关键词:花生品种; 干旱胁迫; 根系特性; 抗旱性 Morphological and Physiological Traits of Root in Different Drought Resistant Peanut Cultivars LI Guang-Hui, WAN Yong-Shan*, LIU Feng-Zhen, ZHANG Kun National Key Laboratory of Crop Biology / College of Agronomy, Shandong Agricultural University, Tai#cod#x02019;an 271018, China Fund: AbstractDrought stress is a serious constraint for peanut production worldwide. It is necessary to identify the drought resistance mechanisms of different peanut cultivars in drought-resistance breeding. Under the artificial water control condition, the peanut root morphological and physiological characteristics under drought stress at seedling stage and pod-setting stage were studied using 12 different drought-resistance peanut cultivars as material. The results showed that drought resistance at seedling stage was basically identical with that at pod-setting stage. According to yield-drought resistance coefficient, 12 peanut cultivars were divided into three grades: high-resistance, including A596, Shanhua 11, and Rugaoxiyangsheng; mid-resistance, including Huayu 20, Nongda 818, Haihua 1, Shanhua 9, and 79266; and weak-resistance, including ICG6848, Baisha 1016, Hua 17, and Penglaiyiwohou. In those peanut cultivars, Shanhua11 can be used as the standard cultivar for high drought resistance identification, and 79266 as the standard cultivar for weak one. The root drought resistance mechanism of 12 peanut cultivars were different, Shanhua 9, Shanhua11 and Huayu 20 presented a larger biomass and strong absorption capacity, while A596, Nongda 818, Shanhua 11, and Rugaoxiyangsheng had strong antioxidant capacity and membrane stability under drought stress. Correlations between drought resistance and root weight, volume, total absorption area per plant, root superoxide dismutase (SOD) activity, malondialdehyde (MDA) content under serious drought stress were significant, also under control condition. Therefore, under drought stress of 40% RWC for 10 to 24 days after germination, the root weight, volume, total absorption area per plant, SOD activity and MDA content could be used for identifies the drought resistance ability of peanut roots, the resistances degree also can be reflected by the indices above under normal water condition. Shanhua 11 can be used as a suitable standard cultivar for root morphological and physiological drought resistance characteristics identification in peanut.
Keyword:Peanut cultivars; Drought stress; Root traits; Drought resistance Show Figures Show Figures
表2 苗期及结荚期干旱胁迫下不同花生品种的抗旱性 Table 2 Drought resistance of peanut cultivars under drought stress at seedling and pod-setting stages
品种 Cultivar
苗期生物量及抗旱性 Drought resistance and biomass at seedling stage
结荚期产量及抗旱性 Drought resistance and yield at pod-setting stage
对照 Control (g plant-1)
轻度干旱 MD (g plant-1)
重度干旱 SD (g plant-1)
轻度干旱 抗旱系数 DC of MD
重度干旱 抗旱系数 DC of SD
对照 Control (kg hm-2)
干旱胁迫 Drought (kg hm-2)
抗旱 系数 DC
抗旱 指数 DI
2011
79266
3.12 d
2.55 ef
1.99 e
0.82 def
0.64 def
4291 c
3452 b
0.80 efg
0.82 de
A596
3.68 c
3.33 abc
2.81 abc
0.91 bc
0.76 ab
3621 ef
3429 b
0.95 ab
0.96 c
ICG6848
3.86 bc
3.27 bcd
2.23 de
0.85 cd
0.58 f
3299 fg
2518 e
0.76 gh
0.57 g
白沙1016 Baisha 1016
3.15 d
2.44 f
1.85 e
0.77 f
0.59 f
3765 de
2751 de
0.73 h
0.59 g
海花1号 Haihua 1
3.58 c
3.04 bcd
2.56 bcd
0.85 cd
0.71 bc
4908 a
4163 a
0.85 def
1.04 bc
花17 Hua 17
4.25 a
3.70 a
2.72 abc
0.87 bcd
0.64 def
4119 cd
3023 cd
0.73 h
0.66 fg
花育20 Huayu 20
4.30 a
3.41 ab
2.90 ab
0.79 ef
0.67 cde
3878 de
3191 bc
0.82 efg
0.78 ef
农大818 Nongda 818
3.54 c
2.90 de
2.49 cd
0.82 def
0.70 bcd
4436 bc
3965 a
0.89 bcd
1.05 bc
蓬莱一窝猴 Penglaiyiwohou
3.12 d
2.92 cde
1.93 e
0.93 ab
0.62 ef
3178 g
2494 e
0.79 fgh
0.58 g
如皋西洋生 Rugaoxiyangsheng
3.71 c
3.69 a
3.05 a
0.99 a
0.82 a
3089 g
2997 cd
0.97 a
0.86 d
山花11 Shanhua 11
3.73 c
3.34 abc
3.01 a
0.89 bcd
0.81 a
4682 ab
4305 a
0.92 abc
1.17 a
山花9号 Shanhua 9
4.15 ab
3.49 ab
2.96 a
0.84 cde
0.71 bc
4979 a
4322 a
0.87 cde
1.11 ab
2012
79266
3.59 d
3.13 ef
2.25 d
0.87 bcd
0.63 d
4173 d
3620 cd
0.87 de
0.88 cd
A596
4.28 b
3.99 bc
3.16 abc
0.93 ab
0.74 b
3818 ef
3684 cd
0.97 ab
1.01 bc
ICG6848
4.13 bc
3.65 cd
2.07 d
0.88 bc
0.50 f
2979 h
2267 h
0.76 g
0.49 f
白沙1016 Baisha 1016
3.49 d
2.91 f
1.87 d
0.83 d
0.53 ef
3749 f
2978 fg
0.79 fg
0.67 e
海花1号 Haihua 1
4.23 bc
3.85 c
2.83 bc
0.91 abc
0.67 c
4767 ab
4190 ab
0.88 de
1.04 b
花17 Hua 17
5.15 a
4.44 a
3.21 ab
0.86 cd
0.62 d
4089 de
3359 de
0.82 ef
0.78 de
花育20 Huayu 20
4.96 a
4.28 ab
3.27 a
0.86 cd
0.66 cd
3822 ef
3599 cde
0.94 abc
0.95 c
农大818 Nongda 818
4.11 bc
3.62 cd
2.80 c
0.88 bc
0.68 c
4268 cd
3908 bc
0.92 bcd
1.01 bc
蓬莱一窝猴 Penglaiyiwohou
3.63 d
3.38 de
2.01 d
0.93 ab
0.55 e
3378 g
2801 g
0.83 ef
0.65 e
如皋西洋生 Rugaoxiyangsheng
3.83 cd
3.73 cd
2.91 abc
0.97 a
0.76 ab
3290 g
3245 ef
0.99 a
0.90 cd
山花11 Shanhua 11
4.14 bc
3.78 cd
3.29 a
0.91 abc
0.79 a
4548 bc
4475 a
0.98 a
1.24 a
山花9号 Shanhua 9
4.25 bc
3.80 cd
2.96 abc
0.89 bc
0.70 c
5045 a
4508 a
0.89 cd
1.13 ab
MD: 中度干旱; SD: 重度干旱; DC: 抗旱系数; DI: 抗旱指数。同一列内标有相同小写字母的值差异未达到5%显著水平。 MD: mild drought; SD: serious drought; DC: drought resistance coefficient; DI: drought resistance index. Values followed by different letters are significantly different at the 5% probability level.
表2 苗期及结荚期干旱胁迫下不同花生品种的抗旱性 Table 2 Drought resistance of peanut cultivars under drought stress at seedling and pod-setting stages
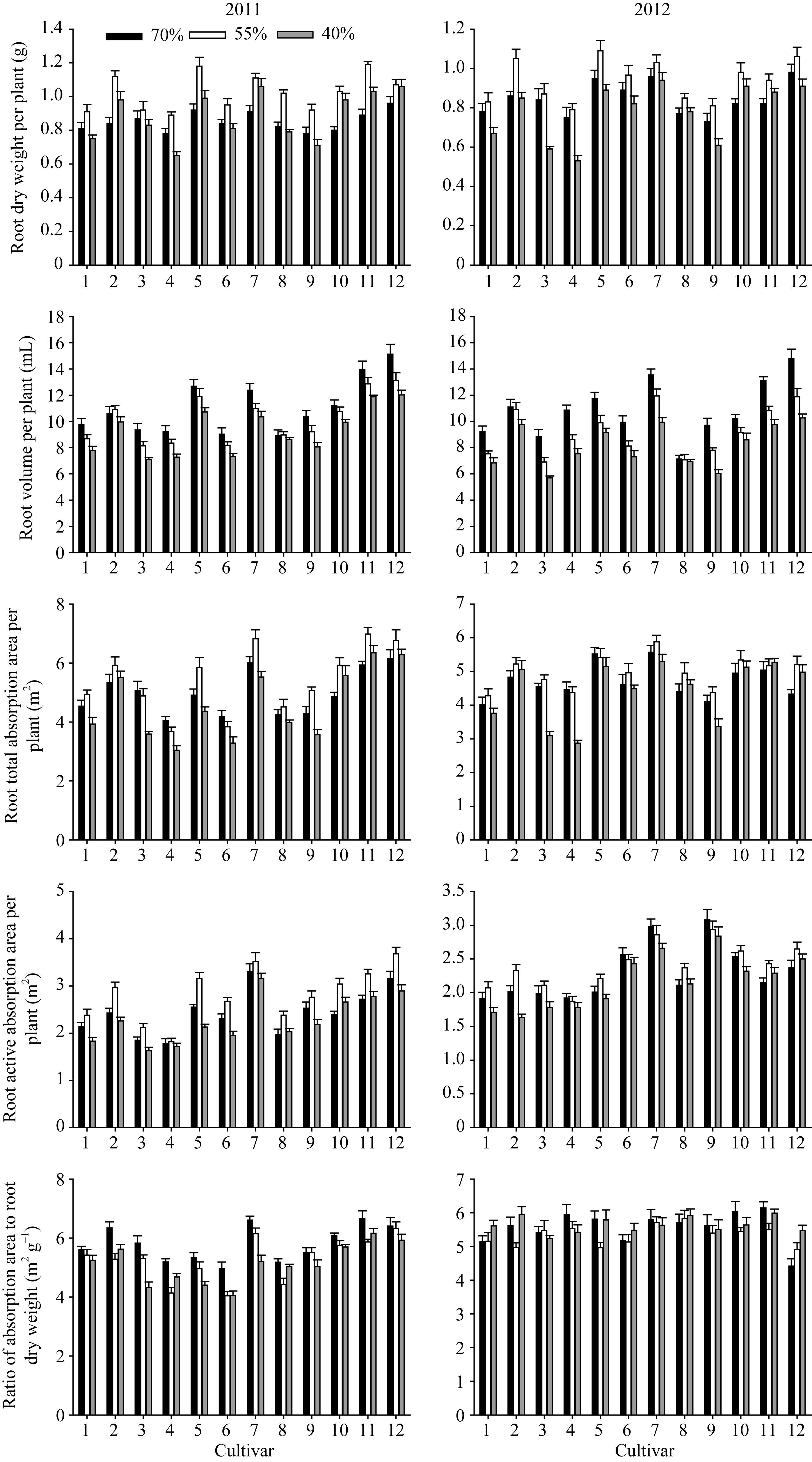
图1 干旱胁迫下不同花生品种苗期根系形态指标品种编号1~12分别代表79266、A596、ICG6848、白沙1016、海花1号、花17、花育20、农大818、蓬莱一窝猴、如皋西洋生、山花11和山花9号。Fig. 1 Roots morphological traits of different peanut cultivars under drought stress at seedling stageThe number of cultivars from 1 to 12 represented 79266, A596, ICG6848, Baisha 1016, Haihua 1, Hua 17, Huayu 20, Nongda 818, Penglaiyiwohou, Rugaoxiyangsheng, Shanhua 11, and Shanhua 9, respectively.
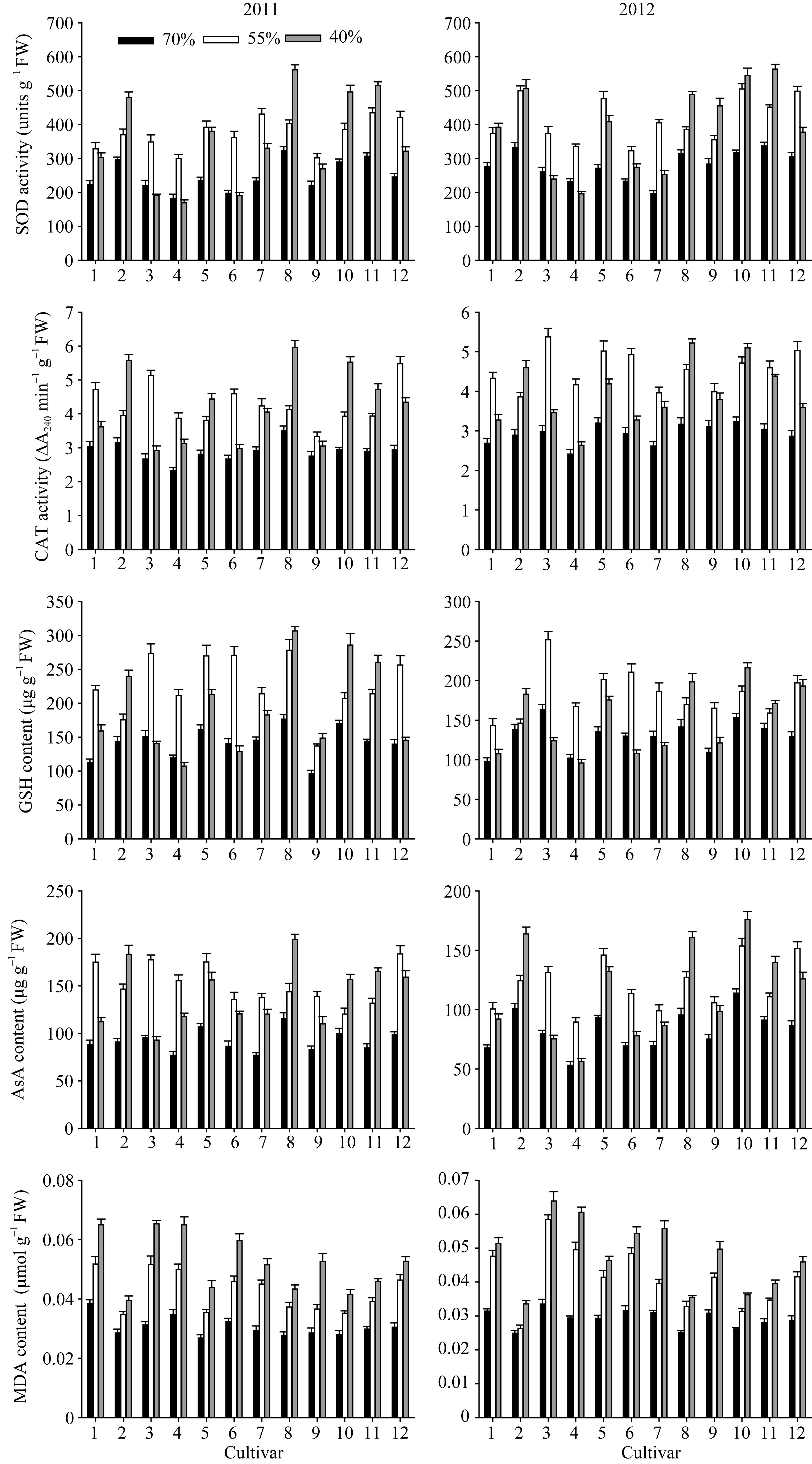
图2 干旱胁迫下不同花生品种苗期根系抗氧化保护相关生理性状品种编号1~12分别代表79266、A596、ICG6848、白沙1016、海花1号、花17、花育20、农大818、蓬莱一窝猴、如皋西洋生、山花11和山花9号。Fig. 2 Roots physiological traits related to antioxidant system of peanut cultivars under drought stress at seedling stageThe number of cultivars from 1 to 12 represented 79266, A596, ICG6848, Baisha 1016, Haihua 1, Hua 17, Huayu 20, Nongda 818, Penglaiyiwohou, Rugaoxiyangsheng, Shanhua 11, and Shanhua 9, respectively.
表3 Table 3 表3(Table 3)
表3 苗期花生根系形态及生理性状与品种抗旱性的相关系数 Table 3 Correlation coefficients of root traits of peanut cultivars with drought resistance
年份 Year
土壤相对含水量 RWC (%)
干重 Dry weight
体积 Volume
总吸收 面积 TAA
比吸收 面积 TAA/DR
活跃吸收 面积 AAA
SOD活性 SOD activity
CAT活性 CAT activity
GSH含量 GSH content
AsA含量 AsA content
MDA含量 MDA content
2011
70
0.19
0.43
0.38
0.52
0.27
0.89**
0.65*
0.46
0.46
-0.58*
55
0.62*
0.63*
0.59*
0.45
0.41
0.49
-0.25
-0.32
-0.24
-0.76**
40
0.57*
0.69*
0.75**
0.75**
0.59*
0.91**
0.88**
0.79**
0.84**
-0.87**
2012
70
0.21
0.33
0.46
0.35
0.58*
0.54
0.31
0.27
0.70*
-0.68*
55
0.61*
0.58*
0.63*
0.04
0.65*
0.73**
-0.32
-0.49
0.28
-0.90**
40
0.78**
0.71**
0.85**
0.63*
0.76**
0.75**
0.60*
0.61*
0.67*
-0.84**
表3 苗期花生根系形态及生理性状与品种抗旱性的相关系数 Table 3 Correlation coefficients of root traits of peanut cultivars with drought resistance
TAA: 总吸收面积; TAA/DR: 比吸收面积; AAA: 活跃吸收面积。*表示 P< 0.05的显著水平,**表示 P< 0.01的显著水平。 TAA: total absorption area; TAA/DR: ratio of total absorption area to root dry weight; AAA: active absorption area.* Significant at P< 0.05;** significant at P< 0.01. 表4 Table 4 表4(Table 4)
表4 苗期不同干旱处理下根系性状间的相关系数 Table 4 Correlation coefficients of root traits under drought stress and control
年份 Year
土壤相对含水量 RWC (%)
干重 Dry weight
体积 Volume
总吸收 面积 TAA
比吸收 面积 TAA/DR
活跃吸收面积 AAA
SOD活性 SOD activity
CAT活性 CAT activity
GSH含量 GSH content
AsA含量 AsA content
MDA含量 MDA content
2011
55
0.67*
0.93**
0.87**
0.52
0.95**
0.61*
0.03
0.56
0.61*
0.81**
40
0.79**
0.95**
0.92**
0.67*
0.91**
0.97**
0.69*
0.61*
0.70*
0.85**
2012
55
0.71*
0.84**
0.66*
0.43
0.91**
0.46
0.41
0.59
0.76*
0.86**
40
0.89**
0.93**
0.86**
0.56
0.92**
0.81**
0.76*
0.58
0.95**
0.91**
表4 苗期不同干旱处理下根系性状间的相关系数 Table 4 Correlation coefficients of root traits under drought stress and control
姜慧芳, 任小平. 干旱胁迫对花生叶片SOD活性和蛋白质的影响. 作物学报, 2004, 30: 169-174JiangH F, RenX P. The effect on SOD activity and protein content in groundnut leaves by drought stress. Acta Agron Sin, 2004, 30: 169-174 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[2]
严美玲, 李向东, 林英杰, 王丽丽, 周录英. 苗期干旱胁迫对不同抗旱花生品种生理特性、产量和品质的影响. 作物学报, 2007, 33: 113-119YanM L, LiX D, LinY J, WangL L, ZhouL Y. Effects of drought during seedling stage on physiological traits, yield and quality of different peanut cultivars. Acta Agron Sin, 2007, 33: 113-119 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[3]
KumarA, SinghP, SinghD P, SinghH, SharmaH C. Differences in osmotic regulation in Brassica species. Ann Bot, 1984, 54: 537-541[本文引用:1][JCR: 0.657]
[4]
UpadhyayaH D. Variability for drought resistance related traits in the mini core collection of peanut. Crop Sci, 2005, 45: 1432-1440[本文引用:1][JCR: 1.513]
[5]
KamoshitaA, BabuR C, BoopathiN M, FukaiS. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res, 2008, 109: 1-23[本文引用:1][JCR: 2.474]
[6]
张智猛, 戴良香, 丁红, 陈殿绪, 杨伟强, 宋文武, 万书波. 中国北方主栽花生品种抗旱性鉴定与评价. 作物学报, 2012, 38: 495-504ZhangZ M, DaiL X, DingH, ChenD X, YangW Q, SongW W, WanS B. Identification and evaluation of drought resistance in different peanut varieties widely grown in northern China. Acta Agron Sin, 2012, 38: 495-504 (in Chinese with English abstract)[本文引用:4][CJCR: 1.667]
[7]
王贺正, 李艳, 马均, 张荣萍, 李旭毅, 汪仁全. 水稻苗期抗旱性指标的筛选. 作物学报, 2007, 33: 1523-1529WangH Z, LiY, MaJ, ZhangR P, LiX Y, WangR Q. Screening indexes of drought resistance during seedling stage in rice. Acta Agron Sin, 2007, 33: 1523-1529 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[8]
王士强, 胡银岗, 佘奎军, 周琳璘, 孟凡磊. 小麦抗旱相关农艺性状和生理生化性状的灰色关联度分析. 中国农业科学, 2007, 40: 2452-2459WangS Q, HuY G, SheK J, ZhouL L, MengF L. Gray relational grade analysis of agronomical and physi-biochemical traits related to drought tolerance in wheat. Sci Agric Sin, 2007, 40: 2452-2459 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[9]
严美玲, 李向东, 矫岩林, 王丽丽. 不同花生品种的抗旱性比较鉴定. 花生学报, 2004, 33: 8-12YanM L, LiX D, JiaoY L, WangL L. Identification of drought resistance in different peanut varieties. J Peanut Sci, 2004, 33: 8-12 (in Chinese with English abstract)[本文引用:4]
[10]
张智猛, 万书波, 戴良香, 宋文武, 陈静, 石运庆. 花生抗旱性鉴定指标的筛选与评价. 植物生态学报, 2011, 35: 100-109ZhangZ M, WanS B, DaiL X, SongW W, ChenJ, ShiY Q. Estimating and screening of drought resistance indexes of peanut. Chin J Plant Ecol, 2011, 35: 100-109 (in Chinese with English abstract)[本文引用:4][CJCR: 1.989]
[11]
谭忠, 朱新亮, 刘文霞, 吴学军. 花生种质资源抗旱性鉴定及综合利用评价. 中国油料, 1997, 19(4): 73-75TanZ, ZhuX L, LiuW X, WuX J. Drought-resistant characterization and evaluation of pre-selected groundnut germplasm. Oil Crops China, 1997, 19(4): 73-75 (in Chinese with English abstract)[本文引用:4]
[12]
姜慧芳, 任小平, 段乃雄. 中国龙生型花生的耐旱性鉴定与综合评价. 中国农业科学, 1999, 32(suppl): 59-63JiangH F, RenX P, DuanN X. Screening and evaluation for drought tolerance in Chinese dragon groundnut. Sci Agric Sin, 1999, 32(suppl): 59-63 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[13]
胡标林, 余守武, 万勇, 张铮, 邱兵余, 谢建坤. 东乡普通野生稻全生育期抗旱性鉴定. 作物学报, 2007, 33: 425-432HuB L, YuS W, WanY, ZhangZ, QiuB Y, XieJ K. Drought-resistance identification of Dongxiang common wild rice (Oryza rufipogon) in whole growth period. Acta Agron Sin, 2007, 33: 425-432 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[14]
杨建昌, 王志琴, 朱庆森. 水稻品种的抗旱性及其生理特性的研究. 中国农业科学, 1995, 28(5): 65-72YangJ C, WangZ Q, ZhuQ S. Drought resistance and its physiological characteristics in rice varieties. Sci Agric Sin, 1995, 28(5): 65-72 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[15]
慕自新, 张岁岐, 梁爱华, 梁宗锁. 玉米整株根系水导与其表型抗旱性的关系研究. 作物学报, 2005, 31: 203-208MuZ X, ZhangS Q, LiangA H, LiangZ S. Relationship between maize root hydraulic conductivity and drought resistance. Acta Agron Sin, 2005, 31: 203-208 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[16]
段舜山, 谷文祥, 张大勇, 李凤民. 半干旱地区小麦群体的根系特征与抗旱性的关系. 应用生态学报, 1997, 8: 134-138DuanS S, GuW X, ZhangD Y, LiF M. Relationship between root system characteristics and drought resistance of wheat populations in semiarid region. Chin J Appl Ecol, 1997, 8: 134-138 (in Chinese with English abstract)[本文引用:3][CJCR: 1.742]
[17]
BenjaminJ G, NielsenD C. Water deficit effects on root distribution of soybean, field pea and chickpea. Field Crops Res, 2006, 97: 248-253[本文引用:1][JCR: 2.474]
[18]
JongrungklangN, ToomsanB, VorasootN, JogloyS, BooteK J, HoogenboomG, PatanothaiA. Rooting traits of peanut genotypes with different yield responses to pre-flowering drought stress. Field Crops Res, 2011, 120: 262-270[本文引用:1][JCR: 2.474]
[19]
赵世杰, 史国安, 董新纯. 植物生理学实验指导. 北京: 中国农业科学技术出版社, 2002. pp45-47ZhaoS J, ShiG A, DongX C. Laboratory Manual of Plant Physiology. Beijing: Chinese Agricultural Science and Technology Press, 2002. pp45-47(in Chinese)[本文引用:1]
[20]
王爱国, 罗广华, 邵从本, 吴淑君, 郭俊彦. 大豆种子超氧物歧化酶的研究. 植物生理学报, 1983, 9: 77-84WangA G, LuoG H, ShaoC B, WuS J, GuoJ Y. A study on the superoxide dismutase of soybean seeds. Acta Phytophysiol Sin, 1983, 9: 77-84 (in Chinese with English abstract)[本文引用:1]
[21]
WakamatsuK, TakahamaU. Changes in peroxidase activity and in peroxidsae isozymes in carrot callus. Physiol Plant, 1993, 88: 167-171[本文引用:1][JCR: 6.555]
[22]
林植芳, 李双顺, 林桂珠, 孙谷畴, 郭俊彦. 水稻叶片的衰老与超氧物歧化酶活性的关系及脂质过氧化作用的关系. 植物学报, 1984, 26: 605-615LinZ F, LiS S, LinG Z, SunG C, GuoJ Y. Superoxide dismutase activity and lipid peroxidation in relation to senescence of rice leaves. Acta Bot Sin, 1984, 26: 605-615 (in Chinese with English abstract)[本文引用:1][JCR: 0.599]
[23]
陈建勋, 王晓峰. 植物生理学实验指导. 广州: 华南理工大学出版社, 2006. pp75-77ChenJ X, WangX F. Laboratory Manual of Plant Physiology. Guangzhou: South China University of Technology Press, 2006. pp75-77(in Chinese)[本文引用:1]
张永清, 苗果园. 水分胁迫条件下有机肥对小麦根苗生长的影响. 作物学报, 2006, 32: 811-816ZhangY Q, MiaoG Y. Effects of manure on root and shoot growth of winter wheat under water stress. Acta Agron Sin, 2006, 32: 811-816 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
杨守萍, 陈加敏, 刘莹, 喻德跃, 盖钧镒. 大豆苗期耐旱性与根系性状的鉴定和分析. 大豆科学, 2005, 24: 176-182YangS P, ChenJ M, LiuY, YuD Y, GaiJ Y. Identification and analysis of drought tolerance and root traits of seedlings in soybeans. Soybean Sci, 2005, 24: 176-182 (in Chinese with English abstract)[本文引用:2]
[27]
梁银丽, 杨翠玲. 不同抗旱型小麦根系形态与生理特性对渗透胁迫的反应. 西北农业学报, 1995, (4): 31-36LiangY L, YangC L. Responses of root system morphology and physiological characters on osmotic stress in drought resistance wheat varieties. Acta Agric Boreali-Occident Sin, 1995, (4): 31-36 (in Chinese with English abstract)[本文引用:1]
[28]
宋海星, 王学立. 玉米根系活力及吸收面积的空间分布变化. 西北农业学报, 2005, 14(1): 137-141SongH X, WangX L. The space distribution of the maize root activity and its absorbing area. Acta Agric Boreali-Occident Sin, 2005, 14(1): 137-141 (in Chinese with English abstract)[本文引用:1]
[29]
刘莹, 盖钧镒, 吕慧能. 大豆根区逆境耐性的种质鉴定及其与根系性状的关系. 作物学报, 2005, 31: 1132-1137LiuY, GaiJ Y, LüH N. Identification of rhizosphere abiotic stress tolerance and related root traits in soybean [Glycine max (L. ) Merr. ]. Acta Agron Sin, 2005, 31: 1132-1137 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[30]
CristinaS, BrankaS, FlaviaN I. Role of phenolics in the antioxidative status of the resurrection plant Ramonda serbica during dehydration and rehydration. Physiol Plant, 2004, 122: 478-485[本文引用:1][JCR: 6.555]
[31]
齐伟, 张吉旺, 王空军, 刘鹏, 董树亭. 干旱胁迫对不同耐旱性玉米杂交种产量和根系生理特性的影响. 应用生态学报, 2010, 21: 48-52QiW, ZhangJ W, WangK J, LiuP, DongS T. Effects of drought stress on the grain yield and root physiological traits of maize varieties with different drought tolerance. Chin J Appl Ecol, 2010, 21: 48-52 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[32]
尚晓颍, 刘化冰, 张小全, 林娟, 段旺军, 杨铁钊. 干旱胁迫对不同烤烟品种根系生长和生理特性的影响. 西北植物学报, 2010, 30: 357-361ShangX Y, LiuH B, ZhangX Q, LinJ, DuanW J, YangT Z. Growth and physiological characteristics of roots in different flue-cured tobacco varieties under drought stress. Acta Bot Boreal-Occident Sin, 2010, 30: 357-361 (in Chinese with English abstract)[本文引用:1]
[33]
王思思, 张吉旺, 刘鹏, 董树亭, 王空军. 干旱对不同玉米品种苗期根系生理生化特性的影响. 山东农业科学, 2009, (6): 36-38WangS S, ZhangJ W, LiuP, DongS T, WangK J. Effect of drought on root physiological and biochemical characters of different maize cultivars during seedling stage. Shand ong Agric Sci, 2009, (6): 36-38 (in Chinese with English abstract)[本文引用:1]
[34]
盖钧镒, 汪越胜, 张孟臣, 王继安, 常汝镇. 中国大豆品种熟期组划分的研究. 作物学报, 2001, 27: 286-292GaiJ Y, WangY S, ZhangM C, WangJ A, ChangR Z. Studies on the classification of maturity groups of soybeans in China. Acta Agron Sin, 2001, 27: 286-292 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[35]
陈加敏. 大豆苗期耐旱性的鉴定及苗期耐旱性和根系性状的遗传研究. 南京农业大学硕士学位论文, 2004ChenJ M. Studies on Identification of Drought Tolerance and Genetic Mechanism of Drought Tolerance and Root Traits of Soybean Seedling. MS Thesis of Nanjing Agricultural University, Nanjing, China, 2004 (in Chinese with English abstract)[本文引用:1]

{kind=link}
{kind=link}