关键词:小麦; 旗叶; 高温胁迫; 氮肥追施后移; 籽粒产量 Effects of Postponed Application of Nitrogen Fertilizer on Yield and Physiological Characteristics of Flag Leaf in Wheat under Post-anthesis Heat Stress JIANG Wen-Wen, YIN Yan-Ping*, WANG Zhen-Lin*, LI Yong, YANG Wei-Bing, PENG Dian-Liang, YANG Dong-Qing, CUI Zheng-Yong, LU Kun-Li, LI Yan-Xia Agronomy College, Shandong Agricultural University / State Key Laboratory of Crop Biology, Tai’an 271018, China Fund: AbstractThis study aimed to disclose the effect and physiological basis of postponed application of nitrogen fertilizer on alleviating the high-temperature damage after anthesis. In a two-year experiment from October 2011 to June 2013, winter wheat (Triticum aestivum L.) cultivars Shannong 16 and Jimai 22 were planted with three nitrogen treatments, namely T0 (total as basal fertilizer), T1 (a half of basal fertilizer and a half of nitrogen applied at jointing stage), and T2 (a half of basal fertilizer and a half applied at booting stage). High-temperature stress was simulated in plastic shed from 11 to 15 days after anthesis. Grain yield and its components, photosynthetic parameters of flag leaf, and activities of some important enzymes were compared among nitrogen treatments. The results indicated that T2 had higher 1000-grain weight and grain yield than T0 and T1 under heat stress. Besides, T2 had the effects on increasing glutamine synthetase (GS) activity, stomatal conductance, photosynthetic rate, and activities of hydrogen peroxidase (CAT) and peroxidase (POD) in flag leaf, stimulating translocation of pre-anthesis assimilate to grains, and increasing the contribution to grain of post-anthesis assimilate. Both cultivars had similar results. In an overall view, T2 is superior to T0 and T1 due to its significant effects on alleviating heat stress damage and increasing grain yield.
Keyword:Wheat; Flag leaf; Heat stress; Postponed application of nitrogen fertilizer; Grain yield Show Figures Show Figures
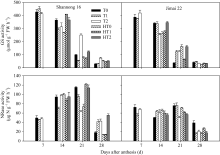
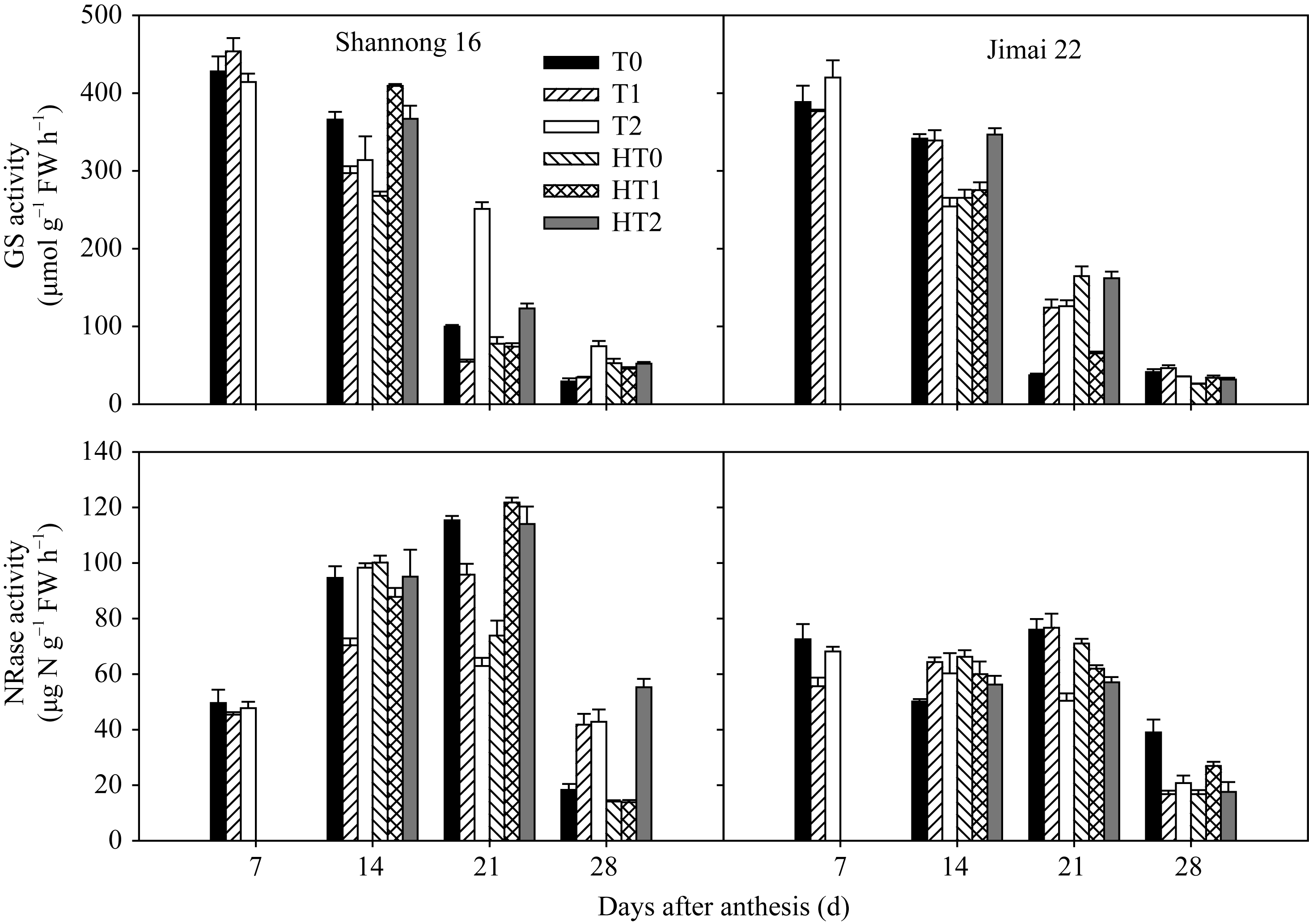
图2 高温胁迫和施氮处理对小麦旗叶谷氨酰胺合成酶(GS)和硝酸还原酶(NR)活性的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 2 Effects of heat stress and nitrogen application on glutamine synthetase (GS) and nitrate reductase (NR) activities in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates.
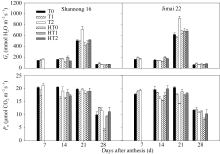
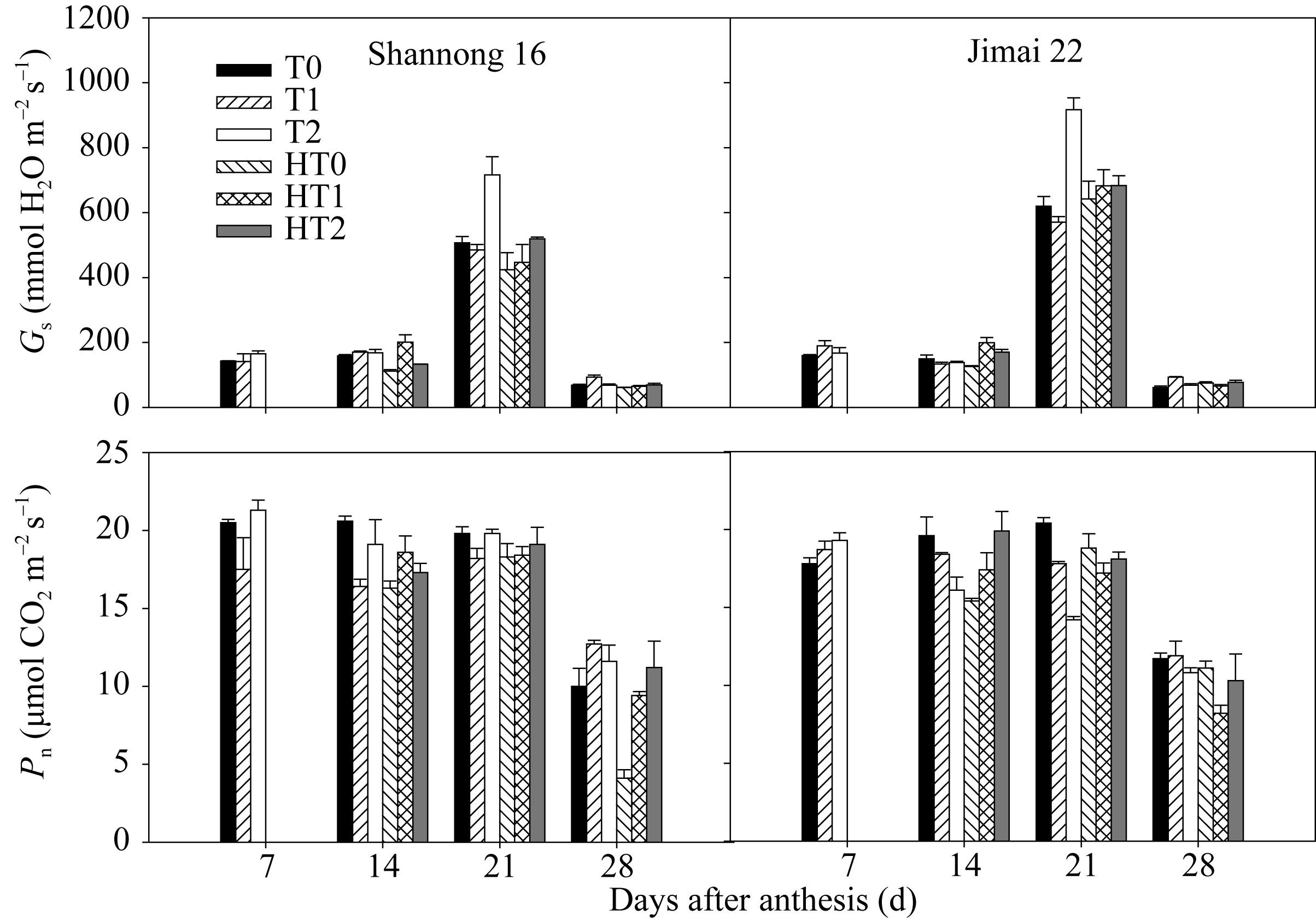
图3 高温胁迫和施氮处理对小麦旗叶气孔导度( Gs)和光合速率( Pn)的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 3 Effects of heat stress and nitrogen application on stomatal conductance ( Gs) and photosynthetic rate ( Pn) in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates.
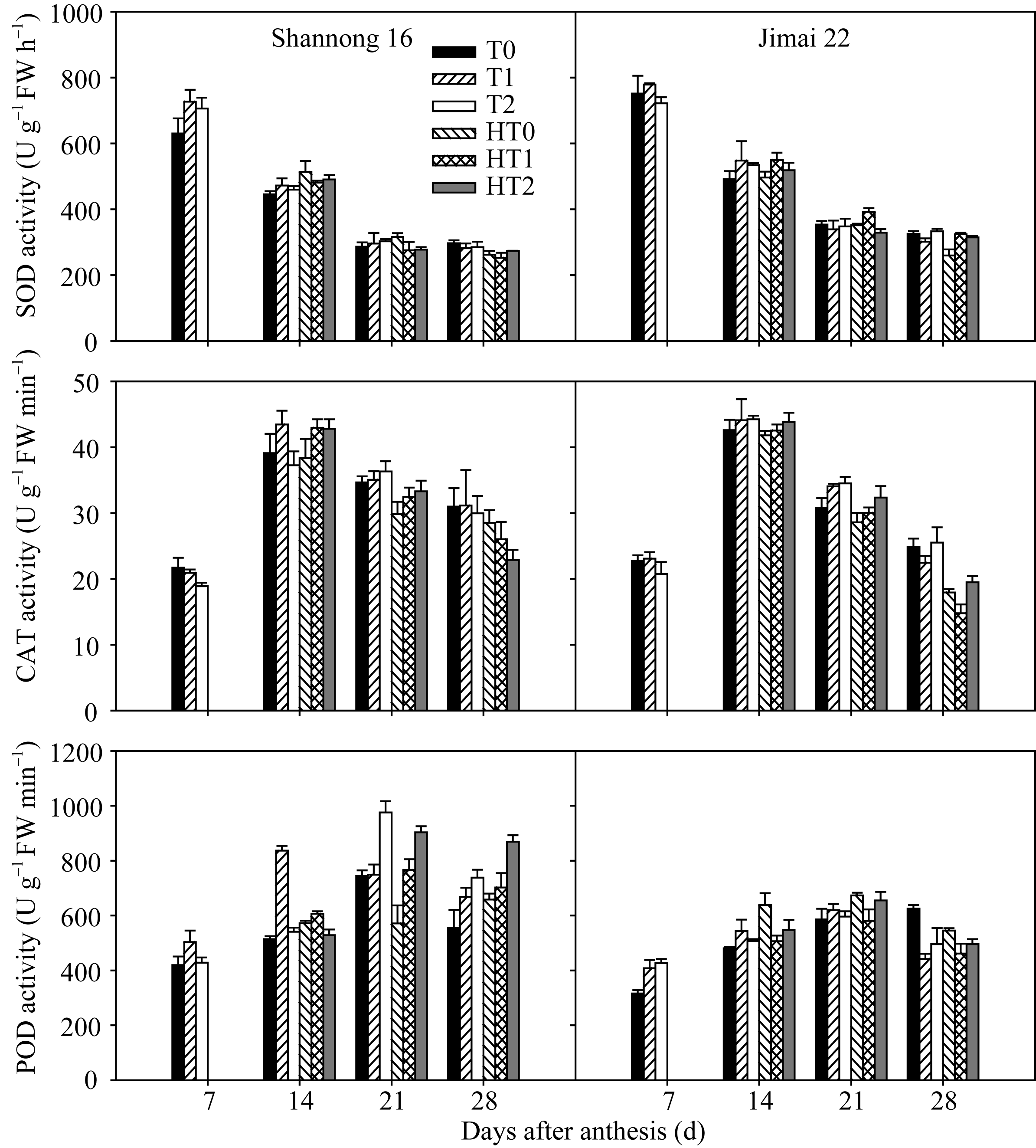
图4 高温胁迫和施氮处理对小麦旗叶超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 4 Effects of heat stress and nitrogen application on activities of superoxide dismutase (SOD), hydrogen peroxidase (CAT), and peroxidase (POD) in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates.
表1 不同处理对小麦开花后营养器官干物质积累量和干物质再分配的影响 Table 1 Effects of different treatments on dry matter accumulation and translocation amount of vegetative organ after anthesis in wheat
年份 Year
处理 Treatment
开花前贮藏的同化物 Pre-anthesis reserves
开花后积累的干物质 Post-anthesis assimilate
向籽粒转运量 Translocated into grains (kg hm-2)
转运率 Percentage (%)
籽粒贡献率 Contribution to grain (%)
积累量 Accumulation (kg hm-2)
籽粒贡献率 Contribution to grain (%)
山农16 Shannong 16
2011-2012
T0
1374.14 c
16.73 b
28.43 bc
3490.55 cd
71.57 ab
T1
1952.07 b
23.28 a
27.17 c
5264.67 a
72.83 a
T2
2765.41 a
26.96 a
37.49 ab
4616.56 ab
62.51 bc
HT0
1927.68 b
23.47 a
43.50 a
2509.24 e
56.50 c
HT1
2036.32 b
24.28 a
43.53 a
2663.59 de
56.47 c
HT2
1626.41 bc
15.85 b
28.15 bc
4167.97 bc
71.85 ab
2012-2013
T0
1513.47 b
18.34 ab
26.21 ab
4264.87 b
73.79 ab
T1
1465.42 b
18.04 ab
20.70 b
5908.82 a
79.30 a
T2
2644.56 a
26.45 a
36.24 ab
4674.61 ab
63.76 ab
HT0
1401.26 b
16.98 ab
35.94 ab
2495.38 d
64.06 ab
HT1
1924.53 ab
23.70 ab
42.93 a
2643.24 cd
57.07 b
HT2
1579.25 b
15.80 b
27.86 ab
4163.40 bc
72.14 ab
济麦22 Jimai 22
2011-2012
T0
1489.31 c
19.71 c
28.47 b
3747.70 a
71.53 a
T1
3096.76 b
33.42 b
47.18 a
3477.08 ab
52.82 b
T2
3582.20 a
37.56 a
49.12 a
3718.47 a
50.88 b
HT0
1639.69 c
21.70 c
34.30 b
3141.72 b
65.70 a
HT1
3161.22 b
34.12 ab
49.08 a
3278.73 ab
50.92 b
HT2
2977.36 b
31.22 b
44.75 a
3686.25 ab
55.25 b
2012-2013
T0
1192.67 b
16.17 b
22.96 c
4061.31 a
77.04 a
T1
3088.26 a
34.18 a
46.90 a
3522.54 a
53.10 c
T2
3130.67 a
33.18 a
48.66 a
3330.58 a
51.34 c
HT0
1432.78 b
19.42 b
30.15 bc
3344.54 a
69.85 ab
HT1
2799.70 a
30.98 a
43.41 ab
3629.78 a
56.59 bc
HT2
2967.50 a
31.45 a
45.57 ab
3586.66 a
54.43 bc
数据后不同字母表示同一年度内处理间有显著差异( P< 0.05)。 Values followed by different letters are significantly different within the same year at P < 0.05.
表1 不同处理对小麦开花后营养器官干物质积累量和干物质再分配的影响 Table 1 Effects of different treatments on dry matter accumulation and translocation amount of vegetative organ after anthesis in wheat
表2 不同处理对小麦籽粒产量及构成因素的影响 Table 2 Effects of different treatments on grain yield and yield components of wheat
处理Treatment
山农16 Shannong 16
济麦22 Jimai 22
穗数 SN (×104 hm-2)
穗粒数 GNS
千粒重 TGW (g)
籽粒产量 GY (kg hm-2)
穗数 SN (×104 hm-2)
穗粒数 GNS
千粒重 TGW (g)
籽粒产量 GY (kg hm-2)
2011-2012
T0
464.76 a
43.90 abc
39.19 bc
7450.00 cd
473.01 a
38.56 a
41.49 bc
8663.67 b
T1
466.41 a
45.10 a
40.64 b
7728.27 bc
479.04 a
38.84 a
45.29 a
9216.80 a
T2
468.06 a
44.78 ab
44.50 a
8090.87 ab
469.14 a
41.16 a
45.38 a
9115.33 a
HT0
464.76 a
40.00 cd
37.53 c
7084.40 d
473.01 a
38.29 a
39.95 c
8117.80 cd
HT1
466.41 a
41.69 bcd
39.21 bc
7439.80 cd
479.04 a
38.38 a
42.48 b
8102.33 cd
HT2
468.06 a
39.06 d
40.70 b
7602.67 c
469.14 a
38.67 a
42.95 b
8251.53 bc
2012-2013
T0
447.07 a
37.97 c
36.90 b
6317.27 c
462.93 a
33.80 ab
32.03 c
7072.60 b
T1
451.73 a
46.77 a
41.66 a
6933.20 ab
458.27 a
37.87 a
36.77 b
7526.77 a
T2
510.53 a
41.87 b
41.94 a
7101.13 a
459.20 a
37.33 a
43.59 a
7668.60 a
HT0
447.07 a
39.57 bc
26.60 d
6033.37 d
462.93 a
32.27 b
30.58 c
6601.80 c
HT1
451.73 a
39.13 bc
26.86 d
6649.17 b
458.27 a
35.43 ab
37.11 b
7040.80 b
HT2
510.53 a
38.87 bc
29.33 c
6895.50 ab
459.20 a
38.33 a
38.34 b
7370.43 ab
数据后不同字母表示同一年度内处理间有显著差异( P< 0.05)。 Values followed by different letters are significantly different within the same year at P < 0.05. SN: spike number; GNS: grain number per spike; TGW: 1000-grain weight; GY: grain yield.
表2 不同处理对小麦籽粒产量及构成因素的影响 Table 2 Effects of different treatments on grain yield and yield components of wheat
徐如强, 孙其信, 张树榛. 小麦耐热性研究现状与展望. 中国农业大学学报, 1998, 3(3): 33-40XuR Q, SunQ X, ZhangS Z. Current status and prospective on the investigation of heat tolerance in wheat. J China Agric Univ, 1998, 3(3): 33-40 (in Chinese with English abstract)[本文引用:1]
[2]
Alexand erL X, ZhangX, PetersonT C, CaesarJ, GleasonB, Klein TankA M G, HaylockM, CollinsD, TrewinB, RahimzadehF, TagipourA, RupaKumar K, RevadekarJ, GriffithsG, VincentL, StephensonD B, BurnJ, AguilarE, BrunetM, TaylorM, NewM, ZhaiP, RusticucciM, Vazquez-AguirreJ L. Global observed changes in daily climate extremes of temperature and precipitation. J Geophys Res Atmospheres, 2006, 111: 1-22[本文引用:1]
[3]
AssengS, FosterI, TurnerN C. The impact of temperature variability on wheat yields. Global Change Biol, 2011, 17: 997-1012[本文引用:1][JCR: 6.91]
[4]
WardlawI F. Factors limiting the rate of dry matter accumulation in the grain of wheat grown at high temperature. Aust J Plant Physiol, 1980, 7: 387-400[本文引用:1]
[5]
李永庚, 于振文, 张秀杰, 高雷明. 小麦产量与品质对灌浆不同阶段高温胁迫的响应. 植物生态学报, 2005, 29: 461-466LiY G, YuZ W, ZhangX J, GaoL M. Response of yield and quality of wheat to heat stress at different grain filling stages. Chin J Plant Ecol, 2005, 29: 461-466 (in Chinese with English abstract)[本文引用:1][CJCR: 1.989]
[6]
岳寿松, 于振文, 余松烈, 许玉敏. 不同生育时期施氮对冬小麦旗叶衰老和粒重的影响. 中国农业科学, 1997, 30(2): 42-46YueS S, YuZ W, YuS L, XuY M. Effects of nitrogen application at different growth stages on the senescence of flag leaves and grain yield in winter wheat (Triticum aestivum L. ). Sci Agric Sin, 1997, 30(2): 42-46 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[7]
SmithC J, WhiteD M. Nitrogen accumulation and redistribution of late applied of N15 labeled fertilizer by wheat. Field Crops Res, 1990, 24: 221-228[本文引用:1][JCR: 2.474]
[8]
PearmenI, ThomasS M. Thorne G N. Effects of nitrogen ferti-lizer on photosynthesis of several varieties of winter wheat. Ann Bot, 1997, 43: 613-621[本文引用:1][JCR: 0.657]
[9]
刘永环, 贺明荣, 王晓英, 张洪华. 不同氮肥基追比例对高温胁迫下小麦籽粒产量和品质的影响. 生态学报, 2009, 29: 5930-5935LiuY H, HeM R, WangX Y, ZhangH H. The effects of topdressing to basal nitrogen ratios on grain yield and quality of winter wheat under heat stresses during grain filling. Acta Ecol Sin, 2009, 29: 5930-5935 (in Chinese with English abstract)[本文引用:1]
[10]
曹云英. 高温对水稻产量与品质的影响及其生理机制. 扬州大学博士学位论文, 2009CaoY Y. Effect of High Temperature on the Quality and Quantity of Rice Yield and Its Physiological Mechanism. PhD Dissertation of Yangzhou University, Yangzhou, China, 2009 (in Chinese with English abstract)[本文引用:1]
[11]
徐晓玲, 王志敏, 张俊平. 灌浆期热胁迫对不同绿色器官光合性能的影响. 植物学报, 2001, 43: 571-577XuX L, WangZ M, ZhangJ P. Effect of heat stress on photosynthetic characteristics of different green organs of winter wheat during grain-filling stage. Acta Bot Sin, 2001, 43: 571-577 (in Chinese with English abstract)[本文引用:1][JCR: 0.599]
[12]
Perez-SobaM, StulenL, van der EerdenL J M. Effect of atmospheric ammonia on the nitrogen metabolism of Scots pine (Pinus sylvestris) needles. Physiol Plant, 1994, 90: 629-636[本文引用:1][JCR: 6.555]
[13]
SilveiraJ A G, MatosJ C S, CecattoV M, ViegasR A, OliveiraJ T A. Nitrate reductase activity, distribution, and response to nitrate in two contrasting Phaseolus species inoculated with Rhizobium spp. Environ Exp Bot, 2001, 46: 37-46[本文引用:1][JCR: 2.578]
[14]
TanW, LiuJ, DaiT, JingQ, CaoW, JiangD. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis water-logging. Photosynthethetica, 2008, 46: 21-27[本文引用:2]
[15]
KlapheckS, ZimmerI, CosseH. Scavenging of hydrogen pero-xide in the endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol, 1990, 31: 1005-1013[本文引用:1][JCR: 4.134]
WangG Y, LiuJ M, ZhangY, YuB S, ShenZ Y. Studies on ultrstructure in common bean leaves during heat acclimation and heat stress. J Agric Biotech, 1999, 7: 151-156[本文引用:1][CJCR: 0.635]
[18]
LiuX, HuangB. Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Sci, 2000, 40: 503-513[本文引用:1][JCR: 1.513]
[19]
XuS, LiJ L, ZhangX Q, WeiH, CuiL G. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ Exp Bot, 2006, 56: 274-285[本文引用:1][JCR: 2.578]
[20]
WangX, CaiJ, JiangD, LiuF L, DaiT B, CaoW X. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J Plant Physiol, 2011, 168: 585-593[本文引用:1][JCR: 2.699]
[21]
段骅, 俞正华, 徐云姬, 王志琴, 刘立军, 杨建昌. 灌溉方式对减轻水稻高温危害的作用. 作物学报, 2012, 38: 107-120DuanH, YuZ H, XuY J, WangZ Q, LiuL J, YangJ C. Role of irrigation patterns in reducing harms of high temperature to rice. ActaAgron Sin, 2012, 38: 107-120 (in Chinese with English abstract)[本文引用:1]
[22]
于振文, 田奇卓, 潘庆民, 岳寿松, 王东, 段藏禄, 段玲玲, 王志军, 牛运生. 黄淮麦区冬小麦超高产栽培的理论与实践. 作物学报, 2002, 28: 577-585YuZ W, TianQ Z, PanQ M, YueS S, WangD, DuanZ L, DuanL L, WangZ J, NiuY S. Theory and practice on cultivation of super high yield of winter wheat in the wheat fields of Yellow River and Huaihe River district. Acta Agron Sin, 2002, 28: 577-585 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[23]
潘庆民, 于振文. 追氮时期对冬小麦籽粒品质和产量的影响. 麦类作物学报, 2002, 22(2): 65-69PanQ M, YuZ W. Effects of nitrogen topdressing stage on grain quality and yield of winter wheat. J Triticeae Crops, 2002, 22(2): 65-69 (in Chinese with English abstract)[本文引用:1]
[24]
王月福, 姜东, 于振文, 曹卫星. 高低土壤肥力下小麦基施和追施氮肥的利用效率和增产效应, 作物学报, 2003, 29: 491-495WangY F, JiangD, YuZ W, CaoW X. Nitrogen use efficiency and yield of wheat with basal and top-dressed nitrogen fertilizers in soils with different fertility. Acta Agron Sin, 2003, 29: 491-495 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[25]
季书勤, 赵淑章, 吕凤荣, 刘媛媛, 王绍中. 小麦前氮后移的施肥效应与有效使用条件. 作物学报, 1998, 24: 889-893JiS Q, ZhaoS Z, LüF R, LiuY Y, WangS Z. The effect of nitrogen application under different conditions at late growth stage of wheat. Acta Agron Sin, 1998, 24: 889-893 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
RuizJ M, RiveroR M, GarciaP C, BaghourM, RomeroL. Role of CaCl2 in nitrate assimilation in leaves and roots of tobacco plants (Nicotiana tabacum L. ). Plant Sci, 1999, 141: 107-115[本文引用:1][JCR: 2.922]
[27]
SolomonsonL P, SpeharA M. Model for the regulation of nitrate assimilation. Nature, 1977, 265: 373-375[本文引用:1][JCR: 38.597]
[28]
BenJ M, DimahZ H. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J Exp Bot, 2002, 53: 979-987[本文引用:1][JCR: 5.242]
[29]
NoctorG, FoverC H. Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Mol Biol, 1998, 49: 249-279[本文引用:1][JCR: 25.962]
[30]
BlumA, EberconA. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci, 1981, 21: 43-47[本文引用:1][JCR: 1.513]
[31]
张生杰, 黄元炯, 任庆成, 张小全, 杨志晓, 杨铁钊. 氮素对不同品种烤烟叶片衰老、光合特性及产量和品质的影响. 应用生态学报, 2010, 21: 668-674ZhangS J, HuangY J, RenQ C, ZhangX Q, YangZ X, YangT Z. Effects of nitrogen fertilization on leaf senescence, photosynthetic characteristics, yield, and quality of different flue-cured tobacco varieties. Chin J Appl Ecol, 2010, 21: 668-674 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[32]
EvansL T, DunstoneR L. Some physiological aspects of evolution in wheat. Aust J Biol Sci, 1970, 23: 725-741[本文引用:1]
[33]
韩占江, 于振文, 王东, 张永丽. 测墒补灌对冬小麦干物质积累与分配及水分利用效率的影响. 作物学报, 2010, 36: 457-465HanZ J, YuZ W, WangD, ZhangY L. Effects of supplemental irrigation based on testing soil moisture on dry matter accumulation and distribution and water use efficiency in winter wheat. Acta Agron Sin, 2010, 36: 457-465 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
 , 尹燕枰
, 尹燕枰


{kind=link}
{kind=link}
{kind=link}
{kind=link}