.85 eD
41.00 cC
9.7 dCD
3.57 dD
0.57 fF
40
14.0 gF
0.91 cB
36.47 eE
8.3 eE
2.78 gG
0.42 gG
60
12.5 hG
0.93 aA
33.37 fF
7.0 fF
2.32 hH
0.36 hH
扬辐粳8号Yangfujing 8
0 (CK)
18.9 bB
0.82 fE
47.37 aA
11.7 bAB
3.92 bB
0.73 bB
20
19.5 aA
0.85 eD
47.67 aA
12.7 aA
3.94 aA
0.75 aA
40
16.3 dD
0.89 dC
41.97 bB
9.7 dCD
3.53 eE
0.62 dD
60
14.2 fF
0.92 bB
38.80 dD
9.0 deDE
3.40 fF
0.57 eE
基因型 F值 F-value of genotype (A)
2171.5**
47.2**
6555.1**
80.7**
594900.0**
48220.0**
浓度 F值 F-value of concentration (B)
1538.0**
907.3**
3483.2**
60.9**
339200.0**
17020.0**
基因型×浓度 F值 F-value of A×B
101.9**
12.9**
20.1**
4.7*
59660.0**
903.6**
同列中标以不同大、小写字母的值分别在0.01和0.05水平差异显著。 Values followed by different letters within a column are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively.
表1 TCB胁迫对2种基因型水稻分蘖盛期各形态指标的影响 Table 1 Effect of TCB on morphological index in the two rice genotypes at maximum tillering stage
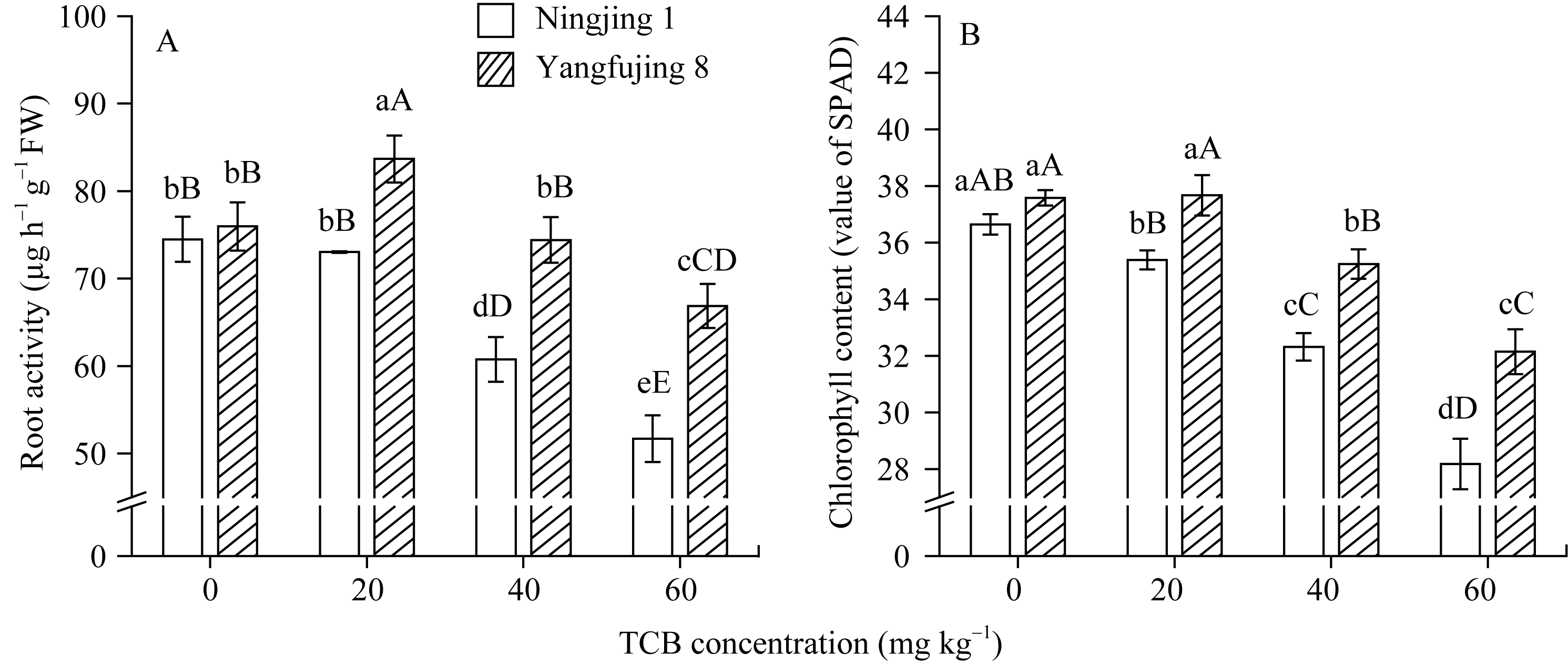
图2 TCB胁迫对分蘖盛期水稻根系活力和叶片叶绿素含量(SPAD值)的影响柱上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 2 Effect of TCB on root activity and chlorophyll content (value of SPAD) of leaves in the two rice genotypes at top tillering stageBars superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively.
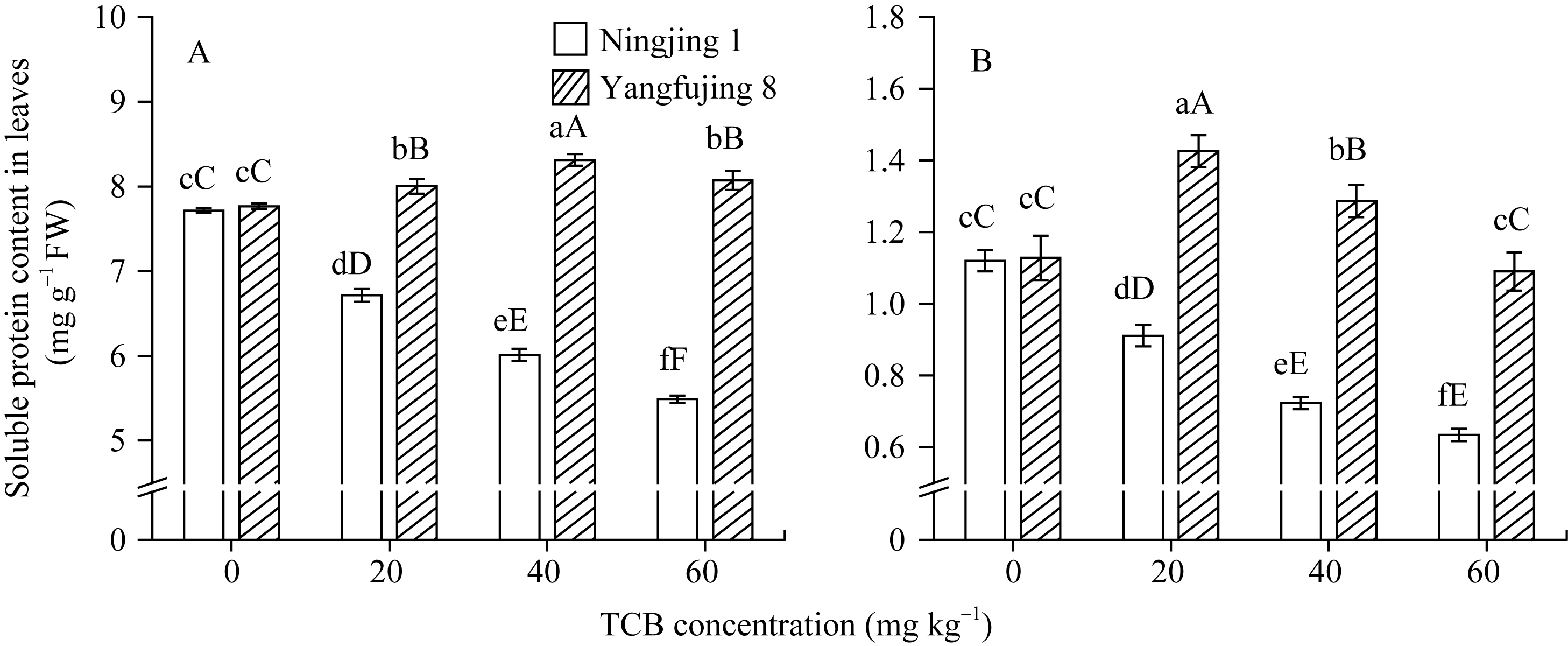
图3 TCB胁迫对分蘖盛期水稻叶片(A)和根系(B)可溶性蛋白质含量的影响柱上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 3 Effect of TCB on the soluble protein content in rice leaves (A) and roots (B) at top tillering stageBars superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively.
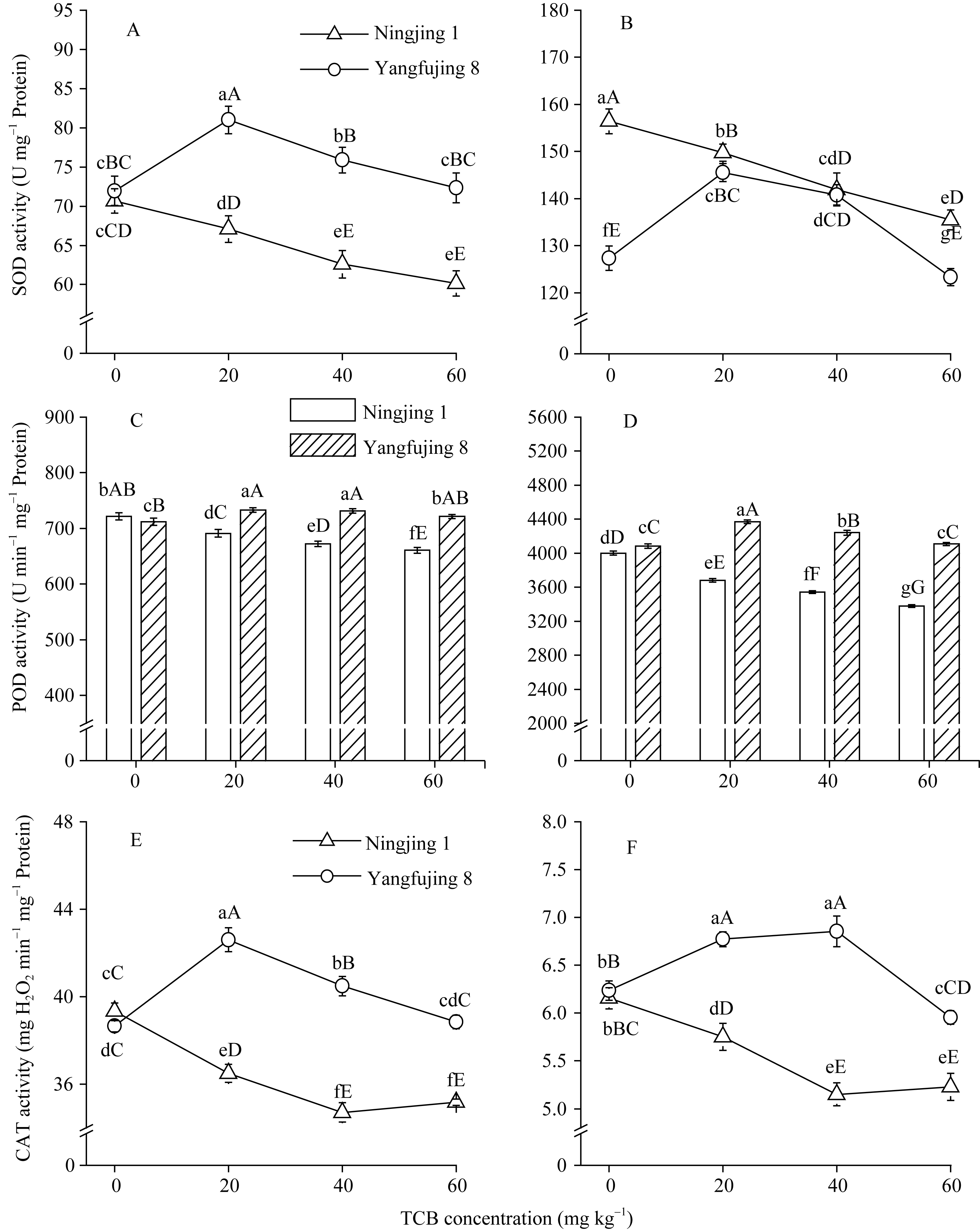
图4 TCB胁迫对分蘖盛期水稻叶片(A, C, E)和根系(B, C, D)抗氧化酶活性的影响柱或曲线上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 4 Effect of TCB on the activity of antioxidant enzymes in rice leaves (A, C, E) and roots (B, C, D) at top tillering stageBars or curves superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively.
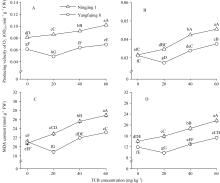
Enzymes activities in the curve with different letters are significantly different at P<0.01 (capital) and P<0.05 (lowercase), respectively. 诱导表达, 导致其受到的毒害作用小于宁粳1号。 TCB对水稻苗期株高[ 24]、地上部生物量[ 24]和成熟期有效分蘖数[ 37]抑制作用显著, 本研究显示TCB对分蘖盛期水稻的地上部也有明显的影响。敏感基因型宁粳1号的株高、地上部干物重、单穴分蘖数在TCB胁迫下均显著低于对照, 而耐性基因型扬辐粳8号在低浓度TCB胁迫下, 株高、单穴分蘖数、地上部干物重与对照相比, 增幅分别为0.6%、8.6%和0.6%, 说明扬辐粳8号对低浓度的TCB胁迫有较强的耐受性, 但当TCB浓度过高时, 对水稻植株的生物毒性加大, 影响了细胞的分裂与生长, 对营养物质的吸收和利用产生干扰, 从而使植株的长势长相异常。 光合作用是绿色植物生长发育的物质基础和能量来源。低温、干旱、盐胁迫、重金属等逆境条件均能影响作物的光合作用, 最终导致减产歉收。王泽港等[ 24, 26]研究发现TCB的添加会导致水稻
当大、小写字母不同时分别表示0.01和0.05差异显著水平。
Values of A, B, C, and D with different letters are significantly different at P<0.01 (capital) and P<0.05 (lowercase), respectively. 幼苗期叶绿素含量、Rubisco酶活性的下降及抽穗期水稻叶片叶绿素含量、净光合速率( Pn)、最大光能转换效率( Fv/ Fm)、光合电子传递的量子化效率( ΦPSII)、光系统PSII的潜在活性( Fv/ Fo)、光化学猝灭系数( qP)及非光化学猝灭系数( qN)呈递减趋势; 杜青平、张波等[ 5, 38]发现水稻和小麦幼苗叶片中叶绿素a、b和叶绿素总含量均随TCB浓度的增加而呈降低趋势。本研究表明, 低浓度TCB使分蘖盛期宁粳1号叶 绿素含量比对照降低3.4%, 而对扬辐粳8号却增加0.2%。中高浓度的TCB胁迫使水稻光合器官受到伤害[ 26], 对两品种分蘖盛期的叶绿素含量均有极显著的抑制作用, 从而引起光合作用减弱, 干物重降低。 蛋白质是植物生长发育、新陈代谢过程中的重要物质。已有研究表明TCB能诱导水稻和小麦幼苗蛋白质的表达[ 23, 39], 主要包括解毒酶、细胞壁物质、激素、原初或次生代谢与调节等的相关酶, 进而导致植株体内蛋白质含量的增加[ 25]。本研究显示, 不同浓度TCB胁迫下, 分蘖盛期宁粳1号叶片和根系可溶性蛋白质分别与各自对照相比, 叶片的降幅(13.0%、22.1%、28.8%)远小于根系(18.7%、35.4%、43.4%); 扬辐粳8号叶片可溶性蛋白质的表达受到了诱导, 增幅分别为3.1%、7.1%、3.9%。中低浓度TCB胁迫下, 扬辐粳8号根系可溶性蛋白质含量的增幅分别为14.1%、26.4%; 而高浓度胁迫时, 根系可溶性蛋白质含量与对照相比差异不显著。说明扬辐粳8号对TCB胁迫的耐性强, 中低浓度能够充分诱导适应逆境相关蛋白及酶的表达, 而宁粳1号耐性较差, 蛋白合成受阻或者分解加快, 导致蛋白质含量较低, 对有机污染胁迫适应性差。 超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等是植物体内清除活性氧的重要抗氧化酶, 也是酶保护系统的重要组分。酶保护系统启动后, 能及时清除一定数量过剩的活性氧和过氧化物, 从而减轻过氧化作用的伤害, 提高细胞的抗逆性。丙二醛(MDA)含量反映了细胞膜脂过氧化程度及器官的衰老情况。高浓度TCB胁迫下, 水稻敏感品种幼苗的MDA含量显著增加, SOD、POD、CAT活性有所降低, 根系受到的伤害明显大于叶片[ 5, 25]。张国良等[ 22]认为小麦敏感品种根系的SOD、POD活性在低浓度TCB胁迫下有所升高, 而CAT活性却显著降低。刘宛等[ 20, 21]发现TCB能够使萌发的大豆种子活性氧代谢紊乱, 超氧阴离子自由基( )产生速率显著增加, 丙二醛大量积累; 随着胁迫时间的延长, 大豆种子POD活性持续升高, SOD和CAT活性均表现为先升高后降低趋势。本研究结果表明, 分蘖盛期宁粳1号和扬辐粳8号对TCB胁迫的酶学响应有明显的基因型差异。TCB胁迫下, 宁粳1号3种抗氧化酶活性有所降低, 导致其酶保护系统清除活性氧的能力下降, 最终导致MDA过量积累, 膜脂过氧化加剧。扬辐粳8号在低浓度TCB胁迫下可以通过极显著提高3种酶的活性增强其抗氧化的能力, 进而促进其生长; 而中等浓度TCB胁迫时, 虽然3种酶活性均显著高于对照, 但 产生速率仍显著增高, 也导致MDA大量积累, 说明扬辐粳8号对胁迫的自我调节能力有一定限度, 当TCB浓度超过一定界限时, 虽然抗氧化能力与对照相比有所增强, 但不足以弥补TCB胁迫对生长造成的伤害; 而高浓度胁迫时, 叶片中3种酶活性变化不明显, 而根系中SOD、CAT活性显著降低, 活性氧大量积累, 膜脂过氧化加剧, 植株代谢紊乱, 生长明显受抑。 低浓度TCB胁迫对扬辐粳8号的生长发育有明显的促进作用, 可能主要有以下三方面原因。(1)植株的解毒系统适应性强, 抗氧化酶活性有所提高、谷胱甘肽-S-转移酶和顺式还原酮加双氧酶(ARD)受到诱导表达[ 36], 有利于清除活性氧和有毒物质、维持细胞结构的稳定, 进而增强植株的抗逆能力; (2) MicroRNA是一类与植物适应逆境胁迫有关的调控基因表达的小分子RNA[ 40], CB等有机污染胁迫可能诱导或下调水稻相关MicroRNA的表达, 从而调节植株的代谢过程; (3) 低浓度TCB能增加以其为碳源的微生物数量, 提高土壤肥力[ 41, 42], 而扬辐粳8号对TCB胁迫有一定耐受限度, 在低浓度TCB下仍能正常吸收与利用养分。 4 结论不同浓度TCB胁迫下敏感基因型和耐性基因型水稻的生长和生理特性差异显著, 敏感基因型随胁迫浓度的增加生长和生理活性受抑程度显著递增; 而耐性基因型在低浓度TCB胁迫下, 植株解毒能力 较强, 最长根长、地上部和地下部干物重均极显著高于对照, 根系活力、叶片和根系可溶性蛋白含量较高, 酶保护系统清除活性氧的能力较强, 膜脂过氧化程度较低。中高浓度TCB胁迫才对耐性基因型水稻的生长表现出明显的抑制作用。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
MiguelA S, FaureM, RavanelP, RavetonM. Biological responses of maize (Zea mays) plants exposed to chlorobenzenes. Case study of monochloro-1, 4-dichloro- and 1, 2, 4-trichloro- benzenes. Ecotoxicology, 2012, 21: 315-324[本文引用:1][JCR: 2.773]
[2]
周文敏, 傅德黔, 孙宗光. 中国水中优先控制污染物黑名单的确定. 环境科学研究, 1991, 4(6): 9-12ZhouW M, FuD Q, SunZ G. Determination of black list of China’s priority pollutants in water. Res Environ Sci, 1991, 4(6): 9-12 (in Chinese with English abstract)[本文引用:1][CJCR: 1.295]
[3]
OliverB G, NiimiA J. Bioconcentration of chlorobenzenes from water by rainbow trout: correlations with partition coefficients and environmental residues. Environ Sci Technol, 1983, 17: 287-291[本文引用:1][JCR: 5.228]
[4]
魏泰莉, 贾晓珊, 杜青平, 赖子尼, 庞世勋, 谢文平, 杨婉玲. 珠江口水、沉积物及水生动物中氯苯类有机物的含量及分布. 环境科学学报, 2007, 27: 1717-1726WeiT L, JiaX S, DuQ P, LaiZ N, PangS X, XieW P, YangW L. Chlorobenzenes in the waters, sediments and aquatic animals from the Pearl River Estuary. Acta Sci Circumstantiae, 2007, 27: 1717-1726 (in Chinese with English abstract)[本文引用:3][CJCR: 1.689]
[5]
杜青平, 贾晓珊, 袁保红. 1, 2, 4-三氯苯对水稻种子萌发及幼苗生长的毒性机理. 应用生态学报, 2006, 17: 2185-2188DuQ P, JiaX S, YuanB H. Toxic effects of 1, 2, 4-trichlorobenzene on rice seed germination and seedling growth. Chin J Appl Ecol, 2006, 17: 2185-2188 (in Chinese with English abstract)[本文引用:5][CJCR: 1.742]
[6]
宋洋, 王芳, 蒋新. 微生物降解1, 2, 4-三氯苯研究进展. 土壤, 2011, 43: 343-349SongY, WangF, JiangX. Microbial degradation of 1, 2, 4-trichlorobenzene: a review. Soils, 2011, 43: 343-349 (in Chinese with English abstract)[本文引用:1][CJCR: 1.507]
[7]
周霞, 余刚, 黄俊, 张祖麟, 胡洪营. 北京东南郊化工区土壤和植物中氯苯类有机物的残留及分布特征. 环境科学, 2007, 28: 249-254ZhouX, YuG, HuangJ, ZhangZ L, HuH Y. Residues and distribution characters of chlorobenzenes in soil and plants from Beijing Southeast Chemical Industry Zone. Environ Sci, 2007, 28: 249-254 (in Chinese with English abstract)[本文引用:2][CJCR: 1.541]
[8]
崔健, 都基众, 杨泽, 马宏伟, 李霄. 沈阳市城郊土壤有机污染特征. 生态学杂志, 2011, 30: 2472-2477CuiJ, DuJ Z, YangZ, MaH W, LiX. Characteristics of soil organic pollution in Shenyang suburbs of Liaoning Province. Chin J Ecol, 2011, 30: 2472-2477 (in Chinese with English abstract)[本文引用:1][CJCR: 1.729]
[9]
蔡全英, 莫测辉, 吴启堂, 李桂荣. 微生物方法降低城市污泥的重金属含量研究进展. 环境化学, 2002, 21(2): 139-143CaiQ Y, MoC H, WuQ T, LiG R. Preliminary study on the content of chlorobenzenes in selected municipal sludge of China. Environ Chem, 2002, 21(2): 139-143 (in Chinese with English abstract)[本文引用:1][JCR: 2.652]
[10]
刘玉萍. 松花江水体氯苯类污染物的污染研究. 环境科学与管理, 2006, 31(5): 91-92LiuY P. Study of Chlorobenzene’s pollution in the Songhua River. Environ Sci Manag, 2006, 31(5): 91-92 (in Chinese with English abstract)[本文引用:1][CJCR: 0.4805]
[11]
朱明吉, 郭志顺. 三峡库区重庆段有机污染研究. 三峡环境与生态, 2009, 2(2): 5-11ZhuM J, GuoZ S. Research on organic pollution in Chongqing section of the Three Gorges Reservoir. Environ Ecol Three Gorges, 2009, 2(2): 5-11 (in Chinese with English abstract)[本文引用:1]
[12]
石建省, 王昭, 张兆吉, 费宇红, 李亚松, 张凤娥, 陈京生, 钱永. 华北平原地下水有机污染特征初步分析. 生态环境学报, 2011, 20: 1695-1699ShiJ S, WangZ, ZhangZ J, FeiY H, LiY S, ZhangF E, ChenJ S, QianY. Preliminary analysis on the organic contamination of groundwater in the North China Plain. Ecol Environ Sci, 2011, 20: 1695-1699 (in Chinese with English abstract)[本文引用:1]
[13]
LeeC L, SongH J, FanM D. Concentrations of chlorobenzenes, hexachlorobutadiene and heavy metals in surficial sediments of Kaohsiung coast, Taiwan. Chemosphere, 2000, 41: 889-899[本文引用:1][JCR: 3.137]
[14]
ZhangJ Y, ZhaoW, PanJ, QiuL M, ZhuY M. Tissue-dependent distribution and accumulation of chlorobenzenes by vegetables in urban area. Environ Int, 2005, 31: 855-860[本文引用:1][JCR: 6.248]
[15]
JaimeD, ManuelR, MarioD. 1, 2, 4-trichlorobenzene flow characteristics in saturated homogeneous and stratified porous media. Water Air Soil Poll, 2006, 177: 3-17[本文引用:1][JCR: 1.748]
[16]
GB5749/2006, 生活饮用水卫生标准. 北京: 中华人民共和国卫生部/中国国家标准化管理委员会, 2006GB5749/2006, Stand ard for Drinking Water Quality. Beijing: Ministry of Public Health of the People’s Republic of China/ Stand ardization Administration of the People’s Republic of China, 2006 (in Chinese)[本文引用:1]
[17]
DB11/307-2005, 水污染物排放标准. 北京: 北京市环境保护局/北京市质量技术监督局, 2005DB11/307-2005, Discharge Stand ard of Water Pollutants. Beijing: Beijing Municipal Environmental Protection Bureau/Beijing Municipal Administration of Quality and Technology Supervision, 2005 (in Chinese)[本文引用:1]
[18]
杨继富. 污水灌溉农业问题与对策. 水资源保护, 2000, (2): 4-8YangJ F. The problem on agricultural sewage irrigation and countermeasures. Water Resour Prot, 2000, (2): 4-8 (in Chinese with English abstract)[本文引用:1]
[19]
MacleodM, MackayD. An assessment of the environmental fate and exposure of benzene and the ehlorobenzenes in Canada. Chemosphere, 1999, 38: 1777-1796[本文引用:1][JCR: 3.137]
[20]
刘宛, 孙铁珩, 李培军, 周启星, 梁文举, 台培东, 许华夏, 张海荣. 1, 2, 4-三氯苯胁迫对萌发大豆种子中活性氧的影响. 应用生态学报, 2002, 13: 1655-1658LiuW, SunT H, LiP J, ZhouQ X, LiangW J, TaiP D, XuH X, ZhangH R. Effects of 1, 2, 4-trichlorobenzene stress on active oxygen in germinated soybean seeds. Chin J Appl Ecol, 2002, 13: 1655-1658 (in Chinese with English abstract)[本文引用:2][CJCR: 1.742]
[21]
刘宛, 孙铁珩, 李培军, 周启星, 台培东, 张春桂, 许华夏, 张海荣. 1, 2, 4-三氯苯胁迫对大豆下胚轴膜脂过氧化的影响. 农业环境保护, 2002, 21: 413-416LiuW, SunT H, LiP J, ZhouQ X, TaiP D, ZhangC G, XuH X, ZhangH R. Effects of 1, 2, 4-trichlorobenzene stress on membrane lipid peroxidation in soybean hypocotyls. J Agro-Environ Prot, 2002, 21: 413-416 (in Chinese with English abstract)[本文引用:2]
[22]
张国良, 陈文军, 王显, 金添, 戴其根, 孙国荣, 许轲, 霍中洋, 张洪程. 小麦苗期对1, 2, 4-三氯苯胁迫的生理响应. 生态学报, 2008, 28: 4388-4395ZhangG L, ChenW J, WangX, JinT, DaiQ G, SunG R, XuC, HuoZ Y, ZhangH C. Physiological reaction of wheat seedling to 1, 2, 4-trichlorobenzene stress. Acta Ecol Sin, 2008, 28: 4388-4395 (in Chinese with English abstract)[本文引用:2]
[23]
丁艳, 葛才林, 王泽港, 杜庆才. 小麦幼苗对镉和1, 2, 4-三氯苯污染的响应. 中国农业大学学报, 2011, 16: 48-52DingY, GeC L, WangZ G, DuQ C. Response of Cd and 1, 2, 4-trichlorobenzene pollutants on growth of wheat seedlings. J China Agric Univ, 2011, 16: 48-52 (in Chinese with English abstract)[本文引用:2]
[24]
王泽港, 葛才林, 万定珍, 郦志文, 罗时石, 杨建昌. 1, 2, 4-三氯苯和萘对水稻幼苗生长的影响. 农业环境科学学报, 2006, 25: 1402-1407WangZ G, GeC L, WanD Z, LiZ W, LuoS S, YangJ C. Effects of 1, 2, 4-trichlorobenzene and naphthalene on growth of rice seedling. J Agro-Environ Sci, 2006, 25: 1402-1407 (in Chinese with English abstract)[本文引用:5][CJCR: 1.178]
[25]
张国良, 陈文军, 仇利民, 孙国荣, 戴其根, 张洪程. 不同基因型水稻苗期对1, 2, 4-三氯苯胁迫的生理响应. 作物学报, 2009, 35: 733-740ZhangG L, ChenW J, QiuL M, SunG R, DaiQ G, ZhangH C. Physiological response to 1, 2, 4-trichlorobenzene stress of different rice genotypes in seedlings. Acta Agron Sin, 2009, 35: 733-740 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[26]
王泽港, 骆剑峰, 高红明, 万定珍, 葛才林, 罗时石, 杨建昌. 1, 2, 4-三氯苯和萘对水稻抽穗期叶片光合特性的影响. 中国农业科学, 2005, 38: 1113-1119WangZ G, LuoJ F, GaoH M, WanD Z, GeC L, LuoS S, YangJ C. Effects of 1, 2, 4-trichlorobenzene and naphthalene stress on photosynthetic characteristics of rice at heading period. Sci Agric Sin, 2005, 38: 1113-1119 (in Chinese with English abstract)[本文引用:3][CJCR: 1.889]
[27]
陈文军, 张国良, 孙国荣, 戴其根, 张洪程, 陶金飞, 孙洁, 严林锋. 水稻耐1, 2, 4-三氯苯胁迫基因型的苗期筛选. 农业环境科学学报, 2008, 27: 1003-1008ChenW J, ZhangG L, SunG R, DaiQ G, ZhangH C, TaoJ F, SunJ, YanL F. Screening of tolerant rice genotypes to 1, 2, 4- trichlorobenzene stress at seedling stage. J Agro-Environ Sci, 2008, 27: 1003-1008 (in Chinese with English abstract)[本文引用:2][CJCR: 1.178]
邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2000. pp173-174ZouQ. The Guidance of Plant Physiological Experiment. Beijing: China Agriculture Press, 2000. pp173-174(in Chinese)[本文引用:1]
[30]
高俊凤. 植物生理学实验指导. 北京: 高等教育出版社, 2006. pp215-216GaoJ F. The Guidance of Plant Physiological Experiment. Beijing: Higher Education Press, 2006. pp215-216(in Chinese)[本文引用:1]
[31]
郝建军. 植物生理学实验技术. 北京: 化学工业出版社, 2007. pp159-160HaoJ J. Experimental technique of Plant Physiological. Beijing: Chemical Industry Press, 2007. pp159-160(in Chinese)[本文引用:1]
[32]
李忠光, 龚明. 植物中超氧阴离子自由基测定方法的改进. 云南植物研究, 2005, 27: 211-216LiZ G, GongM. Improvement of measurement method for superoxide anion radical in plant. Acta Bot Yunnanica, 2005, 27: 211-216 (in Chinese with English abstract)[本文引用:1]
[33]
WangM J, JonesK C. Behavior and fate of chlorobenzenes (CBs) introduced into soil-plant systems by sewage sludge application: a review. Chemosphere, 1994, 28: 1325-1360[本文引用:1][JCR: 3.137]
[34]
HeY W, SunT H, OuZ Q, YedilerAKettrupA. Fate of 1, 2, 4-trichlorobenzene (1, 2, 4-TCB) in soil-rice paddy system. Chemosphere, 1996, 32: 1381-1389[本文引用:1][JCR: 3.137]
[35]
于开源, 苏玉红, 刘涛, 梁永超. 三氯乙烯和三氯苯在水-玉米- 空气系统中的传输过程研究. 农业环境科学学报, 2010, 29: 1702-1705YuK Y, SuY H, LiuT, LiangY C. Transport processes of trichloroethylene and trichlorobenzene across the water-corn-air interface. J Agro-Environ Sci, 2010, 29: 1702-1705 (in Chinese with English abstract)[本文引用:1][CJCR: 1.178]
[36]
GeC L, WanD Z, WangZ G, DingY, WangY L, ShangQ, MaF, LuoS S. Response of rice roots to 1, 2, 4-trichlorobenzene stress. Acta Agron Sin, 2007, 33: 1991-2000[本文引用:3][CJCR: 1.667]
[37]
金阳. 1, 2, 4-三氯苯对水稻伤害机理及水稻耐性机制的研究. 扬州大学硕士学位论文, 2003. pp20-25JinY. Study on the Mechanisms of l, 2, 4-Triehlorobenzene Damaging Rice. MS Thesis of Yangzhou University, Yangzhou, China, 2003 (in Chinese with English abstract)[本文引用:1]
[38]
张波. 小麦对氧乐果和1, 2, 4-三氯苯响应的生物学机理. 石河子大学硕士学位论文, 2010. pp18-22ZhangB. Physiological Responses of Wheat Seedlings to Omethoate and 1, 2, 4-trichlorobenzene Stress. MS Thesis of Shihezi University, Shihezi, China, 2010 (in Chinese with English abstract)[本文引用:1]
[39]
GeC L, WanD Z, WangZ G, DingY, WangY L, ShangQ, MaF, LuoS S. A proteomic analysis of rice seedings responding to 1, 2, 4-trichlorobenzene stress. J Environ Sci, 2008, 10: 309-319[本文引用:1]
何耀武, 孙铁琦, 区自清. 1, 2, 4-三氯苯在土壤中的降解. 应用生态学报, 1996, 7: 429-434HeY W, SunT H, OuZ Q. Degradation of 1, 2, 4-trichlorobenzene (1, 2, 4-TCB) in soil. Chin J Appl Ecol, 1996, 7: 429-434 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[42]
李勇, 黄占斌, 王文萍, 黄震, 颜丙磊, 曹杨, 王诗宇. 重金属铅镉对玉米生长及土壤微生物的影响. 农业环境科学学报, 2009, 28: 2241-2245LiY, HuangZ B, WangW P, HuangZ, YanB L, CaoY, WangS Y. Effects of heavy metals lead and cadmium on Zea mays L. growth and the soil microorganism. J Agro-Environ Sci, 2009, 28: 2241-2245 (in Chinese with English abstract)[本文引用:1][CJCR: 1.178]




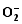
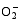
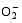
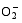
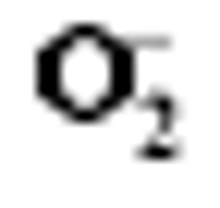
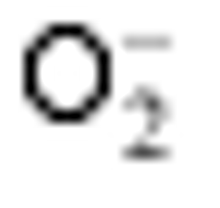

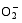

{kind=link}
{kind=link}
{kind=link}
{kind=link}