 ,, 李美娜,广州大学生命科学学院,广州 510006
,, 李美娜,广州大学生命科学学院,广州 510006Progress on genic male sterility gene in soybean
Xiaoyuan Sun, Yifan Wang, Yunhui Wang, Jiayu Lin, Jinhong Li, Yuantao Qiu, Xiaolong Fang, Fanjiang Kong,, Meina Li,School of Life Sciences, Guangzhou University, Guangzhou 510006, China通讯作者: 孔凡江,博士,教授,研究方向:大豆分子育种。E-mail:kongfj@gzhu.edu.cn李美娜,博士,副教授,研究方向:大豆杂种优势利用和生物钟与环境互作。E-mail:limeina@gzhu.edu.cn
编委: 王秀娥
收稿日期:2020-10-23修回日期:2020-12-7网络出版日期:2021-01-20
| 基金资助: |
Received:2020-10-23Revised:2020-12-7Online:2021-01-20
| Fund supported: |
作者简介 About authors
孙小媛,在读硕士研究生,专业方向:植物资源利用。E-mail:

摘要
雄性不育是指植物雄蕊不能正常生长和产生有活力花粉粒的现象。利用雄性不育突变体开展杂交育种工作,是快速提高作物单产的有效途径。目前,通过杂种制种已大幅度提高了水稻(Oryza sativa L.)、玉米(Zea mays L.)和小麦(Triticum aestivum L.)等作物的产量。大豆(Glycine max (L.) Merr.)作为自花授粉作物,通过人工去雄生产杂交种子不仅困难而且经济上不可行。由于适用于杂交种生产的不育系资源短缺,目前大豆还没有实现大规模杂种优势利用。因此,快速实现大豆杂种优势利用迫切需要鉴定稳定的大豆雄性不育系统。本文总结了大豆细胞核雄性不育(genic male sterility, GMS)突变体及不育基因研究进展,同时结合拟南芥(Arabidopsis thaliana)、水稻和玉米中已报道的细胞核雄性不育基因,从反向遗传学的角度,为大豆核雄性不育基因的鉴定提供依据。
关键词:
Abstract
Male sterility refers to the phenomenon that stamens cannot grow normally and produce viable pollen grains in plants. Hybrid seed production by taking advantage of the trait of male sterility is an effective and quick strategy to increase crop yield. Up to date, the yield of rice (Oryza sativa L.), maize (Zea mays L.), wheat (Triticum aestivum L.) and other crops has been greatly increased based on hybrid vigor utilization. Soybean (Glycine max (L.) Merr.) is a self-pollination species, artificial emasculation is not only time-consuming, but also labor-intensive and economically impracticable. So far, large scale hybrid breeding has not been performed in soybean due to the shortage of male sterile lines suitable for hybrid production. Therefore, it is urgent to identify a stable male sterile system for the rapid utilization of heterosis in soybean. In this review, we summarize the progress on the discovery of soybean genic male sterility (GMS) mutants and GMS genes. Combining with the investigation of GMS genes in Arabidopsis, rice and maize, we provide important insights into the identification and potential utilization of GMS genes in soybean in the perspective of reverse genetics.
Keywords:
PDF (1204KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孙小媛, 王一帆, 王韫慧, 蔺佳雨, 李金红, 丘远涛, 方小龙, 孔凡江, 李美娜. 大豆细胞核雄性不育基因研究进展. 遗传[J], 2021, 43(1): 52-65 doi:10.16288/j.yczz.20-357
Xiaoyuan Sun.
大豆(Glycine max (L.) Merr.)原产于中国,是重要的粮食作物和饲用原料。但我国大豆耕种面积逐年缩减、产量增长缓慢,单产水平远远低于世界平均水平,无法满足人们对大豆不断增长的需求。因此,近年来我国大豆超过85%依赖进口,2019年全国大豆进口量达到8851万吨,进口国家主要为巴西和美国。因此,快速提高大豆产量已成为目前我国大豆生产中最迫切的任务。
杂种优势利用是大幅度提高作物单产的有效途径。利用杂种优势,水稻增产55%,小麦增产25%,大麦(Hordeum vulgare L.)增产11%,谷子(Setaria italica)增产68%[1,2]。因杂交植物具有较好的适应性、较强的抗逆性,因此对提高农业产量做出了巨大贡献。大豆是目前少数几个没有大规模利用杂种优势的作物之一,其主要原因是适于杂交种生产的不育系资源短缺。
杂交种子的产生需要具有遗传多样性的亲本异花授粉,因此利用雄性不育和雌性可育的突变体是进行异花授粉、实现大规模杂交制种的前提。大豆是典型的自花授粉作物,通过人工去雄生产杂交种子不仅困难而且经济上不可行。因此,快速实现大豆杂种优势利用迫切需要在大豆中鉴定稳定的雄性不育系统[3]。
研究发现,雄性不育由细胞核基因或线粒体基因控制,前者被称为细胞核雄性不育(nuclear male sterility, NMS或genic male sterility, GMS),后者被称为细胞质雄性不育(cytoplasmic male, CMS)[4,5]。杂交育种技术中的三系法利用CMS系为母本,以保持系为父本繁殖大量保持不育特性的种子;以CMS系为母本,以恢复系为父本,实现大规模生产可育的杂交种子。但三系法也存在缺陷,如恢复系的种质资源狭窄、CMS系与恢复系的遗传多样性差,限制了CMS系的进一步发展[6,7,8,9]。两系法利用光温敏GMS系,如籼稻安农S-1[10],当处于低温环境时可育,而在高温环境育性转变为不育,以常规品种作为恢复系,同时改变光温条件,可实现不育系繁殖和杂交种生产。但随着研究的深入,两系法的问题也日益突出,不育系的育性受光照和温度的影响,随着繁殖代数的增加,育性转变的临界温度发生改变,大大增加了繁殖不育系的难度[6,7,8,9]。随着现代生物技术的发展,研究人员通过分子生物技术和基因工程手段,在水稻中通过利用核不育基因、花粉自我降解基因、红色荧光蛋白筛选标记基因等,获得转基因保持系,利用该保持系繁殖大量非转基因的雄性不育种子可用于商业生产[11]。因此在大豆中利用核不育基因并以此构建雄性不育系统是快速实现大豆杂种优势利用的重要途径。
虽然大豆核雄性不育突变体的发现至今已经有90多年的历史,但是目前仅有ms4基因被克隆,其他核不育基因尚未见报道[12]。对大豆核不育基因相关研究进展进行系统总结有助于深入认识和了解大豆核不育系统,鉴定并挖掘新的雄性不育基因也有助于开发稳定的可用于大豆杂交种子生产的不育系统。本文对大豆核雄性不育基因的相关研究进行了综述,并通过比较拟南芥、水稻和玉米等模式植物中已报道的雄性不育基因,以期为大豆核雄性不育基因的鉴定和研究提供参考。
1 大豆细胞核雄性不育突变体研究进展
1.1 大豆细胞核雄性不育突变体
细胞核雄性不育,其雄性育性大多由细胞核内一对隐性基因控制,少数受显性基因调控,符合孟德尔遗传定律。目前已鉴定出28个大豆细胞核雄性不育突变体,包括雄性和雌性配子都可育的结构型不育、部分雄性不育、雄性和雌性均不育、雄性不育和雌性可育、光敏雄性不育和由单一显性基因控制的雄性不育(表1)。Table 1
表1
表1大豆核雄性不育突变体及基因信息
Table 1
| 大豆GMS 突变体 | 基因 | 基因位置 | 雄性不育类型 | 来源 | 文献 | ||
|---|---|---|---|---|---|---|---|
| 染色体 | 连锁群 | 标记 | |||||
| fs1fs2 | fs1fs2 | - | - | - | 结构型雄性不育 | 自然 | [13] |
| msp | msp | 2 | D1b | GMES4176-Sat_069 (Satt172) | 雄性不育,雌性可育 | 自然 | [14,15] |
| St1 | St1 | - | - | - | 雄性不育,雌性不育 | 自然 | [16,17] |
| St2 | St2 | 11 | B1 | BARCSOYSSR_11_122- BARCSOYSSR_11_137 | 雄性不育,雌性不育 | 自然 | [18,34] |
| St3 | St3 | - | - | - | 雄性不育,雌性不育 | 自然 | [18] |
| St4 | St4 | 1 | D1a | Satt436-Satt468 | 雄性不育,雌性不育 | 自然 | [18,34] |
| St5 | St5 | 13 | F | Satt030-Satt146 | 雄性不育,雌性不育 | 自然 | [19,34] |
| St6 | St6 | 14 | B2 | BARCSOYSSR_14_84- BARCSOYSSR_14_109 | 雄性不育,雌性不育 | 自然 | [18,34] |
| St7 | St7 | 2 | D1b | Satg001-端粒 | 雄性不育,雌性不育 | 自然 | [18,34] |
| w4-mutable line | St8 | 16 | J | Satt132-Satt414 | 雄性不育,雌性不育 | 自然 | [20,36] |
| ms1 | ms1 | 13 | F | Satt516-Satt595 | 雄性不育,雌性可育 | 自然 | [21] |
| ms2 | ms2 | 10 | O | Sat_190-Scaa001 | 雄性不育,雌性可育 | 自然 | [23,41] |
| ms3 | ms3 | 2 | D1b | Satt157-Satt542 | 雄性不育,雌性可育 | 自然 | [24] |
| ms4 | ms4 | 2 | D1b | Glyma.02G243200 | 雄性不育,雌性可育 | 自然 | [12] |
| ms5 | ms5 | 11 | B1 | BARCSOYSSR_11_0122-Sat_270 | 雄性不育,雌性可育 | 诱变 | [26,42] |
| ms6 | ms6 | 13 | F | Satt149-Satt030 | 雄性不育,雌性可育 | 自然 | [27] |
| ms7 | ms7 | 9 | - | - | 雄性不育,雌性可育 | 诱变 | [14,28] |
| ms8 | ms8 | 7 | M | 端粒-Sat_389 | 雄性不育,雌性可育 | 自然 | [15,28] |
| ms9 | ms9 | 3 | N | Satt521-Satt237 | 雄性不育,雌性可育 | 自然 | [28,37] |
| NJ89-1 | ms0 | - | - | - | 雄性不育,雌性可育 | 自然 | [30] |
| 88-428-BY | - | - | - | - | 光敏雄性不育 | 自然 | [31] |
| N7241S | - | - | - | - | 雄性不育,雌性可育 | 自然 | [32] |
| NJS-13H | msNJ | 10 | O | BARCSOYSSR_10_794- BARCSOYSSR_10_819 | 雄性不育,雌性可育 | 自然 | [39] |
| msMOS | - | 2 | D1b | Satt157-Satt698 | 雄性不育,雌性可育 | 自然 | [40] |
| D8804-7 | Introducing exogenous DNA | - | - | - | 雄性不育,雌性不育 | 导入外源DNA | [43] |
| NJS-1H | - | - | - | - | 雄性不育,雌性不育 | 化学诱变 | [44] |
| Wh921 | - | - | - | - | 雄性不育,雌性可育 | 自然 | [45] |
| St-M | Mst-M | 13 | F | W1(Glyma.13G072100)-Satt516 | 雄性不育,雌性可育 | 自然 | [46] |
新窗口打开|下载CSV
fs1fs2是迄今为止在大豆中报道的唯一自然产生的结构型不育突变体,其花粉育性正常,但胚珠发育异常,受双隐性基因fs1和fs2共同调控。fs1fs2通常带有外露的花柱和柱头,花瓣发育异常,阻止花丝的伸长,导致花药与柱头之间距离较远,阻止自花授粉,因此结实率较低[13]。mspmsp是温度敏感型雄性不育,高温环境下雄性生殖力较强[14],其突变体的花药外观各不相同,有的花药为棕色,较小;花药通常开裂,退化的小孢子或花粉凝集成块[15]。St1、St2、St3、St4、St5、St6、St7和St8 (w4-mutable line)属雄性和雌性都不育的突变体,其中St2、St3是染色体联会缺失的突变体,而St4、St5、St6、St7和St8是染色体联会异常的突变体,这类不育系的花粉和胚珠都败育[16,17,18,19,20]。
1971年,Brim和Young在大豆中发现了控制雄性不育性状的突变体ms1[21]。迄今为止,已经发现雄性不育雌性可育的突变体包括ms1、ms2、ms3、ms4、ms5、ms6、ms7、ms8、ms9和NJ89-1。突变体ms1在第二次减数分裂末期细胞质分裂失败,结果产生多核小孢子,最终形成的花粉粒粘附在一起,不能萌发[22]。ms2和ms3均能进行正常的减数分裂,形成四分体,但由于绒毡层发育异常,小孢子随后退化,最终花药中没有产生花粉粒[23,24]。ms4与ms1相似,都是第二次减数分裂末期后不发生胞质分裂引起的败育,但ms4花药略小且偏白色,形成中空小孢子[25]。ms5突变体在花粉发育过程中绒毡层空泡化,小孢子解体,花粉增大[26]。ms6突变体小孢子母细胞在第二次减数分裂末期后退化,药室内壁和绒毡层等发育呈现异常,并且花朵相比其他突变体较小[27]。ms7小孢子的败育主要由于四分体时期胼胝质不解体,结果没有产生花粉粒。ms8育性受环境温度的影响,和ms9相似,结果都产生大量的退化花粉粒,但ms9花粉没有萌发沟[14,28,29]。而NJ89-1小孢子母细胞在第一次减数分裂的早前期开始发生败育,染色体联会异常,结果产生异常的多分体(polyad),其雄性育性受隐性基因ms0调控 [30]。
光敏雄性不育系88-428-BY在短日照条件下,小孢子发育减数分裂过程中染色体行为异常,最终形成异常花粉粒,导致雄性不育[31]。N7241S是目前在大豆中报道的唯一由显性基因调控的雄性不育、雌性可育突变体,该突变体的小孢子四分体表现正常,而成熟花粉粒细胞质分布不均,发生皱缩使花粉完全败育[32]。
以上不育系由于花器官结构异常或大量可育株的存在等问题,很难应用于杂交种生产。目前在优势大豆选育工作中较多使用ms0~ms9核不育突变体材料[33]。
1.2 大豆细胞核雄性不育基因定位与克隆
大多数核不育基因(以下简称GMS基因)已被初步定位到大豆染色体上。控制大豆雄性和雌性均不育的GMS突变体中,St2基因位于11号染色体BARCSOYSSR_11_122-BARCSOYSSR_11_137标记之间,物理距离为~398 kb,包含57个预测基因[34]。St4基因位于1号染色体Satt436-Satt468标记之间,物理距离约为164 kb,包含17个预测基因,该区域中的Glyma.01G159200基因编码具有RWP-RK结构域(RKD)的转录因子,已知RKD转录因子在雄配子发生、卵细胞功能控制和细胞分裂中发挥重要作用[35],因此该基因可能是St4的候选基因。St5位于13号染色体Satt146-Satt030之间[34],St6在14号染色体BARCSOYSSR_14_84-BARCSOYSSR_14_109标记之间,St7在2号染色体Satg001和端粒之间[34]。St8位于16号染色体分子连锁群J上的SSR标记Satt132-Satt414之间,与两个标记的距离分别为7.8 cM和3.4 cM[36]。在大豆中报道的控制雄性不育雌性可育突变体的候选基因(ms1、ms2、ms3、ms4、ms5、ms6、ms7、ms8、ms9、msMOS、msp和msNJ)分别定位到大豆的7条不同染色体上[37,38,39]。其中,ms1和ms6基因定位于13号染色体[38]。ms2基因位于10号染色体Sat_190-Scaa001标记之间,与两个标记距离分别为6.9和9.0 cM[37]。ms3、ms4、msMOS和msp均定位在2号染色体上[15,38,40]。ms1、ms3、ms4和ms6基因分别接近Satt516、Satt157、Satt542 BARCSOYSSR_ 02_1539和AW186493标记[38,40]。ms5、ms7、ms8和ms9分别位于11、9、7、3号染色体上[15,26,37,38]。msNJ定位在10号染色体SSR标记BARCSOYSSR_ 10_794-BARCSOYSSR_10_819之间[39]。虽然大豆核雄性不育基因的挖掘与定位已被广泛研究和报道,但目前仅有ms4基因被克隆。Thu等[12]将ms4位点精细定位到216 kb的区域,该区域包含23个基因,通过测序发现其中Glyma.02G243200第3外显子有一个碱基插入,导致蛋白编码提前终止。利用拟南芥同源基因mmd1突变体与大豆Glyma.02G243200进行功能互补验证,证明Glyma.02G243200编码大豆MS4蛋白,该蛋白是具有PHD结构域的转录因子。该研究首次通过图位克隆的方法定位到大豆核雄性不育基因,但并没有进行深入的功能研究。
2 从反向遗传学角度,同源序列比对鉴定大豆GMS基因
基因之间在序列(蛋白质、DNA或RNA)水平上的相似度越高,往往在功能上也有很高的保守性。有研究报道,在玉米GMS基因中有20%拟南芥GMS同源基因和30%水稻GMS同源基因,在调控雄性不育方面具有相似的功能[47]。相比于大豆,近几十年来在拟南芥和水稻中发现了更多的GMS基因,分别鉴定到至少40个GMS基因[47]。本文根据拟南芥、水稻和玉米GMS基因,在大豆中找到了122个氨基酸相似性大于40%的同源基因(表2),同时结合公共数据库中大豆不同组织中的基因表达数据(http://venanciogroup.uenf.br/resources/),通过聚类分析将这122个基因分成3类(I、II、III),其中第一类可进一步分为两个子类(图1)。Table 2
表2
表2拟南芥、水稻和玉米GMS基因与大豆同源基因
Table 2
| 拟南芥GMS基因 | 水稻GMS基因 | 玉米GMS基因 | 大豆同源基因 |
|---|---|---|---|
| Ms1 | PTC1 | ms7 | Glyma.01G047400、Glyma.02G107600 |
| Ms2 | DPW | ms6021 | Glyma.02G169100 |
| TDF1 | OsTDF1 | ms9 | Glyma.19G017900、Glyma.13G066600 |
| DYT1 | UDT1 | ms32 | Glyma.02G059800、Glyma.15G064000、Glyma.16G142600 |
| CYP704B1 | OsCYP704B2 | ms26 | Glyma.07G043300 |
| CYP703A2 | OsCYP703A3 | apv1 | Glyma.12G160400、Glyma.13G260500 |
| TPD1 | MIL2/OsTDL1a | mac1 | Glyma.04G198200、Glyma.06G167300、Glyma.08G010200 |
| TGA10 | OsTGA10 | Glyma.03G142400 | |
| EMS1 | MSP1 | Glyma.20G156700 | |
| MYB33 | OsGAMYB | Glyma.13G187500、Glyma.15G225300 | |
| MYB65 | |||
| DEX1 | OsDEX1 | Glyma.03G076900、Glyma.01G111300 | |
| AMS | TDR | Glyma.10G281800 | |
| ACOS5 | OsACOS12 | Glyma.08G329700 | |
| CalS5 | GSL5 | Glyma.04G213100 | |
| LAP5, PKSB | OsLAP5/OsPKS2 | Glyma.11G097900 | |
| LAP6, PKSA | OsLAP6/OsPKS1 | Glyma.01G073600 | |
| ABCG26 | OsABCG15/PDA1 | Glyma.18G074800 | |
| KNS4/UPEX1 | ms8 | Glyma.09G233700、Glyma.12G003100、Glyma.14G166300、 Glyma.13G084600、Glyma.04G254100 | |
| MMD1 | Glyma.02G243200(MS4)、Glyma.14G212300 | ||
| TGA9 | Glyma.12G184500、Glyma.13G316900, Glyma.12G088700、Glyma.11G183700 | ||
| MYB80 | Glyma.08G336500、Glyma.18G071600 | ||
| ATGPAT1 | Glyma.14G028300、Glyma.02G286500 | ||
| AtGPAT6 | Glyma.18G197800、Glyma.07G146800、Glyma.01G113200 | ||
| TEK | Glyma.02G285500 | ||
| ARF17 | Glyma.04G254200 | ||
| RBOHE | Glyma.15G182000、Glyma.09G073200 | ||
| CDKG1 | Glyma.05G145800、Glyma.08G102600 | ||
| AtSK32 | Glyma.07G076700、Glyma.03G016500 | ||
| LBD10 | Glyma.04G106400、Glyma.11G058400、Glyma.01G183900 | ||
| LBD27 | Glyma.08G192600 | ||
| ROXY1 | Glyma.16G052800 | ||
| ROXY2 | Glyma.19G096600 | ||
| MYB26 | Glyma.07G008500 | ||
| RPG1 | Glyma.19G009900 | ||
| DRL1/TKPR1 | Glyma.15G018500 | ||
| NEF1 | Glyma.18G298700 | ||
| CER3/FLP1/WAX2 | Glyma.17G069100、Glyma.13G091200 | ||
| CDM1 | Glyma.10G269300、Glyma.20G121800 | ||
| NPU | Glyma.18G215000 | ||
| TES | Glyma.07G096500、Glyma.09G181200、Glyma.13G114200、 Glyma.17G045600 | ||
| MIL1 | ms22 | Glyma.16G052800、Glyma.19G096600、Glyma.05G142700 | |
| TIP2/bHLH142 | ms23 | Glyma.16G090900、Glyma.08G089100、Glyma.08G269300 | |
| OsNP1 | IPE1 | Glyma.05G050100、 | |
| OsGPAT3 | ms33 | Glyma.10G011000、Glyma.03G221100、Glyma.19G218100、 Glyma.02G010600、Glyma.03G221300、Glyma.14G167300、 Glyma.14G167400、Glyma.13G085700 | |
| OsSTRL2 | ms45 | Glyma.13G367400、Glyma.15G005900、Glyma.15G029300 | |
| OsG1 | Glyma.11G095100 | ||
| OsC6 | Glyma.20G146800 | ||
| OsABCG26 | Glyma.09G160000、Glyma.16G209400 | ||
| OsDPW2 | Glyma.19G096100 | ||
| EAT1/DTD | Glyma.05G134400 | ||
| OsFIGNL1 | Glyma.19G071600 | ||
| WDA1 | Glyma.07G121500、Glyma.03G101700、 Glyma.03G101800、 Glyma.07G114200、Glyma.03G101200 | ||
| OsFTIP7 | Glyma.19G145400、Glyma.03G142500 | ||
| OsDTC1 | Glyma.18G207500 | ||
| OsDTM1 | Glyma.04G102200、Glyma.06G103500 | ||
| OsCSA | Glyma.01G049600 | ||
| MTR1 | Glyma.05G181300、Glyma.08G139000 | ||
| OsRAFTIN | Glyma.06G081100 | ||
| OsUAM3 | Glyma.02G120900、Glyma.01G063000 | ||
| OsGT1 | Glyma.06G029300、Glyma.17G253700 | ||
| CAP1 | Glyma.17G194400 | ||
| OsADF | Glyma.02G294800 | ||
| OsAPI5 | Glyma.07G203600 | ||
| Ocl4 | Glyma.01G014600、Glyma.09G207500 | ||
| IG1 | Glyma.U012500、Glyma.15G228100、Glyma.13G191000 | ||
| ms30 | Glyma.02G273900 | ||
| Ms44 | Glyma.09G144000 |
新窗口打开|下载CSV
图1
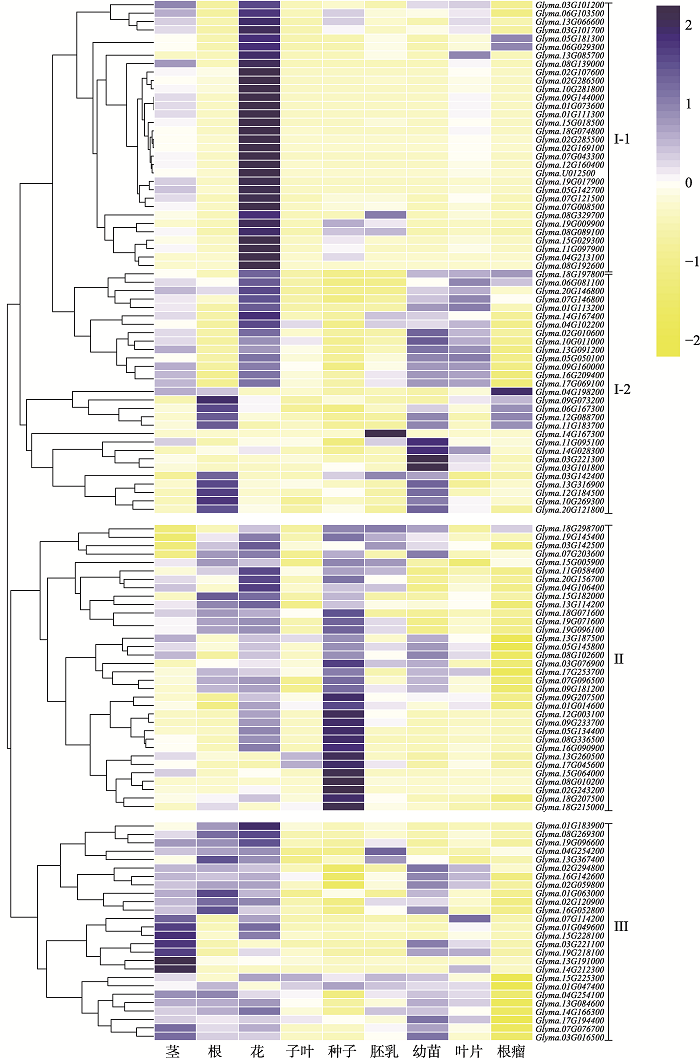 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1大豆同源GMS基因在不同组织中表达聚类热图
不同颜色深浅表示不同基因在不同组织表达量的高低,蓝色越深则表达量越高,黄色越深则表达量越低;I:I-1类基因在花中优势表达,I-2类基因在子叶和胚乳中几乎不表达;II:在根瘤中几乎不表达的一类基因;III:在子叶、胚乳和根瘤中几乎不表达的一类基因。
Fig. 1Clustered heat map of expressions of soybean homologs of GMS genes in different tissues
32个I类(I-1)基因在花中优势表达,包括拟南芥ATGPAT1[48]、DRL1/TKPR1[49]、MYB26[50]、TEK[51]、CYP704B1[52]、CYP703A2[53]、PKSA/LAP6[54]、DEX1[55]、RPG1[56]、LBD27[57]和AMS[58],水稻OsTDF1[59]、OsABCG15/PDA1[60]、PTC1[61]、OsPKS2/OsLAP5[62]、OsACOS12[63]、MTR1[64]、TIP2/bHLH142[65]、OsSTRL2[66]、OsGT1[67]和GSL5[68],玉米IG1、MS6021、MS33、MS44、MS23等大豆同源基因[47]。其中绝大部分GMS基因参与了脂质代谢过程,如拟南芥MS2、CYP704B1和CYP703A分别编码脂肪酰基辅酶A合成酶(ACOS)、长链脂肪酸ω-羟化酶(CYP704B)和细胞色素P450脂肪酸羟化酶(CYP703A),在孢粉素合成和花粉外壁形成中扮演重要角色;一旦发生突变,将导致乌氏体形成缺陷,花粉壁发育不完整;它们分别与水稻OsDPW、OsCYP704B2和OsCYP703A3及玉米MS6021、MS26和APV1同源[69]。参与脂质代谢的基因中也包含一些重要转录因子,其中拟南芥MS1[70]与水稻PTC1和玉米MS7[71]同源,编码具有PHD结构域的转录因子,调控绒毡层细胞程序性死亡及花药晚期发育。拟南芥AMS和水稻TDR[65]同源,编码bHLH转录因子,形成DYT1-TDF1-AMS-Ms188- Ms1遗传调控网络,影响绒毡层细胞发育和减数分裂后小孢子形成过程[58,72]。玉米MS23是花药体细胞层分化所必须的转录因子,其在水稻中同源基因TIP2/bHLH142编码的转录因子在绒毡层细胞程序性死亡和花粉发育中起着关键作用[73]。
29个I类基因(I-2)在子叶和胚乳中几乎不表达。34 个II类基因在胚乳和花中的表达量较高,在子叶和根瘤中几乎不表达。26个III类基因在不同组织均有表达,但大多数基因在子叶、胚乳和根瘤中几乎不表达。综上所述,I类基因(I-1)更有可能是大豆中的GMS基因。
利用大豆中122个可能的GMS基因,结合大豆中已经报道的GMS基因所在的区间,本文将部分基因绘制在大豆染色体上(图2)。结果显示,基因Glyma.02G107600和Glyma.02G120900在ms3位点所在的区间内,其中拟南芥MS1[70]与Glyma.02G107600同源,在花粉发育过程中影响绒毡层发育,与大豆ms3突变体表型相似。基因Glyma.10G269300和Glyma.10G281800在ms2位点所在的区间内,其中Glyma.10G269300与拟南芥CDM1[74]同源,在小孢子形成过程中调控胼胝质代谢,影响花粉外壁形成;而Glyma.10G281800与拟南芥AMS、水稻TDR基因同源,编码bHLH转录因子,调控绒毡层细胞发育和减数分裂后小孢子形成,ams突变体产生膨大绒毡层和败育小孢子,tdr突变体绒毡层退化和细胞程序性死亡延迟,导致小孢子凹陷败育[58]。大豆ms2突变体也是由于绒毡层发育异常导致小孢子败育,因此Glyma.10G281800可能是大豆ms2的候选基因。13号染色体的Glyma. 13G084600、Glyma.13G085700和Glyma.13G091200在mst-M所在的区间内,其中Glyma.13G084600与拟南芥KNS4同源,KNS4基因是CAZy糖基转移酶31基因家族成员,KNS4突变导致阿拉伯半乳聚糖蛋白(AGP)/果胶聚糖合成缺陷,参与拟南芥花粉外壁结构和形态建成,从而影响花粉活力[75]。Glyma.13G085700与水稻OsGPAT3[76]同源,与绒毡层发育、脂质代谢相关。拟南芥WAX2[77]与Glyma.13G091200同源,通过调控角质层发育影响花粉育性,其中Glyma.13G084600拟南芥同源基因KNS4突变体的表型与大豆mst-M的突变体表型更为接近,可能是mst-M的候选基因。此外,Glyma.13G066600和Glyma.13G114200分别在ms6和ms1位点所在的区间内,其中Glyma.13G066600在拟南芥中的同源基因TDF1编码R2R3 MYB转录因子,在绒毡层、减数分裂细胞和小孢子中高度表达,可能是控制胼胝质降解的关键基因[78]。大豆Glyma.13G114200与拟南芥TES[79]和烟草NACK2[80]基因同源,拟南芥TES基因编码驱动蛋白,与细胞分裂时期微管运动有关,tes突变体在小孢子四分体时期胞质分裂失败,结果产生体积较大的败育花粉粒;而烟草NACK2突变体中NPK1的定位与活性受到影响,抑制了成膜体和细胞板的扩张,从而产生了具有不完整细胞板的多核细胞,该表型与大豆ms1突变体极为相似,提示Glyma.13G114200很有可能是大豆ms1的候选基因。Glyma.02G243200编码具PHD结构域的转录因子,位于ms4所在的区间,已经通过基因功能互补和CRISPR/Cas9技术,证实该基因就是大豆中的ms4[12]。
通过对以上大豆GMS基因候选基因的预测,再利用CRISPR/Cas9或RNAi等反向遗传学手段,对这些可能的大豆GMS基因进行功能验证,以筛选大豆完全雄性不育系,有望创造出更多有价值的大豆雄性不育材料用于杂交育种。
图2
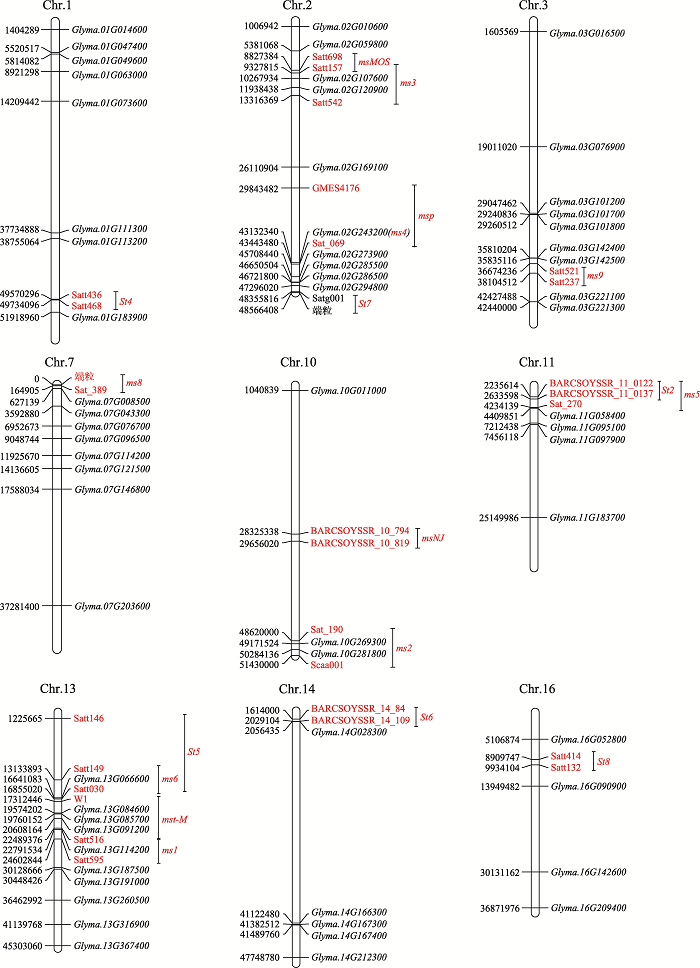 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2大豆不育位点及GMS同源基因所在的染色体位置图
红色斜体符号ms1、ms2、ms3、ms4、ms5、ms6、ms8、ms9、msp、msMOS、mst-M、msNJ、St2、St4、St5、St6和St8为基因;其余红色符号为分子标记。
Fig. 2Chromosomal locations of sterility locus and homologs of GMS genes in soybean
3 结语与展望
大豆中虽然已经鉴定很多细胞核雄性不育突变体,但是基因克隆和分子机理的研究报道较少。植物之间存在着雄性生殖发育调控网络的保守性和多样性[47],利用模式植物GMS基因信息进行分析是发现大豆中GMS基因的有效方法,通过鉴定这些GMS基因在大豆花药中优势表达的同源基因,再利用CRISPR/Cas9基因编辑技术和功能互补实验进行进一步的验证,可鉴定新的大豆核雄性不育基因。这不仅有助于人们了解雄性不育的分子机制和大豆生殖生物学,更重要的是可以开发稳定的能够用于杂交种子生产的大豆不育系统。大豆中很多核不育系由于其中存在大量可育植株,并且花器官结构存在异常,很难应用于杂交种的生产[81]。随着三系杂交和两系杂交育种技术在应用中显现的问题日益突出[6,7,8,9],研究人员利用现代生物技术,已开发了利用转基因手段保持或繁殖非转基因的植物核雄性不育系的方法[82]。其中包括玉米的多控不育技术、新型玉米基因工程不育制种技术和水稻的智能不育技术,综合了转基因技术、荧光蛋白筛选技术和花粉失活技术,将花粉育性恢复基因、花粉自我降解基因Zm-AA1或Dam、荧光蛋白筛选标记基因DsRed或mCherry、抗除草剂基因bar组合构建遗传转化载体,通过遗传转化将转基因元件导入隐性核不育系中,获得可育的转基因保持系,该保持系自花授粉以1∶1的比例获得可育转基因种子和不育非转基因种子,可通过荧光分选出来;非转基因不育种子不发荧光,因此可以有效解决雄性不育种子保持和繁殖的问题,以期用于大规模商业生产。
叶绿体转化和利用种子成熟后特异表达启动子可能是利用作物核雄性不育基因的有效途径[83]。通过叶绿体转化,如构建定点整合载体、利用叶绿体特异的启动子和终止子等将育性恢复基因转移到雄性不育株的胞质中,实现外源花粉育性恢复基因的表达,获得可育的保持系,该保持系可作为父本与不育系进行杂交,生产杂交种子[83,84]。另外,可以通过人工构建的启动子调控外源基因的表达,将种子成熟特异表达启动子、毒素致死基因和花粉育性恢复基因作为紧密连锁的元件通过遗传转化导入不育株,获得转基因植株。该转基因株系在早期是可育的,可通过自花授粉生产繁殖不育系,在转基因植株种子成熟后,致死基因将发挥作用,导致带有育性恢复基因的转基因种子死亡,因此后代成活的不育种子不具有转基因[84,85]。
通过现代生物技术手段,有望充分利用大豆核不育基因构建相似的大豆核雄性不育杂交系统,有利于在大豆中更快、更好的实现杂种优势利用。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.tplants.2017.10.001URLPMID:29126789 [本文引用: 1]

In many plant species, male-sterile female lines with cytoplasmic male sterility (CMS) or nuclear-controlled environment-sensitive genic male sterility (EGMS) have long been used to efficiently produce hybrids that harness hybrid vigor or heterosis. However, the underlying molecular mechanisms for these applications have only recently been uncovered in a few species. We provide here an update on the understanding of cytoplasmic-nuclear communication based on the discovery of mitochondrial CMS genes and their corresponding nuclear fertility determinants. Recent findings that uncover diverse mechanisms such as epigenetic, transcriptional, and post-transcriptional controls of EGMS by temperature and photoperiod signals are also reviewed. Furthermore, translational research that applies basic knowledge of plant male fertility control to hybrid seed production practice is highlighted.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/pbr.v138.6URL [本文引用: 1]
DOI:10.1146/annurev-arplant-050213-040119URL [本文引用: 1]
DOI:10.1016/j.mito.2014.04.004URL [本文引用: 1]
Cytoplasmic male sterility and its fertility restoration via nuclear genes offer the possibility to understand the role of mitochondria during microsporogenesis. In most cases rearrangements in the mitochondrial DNA involving known mitochondrial genes as well as unknown sequences result in the creation of new chimeric open reading frames, which encode proteins containing transmembrane domains. So far, most of the CMS systems have been characterized via restriction fragment polymorphisms followed by transcript analysis. However, whole mitochondrial genome sequence analyses comparing male sterile and fertile cytoplasm open options for deeper insights into mitochondrial genome rearrangements. We more and more start to unravel how mitochondria are involved in triggering death of the male reproductive organs. Reduced levels of ATP accompanied by increased concentrations of reactive oxygen species, which are produced more under conditions of mitochondrial dysfunction, seem to play a major role in the fate of pollen production. Nuclear genes, so called restorer-of-fertility are able to restore the male fertility. Fertility restoration can occur via pentatricopeptide repeat (PPR) proteins or via different mechanisms involving non-PPR proteins. (C) 2014 Elsevier B.V. and Mitochondria Research Society.
DOI:10.1186/s12284-014-0013-6URLPMID:26055995 [本文引用: 3]
The exploitation of male sterility systems has enabled the commercialization of heterosis in rice, with greatly increased yield and total production of this major staple food crop. Hybrid rice, which was adopted in the 1970s, now covers nearly 13.6 million hectares each year in China alone. Various types of cytoplasmic male sterility (CMS) and environment-conditioned genic male sterility (EGMS) systems have been applied in hybrid rice production. In this paper, recent advances in genetics, biochemistry, and molecular biology are reviewed with an emphasis on major male sterility systems in rice: five CMS systems, i.e., BT-, HL-, WA-, LD- and CW- CMS, and two EGMS systems, i.e., photoperiod- and temperature-sensitive genic male sterility (P/TGMS). The interaction of chimeric mitochondrial genes with nuclear genes causes CMS, which may be restored by restorer of fertility (Rf) genes. The PGMS, on the other hand, is conditioned by a non-coding RNA gene. A survey of the various CMS and EGMS lines used in hybrid rice production over the past three decades shows that the two-line system utilizing EGMS lines is playing a steadily larger role and TGMS lines predominate the current two-line system for hybrid rice production. The findings and experience gained during development and application of, and research on male sterility in rice not only advanced our understanding but also shed light on applications to other crops.
DOI:10.1093/aob/mcm121URLPMID:17704538 [本文引用: 3]
BACKGROUND: China has been successful in breeding hybrid rice strains, but is now facing challenges to develop new hybrids with high-yielding potential, better grain quality, and tolerance to biotic and abiotic stresses. This paper reviews the most significant advances in hybrid rice breeding in China, and presents a recent study on fine-mapping quantitative trait loci (QTLs) for yield traits. SCOPE: By exploiting new types of male sterility, hybrid rice production in China has become more diversified. The use of inter-subspecies crosses has made an additional contribution to broadening the genetic diversity of hybrid rice and played an important role in the breeding of super rice hybrids in China. With the development and application of indica-inclined and japonica-inclined parental lines, new rice hybrids with super high-yielding potential have been developed and are being grown on a large scale. DNA markers for subspecies differentiation have been identified and applied, and marker-assisted selection performed for the development of restorer lines carrying disease resistance genes. The genetic basis of heterosis in highly heterotic hybrids has been studied, but data from these studies are insufficient to draw sound conclusions. In a QTL study using stepwise residual heterozygous lines, two linked intervals harbouring QTLs for yield traits were resolved, one of which was delimited to a 125-kb region. CONCLUSIONS: Advances in rice genomic research have shed new light on the genetic study and germplasm utilization in rice. Molecular marker-assisted selection is a powerful tool to increase breeding efficiency, but much work remains to be done before this technique can be extended from major genes to QTLs.
[本文引用: 3]
[本文引用: 3]
URL [本文引用: 3]
The main problems about research and application of two-line hybrid rice were reviewed, including the confusing nomenclature and male sterile lines classification, the unclear characteristics of photoperiod and temperature responses and the unsuitable site selection for male sterile line and hybrid rice seed production. In order to efficiently and accurately use dual-purpose genic male sterile lines, four types, including PTGMS (photo-thermo-sensitive genic male sterile rice), TGMS (thermo-sensitive genic male sterile rice), reverse PTGMS and reverse TGMS, were proposed. A new idea for explaining the mechanism of sterility in dual-purpose hybrid rice was proposed. The transition from sterile to fertile was involved in the cooperative regulation of major-effect sterile genes and photoperiod and/or temperature sensitive ones. The minor-effect genes with accumulative effect on sterility were important factors that affected the critical temperature of sterility transfer. In order to make better use of dual-purpose lines, the characterization of responses to photoperiod and temperature of PTGMS should be made and the identification method for the characterization of photoperiod and temperature responses of PTGMS should also be put forward. The optimal ecological site for seed production could be determined according to the historical climate data and the requirements for the meteorological conditions during the different periods of seed production.
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1613792113URLPMID:27864513 [本文引用: 1]
The breeding and large-scale adoption of hybrid seeds is an important achievement in agriculture. Rice hybrid seed production uses cytoplasmic male sterile lines or photoperiod/thermo-sensitive genic male sterile lines (PTGMS) as female parent. Cytoplasmic male sterile lines are propagated via cross-pollination by corresponding maintainer lines, whereas PTGMS lines are propagated via self-pollination under environmental conditions restoring male fertility. Despite huge successes, both systems have their intrinsic drawbacks. Here, we constructed a rice male sterility system using a nuclear gene named Oryza sativa No Pollen 1 (OsNP1). OsNP1 encodes a putative glucose-methanol-choline oxidoreductase regulating tapetum degeneration and pollen exine formation; it is specifically expressed in the tapetum and miscrospores. The osnp1 mutant plant displays normal vegetative growth but complete male sterility insensitive to environmental conditions. OsNP1 was coupled with an alpha-amylase gene to devitalize transgenic pollen and the red fluorescence protein (DsRed) gene to mark transgenic seed and transformed into the osnp1 mutant. Self-pollination of the transgenic plant carrying a single hemizygous transgene produced nontransgenic male sterile and transgenic fertile seeds in 1:1 ratio that can be sorted out based on the red fluorescence coded by DsRed Cross-pollination of the fertile transgenic plants to the nontransgenic male sterile plants propagated the male sterile seeds of high purity. The male sterile line was crossed with approximately 1,200 individual rice germplasms available. Approximately 85% of the F1s outperformed their parents in per plant yield, and 10% out-yielded the best local cultivars, indicating that the technology is promising in hybrid rice breeding and production.
DOI:10.1186/s12870-019-1979-4URLPMID:31455245 [本文引用: 4]
BACKGROUND: Male sterility has tremendous scientific and economic importance in hybrid seed production. Identification and characterization of a stable male sterility gene will be highly beneficial for making hybrid seed production economically feasible. In soybean, eleven male-sterile, female-fertile mutant lines (ms1, ms2, ms3, ms4, ms5, ms6, ms7, ms8, ms9, msMOS, and msp) have been identified and mapped onto various soybean chromosomes, however the causal genes responsible for male sterility are not isolated. The objective of this study was to identify and functionally characterize the gene responsible for the male sterility in the ms4 mutant. RESULTS: The ms4 locus was fine mapped to a 216 kb region, which contains 23 protein-coding genes including Glyma.02G243200, an ortholog of Arabidopsis MALE MEIOCYTE DEATH 1 (MMD1), which is a Plant Homeodomain (PHD) protein involved in male fertility. Isolation and sequencing of Glyma.02G243200 from the ms4 mutant line showed a single base insertion in the 3rd exon causing a premature stop codon resulting in truncated protein production. Phylogenetic analysis showed presence of a homolog protein (MS4_homolog) encoded by the Glyma.14G212300 gene. Both proteins were clustered within legume-specific clade of the phylogenetic tree and were likely the result of segmental duplication during the paleoploidization events in soybean. The comparative expression analysis of Ms4 and Ms4_homologs across the soybean developmental and reproductive stages showed significantly higher expression of Ms4 in early flowering (flower bud differentiation) stage than its homolog. The functional complementation of Arabidopsis mmd1 mutant with the soybean Ms4 gene produced normal stamens, successful tetrad formation, fertile pollens and viable seeds, whereas the Ms4_homolog was not able to restore male fertility. CONCLUSIONS: Overall, this is the first report, where map based cloning approach was employed to isolate and characterize a gene responsible for the male-sterile phenotype in soybean. Characterization of male sterility genes may facilitate the establishment of a stable male sterility system, highly desired for the viability of hybrid seed production in soybean. Additionally, translational genomics and genome editing technologies can be utilized to generate new male-sterile lines in other plant species.
DOI:10.1002/ajb2.1982.69.issue-5URL [本文引用: 2]
DOI:10.1007/BF00023200URL [本文引用: 4]
DOI:10.1093/jhered/esq100URL [本文引用: 5]
In soybean [Glycine max (L.) Merr.], manual cross-pollination to produce large quantities of hybrid seed is difficult and time consuming. Identification of an environmentally stable male-sterility system could make hybrid seed production commercially valuable. In soybean, 2 environmentally sensitive male-sterile, female-fertile mutants (ms8 and msp) have been identified. Inheritance studies showed that sterility in both mutants is inherited as a single gene. The objectives of this study were to 1) confirm that msp and ms8 are independent genes; 2) identify the soybean chromosomes that contain the msp and the ms8 genes using bulked segregant analyses (BSAs); and 3) make a genetic linkage map of the regions containing these genes. Mapping populations consisting of 176 F(2) plants for ms8 and 134 F(2) plants for msp were generated. BSA revealed that Sat_389 and Satt172 are closely associated markers with ms8 and msp, respectively. Map location of Sat_389 suggested that the ms8 gene is located on chromosome 7; molecular linkage group (MLG) M. Map location of Satt172 indicated that the msp gene is located on chromosome 2 (MLG Dlb). Genetic linkage maps developed using F(2) populations revealed that ms8 is flanked by a telomere and Sat_389 and msp is flanked by Sat_069 and GMES4176. The region between the telomere and Sat_389 is physically 160 Kb. Soybean sequence information revealed that there are 13 genes present in that region. Protein BLASTP analyses revealed that homologs of 3 of the 13 genes are known to a play role in cell division, suggesting putative candidates for ms8.
DOI:10.1016/S2095-3119(18)62054-4URL [本文引用: 2]
DOI:10.1104/pp.3.2.223URLPMID:16652565 [本文引用: 2]
DOI:10.2135/cropsci1964.0011183X000400040027xURL [本文引用: 6]
DOI:10.1093/oxfordjournals.jhered.a109780URL [本文引用: 2]
DOI:10.1093/jhered/91.5.378URLPMID:10994704 [本文引用: 2]
A male-sterile, female-sterile soybean mutant (w4-m sterile) was identified among progeny of germinal revertants of a gene-tagging study. Our objectives were to determine the genetics (inheritance, allelism, and linkage) and the cytology (microsporogenesis and microgametogenesis) of the w4-m sterile. The mutant was inherited as a single recessive nuclear gene and was nonallelic to known male-sterile, female-sterile mutants st2 st2, st3 st3, st4 st4, st5 st5, and st6 st6 st7 st7. No linkage was detected between the w4-m sterile and the w4w4, y10 y10, y11 y11, y20 y20, fr1 fr1, and fr2 fr2 mutants. Homologous chromosome pairing was complete in fertile plants. Chromosome pairing, as observed in squash preparation, was almost completely absent in sterile plants. Developmentally microsporogenesis proceeded normally in both the fertile and the w4-m sterile through the early microspore stage. Then the tapetal cells of the w4-m sterile surrounding the young microspores developed different-size vacuoles. These tapetal cells became smaller in size and separated from each other. Some of the microspores of the w4-m sterile also became more vacuolate prematurely and sometimes they collapsed, usually by the late microspore stage. In the w4-m sterile the microspore walls remained thinner and structurally different from the microspore walls of fertile plants. No pollen was formed in the mutant plants, even though some of the male cells reached the pollen stage, although without normal filling. The w4-m sterile was designated st8st8 and assigned Soybean Genetic Type Collection number T352.
DOI:10.2135/cropsci1971.0011183X001100040032xURL [本文引用: 2]
DOI:10.1002/ajb2.1979.66.issue-3URL [本文引用: 1]
DOI:10.1002/ajb2.1985.72.issue-11URL [本文引用: 2]
DOI:10.1093/oxfordjournals.jhered.a109383URL [本文引用: 2]
DOI:10.1093/oxfordjournals.jhered.a109621URL [本文引用: 1]
DOI:10.3390/plants2030441URLPMID:27137386 [本文引用: 3]
In soybean, genic male sterility can be utilized as a tool to develop hybrid seed. Several male-sterile, female-fertile mutants have been identified in soybean. The male-sterile, female-fertile ms5 mutant was selected after fast neutron irradiation. Male-sterility due to ms5 was associated with the
[本文引用: 2]
DOI:10.2135/cropsci2000.40178xURL [本文引用: 4]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 2]
A spontaneous soybean male-sterile mutant N7241S was discovered and identified from a landrace Fuyangsilijia (N7241). The results of pollen germination experiments showed that the pollens of N7241S plants were completely aborted, while the female fertility of the N7241S sterile plants was normal according to parallel cross tests. The male-sterile plants performed relatively better in outcrossing ability than those of recessive monogenic mutants, such as ms1, ms2, etc. Therefore, N7241S is a male-sterile female-fertile mutant. Genetical analyses of different segregational populations between N7241S male-sterile plants and five normal parents showed that the male sterility of N7241S was controlled by a dominant gene. Strategies and methods of N7241S dominant male sterility to be used in recurrent selection as well as hybrid seed production were discussed.
URL [本文引用: 2]
A spontaneous soybean male-sterile mutant N7241S was discovered and identified from a landrace Fuyangsilijia (N7241). The results of pollen germination experiments showed that the pollens of N7241S plants were completely aborted, while the female fertility of the N7241S sterile plants was normal according to parallel cross tests. The male-sterile plants performed relatively better in outcrossing ability than those of recessive monogenic mutants, such as ms1, ms2, etc. Therefore, N7241S is a male-sterile female-fertile mutant. Genetical analyses of different segregational populations between N7241S male-sterile plants and five normal parents showed that the male sterility of N7241S was controlled by a dominant gene. Strategies and methods of N7241S dominant male sterility to be used in recurrent selection as well as hybrid seed production were discussed.
DOI:10.2135/cropsci2011.08.0410URL [本文引用: 1]
Cultivated soybean [Glycine max (L.) Merr.] is a major source of oil and protein in global food production. As demand for soy-based products increases, it is essential for breeders to find ways to increase yield beyond conventional methods. Alternative methods of cultivar development and improvement must be identified. Our objectives were to determine if the ms9 locus conferring male sterility in soybean was environmentally sensitive and, if it were, to determine which environments could be used in hybrid seed production. This was accomplished through characterization of pollen phenotypes and selfed seed set among progeny of soybean genotype T359H (Genetic Type Collection number T359H), a heterozygous maintainer line for the ms9 locus. T359H was evaluated in seven environmental regimens in growth chamber experiments. We used a split-plot design to test the effect of temperature regimens and male-sterile phenotypic classifications within chambers. As day temperature increased from 30 to 35 degrees C, selfed seed set on male-sterile, female-fertile plants decreased. Conversely, night temperature affected neither fertile nor male-sterile, female-fertile selfed seed set. This suggests that ms9 can be used in hybrid seed production in controlled environments where day temperature is 35 degrees C during the flowering. These temperature conditions and their effect on ms9 may provide an alternative method to produce hybrid seed in soybean cultivar development programs.
DOI:10.1139/gen-2015-0044URLPMID:26213292 [本文引用: 8]
In soybean, asynaptic and desynaptic mutants lead to abnormal meiosis and fertility reduction. Several male-sterile, female-sterile mutants have been identified and studied in soybean, however, some of these mutants have not been mapped to locations on soybean chromosomes. The objectives of this study were to molecularly map five male-sterile, female-sterile genes (st2, st4, st5, st6, and st7) in soybean and compare the map locations of these genes with already mapped sterility genes. Microsatellite markers were used in bulked segregant analyses to locate all five male-sterile, female-sterile genes to soybean chromosomes, and markers from the corresponding chromosomes were used on F2 populations to generate genetic linkage maps. The st2, st4, st5, st6, and st7 genes were located on molecular linkage group (MLG) B1 (chromosome 11), MLG D1a (chromosome 01), MLG F (chromosome 13), MLG B2 (chromosome 14), and D1b (chromosome 02), respectively. The st2, st4, st5, st6, and st7 genes were flanked to 10.3 ( approximately 399 kb), 6.3 ( approximately 164 kb), 3.9 ( approximately 11.8 Mb), 11.0 ( approximately 409 kb), and 5.3 cM ( approximately 224 kb), and the flanked regions contained 57, 17, 362, 52, and 17 predicted genes, respectively. Future characterization of candidate genes should facilitate identification of the male- and female-fertility genes, which may provide vital insights on structure and function of genes involved in the reproductive pathway in soybean.
DOI:10.1093/jxb/eru261URLPMID:24987011 [本文引用: 1]
The plant specific RWP-RK family of transcription factors, initially identified in legumes and Chlamydomonas, are found in all vascular plants, green algae, and slime molds. These proteins possess a characteristic RWP-RK motif, which mediates DNA binding. Based on phylogenetic and domain analyses, we classified the RWP-RK proteins of six different species in two subfamilies: the NIN-like proteins (NLPs), which carry an additional PB1 domain at their C-terminus, and the RWP-RK domain proteins (RKDs), which are divided into three subgroups. Although, the functional analysis of this family is still in its infancy, several RWP-RK proteins have a key role in regulating responses to nitrogen availability. The nodulation-specific NIN proteins are involved in nodule organogenesis and rhizobial infection under nitrogen starvation conditions. Arabidopsis NLP7 in particular is a major player in the primary nitrate response. Several RKDs act as transcription factors involved in egg cell specification and differentiation or gametogenesis in algae, the latter modulated by nitrogen availability. Further studies are required to extend the general picture of the functional role of these exciting transcription factors.
DOI:10.1093/jhered/esg085URLPMID:14557397 [本文引用: 2]
Soybean male-sterile, female-sterile mutant genes have been identified by genetic and cytological studies. The St8 gene has been identified as an asynaptic mutation resulting in male and female sterility. This mutant gene was derived from a gene-tagging study using the soybean w4-mutable line. In this report we identified the genetic map position of st8 via restriction fragment length polymorphism (RFLP) and simple sequence repeat (SSR) markers. The St8 gene mutation was located between RFLP marker E107 and SSR markers Satt132, Sct_065, and Satt414 on molecular linkage group J and linked to each by 7.8 cM and 3.4 cM, respectively.
DOI:10.2135/cropsci2006.03.0143URL [本文引用: 4]
DOI:10.1139/gen-2014-0018URL [本文引用: 5]
In soybean, an environmentally stable male sterility system is vital for making hybrid seed production commercially viable. Eleven male-sterile, female-fertile mutants (ms1, ms2, ms3, ms4, ms5, ms6, ms7, ms8, ms9, msMOS, and msp) have been identified in soybean. Of these, eight (ms2, ms3, ms5, ms7, ms8, ms9, msMOS, and msp) have been mapped to soybean chromosomes. The objectives of this study were to (i) locate the ms1, ms4, and ms6 genes to soybean chromosomes; (ii) generate genetic linkage maps of the regions containing these genes; and (iii) develop a comprehensive map of all known male-sterile, female-fertile genes in soybean. The bulked segregant analysis technique was used to locate genes to soybean chromosomes. Microsatellite markers from the corresponding chromosomes were used on F-2 populations to generate genetic linkage maps. The ms1 and ms6 genes were located on chromosome 13 (molecular linkage group F) and ms4 was present on chromosome 2 (molecular linkage group D1b). Molecular analyses revealed markers Satt516, BARCSOYSSR_02_1539, and AW186493 were located closest to ms1, ms4, and ms6, respectively. The ms1 and ms6 genes, although present on the same chromosome, were independently assorting with a genetic distance of 73.7 cM. Using information from this study and compiled information from previously published male sterility genes in soybean, a comprehensive genetic linkage map was generated. Eleven male sterility genes were present on seven soybean chromosomes. Four genes were present in two regions on chromosome 2 (molecular linkage group D1b) and two genes were present on chromosome 13 (molecular linkage group F).
DOI:10.1007/s00497-019-00377-6URLPMID:31620875 [本文引用: 3]
Nuclear male sterility (NMS) is a potential characteristic in crop recurrent selection and hybrid breeding. Mapping of nuclear male-sterile genes is key to utilizing NMS. Previously, we discovered a spontaneous soybean (Glycine max [L.] Merr.) male-sterile female-fertile mutant NJS-13H, which was conferred by a single recessive gene, designated msNJ. In this study, the msNJ was mapped to Chromosome 10 (LG O), and narrowed down between two SSR (simple sequence repeats) markers, BARCSOYSSR_10_794 and BARCSOYSSR_10_819 using three heterozygote-derived segregating populations, i.e., (NJS-13H x NN1138-2)F2, (NJS-13H x N2899)F2 and (NJS-13H)SPAG (segregating populations in advanced generations). This region spans approximately 1.32 Mb, where 27 genes were annotated according to the soybean reference genome sequence (Wm82.a2.v1). Among them, four genes were recognized as candidate genes for msNJ. Comparing to the physical locations of all the known male-sterile loci, msNJ is demonstrated to be a new male-sterile locus. This result may help the utilization and cloning of the gene.
DOI:10.1093/jhered/esp054URLPMID:19617521 [本文引用: 3]
Soybean [Glycine max (L.) Merr.] is self-pollinated. To produce large quantities of hybrid seed, insect-mediated cross-pollination is necessary. An efficient nuclear male-sterile system for hybrid seed production would benefit from molecular and/or phenotypic markers linked to male fertility/sterility loci to facilitate early identification of phenotypes. Nuclear male-sterile, female-fertile ms3 mutant is a single recessive gene and displays high outcrossed seed set with pollinators. Our objective was to map the ms3 locus. A segregating population of 150 F(2) plants from Minsoy (PI 27890) x T284H, Ms3ms3 (A00-68), was screened with 231 simple sequence repeat markers. The ms3 locus mapped to molecular linkage group (MLG) D1b (Gm02) and is flanked by markers Satt157 and Satt542, with a distance of 3.7 and 12.3 cM, respectively. Female-partial sterile-1 (Fsp1) and the Midwest Oilseed male-sterile (msMOS) mutants previously were located on MLG D1b. msMOS and Fsp1 are independent genes located very close to each other. All 3 genes are located in close proximity of Satt157. We believe that this is the first report of clustering of fertility-related genes in plants. Characterization of these closely linked genes may help in understanding the evolutionary relationship among them.
DOI:10.1093/oxfordjournals.jhered.a110338URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3389/fpls.2019.00001URL [本文引用: 1]
DOI:10.1016/j.molp.2019.01.014URLPMID:30690174 [本文引用: 4]
As one of the most important crops, maize not only has been a source of the food, feed, and industrial feedstock for biofuel and bioproducts, but also became a model plant system for addressing fundamental questions in genetics. Male sterility is a very useful trait for hybrid vigor utilization and hybrid seed production. The identification and characterization of genic male-sterility (GMS) genes in maize and other plants have deepened our understanding of the molecular mechanisms controlling anther and pollen development, and enabled the development and efficient use of many biotechnology-based male-sterility (BMS) systems for crop hybrid breeding. In this review, we summarize main advances on the identification and characterization of GMS genes in maize, and construct a putative regulatory network controlling maize anther and pollen development by comparative genomic analysis of GMS genes in maize, Arabidopsis, and rice. Furthermore, we discuss and appraise the features of more than a dozen BMS systems for propagating male-sterile lines and producing hybrid seeds in maize and other plants. Finally, we provide our perspectives on the studies of GMS genes and the development of novel BMS systems in maize and other plants. The continuous exploration of GMS genes and BMS systems will enhance our understanding of molecular regulatory networks controlling male fertility and greatly facilitate hybrid vigor utilization in breeding and field production of maize and other crops.
DOI:10.1105/tpc.012427URLPMID:12897259 [本文引用: 1]
Membrane-bound glycerol-3-phosphate acyltransferase (GPAT; EC 2.3.1.15) mediates the initial step of glycerolipid biosynthesis in the extraplastidic compartments of plant cells. Here, we report the molecular characterization of a novel GPAT gene family from Arabidopsis, designated AtGPAT. The corresponding polypeptides possess transmembrane domains and GPAT activity when expressed heterologously in a yeast lipid mutant. The functional significance of one isoform, AtGPAT1, is the focus of the present study. Disruption of the AtGPAT1 gene causes a massive pollen development arrest, and subsequent introduction of the gene into the mutant plant rescues the phenotype, illustrating a pivotal role for AtGPAT1 in pollen development. Microscopic examinations revealed that the gene lesion results in a perturbed degeneration of the tapetum, which is associated with altered endoplasmic reticulum profiles and reduced secretion. In addition to the sporophytic effect, AtGPAT1 also exerts a gametophytic effect on pollen performance, as the competitive ability of a pollen grain to pollinate is dependent on the presence of an AtGPAT1 gene. Deficiency in AtGPAT1 correlates with several fatty acid composition changes in flower tissues and seeds. Unexpectedly, however, a loss of AtGPAT1 causes no significant change in seed oil content.
DOI:10.1105/tpc.110.080036URLPMID:21193572 [本文引用: 1]
The precise structure of the sporopollenin polymer that is the major constituent of exine, the outer pollen wall, remains poorly understood. Recently, characterization of Arabidopsis thaliana genes and corresponding enzymes involved in exine formation has demonstrated the role of fatty acid derivatives as precursors of sporopollenin building units. Fatty acyl-CoA esters synthesized by ACYL-COA SYNTHETASE5 (ACOS5) are condensed with malonyl-CoA by POLYKETIDE SYNTHASE A (PKSA) and PKSB to yield alpha-pyrone polyketides required for exine formation. Here, we show that two closely related genes encoding oxidoreductases are specifically and transiently expressed in tapetal cells during microspore development in Arabidopsis anthers. Mutants compromised in expression of the reductases displayed a range of pollen exine layer defects, depending on the mutant allele. Phylogenetic studies indicated that the two reductases belong to a large reductase/dehydrogenase gene family and cluster in two distinct clades with putative orthologs from several angiosperm lineages and the moss Physcomitrella patens. Recombinant proteins produced in bacteria reduced the carbonyl function of tetraketide alpha-pyrone compounds synthesized by PKSA/B, and the proteins were therefore named TETRAKETIDE alpha-PYRONE REDUCTASE1 (TKPR1) and TKPR2 (previously called DRL1 and CCRL6, respectively). TKPR activities, together with those of ACOS5 and PKSA/B, identify a conserved biosynthetic pathway leading to hydroxylated alpha-pyrone compounds that were previously unknown to be sporopollenin precursors.
DOI:10.1105/tpc.106.046391URLPMID:17329564 [本文引用: 1]
The Arabidopsis thaliana MYB26/MALE STERILE35 (MS35) gene is critical for the development of secondary thickening in the anther endothecium and subsequent dehiscence. MYB26 is localized to the nucleus and regulates endothecial development and secondary thickening in a cell-specific manner in the anther. MYB26 expression is seen in anthers and also in the style and nectaries, although there is no effect on female fertility in the ms35 mutant. MYB26 expression in anthers occurs early during endothecial development, with maximal expression during pollen mitosis I and bicellular stages, indicating a regulatory role in specifying early endothecial cell development. Overexpression of MYB26 results in ectopic secondary thickening in both Arabidopsis and tobacco (Nicotiana tabacum) plants, predominantly within the epidermal tissues. MYB26 regulates a number of genes linked to secondary thickening, including IRREGULAR XYLEM1 (IRX1), IRX3, IRX8, and IRX12. Changes in expression were also detected in two NAC domain genes, NAC SECONDARY WALL-PROMOTING FACTOR1 (NST1) and NST2, which have been linked to secondary thickening in the anther endothecium. These data indicate that MYB26 regulates NST1 and NST2 expression and in turn controls the process of secondary thickening. Therefore, MYB26 appears to function in a regulatory role involved in determining endothecial cell development within the anther and acts upstream of the lignin biosynthesis pathway.
DOI:10.1038/ncomms4855URL [本文引用: 1]
DOI:10.1104/pp.109.144469URLPMID:19700560 [本文引用: 1]
Sporopollenin is the major component of the outer pollen wall (exine). Fatty acid derivatives and phenolics are thought to be its monomeric building blocks, but the precise structure, biosynthetic route, and genetics of sporopollenin are poorly understood. Based on a phenotypic mutant screen in Arabidopsis (Arabidopsis thaliana), we identified a cytochrome P450, designated CYP704B1, as being essential for exine development. CYP704B1 is expressed in the developing anthers. Mutations in CYP704B1 result in impaired pollen walls that lack a normal exine layer and exhibit a characteristic striped surface, termed zebra phenotype. Heterologous expression of CYP704B1 in yeast cells demonstrated that it catalyzes omega-hydroxylation of long-chain fatty acids, implicating these molecules in sporopollenin synthesis. Recently, an anther-specific cytochrome P450, denoted CYP703A2, that catalyzes in-chain hydroxylation of lauric acid was also shown to be involved in sporopollenin synthesis. This shows that different classes of hydroxylated fatty acids serve as essential compounds for sporopollenin formation. The genetic relationships between CYP704B1, CYP703A2, and another exine gene, MALE STERILITY2, which encodes a fatty acyl reductase, were explored. Mutations in all three genes resulted in pollen with remarkably similar zebra phenotypes, distinct from those of other known exine mutants. The double and triple mutant combinations did not result in the appearance of novel phenotypes or enhancement of single mutant phenotypes. This implies that each of the three genes is required to provide an indispensable subset of fatty acid-derived components within the sporopollenin biosynthesis framework.
DOI:10.1105/tpc.106.045948URLPMID:17496121 [本文引用: 1]
CYP703 is a cytochrome P450 family specific to land plants. Typically, each plant species contains a single CYP703. Arabidopsis thaliana CYP703A2 is expressed in the anthers of developing flowers. Expression is initiated at the tetrad stage and restricted to microspores and to the tapetum cell layer. Arabidopsis CYP703A2 knockout lines showed impaired pollen development and a partial male-sterile phenotype. Scanning electron and transmission electron microscopy of pollen from the knockout plants showed impaired pollen wall development with absence of exine. The fluorescent layer around the pollen grains ascribed to the presence of phenylpropanoid units in sporopollenin was absent in the CYP703A2 knockout lines. Heterologous expression of CYP703A2 in yeast cells demonstrated that CYP703 catalyzes the conversion of medium-chain saturated fatty acids to the corresponding monohydroxylated fatty acids, with a preferential hydroxylation of lauric acid at the C-7 position. Incubation of recombinant CYP703 with methanol extracts from developing flowers confirmed that lauric acid and in-chain hydroxy lauric acids are the in planta substrate and product, respectively. These data demonstrate that in-chain hydroxy lauric acids are essential building blocks in sporopollenin synthesis and enable the formation of ester and ether linkages with phenylpropanoid units. This study identifies CYP703 as a P450 family specifically involved in pollen development.
DOI:10.1105/tpc.110.080028URLPMID:21193570 [本文引用: 1]
Plant type III polyketide synthases (PKSs) catalyze the condensation of malonyl-CoA units with various CoA ester starter molecules to generate a diverse array of natural products. The fatty acyl-CoA esters synthesized by Arabidopsis thaliana ACYL-COA SYNTHETASE5 (ACOS5) are key intermediates in the biosynthesis of sporopollenin, the major constituent of exine in the outer pollen wall. By coexpression analysis, we identified two Arabidopsis PKS genes, POLYKETIDE SYNTHASE A (PKSA) and PKSB (also known as LAP6 and LAP5, respectively) that are tightly coexpressed with ACOS5. Recombinant PKSA and PKSB proteins generated tri-and tetraketide alpha-pyrone compounds in vitro from a broad range of potential ACOS5-generated fatty acyl-CoA starter substrates by condensation with malonyl-CoA. Furthermore, substrate preference profile and kinetic analyses strongly suggested that in planta substrates for both enzymes are midchain- and omega-hydroxylated fatty acyl-CoAs (e.g., 12-hydroxyoctadecanoyl-CoA and 16-hydroxyhexadecanoyl-CoA), which are the products of sequential actions of anther-specific fatty acid hydroxylases and acyl-CoA synthetase. PKSA and PKSB are specifically and transiently expressed in tapetal cells during microspore development in Arabidopsis anthers. Mutants compromised in expression of the PKS genes displayed pollen exine layer defects, and a double pksa pksb mutant was completely male sterile, with no apparent exine. These results show that hydroxylated alpha-pyrone polyketide compounds generated by the sequential action of ACOS5 and PKSA/B are potential and previously unknown sporopollenin precursors.
URLPMID:11743117 [本文引用: 1]
To identify factors that are required for proper pollen wall formation, we have characterized the T-DNA-tagged, dex1 mutation of Arabidopsis, which results in defective pollen wall pattern formation. This study reports the isolation and molecular characterization of DEX1 and morphological and ultrastructural analyses of dex1 plants. DEX1 encodes a novel plant protein that is predicted to be membrane associated and contains several potential calcium-binding domains. Pollen wall development in dex1 plants parallels that of wild-type plants until the early tetrad stage. In dex1 plants, primexine deposition is delayed and significantly reduced. The normal rippling of the plasma membrane and production of spacers observed in wild-type plants is also absent in the mutant. Sporopollenin is produced and randomly deposited on the plasma membrane in dex1 plants. However, it does not appear to be anchored to the microspore and forms large aggregates on the developing microspore and the locule walls. Based on the structure of DEX1 and the phenotype of dex1 plants, several potential roles for the protein are proposed.
DOI:10.1104/pp.108.118026URLPMID:18434608 [本文引用: 1]
During microsporogenesis, the microsporocyte (or microspore) plasma membrane plays multiple roles in pollen wall development, including callose secretion, primexine deposition, and exine pattern determination. However, plasma membrane proteins that participate in these processes are still not well known. Here, we report that a new gene, RUPTURED POLLEN GRAIN1 (RPG1), encodes a plasma membrane protein and is required for exine pattern formation of microspores in Arabidopsis (Arabidopsis thaliana). The rpg1 mutant exhibits severely reduced male fertility with an otherwise normal phenotype, which is largely due to the postmeiotic abortion of microspores. Scanning electron microscopy examination showed that exine pattern formation in the mutant is impaired, as sporopollenin is randomly deposited on the pollen surface. Transmission electron microscopy examination further revealed that the primexine formation of mutant microspores is aberrant at the tetrad stage, which leads to defective sporopollenin deposition on microspores and the locule wall. In addition, microspore rupture and cytoplasmic leakage were evident in the rpg1 mutant, which indicates impaired cell integrity of the mutant microspores. RPG1 encodes an MtN3/saliva family protein that is integral to the plasma membrane. In situ hybridization analysis revealed that RPG1 is strongly expressed in microsporocyte (or microspores) and tapetum during male meiosis. The possible role of RPG1 in microsporogenesis is discussed.
DOI:10.1111/tpj.12767URLPMID:25611322 [本文引用: 1]
During male gametophyte development in Arabidopsis thaliana, the microspores undergo an asymmetric division to produce a vegetative cell and a generative cell, which undergoes a second division to give rise to two sperm cells. SIDECAR POLLEN/LATERAL ORGAN BOUNDARIES DOMAIN (LBD) 27 plays a key role in the asymmetric division of microspores. Here we provide molecular genetic evidence that a combinatorial role of LBD10 with LBD27 is crucial for male gametophyte development in Arabidopsis. Expression analysis, genetic transmission and pollen viability assays, and pollen development analysis demonstrated that LBD10 plays a role in the male gametophyte function primarily at germ cell mitosis. In the mature pollen of lbd10 and lbd10 expressing a dominant negative version of LBD10, LBD10:SRDX, aberrant microspores such as bicellular and smaller tricellular pollen appeared at a ratio of 10-15% with a correspondingly decreased ratio of normal tricellular pollen, whereas in lbd27 mutants, 70% of the pollen was aborted. All pollen in the lbd10 lbd27 double mutants was aborted and severely shrivelled compared with that of the single mutants, indicating that LBD10 and LBD27 are essential for pollen development. Gene expression and subcellular localization analyses of LBD10:GFP and LBD27:RFP during pollen development indicated that posttranscriptional and/or posttranslational controls are involved in differential accumulation and subcellular localization of LBD10 and LBD27 during pollen development, which may contribute in part to combinatorial and distinct roles of LBD10 with LBD27 in microspore development. In addition, we showed that LBD10 and LBD27 interact to form a heterodimer for nuclear localization.
DOI:10.1105/tpc.109.071803URLPMID:20118226 [本文引用: 3]
The Arabidopsis thaliana ABORTED MICROSPORES (AMS) gene encodes a basic helix-loop-helix (bHLH) transcription factor that is required for tapetal cell development and postmeiotic microspore formation. However, the regulatory role of AMS in anther and pollen development has not been fully defined. Here, we show by microarray analysis that the expression of 549 anther-expressed genes was altered in ams buds and that these genes are associated with tapetal function and pollen wall formation. We demonstrate that AMS has the ability to bind in vitro to DNA containing a 6-bp consensus motif, CANNTG. Moreover, 13 genes involved in transportation of lipids, oligopeptides, and ions, fatty acid synthesis and metabolism, flavonol accumulation, substrate oxidation, methyl-modification, and pectin dynamics were identified as direct targets of AMS by chromatin immunoprecipitation. The functional importance of the AMS regulatory pathway was further demonstrated by analysis of an insertional mutant of one of these downstream AMS targets, an ABC transporter, White-Brown Complex homolog, which fails to undergo pollen development and is male sterile. Yeast two-hybrid screens and pull-down assays revealed that AMS has the ability to interact with two bHLH proteins (AtbHLH089 and AtbHLH091) and the ATA20 protein. These results provide insight into the regulatory role of the AMS network during anther development.
DOI:10.1007/s11434-015-0810-3URL [本文引用: 1]
DOI:10.1104/pp.15.00262URLPMID:26392263 [本文引用: 1]
Male reproduction in higher plants requires the support of various metabolites, including lipid molecules produced in the innermost anther wall layer (the tapetum), but how the molecules are allocated among different anther tissues remains largely unknown. Previously, rice (Oryza sativa) ATP binding cassette G15 (ABCG15) and its Arabidopsis (Arabidopsis thaliana) ortholog were shown to be required for pollen exine formation. Here, we report the significant role of OsABCG26 in regulating the development of anther cuticle and pollen exine together with OsABCG15 in rice. Cytological and chemical analyses indicate that osabcg26 shows reduced transport of lipidic molecules from tapetal cells for anther cuticle development. Supportively, the localization of OsABCG26 is on the plasma membrane of the anther wall layers. By contrast, OsABCG15 is polarly localized in tapetal plasma membrane facing anther locules. osabcg26 osabcg15 double mutant displays an almost complete absence of anther cuticle and pollen exine, similar to that of osabcg15 single mutant. Taken together, we propose that OsABCG26 and OsABCG15 collaboratively regulate rice male reproduction: OsABCG26 is mainly responsible for the transport of lipidic molecules from tapetal cells to anther wall layers, whereas OsABCG15 mainly is responsible for the export of lipidic molecules from the tapetal cells to anther locules for pollen exine development.
DOI:10.1104/pp.111.175760URL [本文引用: 1]
In higher plants, timely degradation of tapetal cells, the innermost sporophytic cells of the anther wall layer, is a prerequisite for the development of viable pollen grains. However, relatively little is known about the mechanism underlying programmed tapetal cell development and degradation. Here, we report a key regulator in monocot rice (Oryza sativa), PERSISTANT TAPETAL CELL1 (PTC1), which controls programmed tapetal development and functional pollen formation. The evolutionary significance of PTC1 was revealed by partial genetic complementation of the homologous mutation MALE STERILITY1 (MS1) in the dicot Arabidopsis (Arabidopsis thaliana). PTC1 encodes a PHD-finger (for plant homeodomain) protein, which is expressed specifically in tapetal cells and microspores during anther development in stages 8 and 9, when the wild-type tapetal cells initiate a typical apoptosis-like cell death. Even though ptc1 mutants show phenotypic similarity to ms1 in a lack of tapetal DNA fragmentation, delayed tapetal degeneration, as well as abnormal pollen wall formation and aborted microspore development, the ptc1 mutant displays a previously unreported phenotype of uncontrolled tapetal proliferation and subsequent commencement of necrosis-like tapetal death. Microarray analysis indicated that 2,417 tapetum-and microspore- expressed genes, which are principally associated with tapetal development, degeneration, and pollen wall formation, had changed expression in ptc1 anthers. Moreover, the regulatory role of PTC1 in anther development was revealed by comparison with MS1 and other rice anther developmental regulators. These findings suggest a diversified and conserved switch of PTC1/MS1 in regulating programmed male reproductive development in both dicots and monocots, which provides new insights in plant anther development.
DOI:10.1111/jipb.12574URL [本文引用: 1]
Lipid and phenolic metabolism are important for pollen exine formation. In Arabidopsis, polyketide synthases (PKSs) are essential for both sporopollenin biosynthesis and exine formation. Here, we characterized the role of a polyketide synthase (OsPKS2) in male reproduction of rice (Oryza sativa). Recombinant OsPKS2 catalyzed the condensation of fatty acyl-CoA with malonyl-CoA to generate triketide and tetraketide α-pyrones, the main components of pollen exine. Indeed, the ospks2 mutant had defective exine patterning and was male sterile. However, the mutant showed no significant reduction in sporopollenin accumulation. Compared with the WT (wild type), ospks2 displayed unconfined and amorphous tectum and nexine layers in the exine, and less organized Ubisch bodies. Like the pksb/lap5 mutant of the Arabidopsis ortholog, ospks2 showed broad alterations in the profiles of anther-related phenolic compounds. However, unlike pksb/lap5, in which most detected phenolics were substantially decreased, ospks2 accumulated higher levels of phenolics. Based on these results and our observation that OsPKS2 is unable to fully restore the exine defects in the pksb/lap5, we propose that PKS proteins have functionally diversified during evolution. Collectively, our results suggest that PKSs represent a conserved and diversified biochemical pathway for anther and pollen development in higher plants.
DOI:10.1186/s12870-016-0943-9URLPMID:27871243 [本文引用: 1]
BACKGROUND: Sporopollenin is a major component of the pollen exine pattern. In Arabidopsis, acyl-CoA synthetase5 (ACOS5) is involved in sporopollenin precursor biosynthesis. In this study, we identified its orthologue, OsACOS12, in rice (Oryza sativa) and compared the functional conservation of ACOS in rice to Arabidopsis. RESULTS: Sequence analysis showed that OsACOS12 shares 63.9 % amino acid sequence identity with ACOS5. The osacos12 mutation caused by a pre-mature stop codon in LOC_Os04g24530 exhibits defective sexine resulting in a male sterile phenotype in rice. In situ hybridization shows that OsACOS12 is expressed in tapetal cells and microspores at the transcript level. The localization of OsACOS12-GFP demonstrated that OsACOS12 protein is accumulated in tapetal cells and anther locules. OsACOS12 driven by the ACOS5 promoter could partially restore the male fertility of the acos5 mutant in Arabidopsis. CONCLUSIONS: OsACOS12 is an orthologue of ACOS5 that is essential for sporopollenin synthesis in rice. ACOS5 and OsACOS12 are conserved for pollen wall formation in monocot and dicot species.
DOI:10.1016/j.devcel.2012.04.011URLPMID:22698279 [本文引用: 1]
In flowering plants, formation of the haploid male gametophytes in anthers requires the interaction between reproductive cells and the neighboring somatic cells, yet the underlying mechanism remains poorly understood. Here, we reveal the crucial role of a fasciclin glycoprotein, MICROSPORE AND TAPETUM REGULATOR1 (MTR1), in controlling the development of sporophytic and reproductive cells in rice (Oryza sativa). MTR1 is specifically expressed in the male reproductive cells, yet its mutant exhibits defects in both tapetum and microspore development, causing complete male sterility. We also demonstrate that the fasciclin domains, N-glycolation, and N-terminal signal peptide-mediated plasma membrane localization of MTR1 are required for normal anther development and pollen fertility. Our findings show that rice male reproductive cells secrete the MTR1 protein to control the development of reproductive cells and their adjacent somatic cells, thus providing novel insights into the mechanism of plant male reproductive development.
DOI:10.1105/tpc.114.123745URL [本文引用: 2]
In male reproductive development in plants, meristemoid precursor cells possessing transient, stem cell-like features undergo cell divisions and differentiation to produce the anther, the male reproductive organ. The anther contains centrally positioned microsporocytes surrounded by four distinct layers of wall: the epidermis, endothecium, middle layer, and tapetum. Here, we report that the rice (Oryza sativa) basic helix-loop-helix ( bHLH) protein TDR INTERACTING PROTEIN2 (TIP2) functions as a crucial switch in the meristemoid transition and differentiation during early anther development. The tip2 mutants display undifferentiated inner three anther wall layers and abort tapetal programmed cell death, causing complete male sterility. TIP2 has two paralogs in rice, TDR and EAT1, which are key regulators of tapetal programmed cell death. We revealed that TIP2 acts upstream of TDR and EAT1 and directly regulates the expression of TDR and EAT1. In addition, TIP2 can interact with TDR, indicating a role of TIP2 in later anther development. Our findings suggest that the bHLH proteins TIP2, TDR, and EAT1 play a central role in regulating differentiation, morphogenesis, and degradation of anther somatic cell layers, highlighting the role of paralogous bHLH proteins in regulating distinct steps of plant cell-type determination.
DOI:10.1038/s41598-017-07064-4URLPMID:28761138 [本文引用: 1]
Strictosidine synthase (STR) plays an important role in the biosynthesis of terpenoid indole alkaloids (TIAs) and is expressed in a range of active meristematic tissues of higher plants. STR proteins are involved in different physiological and biochemical pathways. However, the function of STR proteins in rice development remains poorly understood. In this study, we identified 21 possible STR-like (OsSTRL) family members in rice genome and found that only one gene, OsSTRL2, exhibited a pre-emergency specific florescence expression pattern. Tissue-specific expression profile analysis, beta-glucuronidase histochemical (GUS) staining and RNA in situ hybridization confirmed that OsSTRL2 was highly expressed in tapetal cells and microspores. Comparative protein sequence analysis indicated that OsSTRL2 lacked the key catalytic residue found in a typical STR (STR1), although it possessed conserved beta-propellers and alpha-helices formed the basic structure of STR1. OsSTRL2 knockout mutant resulted to male sterility because of the defects in anther development and pollen wall formation. Subcellular localization of OsSTRL2-YFP revealed that the OsSTRL2 protein was primarily localized in the endoplasmic reticulum (ER). Therefore, OsSTRL2 is an atypical strictosidine synthase that plays crucial roles in regulating anther development and pollen wall formation in rice.
DOI:10.1104/pp.112.210948URL [本文引用: 1]
The pollen wall consists of an exine and an intine. The mechanism underlying its formation is not well understood. Glycosyltransferases catalyze the modification of biological molecules by attaching a single or multiple sugars and play key roles in a wide range of biological processes. We examined the role of GLYCOSYLTRANSFERASE1 (OsGT1) in pollen wall development in rice (Oryza sativa). This gene is highly expressed in mature pollen, and plants containing alleles caused by transfer DNA insertion do not produce homozygous progeny. Reciprocal crosses between OsGT1/osgt1 and the wild type indicated that the mutation leads to a male gametophyte defect. Microscopic analyses revealed that osgt1 pollen developed normally to the pollen mitosis stage but failed to produce mature grains. In osgt1 pollen, intine structure was disrupted. In addition, starch and protein levels were much lower in the mutant grains. Recombinant OsGT1 transferred glucose from UDP-glucose to the third and seventh positions of quercetin, a universal substrate of glycosyltransferases. Consistent with the role of OsGT1, an OsGT1-green fluorescent protein fusion protein was localized to the Golgi apparatus. Taken together, our results suggest that OsGT1 is a Golgi-localized glycosyltransferase essential for intine construction and pollen maturation, providing new insight into male reproductive development.
DOI:10.1093/pcp/pcu193URLPMID:25520407 [本文引用: 1]
Callose plays an important role in pollen development in flowering plants. In rice, 10 genes encoding putative callose synthases have been identified; however, none of them has been functionally characterized. In this study, a rice Glucan Synthase-Like 5 (GSL5) knock-out mutant was isolated that exhibited a severe reduction in fertility. Pollen viability tests indicated that the pollen of the mutant was abnormal while the embryo sac was normal. Further, GSL5-RNA interference transgenic plants phenocopied the gsl5 mutant. The RNA expression of GSL5 was found to be knocked out in the gsl5 mutant and knocked down in GSL5-RNA interference transgenic plants by real-time reverse transcripion-PCR (RT-PCR) analysis. The male sterility of the mutant was due to abnormal microspore development; an analysis of paraffin sections of the mutant anthers at various developmental stages revealed that abnormal microspore development began in late meiosis. Both the knock-out and knock-down of GSL5 caused a lack of callose in the primary cell wall of meiocytes and in the cell plate of tetrads. As a result, the callose wall of the microspores was defective. This was demonstrated by aniline blue staining and an immunogold labeling assay; the microspores could not maintain their shape, leading to premature swelling and even collapsed microspores. These data suggest that the callose synthase encoded by GSL5 plays a vital role in microspore development during late meiosis and is essential for male fertility in rice.
DOI:10.1016/j.molp.2020.05.009URLPMID:32434071 [本文引用: 1]
Fatty acids and their derivatives are essential building blocks for anther cuticle and pollen wall formation. Disruption of lipid metabolism during anther and pollen development often leads to genic male sterility (GMS). To date, many lipid metabolism-related GMS genes that are involved in the formation of anther cuticle, pollen wall, and subcellular organelle membranes in anther wall layers have been identified and characterized. In this review, we summarize recent progress on characterizing lipid metabolism-related genes and their roles in male fertility and other aspects of reproductive development in plants. On the basis of cloned GMS genes controlling biosynthesis and transport of anther cutin, wax, sporopollenin, and tryphine in Arabidopsis, rice, and maize as well as other plant species, updated lipid metabolic networks underlying anther cuticle development and pollen wall formation were proposed. Through bioinformatics analysis of anther RNA-sequencing datasets from three maize inbred lines (Oh43, W23, and B73), a total of 125 novel lipid metabolism-related genes putatively involved in male fertility in maize were deduced. More, we discuss the pathways regulating lipid metabolism-related GMS genes at the transcriptional and post-transcriptional levels. Finally, we highlight recent findings on lipid metabolism-related genes and their roles in other aspects of plant reproductive development. A comprehensive understanding of lipid metabolism, genes involved, and their roles in plant reproductive development will facilitate the application of lipid metabolism-related genes in gene editing, haploid and callus induction, molecular breeding and hybrid seed production in crops.
DOI:10.1105/tpc.107.054536URLPMID:18032630 [本文引用: 2]
The Arabidopsis thaliana MALE STERILITY1 (MS1) gene encodes a nuclear protein with Leu zipper-like and PHD-finger motifs and is important for postmeiotic pollen development. Here, we examined MS1 function using both cell biological and molecular biological approaches. We introduced a fusion construct of MS1 and a transcriptional repression domain (MS1-SRDX) into wild-type Arabidopsis, and the transgenic plants showed a semisterile phenotype similar to that of ms1. Since the repression domain can convert various kinds of transcriptional activators to dominant repressors, this suggested that MS1 functioned as a transcriptional activator. The Leu zipper-like region and the PHD motif were required for the MS1 function. Phenotypic analysis of the ms1 mutant and the MS1-SRDX transgenic Arabidopsis indicated that MS1 was involved in formation of pollen exine and pollen cytosolic components as well as tapetum development. Next, we searched for MS1 downstream genes by analyzing publicly available microarray data and identified 95 genes affected by MS1. Using a transgenic ms1 plant showing dexamethasone-inducible recovery of fertility, we further examined whether these genes were immediately downstream of MS1. From these results, we discuss a role of MS1 in pollen and tapetum development and the conservation of MS1 function in flowering plants.
DOI:10.1111/pbi.12786URLPMID:28678349 [本文引用: 1]
Although hundreds of genetic male sterility (GMS) mutants have been identified in maize, few are commercially used due to a lack of effective methods to produce large quantities of pure male-sterile seeds. Here, we develop a multicontrol sterility (MCS) system based on the maize male sterility 7 (ms7) mutant and its wild-type Zea mays Male sterility 7 (ZmMs7) gene via a transgenic strategy, leading to the utilization of GMS in hybrid seed production. ZmMs7 is isolated by a map-based cloning approach and encodes a PHD-finger transcription factor orthologous to rice PTC1 and Arabidopsis MS1. The MCS transgenic maintainer lines are developed based on the ms7-6007 mutant transformed with MCS constructs containing the (i) ZmMs7 gene to restore fertility, (ii) alpha-amylase gene ZmAA and/or (iii) DNA adenine methylase gene Dam to devitalize transgenic pollen, (iv) red fluorescence protein gene DsRed2 or mCherry to mark transgenic seeds and (v) herbicide-resistant gene Bar for transgenic seed selection. Self-pollination of the MCS transgenic maintainer line produces transgenic red fluorescent seeds and nontransgenic normal colour seeds at a 1:1 ratio. Among them, all the fluorescent seeds are male fertile, but the seeds with a normal colour are male sterile. Cross-pollination of the transgenic plants to male-sterile plants propagates male-sterile seeds with high purity. Moreover, the transgene transmission rate through pollen of transgenic plants harbouring two pollen-disrupted genes is lower than that containing one pollen-disrupted gene. The MCS system has great potential to enhance the efficiency of maize male-sterile line propagation and commercial hybrid seed production.
DOI:10.1111/nph.14790URLPMID:28940573 [本文引用: 1]
Tapetum development and pollen production are regulated by a complex transcriptional network that consists of a group of tapetum-specific Arabidopsis transcription factors (TFs). Among these TFs, DEFECTIVE IN TAPETAL DEVELOPMENT AND FUNCTION 1 (TDF1) encodes an R2R3 MYB factor, and ABORTED MICROSPORE (AMS) encodes a basic helix-loop-helix (bHLH) factor. However, knowledge regarding the regulatory role of TDF1 in anther development remains limited. Here, we discovered that TDF1 directly regulates AMS via an AACCT cis-element. We found the precocious AMS transcript and absence of AMS protein in ams(-/-) gpTDF1:AMS-FLAG lines, suggesting the timing of the TDF1-regulated AMS expression is a prerequisite for AMS functioning. We found that TDF1 interacts with AMS. Additionally, the TDF1-AMS complex additively promotes the expression of AMS-regulated genes, suggesting that TDF1 and AMS regulate the downstream genes through a feed-forward loop. EPXB5, encoding a beta-expansin family protein, is another direct target of TDF1, and it is highly expressed in the tapetum and pollen grains. The TDF1-AMS complex acts in concert to activate EXPB5 expression through a feed-forward loop. The identification of the regulatory pathway between TDF1 and AMS provides an interlocked feed-forward loop circuit that precisely regulates the transcriptional cascades that support anther development.
DOI:10.1105/tpc.114.126292URL [本文引用: 1]
Male sterility plays an important role in F1 hybrid seed production. We identified a male-sterile rice (Oryza sativa) mutant with impaired pollen development and a single T-DNA insertion in the transcription factor gene bHLH142. Knockout mutants of bHLH142 exhibited retarded meiosis and defects in tapetal programmed cell death. RT-PCR and in situ hybridization analyses showed that bHLH142 is specifically expressed in the anther, in the tapetum, and in meiocytes during early meiosis. Three basic helix-loop-helix transcription factors, UDT1 (bHLH164), TDR1 (bHLH5), and EAT1/DTD1 (bHLH141) are known to function in rice pollen development. bHLH142 acts downstream of UDT1 and GAMYB but upstream of TDR1 and EAT1 in pollen development. In vivo and in vitro assays demonstrated that bHLH142 and TDR1 proteins interact. Transient promoter assays demonstrated that regulation of the EAT1 promoter requires bHLH142 and TDR1. Consistent with these results, 3D protein structure modeling predicted that bHLH142 and TDR1 form a heterodimer to bind to the EAT1 promoter. EAT1 positively regulates the expression of AP37 and AP25, which induce tapetal programmed cell death. Thus, in this study, we identified bHLH142 as having a pivotal role in tapetal programmed cell death and pollen development.
[本文引用: 1]
DOI:10.1104/pp.16.01385URLPMID:27837085 [本文引用: 1]
Pollen exine is essential for protection from the environment of the male gametes of seed-producing plants, but its assembly and composition remain poorly understood. We previously characterized Arabidopsis (Arabidopsis thaliana) mutants with abnormal pollen exine structure and morphology that we named kaonashi (kns). Here we describe the identification of the causal gene of kns4 that was found to be a member of the CAZy glycosyltransferase 31 gene family, identical to UNEVEN PATTERN OF EXINE1, and the biochemical characterization of the encoded protein. The characteristic exine phenotype in the kns4 mutant is related to an abnormality of the primexine matrix laid on the surface of developing microspores. Using light microscopy with a combination of type II arabinogalactan (AG) antibodies and staining with the arabinogalactan-protein (AGP)-specific beta-Glc Yariv reagent, we show that the levels of AGPs in the kns4 microspore primexine are considerably diminished, and their location differs from that of wild type, as does the distribution of pectin labeling. Furthermore, kns4 mutants exhibit reduced fertility as indicated by shorter fruit lengths and lower seed set compared to the wild type, confirming that KNS4 is critical for pollen viability and development. KNS4 was heterologously expressed in Nicotiana benthamiana, and was shown to possess beta-(1,3)-galactosyltransferase activity responsible for the synthesis of AG glycans that are present on both AGPs and/or the pectic polysaccharide rhamnogalacturonan I. These data demonstrate that defects in AGP/pectic glycans, caused by disruption of KNS4 function, impact pollen development and viability in Arabidopsis.
DOI:10.1093/jxb/erw445URLPMID:28082511 [本文引用: 1]
Lipid molecules are key structural components of plant male reproductive organs, such as the anther and pollen. Although advances have been made in the understanding of acyl lipids in plant reproduction, the metabolic pathways of other lipid compounds, particularly glycerolipids, are not fully understood. Here we report that an endoplasmic reticulum-localized enzyme, Glycerol-3-Phosphate Acyltransferase 3 (OsGPAT3), plays an indispensable role in anther development and pollen formation in rice. OsGPAT3 is preferentially expressed in the tapetum and microspores of the anther. Compared with wild-type plants, the osgpat3 mutant displays smaller, pale yellow anthers with defective anther cuticle, degenerated pollen with defective exine, and abnormal tapetum development and degeneration. Anthers of the osgpat3 mutant have dramatic reductions of all aliphatic lipid contents. The defective cuticle and pollen phenotype coincide well with the down-regulation of sets of genes involved in lipid metabolism and regulation of anther development. Taking these findings together, this work reveals the indispensable role of a monocot-specific glycerol-3-phosphate acyltransferase in male reproduction in rice.
DOI:10.1105/tpc.010926URLPMID:12724542 [本文引用: 1]
Insertional mutagenesis of Arabidopsis ecotype C24 was used to identify a novel mutant, designated wax2, that had alterations in both cuticle membrane and cuticular waxes. Arabidopsis mutants with altered cuticle membrane have not been reported previously. Compared with the wild type, the cuticle membrane of wax2 stems weighed 20.2% less, and when viewed using electron microscopy, it was 36.4% thicker, less opaque, and structurally disorganized. The total wax amount on wax2 leaves and stems was reduced by >78% and showed proportional deficiencies in the aldehydes, alkanes, secondary alcohols, and ketones, with increased acids, primary alcohols, and esters. Besides altered cuticle membranes, wax2 displayed postgenital fusion between aerial organs (especially in flower buds), reduced fertility under low humidity, increased epidermal permeability, and a reduction in stomatal index on adaxial and abaxial leaf surfaces. Thus, wax2 reveals a potential role for the cuticle as a suppressor of postgenital fusion and epidermal diffusion and as a mediator of both fertility and the development of epidermal architecture (via effects on stomatal index). The cloned WAX2 gene (verified by three independent allelic insertion mutants with identical phenotypes) codes for a predicted 632-amino acid integral membrane protein with a molecular mass of 72.3 kD and a theoretical pI of 8.78. WAX2 has six transmembrane domains, a His-rich diiron binding region at the N-terminal region, and a large soluble C-terminal domain. The N-terminal portion of WAX2 is homologous with members of the sterol desaturase family, whereas the C terminus of WAX2 is most similar to members of the short-chain dehydrogenase/reductase family. WAX2 has 32% identity to CER1, a protein required for wax production but not for cuticle membrane production. Based on these analyses, we predict that WAX2 has a metabolic function associated with both cuticle membrane and wax synthesis. These studies provide new insight into the genetics and biochemistry of plant cuticle production and elucidate new associations between the cuticle and diverse aspects of plant development.
DOI:10.1111/j.1365-313X.2008.03500.xURLPMID:18397379 [本文引用: 1]
In Arabidopsis, the tapetum plays important roles in anther development by providing enzymes for callose dissolution and materials for pollen-wall formation, and by supplying nutrients for pollen development. Here, we report the identification and characterization of a male-sterile mutant, defective in tapetal development and function 1 (tdf1), that exhibits irregular division and dysfunction of the tapetum. The TDF1 gene was characterized using a map-based cloning strategy, and was confirmed by genetic complementation. It encodes a putative R2R3 MYB transcription factor, and is highly expressed in the tapetum, meiocytes and microspores during anther development. Callose staining and gene expression analysis suggested that TDF1 may be a key component in controlling callose dissolution. Semi-quantitative and quantitative RT-PCR analysis showed that TDF1 acts downstream of DYT1 and upstream of AMS and AtMYB103 in the transcriptional regulatory networks that regulate tapetal development. In conclusion, our results show that TDF1 plays a vital role in tapetal differentiation and function.
DOI:10.1046/j.1365-313x.2003.01713.xURLPMID:12694597 [本文引用: 1]
A key step in pollen formation is the segregation of the products of male meiosis into a tetrad of microspores, each of which develops into a pollen grain. Separation of microspores does not occur in tetraspore (tes) mutants of Arabidopsis thaliana, owing to the failure of male meiotic cytokinesis. tes mutants thus generate large 'tetraspores' containing all the products of a single meiosis. Here, we report the positional cloning of the TES locus and details of the role played by the TES product in male cytokinesis. The predicted TES protein includes an N-terminal domain homologous to kinesin motors and a C-terminus with little similarity to other proteins except for a small number of plant kinesins. These include the Arabidopsis HINKEL protein and NACK1 and two from tobacco (Nishihama et al., 2002), which are involved in microtubule organization during mitotic cytokinesis. Immunocytochemistry shows that the characteristic radial arrays of microtubules associated with male meiotic cytokinesis fail to form in tes mutants. The TES protein therefore is likely to function as a microtubule-associated motor, playing a part either in the formation of the radial arrays that establish spore domains following meiosis, or in maintaining their stability.
DOI:10.1016/s0092-8674(02)00691-8URLPMID:11955449 [本文引用: 1]
The tobacco mitogen-activated protein kinase kinase kinase NPK1 regulates lateral expansion of the cell plate at cytokinesis. Here, we show that the kinesin-like proteins NACK1 and NACK2 act as activators of NPK1. Biochemical analysis suggests that direct binding of NACK1 to NPK1 stimulates kinase activity. NACK1 is accumulated specifically in M phase and colocalized with NPK1 at the phragmoplast equator. Overexpression of a truncated NACK1 protein that lacks the motor domain disrupts NPK1 concentration at the phragmoplast equator and cell plate formation. Incomplete cytokinesis is also observed when expression of NACK1 and NACK2 is repressed by virus-induced gene silencing and in embryonic cells from Arabidopsis mutants in which a NACK1 ortholog is disrupted. Thus, we conclude that expansion of the cell plate requires NACK1/2 to regulate the activity and localization of NPK1.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
Spontaneous genic male sterility will be a best genetic tool for hybrid seed production if uniform male sterile (ms, hereafter) lines are available. Some strategies were suggested to obtain such lines by means of cloning and transformation of male fertility (MF, hereafter) genes of spontaneous genic male sterility: (1) transferring MF gene into plastid genome of ms plants to suppress ms gene. The male fertile transgenic plants can be used as the maintainers to reproduce the genic male sterile lines when crossed to ms plants. (2) To comprise a lethal gene such as ribosomal inhibitor protein gene to transiently activate promoters that are active only in late embryogensis, then splice it with the MF gene, and transfer them into ms plants. The spikelets of transgenic plants may be fertile at heading stage and killed by the lethal gene when matured. When the transgenic plants are crossed with ms plants, their hybrid seeds with MF gene will be killed. The others with same nuclear as ms plants will survive. Immature seeds with the lethal gene can be harvested before maturing as the maintainer. (3) Gene switch system such as Cre/lox can snip out a stretch of DNA in plant genome. Its repressor protein prevents Cre recombinase gene from being switched on. Such stimulant as the antibiotic tetracycline can block the repressor protein. We can splice a DNA sequence that can be recognized by Cre recombinase to each end of a MF gene. By transferring this stretch of DNA and the gene switch system into ms plants, we can obtain a transgenic plant having homozygous MF gene, ms gene and the gene switch system. The transgenic plant may reproduce seeds like a conventional variety. When it is treated with the stimulant, the repressor protein will be blocked. The recombinase gene can work and snip out the MF gene. The fertile transgenic plants become male sterile then and can be used as the female parent for hybrid seeds production. (4)Take the native promoter from MF gene and replace it with a promoter that only functions in response to a chemical (such as antibiotic tetracycline), transfer the recombinant into ms line. The transgenic line would be fertile when the inducing chemical is present and used for the multiplication of ms plants. For hybrid seeds production the chemical will not be applied. Artificial male sterile genes and their restoration genes are also suggested to be used in the strategies.
URL [本文引用: 2]
Spontaneous genic male sterility will be a best genetic tool for hybrid seed production if uniform male sterile (ms, hereafter) lines are available. Some strategies were suggested to obtain such lines by means of cloning and transformation of male fertility (MF, hereafter) genes of spontaneous genic male sterility: (1) transferring MF gene into plastid genome of ms plants to suppress ms gene. The male fertile transgenic plants can be used as the maintainers to reproduce the genic male sterile lines when crossed to ms plants. (2) To comprise a lethal gene such as ribosomal inhibitor protein gene to transiently activate promoters that are active only in late embryogensis, then splice it with the MF gene, and transfer them into ms plants. The spikelets of transgenic plants may be fertile at heading stage and killed by the lethal gene when matured. When the transgenic plants are crossed with ms plants, their hybrid seeds with MF gene will be killed. The others with same nuclear as ms plants will survive. Immature seeds with the lethal gene can be harvested before maturing as the maintainer. (3) Gene switch system such as Cre/lox can snip out a stretch of DNA in plant genome. Its repressor protein prevents Cre recombinase gene from being switched on. Such stimulant as the antibiotic tetracycline can block the repressor protein. We can splice a DNA sequence that can be recognized by Cre recombinase to each end of a MF gene. By transferring this stretch of DNA and the gene switch system into ms plants, we can obtain a transgenic plant having homozygous MF gene, ms gene and the gene switch system. The transgenic plant may reproduce seeds like a conventional variety. When it is treated with the stimulant, the repressor protein will be blocked. The recombinase gene can work and snip out the MF gene. The fertile transgenic plants become male sterile then and can be used as the female parent for hybrid seeds production. (4)Take the native promoter from MF gene and replace it with a promoter that only functions in response to a chemical (such as antibiotic tetracycline), transfer the recombinant into ms line. The transgenic line would be fertile when the inducing chemical is present and used for the multiplication of ms plants. For hybrid seeds production the chemical will not be applied. Artificial male sterile genes and their restoration genes are also suggested to be used in the strategies.
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}