关键词:普通菜豆; 干旱胁迫; 生理特性; 评价指标 Physiological Characteristics of Drought Resistance in Common Bean (Phaseolus vulgarisL.) LI Long, WANG Lan-Fen, WU Jing, JING Rui-Lian, WANG Shu-Min* National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund: AbstractThree common bean cultivars, Yuejindou (drought-resistant), 260205 (drought-resistant), and Naihuayundou (drought- susceptible) were evaluated with two treatments (drought stress and normal water supply) in pot experiments. We determined physiological and biochemical parameters during growth stage and yield and traits related to yield after harvest, analyzed the changes of all parameters and indicators under drought stress. The result showed that the root dry weight of Yuejindou and 260205 was 20.2% and 20.6% of the total biomass and the pod dry weight was 30.0% and 28.9% of the total biomass especially at 36 days of drought treatment, while, the root dry weight and pod dry weight only 10.6% and 17.1% of the total biomass in Naihuayundou, indicating that effective photosynthate distribution is significantly correlated with drought resistance in common bean. The water use efficiency in the drought-resistant cultivar (Yuejindou) increased by 230.5% compared with control, much higher than the 60% increase in drought-susceptible cultivar (Naihuayundou). The drought-resistant cultivars had an efficient CO2 diffusion and fixation in leaf tissues, and an effective water-use. In drought-resistant cultivars, antioxidant enzyme and photorespiration played a significant role in reactive oxygen scavenging; proline and soluble sugar contributed to the maintenance of relative water content in leaves under low water potential. All together, our results indicated that the competence of drought-resistant cultivars to maintain seed production under drought stress relies on effective adjustments in morphology, stomatal conductance, osmosis and antioxidant capacity.
Keyword:Common bean; Drought stress; Physiological characteristics; Evaluation indices Show Figures Show Figures
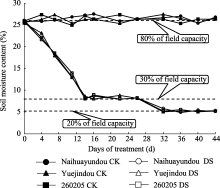
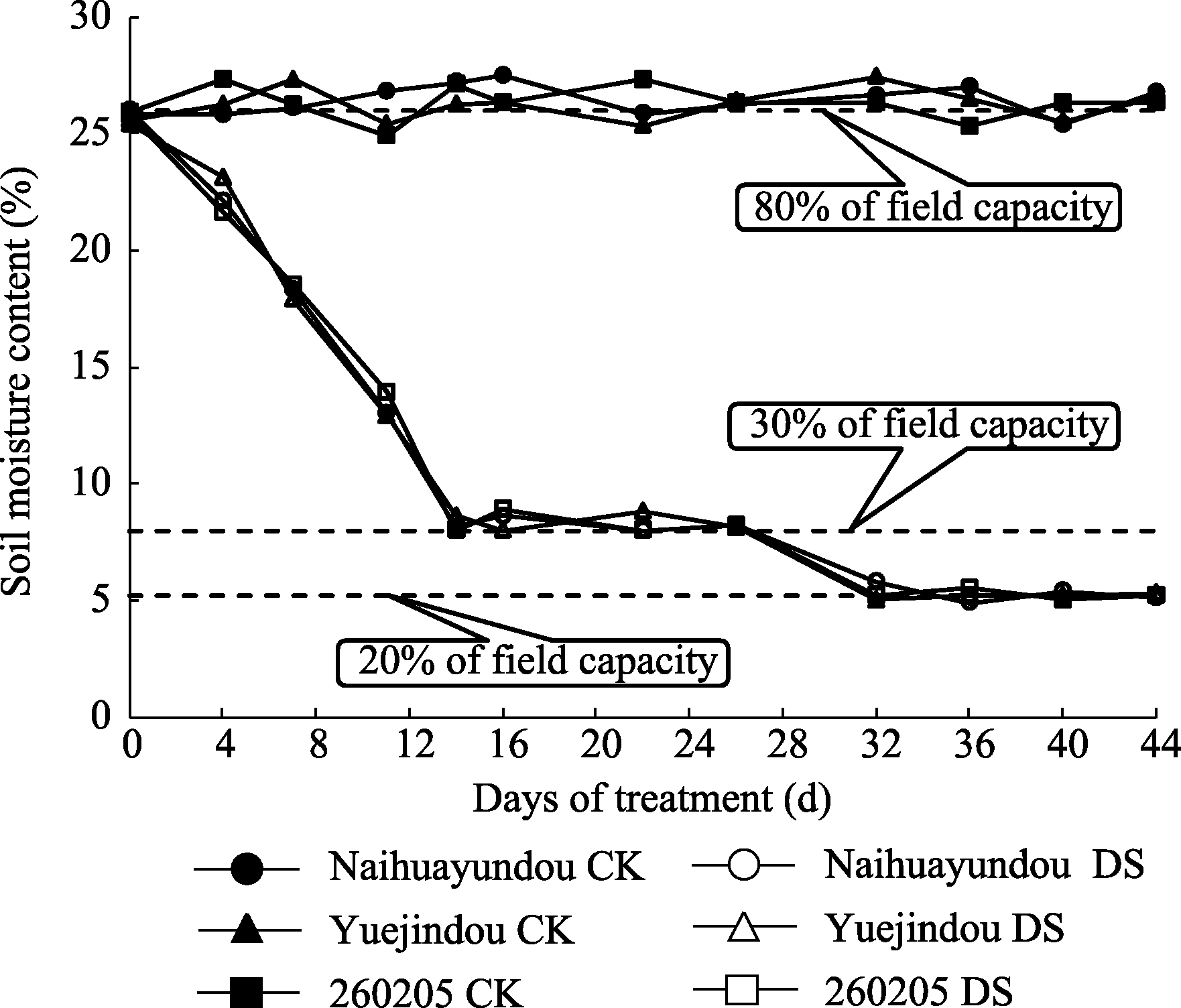
图1 不同处理土壤含水量变化CK(对照): 80%田间持水量; DS(干旱处理): 干旱胁迫前期保持30%田间持水量, 干旱胁迫后期保持20%田间持水量。Fig. 1 Changes of soil moisture content in different treatmentsCK (control): 80% field capacity; DS (drought stress): 30% of field capacity at early stage, 20% of field capacity at later stage.
表1 干旱胁迫下不同品种产量及其构成因素 Table 1 Yield and its components in different common bean cultivars under drought treatments
品种 Cultivar
处理 Treatment
单株荚数 Pod number per plant
单株粒数 Grain number per plant
荚长 Pod length (cm)
单株荚重 Pod weight per plant (g)
单荚粒数 Grain number per pod
单株产量 Yield per plant (g)
奶花芸豆 Naihuayundou
正常浇水CK
3.5±0.28 Aa
9.2±1.05 Aa
14.4±2.49 Aa
2.1±0.27 Aa
2.6±0.19 Aa
5.8±0.61 Aa
干旱胁迫DS
1.0±0.00 Bb
1.1±0.14 Bb
7.0±1.94 Bb
0.3±0.06 Bb
1.1±0.14 Bb
0.6±0.03 Bb
下降比例(%)
71.43
88.44
51.46
87.26
59.54
88.89
260205
正常浇水CK
9.5±1.38 Aa
31.5±4.53 Aa
7.4±0.49 Aa
1.7±0.20 Aa
3.3±0.22 Aa
5.8±0.62 Aa
干旱胁迫DS
3.4±0.83 Bb
7.2±0.78 Bb
6.6±0.47 Ba
0.3±0.05 Bb
2.2±0.51 Bb
2.1±0.12 Bb
下降比例(%)
64.32
77.24
11.46
81.33
33.43
63.70
跃进豆 Yuejindou
正常浇水CK
7.6±1.26 Aa
30.2±5.11 Aa
8.5±0.59 Aa
1.8±0.11 Aa
4.0±0.51 Aa
5.6±0.62 Aa
干旱胁迫DS
2.9±0.33 Bb
8.1±1.00 Bb
7.6±0.57 Ba
0.3±0.03 Bb
2.8±0.29 Ba
2.1±0.15 Bb
下降比例(%)
61.11
73.28
10.59
84.09
31.76
63.13
采用Duncan’s法, 不同小写字母表示差异显著( P<0.05), 不同大写字母表示差异极显著( P<0.01); CK: 对照; DS: 干旱处理。 Using Duncan’s test, values followed by different lowercases and capitals are significantly different at 0.05 and 0.01 probability levels, respectively. CK: control; DS: drought stress.
表1 干旱胁迫下不同品种产量及其构成因素 Table 1 Yield and its components in different common bean cultivars under drought treatments
表2 Table 2 表2(Table 2)
表2 干旱胁迫下产量构成因素与单株产量的相关性 Table 2 Correlation coefficients between yield component and yield per plant under drought stress
品种 Cultivar
单株荚数 Pod number per plant
单株粒数 Grain number per plant
荚长 Pod length
单株荚重 Pod weight per plant
单荚粒数 Grain number per pod
奶花芸豆 Naihuayundou
0.18
0.78**
0.29
0.46*
0.78**
260250
0.81**
0.80**
0.22
0.48*
0.80**
跃进豆 Yuejindou
0.81**
0.83**
0.61*
0.80**
0.83**
* 表示在0.05水平上相关,** 表示在0.01水平上相关。 * Correlation significant at P<0.05;** Correlation significant at P<0.01.
表2 干旱胁迫下产量构成因素与单株产量的相关性 Table 2 Correlation coefficients between yield component and yield per plant under drought stress
表3 Table 3 表3(Table 3)
表3 干旱胁迫下不同抗旱性品种各器官干重 Table 3 Morphological characteristics of different common bean cultivars under drought treatments
品种 Cultivar
处理时间 Days at treatment
处理 Treatment
茎叶干重 Shoot dry weight (g)
根干重 Root dry weight (g)
荚干重 Pod dry weight (g)
根冠比 Root/shoot
单株总生物量 Biomass per plant (g)
奶花芸豆 Naihuayundou
处理16 d 16DAT
正常浇水CK
10.91±0.78 Aa
2.38±0.28 Aa
—
0.22±0.02 Aa
13.29±0.95 Aa
干旱胁迫DS
4.81±0.14 Bb
0.78±0.08 Bb
—
0.16±0.02 Ba
5.59±0.13 Bb
下降比例(%)
55.87
67.41
—
26.04
57.94
处理36 d 36DAT
正常浇水CK
11.81±1.25 Aa
2.64±0.30 Bb
19.83±1.68 Aa
0.18±0.02 Aa
34.28±1.51 Aa
干旱胁迫DS
5.52±0.19 Bb
0.81±0.02 Bb
1.30±0.19 Bb
0.13±0.01 Ab
7.63±0.34 Bb
下降比例(%)
53.30
69.41
93.43
30.34
77.75
260250
处理16 d 16DAT
正常浇水CK
8.11±0.12 Aa
1.98±0.16 Aa
—
0.24±0.02 Aa
10.09±0.12 Aa
干旱胁迫DS
3.55±0.33 Bb
1.25±0.24 Ba
—
0.36±0.11 Ba
4.80±0.10 Bb
下降比例(%)
56.27
36.97
—
-48.06
52.48
处理36 d 36DAT
正常浇水CK
21.32±0.28 Aa
8.18±1.00 Aa
8.62±0.27 Aa
0.28±0.03 Aa
38.12±0.67 Aa
干旱胁迫DS
4.35±0.22 Bb
1.84±0.04 Bb
2.92±0.13 Bb
0.30±0.01 Ab
9.10±0.26 Bb
下降比例(%)
79.62
77.50
66.18
-7.67
76.12
跃进豆 Yuejindou
处理16 d 16DAT
正常浇水CK
8.72±1.16 Aa
2.91±0.12 Aa
—
0.34±0.04 Aa
11.63±1.25 Aa
干旱胁迫DS
3.26±0.15 Bb
1.81±0.06 Bb
—
0.56±0.04 Bb
5.07±0.10 Bb
下降比例(%)
62.65
37.69
—
-64.87
56.41
处理36 d 36DAT
正常浇水CK
22.66±0.92 Aa
4.26±0.22 Aa
5.73±0.69 Aa
0.16±0.01 Aa
32.64±0.27 Aa
干旱胁迫DS
5.26±0.14 Bb
2.15±0.12 Bb
3.02±0.37 Bb
0.29±0.01 Bb
10.44±0.62 Bb
下降比例(%)
76.77
49.41
47.29
-83.09
68.03
采用Duncan’s法, 不同小写字母表示差异显著( P<0.05), 不同大写字母表示差异极显著( P<0.01); CK: 对照; DS: 干旱处理; DAT: 干旱处理天数。 Using Duncan’s test, values followed by different lowercases and capitals are significantly different at 0.05 and 0.01 probability levels, respectively. CK: control; DS: drought stress; DAT: days after treatment.
表3 干旱胁迫下不同抗旱性品种各器官干重 Table 3 Morphological characteristics of different common bean cultivars under drought treatments
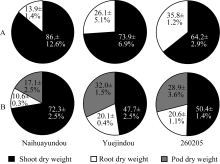
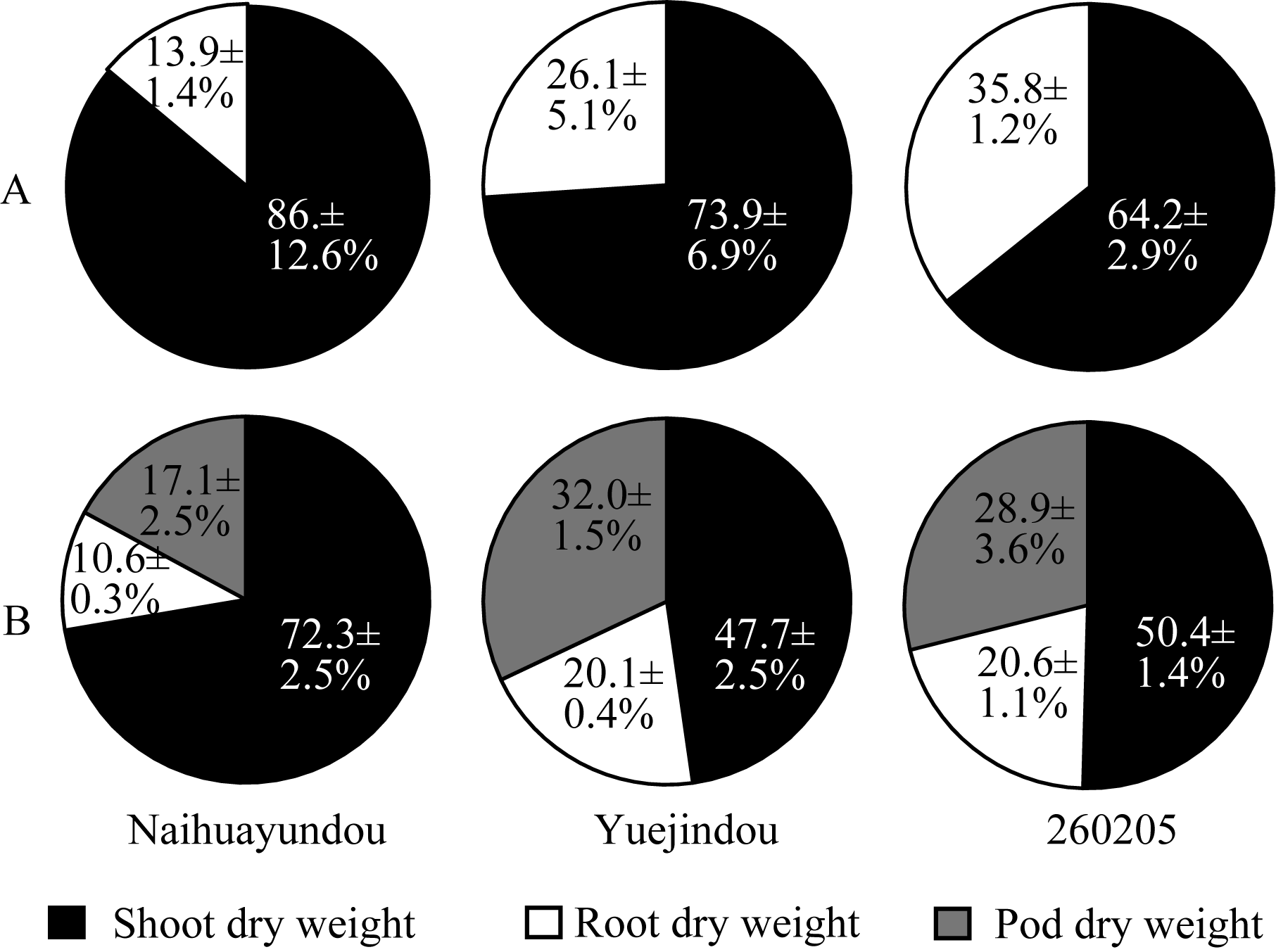
图2 干旱胁迫下不同品种干物质分配A: 干旱胁迫16 d; B: 干旱胁迫36 d。 A: 16 days of drought treatment; B: 36 days of drought treatment.Fig. 2 Dry matter partitioning of different common bean cultivars under drought treatments
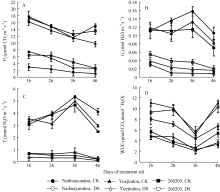
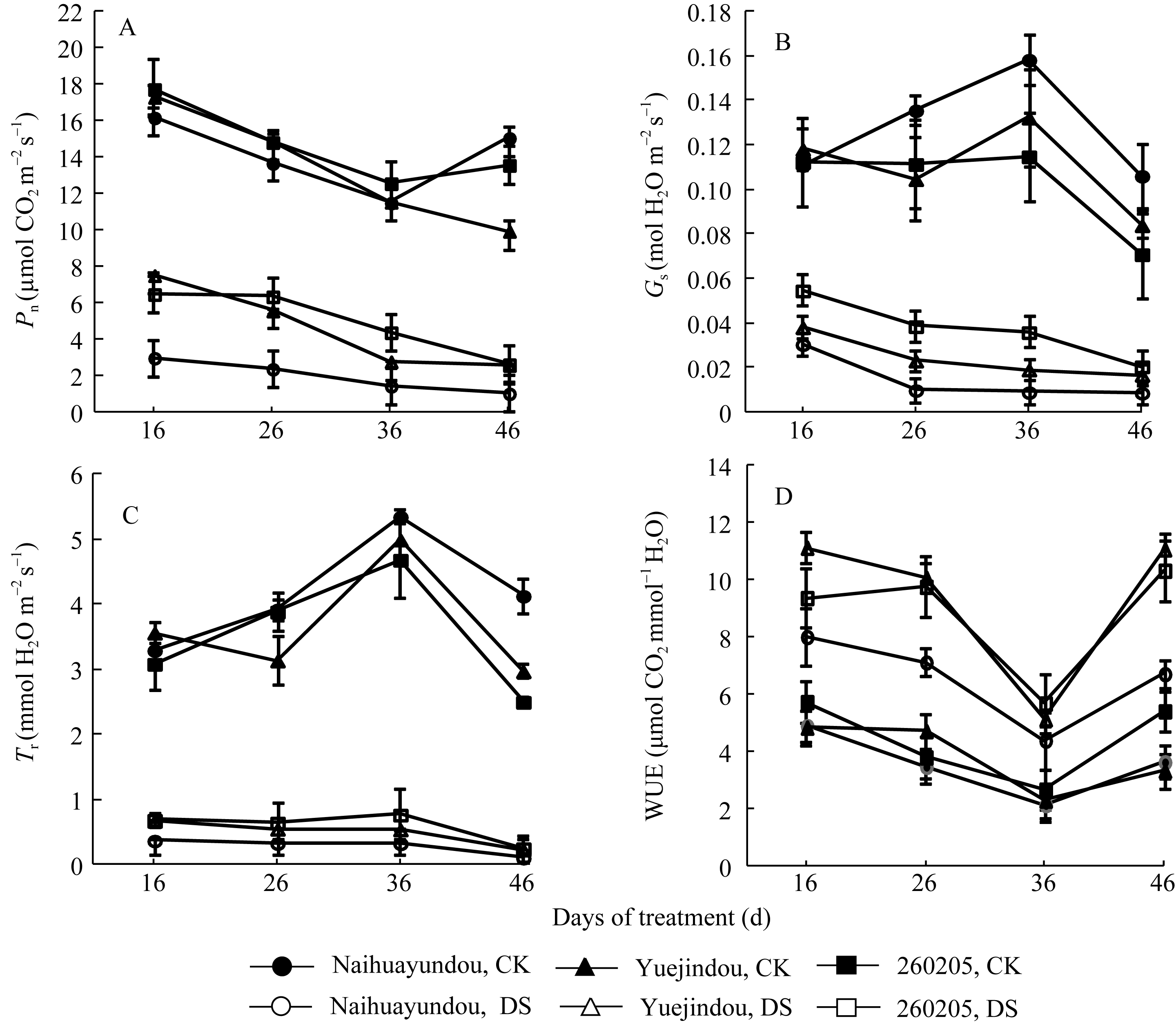
图3 干旱胁迫对不同抗旱性品种光合速率、气孔导度、蒸腾速率和水分利用效率的影响CK: 对照; DS: 干旱处理。CK: control; DS: drought stress.Fig. 3 Effects of drought stress on photosynthetic rate ( Pn), stomatal conductance ( Gs), transpiration rate ( Tr), and water use efficiency(WUE) in different common bean cultivars
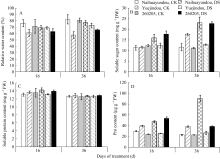
图4 干旱胁迫对不同抗旱性品种相对含水量及渗透调节物质的影响CK: 对照; DS: 干旱处理。CK: control; DS: drought stress.Fig. 4 Effects of drought stress on relative water content and content of osmoregulation substances after treatments in different common bean cultivars
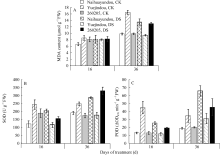
图5 干旱胁迫对不同品种保护酶活性及MDA含量的影响CK: 对照; DS: 干旱处理。CK: control; DS: drought stress.Fig. 5 Effects of drought stress on protective enzyme activities and MDA content in different common bean cultivars
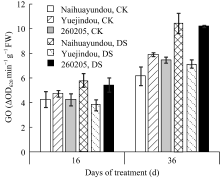
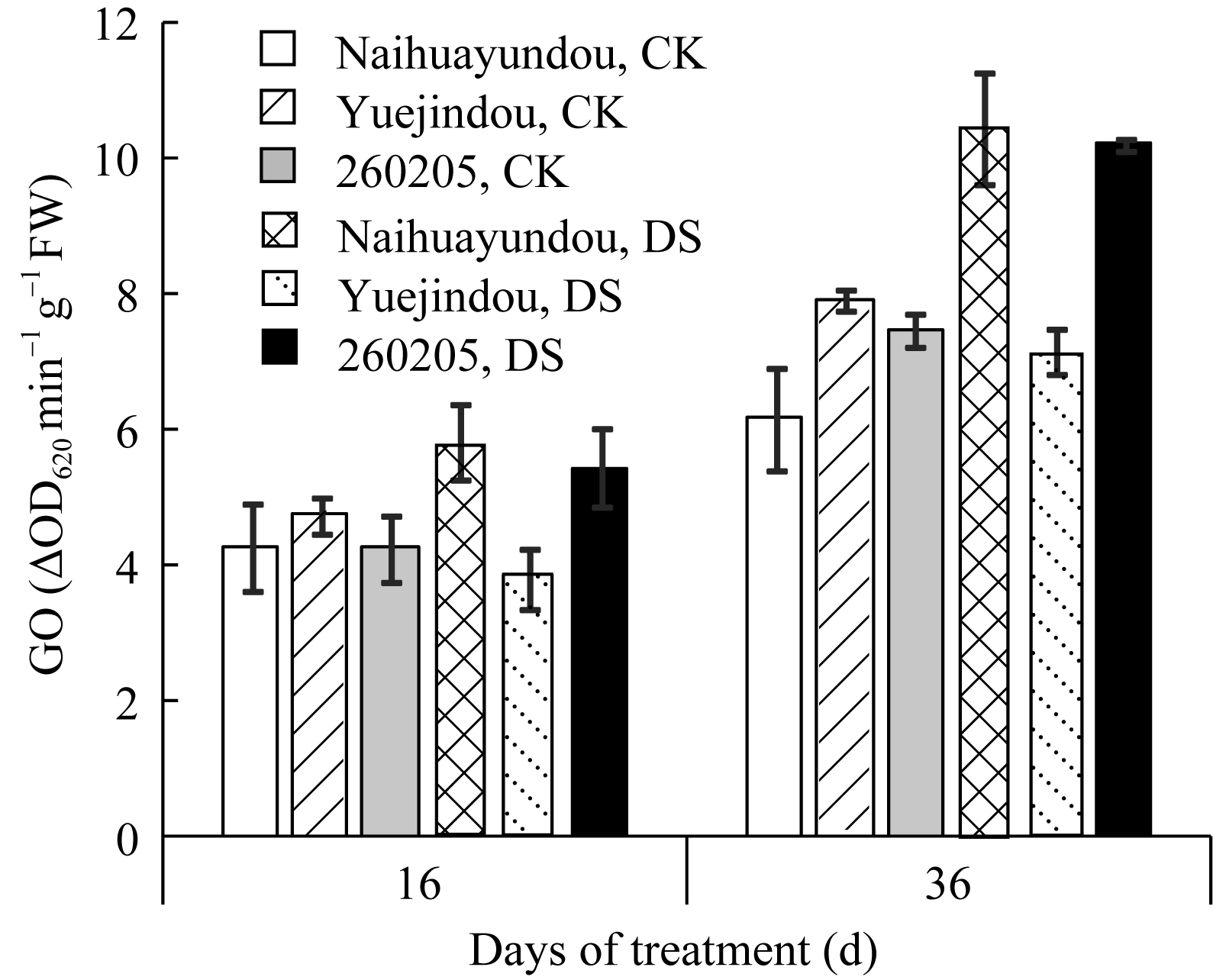
图6 干旱胁迫对不同抗旱性品种乙醇酸氧化酶活性的影响CK: 对照; DS: 干旱处理。CK: control; DS: drought stress.Fig. 6 Effects of drought stress on glycolate oxidase activity in different common bean cultivars
4 结论普通菜豆抗旱性是多种生理调节机制协同作用的结果, 包括光合产物的有效分配, 良好的气孔调节及较高的WUE, 抗氧化酶与光呼吸对有害物质的清除和抑制, 脯氨酸及可溶性糖的渗透调节等。利用单株产量、单株粒数、单株荚重和单荚粒数可以综合评价普通菜豆全生育期抗旱性, 提高抗旱种质筛选的准确性; WUE、脯氨酸含量、叶片RWC、MDA含量以及GO活性在不同的普通菜豆品种间具有稳定的差异, 也可以作为抗旱性评价指标运用于科学研究和育种工作。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Franca M G C, Thi A T O, PimentelC, Rossiello R O P, FodilY Z, LaffrayD. Differences in growth and water relations among Phaseolus vulgaris cultivars in response to induced drought stress. Environ Exp Bot, 2000, 43: 227-237[本文引用:1][JCR: 2.578]
[4]
SinghS P. Broadening the genetic base of common bean cultivars: a review. Crop Sci, 2001, 41: 1659-1675[本文引用:1][JCR: 1.513]
[5]
JonesP G, ThorntonP K. The potential impacts of climate change on maize production in Africa and Latin America in 2055. Global Environ Change, 2003, 13: 51-59[本文引用:1][JCR: 5.236]
[6]
SadeghipourO, AghaeiP. Response of common bean (Phaseolus vulgaris L. ) to exogenous application of salicylic acid (SA) under water stress conditions. Adv Environ Biol, 2012, 6: 1160-1168[本文引用:1]
[7]
CortésA J, ThisD, ChavarroC, MadriñánS, BlairM W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L. ). Theor Appl Genet, 2012, 125: 1069-1085[本文引用:1][JCR: 3.658]
[8]
VelikovaV, YordanovI, TsonevT. Plant responses to drought, acclimation, and stress tolerance. Photosynthetica, 2000, 38: 171-186[本文引用:1][JCR: 0.862]
[9]
ShaoH B, ChuL Y, JaleelC A, ManlvannanP, PanneerselvamR, ShaoM A. Understand ing water deficit stress-induced changes in the basic metabolism of higher plants-biotechnologically and sustainably improving agriculture and ecoenvironment in arid regions of the globe. Crit Rev Biotech, 2009, 29: 131-151[本文引用:1]
[10]
SimsekM, ComlekciogluN, OzturkI. The effects of the regulated deficit irrigation on yield and some yield components of common bean (Phaseolus vulgaris L. ) under semi-arid conditions. Afr J Biotechnol, 2011, 10: 4057-4064[本文引用:1][JCR: 0.573]
[11]
RosalesM A, OcampoE, Rodríguez-ValentínR, Olvera-CarrilloY, Acosta-GallegosJ, CovarrubiasA A. Physiological analysis of common bean (Phaseolus vulgaris L. ) cultivars uncovers characteristics related to terminal drought resistance. Plant Physiol Biochem, 2012, 56: 24-34[本文引用:4]
[12]
WentworthM, MurchieE H, GrayJ E, VillegasD, PastenesC, PintoM, HortonP. Differential adaptation of two varieties of common bean to abiotic stress. J Exp Bot, 2006, 57: 699-709[本文引用:1][JCR: 5.242]
[13]
MohamedM F, Schmitz-EibergerN, KeutgenN, NogaG. Comparative drought postponing and tolerance potentials of two tepary bean lines in relation to seed yield. Afr Crop Sci J, 2005, 13: 49-60[本文引用:1]
[14]
王述民, 张亚芝, 魏淑红. 普通菜豆种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006. pp50-52WangS M, ZhangY Z, WeiS H. Descriptors and data stand ard for common bean (Phaseolus vulgaris L. ). Beijing: China Agriculture Press, 2006. pp50-52(in Chinese)[本文引用:1]
[15]
张仁和, 郭东伟, 张兴华, 路海东, 刘建超, 李凤艳, 郝引川, 薛吉全. 吐丝期干旱胁迫对玉米生理特性和物质生产的影响. 作物学报, 2012, 32: 1884-1890ZhangR H, GuoD W, ZhangX H, LuH D, LiuJ C, LiF Y, HaoY C, XueJ Q. Effects of drought stress on physiological characteristics and dry matter production in maize silking stage. Acta Agron Sin, 2012, 38: 1884-1890 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[16]
徐晨, 凌风楼, 徐克章, 武志海, 刘晓龙, 安久海, 赵兰坡. 盐胁迫对不同水稻品种光合特性和生理生化特性的影响. 中国水稻科学, 2013, 27: 280-286XuC, LingF L, XuK Z, WuZ H, LiuX L, AnJ H, ZhaoL P. Effect of salt stress on photosynthetic characteristics and physiological and biochemical traits of different rice varieties. Chin J Rice Sci, 2013, 27: 280-286 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[17]
邱鹏程, 张闻博, 李灿东, 蒋洪蔚, 刘春燕, 范冬梅, 曾庆力, 胡国华, 陈庆山. 利用选择导入系分析大豆芽期和苗期耐旱性的遗传重叠. 作物学报, 2011, 37: 477-483QiuP C, ZhangW B, LiC D, JiangH W, LiuC Y, FanD M, ZengQ L, HuG H, ChenQ S. Genetic overlap of drought- tolerance loci between germination stage and seedling stage analyzed using introgression lines in soybean. Acta Agron Sin, 2011, 37: 477-483 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
张志良. 植物生理学实验指导. 北京: 高等教育出版社, 1990. pp 208-209, 125-126, 103-104, 100-101, 218-219, 227-229ZhangZ L. Hand book of Plant Physiology Experiment. Beijing: Higher Education Press, 1990. pp 208-209, 125-126, 103-104, 100-101, 218-219, 227-229(in Chinese)[本文引用:1]
[19]
李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2003. pp213-214LiH S. Principle and Technology of Plant in Physiology Biochemical Experiment. Beijing: Higher Education Press, 2003. pp213-214(in Chinese)[本文引用:1]
[20]
Acosta-GallegosJ A, AdamsM W. Plant traits and yield stability of dry bean (Phaseolus vulgaris L. ) cultivars under drought stress. J Agric Sci, 1991, 117: 213-219[本文引用:1][JCR: 2.041]
[21]
Ramirez-VallejoP, KellyJ D. Traits related to drought resistance in common bean. Euphytica, 1998, 99: 127-136[本文引用:1][JCR: 1.643]
[22]
AmedeT, SchubertS, StahrK. Mechanisms of drought resistance in grain legumes: I. Osmotic adjustment. Ethiopian J Sci, 2003, 26: 37-46[本文引用:1]
[23]
Acosta-GallegosJ A, Kohashi-ShibataJ. Effect of water stress on growth and yield of indeterminate dry beans (Phaseolus vulgaris L. ) cultivar. Field Crops Res, 1989, 20: 81-93[本文引用:1][JCR: 2.474]
[24]
Cuéllar-OrtizS M, Arrieta-MontielM P, Acosta-GallegosJ, CovarrubiasA A. Relationship between carbohydrate partioning and drought resistance in common bean. Plant Cell Environ, 2008, 31: 1399-1409[本文引用:1][JCR: 5.135]
[25]
ChavesM M, FlexasJ, PinheiroC. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann Bot, 2009, 103: 551-560[本文引用:2][JCR: 0.657]
[26]
BotaJ, FlexasJ, MedranoH. Is photosynthesis limited by decreased rubisco activity and RuBP content under progressive water stress?New Phytol, 2004, 162: 671-681[本文引用:1][JCR: 6.736]
[27]
云建英, 杨甲定, 赵哈林. 干旱和高温对植物光合作用的影响机制研究进展. 西北植物学报, 2006, 26: 641-648YunJ Y, YangJ D, ZhaoH L. Research progress in the mechanism for drought and high temperature to affect plant photosynthesis. Acta Bot Boreal-Occident Sin, 2006, 26: 641-648 (in Chinese with English abstract)[本文引用:1]
[28]
吴永美, 吕炯章, 王书建, 李润植. 植物抗旱生理生态特性研究进展. 杂粮作物, 2008, 28(2): 90-93WuY M, LüJ Z, WangS J, LiR Z. Research progress on eco-physiological responses of plants to drought conditions. Rain Fed Crops, 2008, 28(2): 90-93 (in Chinese)[本文引用:1][CJCR: 0.5562]
[29]
TurkanI, BorM, OzdemirF, KocaH. Differential responses of lipid peroxidation and antioxidants in the leaves of drought tolerant P. acutifolius gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci, 2005, 168: 223-231[本文引用:1][JCR: 2.922]
[30]
金怡, 刘合芹, 汪得凯, 陶跃之. 植物光呼吸分子机制研究进展. 中国农学通报, 2011, 27(3): 232-236JingY, LiuH Q, WangD K, TaoY Z. The progress of molecular mechanisms of photorespiration in plants. Chin Agric Sci Bull, 2011, 27(3): 232-236[本文引用:1]
[31]
杜伟莉, 高杰, 胡富亮, 郭德林, 张改生, 张仁和, 薛吉全. 玉米叶片光合作用和渗透调节对干旱胁迫的响应. 作物学报, 2013, 39: 530-536DuW L, GaoJ, HuF L, GuoD L, ZhangG S, ZhangR H, XueJ Q. Responses of drought stress on photosynthetic trait and osmotic adjustment in two maize cultivars. Acta Agron Sin, 2013, 39: 530-536[本文引用:1][CJCR: 1.667]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}