 ,江苏大学附属医院妇科,镇江 212001
,江苏大学附属医院妇科,镇江 212001Functional analysis of CG8005 gene in Drosophila testis
Wanyin Chen, Yidan Yan, Xiaojin Luan, Min Wang, Jie Fang,Department of Gynecology, Affiliated Hospital of Jiangsu University, Zhenjiang 212001, China通讯作者: 方杰,博士,副教授,主任医师,研究方向:妇科肿瘤。E-mail:fangjie070@163.com
编委: 史庆华
收稿日期:2020-06-1修回日期:2020-08-11网络出版日期:2020-11-20
| 基金资助: |
Received:2020-06-1Revised:2020-08-11Online:2020-11-20
| Fund supported: |
作者简介 About authors
陈万银,在读硕士研究生,专业方向:妇科肿瘤。E-mail:

摘要
生殖细胞的自我更新及分化过程对于男性不育及生殖细胞肿瘤的发生至关重要,CG8005基因作为果蝇(Drosophila melanogaster)睾丸生殖干细胞的调控因子之一,其功能机制尚不清楚。为了探讨CG8005基因在果蝇睾丸生殖细胞中的生物学功能,本研究首先通过UAS-gal4系统介导的RNAi途径,驱动果蝇睾丸生殖细胞和包囊细胞中UAS-CG8005 RNAi的表达,观察后代雄性果蝇的生育能力;其次,通过免疫荧光染色方法检测对照组和CG8005 RNAi雄性果蝇睾丸中Vasa、IBI、锌指结构域1蛋白(Zn finger homeodomain 1, Zfh1)、眼缺陷蛋白(eyes absent, Eya)、DE-cad、FasIII以及磷酸化组蛋白H3 (phospho-histone H3, PH3)、TUNEL等表达模式;最后,使用小干扰RNA (siRNA)在果蝇 S2 细胞中将 CG8005基因表达沉默,用PH3检测对照组和CG8005 siRNA组中果蝇S2细胞增殖能力,TUNEL及流式凋亡检测技术分析果蝇S2细胞凋亡情况;通过荧光定量PCR检测剪接体亚基的mRNA水平,观察剪接体的相对表达量。结果表明,与对照组果蝇比较,在生殖细胞和包囊细胞中缺失CG8005基因,雄性果蝇的生育能力降低甚至完全丧失;另外,nos-gal4驱动UAS-CG8005 RNAi的雄性果蝇中生殖细胞及生殖融合体消失,并且生殖细胞增殖能力减弱,而tj-gal4驱动UAS-CG8005 RNAi的果蝇睾丸中出现生殖细胞样肿瘤;在果蝇S2细胞中敲减CG8005基因导致细胞凋亡增加,增殖受到抑制;CG8005基因在果蝇S2细胞中沉默引起剪接体亚基的信使RNA水平升高。可见,CG8005基因在果蝇睾丸生殖细胞的自我更新与分化过程中必不可少,其缺失可能导致生殖细胞存活受限甚至生殖细胞样肿瘤的形成;并且CG8005基因可参与果蝇S2细胞的增殖和凋亡,对于细胞生命的维持至关重要,同时可竞争性调节剪接体亚基的mRNA水平。
关键词:
Abstract
The processes of self-renewal and differentiation of germ cells are crucial to the development of male infertility and germ cell tumors. CG8005 gene is one of the regulatory factors of the testicular germ stem cells in Drosophila melanogaster, and its functional mechanism is still unknown. To explore the biological function(s) of CG8005 gene in the germ cell niche of Drosophila testis, first, the UAS-gal4 system was used to drive the expression of UAS-CG8005 RNAi in Drosophila testicular germ cells and cyst cells. Fertility tests were then performed to determine the fertility rate of male flies. Second, the expression patterns of Vasa, IBI, Zn finger homeodomain 1 (Zfh1), eyes absent (Eya), DE-cad, FasIII and Phospho-Histone H3(PH3), and TUNEL were analyzed by immunofluorescence staining in both control and CG8005 RNAi testes. Lastly, small interfering RNA (siRNA) was used to silence the CG8005 gene expression in Drosophila S2 cells; and PH3 was used to detect the proliferation ability of Drosophila S2 cells in the control group and CG8005 siRNA group. Apoptosis of Drosophila S2 cells was analyzed with TUNEL and flow cytometry. To observe the relative expression of the spliceosome, the mRNA levels of the spliceosome subunits were detected by fluorescence quantitative RT-PCR. As compared with the control group, the results showed that deletion of the CG8005 gene in the germ cells and cyst cells of the testis reduced or even completely abolished the fertility of male fruit flies. In addition, nos-gal4 driven UAS-CG8005 RNAi led to loss of fusomes and reduce the proliferative ability of germ cells. Noticeably, tj-gal4-directed UAS-CG8005 RNAi knockdown of CG8005 gene in the testis led to germ cell tumor development. Knockdown of CG8005 gene in Drosophila S2 cells resulted in increase in apoptosis and inhibition of proliferation. Further, the silencing of the CG8005 gene in Drosophila S2 cells caused increases in the mRNA levels of the spliceosome subunits. Hence, CG8005 gene is essential for the self-renewal and differentiation of germ cells in Drosophila testis. Its deletion may lead to restricted germ cell survival and the formation of germ cell-like tumors. CG8005 gene can participate in the regulation of proliferation and apoptosis of Drosophila S2 cells, which is essential for the maintenance of cell life, and might competitively regulate the mRNA levels of spliceosome subunits.
Keywords:
PDF (1421KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
陈万银, 颜一丹, 栾晓瑾, 王敏, 方杰. CG8005基因在果蝇睾丸生殖细胞中的功能分析. 遗传[J], 2020, 42(11): 1122-1132 doi:10.16288/j.yczz.20-163
Wanyin Chen.
通常认为干细胞需要在特定的环境中才能保持自我更新及分化的性状,对于组织稳态及细胞生长至关重要[1,2]。果蝇(Drosophila melanogaster)睾丸中存在有两种干细胞类型,包括生殖干细胞(germline stem cells, GSCs)和体细胞干细胞(cyst stem cells, CySCs)[3,4],它们均与睾丸顶端的中心(Hub)细胞相连,共同构成生殖干细胞微环境(stem cell niche)[5,6,7]。GSCs产生的一个子代细胞经过4轮有丝分裂分化为16个精原细胞,继而经过精母细胞、圆形精子、长形精子和成熟精子完成整个生精过程[8]。两个由CySCs分化而来的包囊细胞包裹着生殖细胞并且支持着生殖细胞的生长[3]。而Hub细胞主要维持着GSCs和CySCs的自我更新及分化[9,10]。
干细胞微环境内的多条重要信号通路调控着干细胞的自我更新及分化过程[8,11],许多研究表明,Hub细胞通过分泌Upd因子在GSCs和CySCs激活JAK-STAT信号通路,维持这两类细胞自我更新的能力[12,13]。另外,Hub细胞通过CySCs激活Hh信号通路来维持干细胞自我更新的性状[14]。此外,大量研究证明多种细胞因子也参与调控干细胞微环境,其中,剪接体亚基与果蝇睾丸干细胞自我更新及分化之间存在密切关系。比如U2A剪接因子通过影响生殖细胞分化从而影响雄性果蝇生育能力[15]。PrP3作为PrP复合体中的关键剪接体成分,对于雄性果蝇生育及生殖系干细胞的自我更新及分化至关重要[16]。
果蝇遗传学研究中常用UAS-gal4系统异位表达转基因果蝇,用于激活UAS下游靶基因。Yu等[6]通过对2881个果蝇睾丸进行RNA干扰(RNAi),共鉴定到221个参与生殖细胞自我更新过程的调控因子,其中CG8005基因的表达具有差异性。CG8005基因在细胞周期进程及信使RNA衰减等过程中发挥作用,通过Flybase数据库(http://flybase.org/)显示CG8005基因与维持细胞壁完整性有关,起到凋亡调节剂的作用,而其对果蝇睾丸生殖细胞凋亡的影响尚不明确。因此,本研究分析了CG8005基因在果蝇睾丸生殖细胞中的生物学功能及CG8005基因对果蝇S2细胞增殖及凋亡的影响,探索了CG8005基因沉默与剪接体亚基之间的潜在联系,为研究CG8005基因的作用机制提供方向。
1 材料与方法
1.1 果蝇品系及饲养
UAS-CG8005 RNAi转基因果蝇购自清华果蝇中心(Tsinghua Fly Center, THFC),该品系来源于TRiP RNAi转基因果蝇库(Transgenic RNAi Project)。gal4转基因果蝇品系信息如下:nos-gal4 (BDSC, #4937),tj-gal4 (DGRC, #104055)。WT品系果蝇为野生型果蝇,作为对照组,来源于THFC果蝇库。本研究经江苏大学附属医院生物医学伦理学委员会批准。所有果蝇均在温度25℃、相对湿度60%的条件下饲养。
1.2 果蝇杂交策略
分别挑选nos-gal4、tj-gal4品系雄性果蝇与UAS-CG8005 RNAi的处女蝇进行杂交。在出生2 d内的F1代中挑选特定基因型(nos>CG8005 RNAi、tj> CG8005 RNAi)的果蝇用于后续实验。nos>CG8005 RNAi和tj>CG8005 RNAi基因型分别指的是在生殖细胞和包囊细胞中敲减CG8005基因。1.3 生育率测试
在F1代果蝇中挑选单只特定基因型(UAS- CG8005 RNAi)的成年雄蝇与3只野生型处女蝇杂交,观察其是否可以获得后代果蝇。通过统计F1代成年雄性果蝇生育比例判断雄性果蝇的生育能力。1.4 细胞培养与转染
利用快冻慢融的原理复苏果蝇S2细胞后,放入DMEM+10%胎牛血清(以色列Bioind公司)配制的培养基中进行培养,培养环境为28℃恒温培养箱(上海精宏实验设备有限公司),根据细胞生长速度和状态,及时进行细胞换液和细胞传代,并以适当密度重新接种至新的培养瓶中继续培养[17]。待细胞生长至80%~90%密度后进行转染。转染步骤如下:由苏州吉玛基因公司设计并合成siRNA序列,使用Lipo2000脂质体(美国Invitrogen公司)转染S2细胞。使用250 μL Opti-MEM (美国Gibco公司)稀释成15 μL的Lipo2000,轻轻混匀后室温放置5 min,在250 μL opti-MEM中加入15 μL小干扰RNA (siRNA),将两管混合物充分混匀后,室温放置20 min,使siRNA终浓度为150 nmol/L作为最终培养基加入细胞培养板中,6 h后更换成完全培养基继续培养至48 h。siRNA序列见表1。Table 1
Table 1siRNA sequences used in this study
| siRNA名称 | 序列(5′→3′) |
|---|---|
| 阴性对照 | F:UUCUCCG AACGUGUCACGUTT |
| R:ACGUGACACGUUCGGAGAATT | |
| siCG8005-1 | F:GGACCAAAUAGACAGCCAUTT |
| R:AUGGCUGUCUAUUUGGUCCTT | |
| siCG8005-2 | F:CCACGUUCAUGGGUUCAUUTT |
| R:AAUGAACCCAUGAACGUGGTT |
新窗口打开|下载CSV
1.5 实时荧光定量PCR (qRT-PCR)检测
果蝇S2细胞总RNA的提取及逆转录反应按照反转录试剂盒说明书(日本TaKaRa公司)进行。将目的DNA及相应的引物与SYBRGreen染料配成10 μL反应体系在反应管中充分混匀,4℃短暂离心,置入Mx3000P荧光定量PCR仪(美国安捷伦公司)中进行反应。反应条件为:95℃预变性10 min,95℃变性10 s,60℃退火20 s,72℃延伸10 s,40个循环。溶解曲线分析:95℃ 15 s,60℃ 1 min,95℃ 30 s,60℃ 15 s。以GAPDH作为内参,以Folds=2-ΔΔCt表示目的基因在实验组与对照组中表达的倍比关系。qRT-PCR引物序列见表2。Table 2
Table 2qRT-PCR primer sequences used in this study
| 基因名称 | 引物序列(5′→3′) |
|---|---|
| GAPDH | F: GTGGTGAACGGCCAGAAGAT |
| R: GCCTTGTCAATGGTGGTGAA | |
| CG8005 | F: CTGGACTCGTGGTGGACATTCTG |
| R: ACACTGAGTAATCCGCTCCATTGC | |
| Prp19 | F: GCTGCCACGAAGGACCTGTTAC |
| R: CCTGTGCGGATATCGGAGAATGC | |
| PrP18 | F: GCTGCTCACCTTCCTGCTCAAG |
| R: AGCGGCTTAACGTATTCCTTAGTCTG | |
| SmB | F: CATGAACTTGATCCTCGGCGACTG |
| R: CCTCTGGCGGCGGTGGTC | |
| SmD1 | F: CACCTGAAGAGCGTTCGGATGAC |
| R: TGTCGTCGATGAGGAGCGTCTC | |
| SmE | F: CCATCAACCTGATCTTCCGTTACCTG |
| R: GCGTCGTCCAGCACCAGATTC | |
| SmF | F: GCTCCGTGACTGGTAATCTTGGC |
| R: TCCTCGTCGTCGTCCTCCATG | |
| SmG | F: TCTGAGCAGCGTGGTTATTGACATC |
| R: CGTAGCATGGCATCGAGTCCTTG |
新窗口打开|下载CSV
1.6 免疫荧光检测果蝇S2细胞的增殖与凋亡
将转染24 h后的果蝇S2细胞放入六孔板中进行种板,孵育24 h后在培养皿中进行免疫染色。具体步骤如下:4%多聚甲醛固定30 min。在含有0.1%Triton X-100(PBST)的1×PBS中洗涤3次,并在5%牛血清白蛋白BSA (上海生工生物工程股份有限公司)中封闭30 min后,将样品与一抗(Vasa购自美国Santa Cruz Biotechnology公司,1B1、Zfh1、Eya、DE-cad、FasIII均购自美国Developmental Studies Hybridoma Bank公司,PH3购自美国Cell Signaling Technology公司)在室温孵育1 h或4℃过夜。之后0.1%PBST中洗涤3次,每次10 min。然后将样品在室温下与二抗(包括488-兔二抗、cy3-小鼠二抗、647-大鼠二抗,均购自美国Molecular Probes and Jackson Immunologicals公司)一起孵育1 h,之后在0.1%PBST中洗涤3次。用Hoechst 33342(美国Invitrogen公司)染色10 min,加入20 μL 80%甘油,盖上盖玻片封片。在奥林巴斯BX51荧光正置显微镜上捕获图像并使用Adobe Photoshop CS5软件处理。使用Image J软件对荧光强度进行定量分析。1.7 流式细胞分析果蝇S2细胞凋亡
将siRNA转染48 h后的果蝇S2细胞,根据Annexin V-Alexa Fluor 647/PI细胞凋亡检测试剂盒(南京福麦斯生物技术有限公司)的说明书进行操作。根据细胞计数取1×106个细胞进行离心,用250 μL结合换缓冲液重悬细胞沉淀,与2.5 μL Annexin V-Alexa Fluor 647和5 μL PI混合,然后将其在黑暗室温下孵育15 min,每个样品加入200 μL PBS,使用FACScan流式细胞仪(美国BD Biosciences公司)分析样品。1.8 统计学分析
采用Graphpad软件(2 结果与分析
2.1 在生殖细胞与包囊细胞中敲减CG8005基因导致雄性果蝇生育率下降
为了探究CG8005基因与雄性果蝇生育能力的相关性,本研究分别在生殖细胞和包囊细胞中表达的gal4来驱动UAS-CG8005 RNAi。生育率测试(表3)结果表明,生殖细胞中敲减CG8005基因(nos> CG8005 RNAi)导致雄性果蝇完全不育,生育率为0.00% (n=59, P<0.001);包囊细胞中敲减CG8005基因(tj>CG8005 RNAi),雄性果蝇生育率下降至19.67% (n=61, P<0.001)。2.2 在生殖细胞中敲低CG8005基因导致生殖细胞缺失
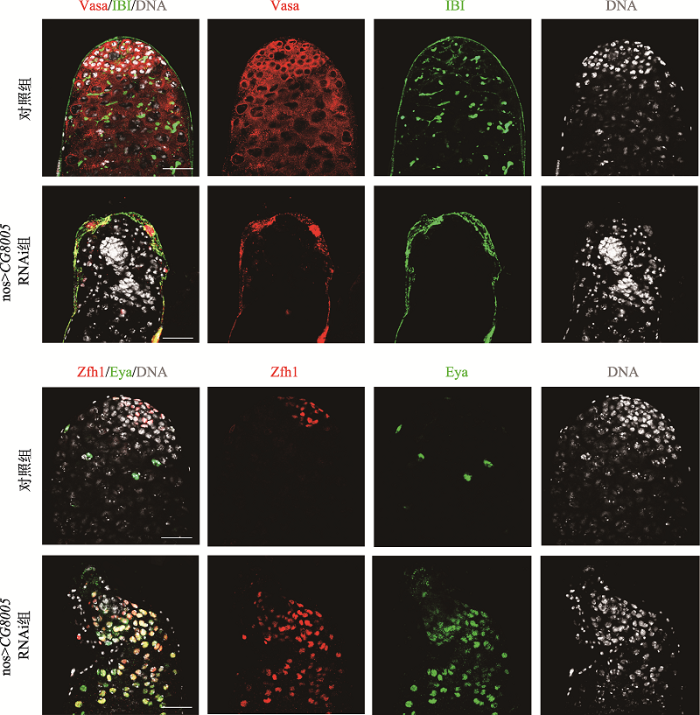
本研究首先利用免疫荧光技术,并通过多种睾丸细胞相关抗体观察CG8005基因在果蝇睾丸中的功能。其中,生殖细胞可以被Vasa抗体标记,融合体(fusome)可用IBI蛋白标记,其在睾丸头部顶端呈点状分布,随着生殖细胞的分化,逐渐发展成树杈状;CySCs可被锌指结构域1蛋白(Zn finger homeodomain 1, Zfh1)标记;成熟的包囊细胞被眼缺陷蛋白(eyes absent, Eya)标记。通过共聚焦显微镜观察结果显示,与对照组相比,nos>CG8005 RNAi组睾丸形态坍塌、内部结构紊乱,呈现小睾丸表型,此时生殖细胞及融合体均全部消失(图1)。为了进一步探索被DNA标记的细胞核来源,利用体细胞相关免疫抗体进行检测,睾丸顶部的CySCs和包囊细胞的细胞数目明显增多,Eya积累与Zfh1信号发生重合(图1)。以上结果提示,CG8005基因在早期生殖细胞中导致生殖细胞缺失。Table 3
表3
表3敲减CG8005基因雄性果蝇生育率的测试结果
Table 3
| 组别 | 总数 | 可育数 | 生育率 | P值 |
|---|---|---|---|---|
| 野生型 | 74 | 71 | 95.94% | - |
| nos>CG8005 RNAi | 59 | 0 | 0 | <0.001 |
| tj>CG8005 RNAi | 61 | 12 | 19.67% | <0.001 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1缺失CG8005基因生殖细胞的表达模式
Vasa抗体(红色)标记生殖细胞,IBI抗体(绿色)标记生殖融合体。Zfh1抗体(红色)标记体细胞干细胞,Eya抗体(绿色)标记包囊细胞,DNA标记细胞核(灰色)。标尺:20 μm。
Fig. 1The expression patterns of germ cells with the knockdown of the CG8005 gene
2.3 在生殖细胞中敲低CG8005基因生殖细胞增殖能力减弱
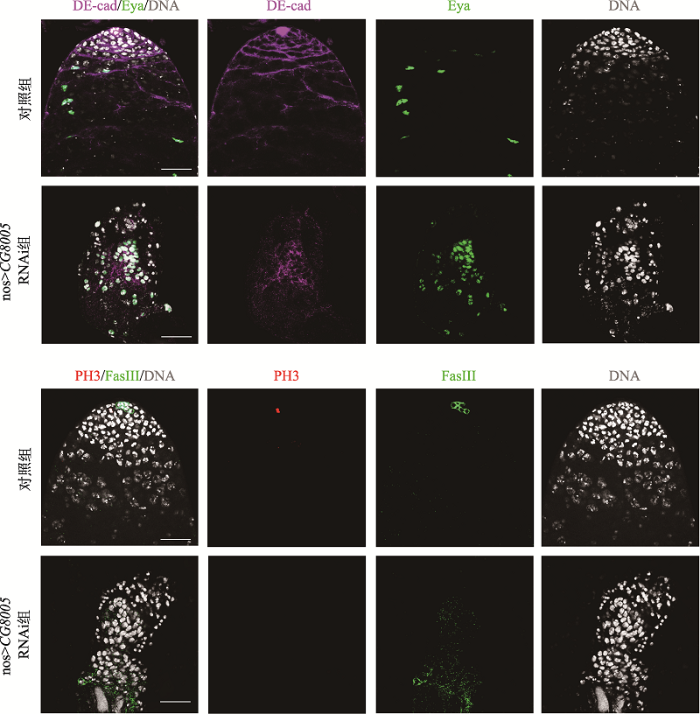
黏附蛋白DE-cad是中心细胞和体细胞的标记物,生殖细胞中缺失CG8005基因,睾丸头部顶端的中心细胞消失不可见,体细胞与增多的包囊细胞同步定位(图2),包囊细胞属于种系而不是生殖系。磷酸化组蛋白 H3 (phospho-histone H3,PH3)作为增殖细胞的标记物,在正常果蝇睾丸早期生殖细胞中仅存在1~3个。本研究进一步利用免疫荧光技术检测生殖细胞消失后其增殖能力的改变,结果显示在对照组果蝇睾丸头部顶端可检测到PH3信号,而nos>CG8005 RNAi组中未检测到PH3表达(图2),提示生殖细胞缺失可能与其增殖能力减弱有关。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2生殖细胞中敲低CG8005基因对生殖细胞增殖能力的影响
体细胞被DE-cad抗体(紫色)识别,Eya抗体(绿色)标记包囊细胞。PH3抗体(红色)检测细胞增殖能力,FasIII抗体(绿色)标记中心细胞,DNA标记细胞核(灰色)。标尺:20 μm。
Fig. 2Effects of CG8005 gene knockdown on the cell proliferation ability in germ cells
2.4 在包囊细胞中敲低CG8005基因影响生殖细胞分化
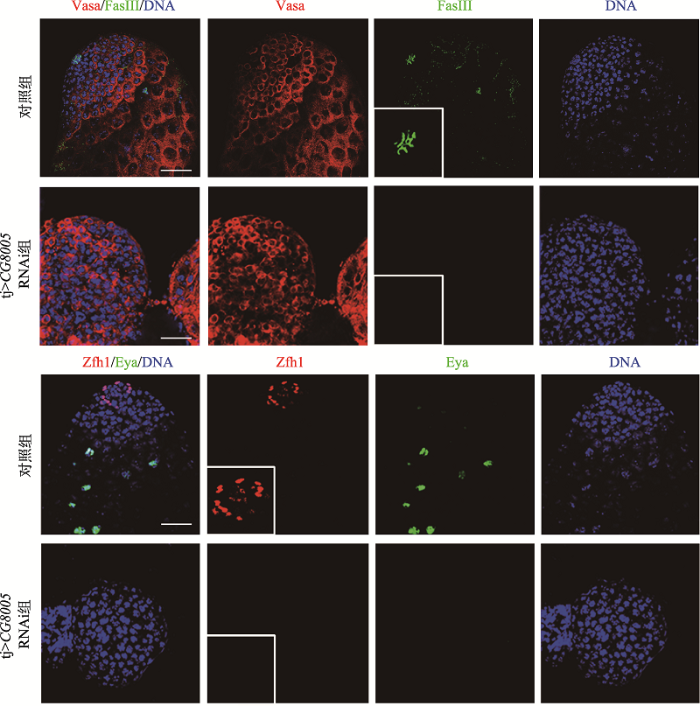
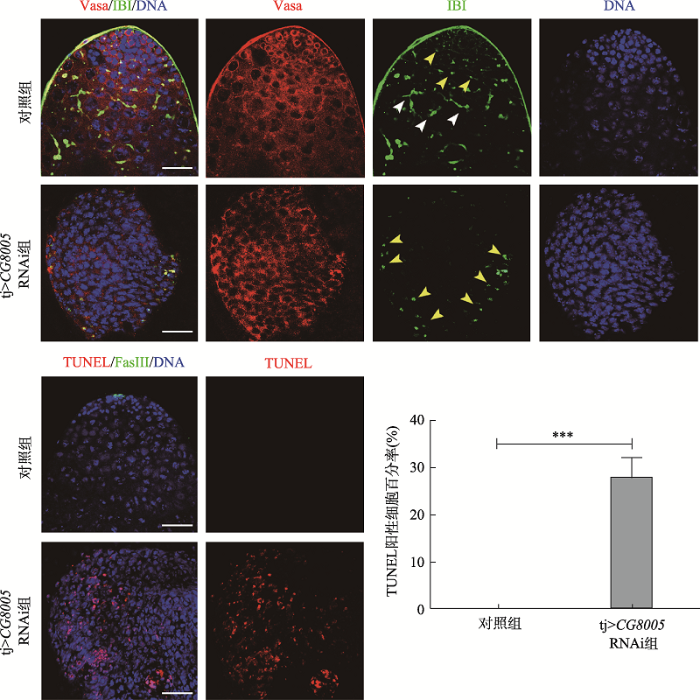
为了探索果蝇睾丸中CG8005基因在包囊细胞中的功能,本研究利用tj-gal4在包囊细胞中敲低CG8005基因。免疫荧光结果显示,对照组睾丸顶部的中心细胞被FasIII抗体识别,而实验组中FasIII信号消失,Zfh1和Eya染色结果显示不存在CySCs和包囊细胞(图3)。有研究表明,中心细胞参与调控生殖干细胞自我更新与分化过程,本研究结果显示,tj>CG8005 RNAi组睾丸失去了正常的形态,中心细胞缺失,并在果蝇睾丸中出现被Vasa抗体标记的异常细胞团块,此时树杈状的融合体变成点状(图4),最终导致细胞过度增殖,形成生殖细胞样肿瘤。这些未分化的生殖细胞团块被TUNEL(末端脱氧核苷酸转移酶介导的dUTP-生物素缺口末端)标记,统计学结果显示凋亡细胞比率近30% (图4),提示这些细胞具有凋亡能力,表明CG8005基因是生殖细胞形成所必需的。以上结果揭示了CG8005基因在包囊细胞中影响生殖细胞的分化过程。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3在包囊细胞中敲低CG8005基因对生殖细胞及体细胞的影响
Vasa抗体(红色)标记生殖细胞,FasIII抗体(绿色,放大)标记中心细胞。Zfh1抗体(红色,放大)标记体细胞干细胞,包囊细胞被Eya抗体(绿色)识别。DNA标记细胞核(蓝色)。标尺:20 μm。
Fig. 3Effects of CG8005 gene knockdown on germ and somatic cells among the cyst cells
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4在包囊细胞中敲低CG8005基因对生殖融合体和细胞凋亡影响
IBI抗体(绿色)标记生殖细胞间融合体,黄色箭头表示点状融合体,白色箭头表示树杈状融合体。TUNEL末端脱氧核苷酸转移酶介导的dUTP-生物素缺口末端。柱状图表示TUNEL阳性细胞百分比。DNA标记细胞核(蓝色)。标尺:20 μm。
Fig. 4Effects of CG8005 gene knockdown on fusomes and apoptosis among the cyst cells
2.5 CG8005基因缺失抑制果蝇S2细胞增殖
为了进一步探究CG8005基因的体外功能,本研究使用两个siRNA (siCG8005-1和siCG8005-2)将CG8005基因在果蝇S2细胞中表达沉默。通过荧光定量PCR检测对照组、siCG8005-1组和siCG8005-2组中CG8005 信使RNA (mRNA)表达水平,结果显示片段siCG8005-1敲减效率52%,而siCG8005-2敲减效率达70%,因此选用片段siCG8005-2进行后续实验(图5A)。在果蝇S2细胞中敲减CG8005基因,用检测细胞增殖能力标记物PH3进行免疫荧光染色,结果显示,与对照组相比,siCG8005-2组PH3阳性细胞减少(图5B),对照组PH3阳性信号百分比为5.8%,实验组下降至4.3%,具有统计学差异(图5C)。以上结果表明,在果蝇S2 细胞中敲减 CG8005基因抑制细胞的增殖。图5
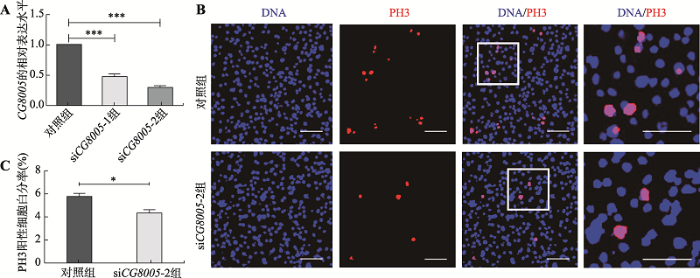 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5在果蝇S2细胞中缺失CG8005基因对细胞增殖能力的影响
A:qRT-PCR法检测在果蝇S2细胞中转染siCG8005-1、siCG8005-2验证干涉效率;B:免疫荧光检测在果蝇S2细胞中转染siCG8005-2细胞的增殖能力;C: PH3阳性细胞百分比。* P <0.05;***P <0.001。标尺:30 μm。
Fig. 5Effects of CG8005 gene knockdown on the proliferation ability of the Drosophila S2 cells
2.6 CG8005基因缺失导致果蝇S2细胞凋亡
运用UAS-gal4系统实现CG8005基因在睾丸中缺失,导致生殖细胞自我更新和分化异常。本研究进一步分析CG8005基因对果蝇S2细胞存活的影响,利用TUNEL免疫荧光技术检测凋亡细胞,结果显示,与对照组相比,siCG8005-2组果蝇S2细胞中TUNEL阳性细胞增加(图6A),实验组中TUNEL阳性细胞百分比从2%上升至7% (图6B)。流式细胞术的检测结果与TUNEL免疫荧光结果一致(图6C),凋亡细胞增加存在统计学意义(图6D)。反映CG8005基因敲减后,果蝇S2细胞凋亡增加。图6
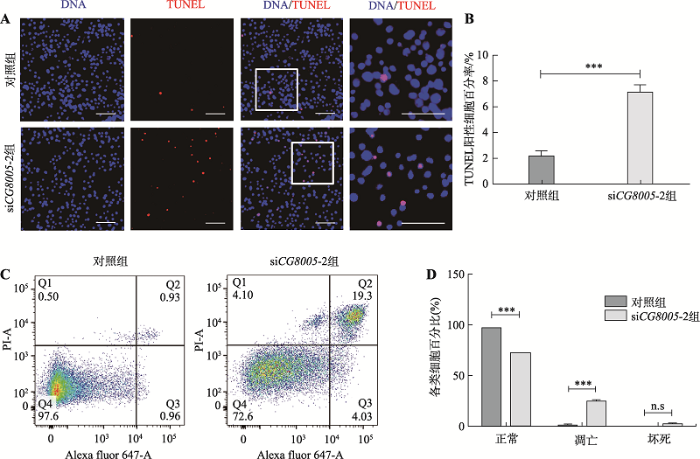 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6在果蝇S2细胞中缺失CG8005基因对细胞凋亡水平的影响
A:免疫荧光检测在果蝇S2细胞中转染siCG8005-2细胞的凋亡情况;B:TUNEL阳性细胞百分比;C:流式检测细胞凋亡情况;D:流式各组细胞百分比。***P <0.001;n.s 无统计学差异。标尺:30 μm。
Fig. 6Effects of CG8005 gene knockdown on the apoptosis level of the Drosophila S2 cells
2.7 CG8005基因沉默影响剪接体亚基的表达
剪接体在疾病的发生中具有重要的意义,主要的剪接体对于mRNA加工和细胞存活至关重要,剪接体亚基U2A的突变会损害精原细胞的分化,阻碍生殖细胞成熟为精子。为了初步探索CG8005基因与剪接体亚基之间的联系,本研究主要利用qRT-PCR方法,对主要剪接体的相对表达量进行分析。结果表明,剪接体亚基(Prp18、Prp19、SmB、SmD1、SmE、SmF和SmG)的mRNA水平均在敲减CG8005基因的果蝇S2细胞中表达上调(图7),提示 CG8005基因沉默可能促进了剪接体亚基的表达,从而影响它们的功能。图7
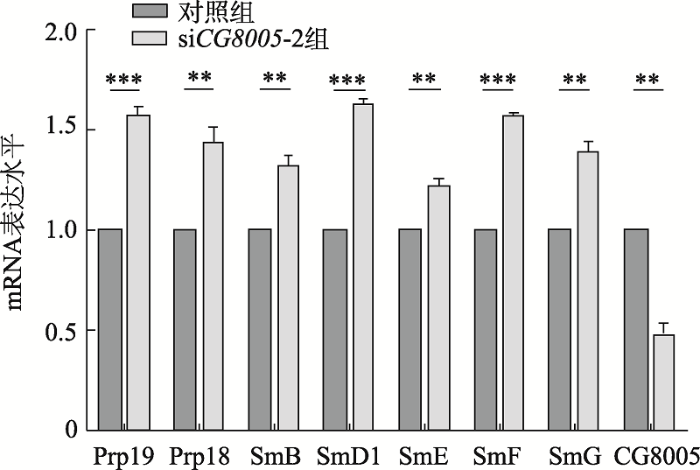 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7CG8005基因沉默对剪接体亚基表达水平的影响
**P <0.01;***P <0.001)。
Fig. 7Effects of CG8005 gene knockdown on the expression of spliceosome subunits
3 讨论
精子发生过程中任何障碍最终可能导致男性不育,其中生殖细胞的功能异常对于男性生育能力尤为重要。并且有证据显示,生殖细胞肿瘤患者在年轻的时候存在生育相关问题,提示不育症可能是生殖细胞肿瘤发病的高危因素[17]。果蝇具有繁殖时间短、遗传操作性强、靶基因的RNAi品系资源丰富等优势,在雄性生殖系统(特别是生殖细胞)的研究中具有优势。果蝇睾丸中GSCs是果蝇精子产生的源头,CySCs为生殖细胞的增殖、生长及分化提供环境,它们的自我更新与分化过程均需要Hub细胞的维持[18]。Yu等[6]运用果蝇模型在睾丸中筛选到221个生殖干细胞调控因子,并进一步对这些调控因子进行深入研究,发现一些调控因子可以通过包囊细胞影响生殖细胞的分化过程,最终形成生殖细胞样肿瘤[6,19]。
CG8005蛋白是一种可能的脱氧苏氨酸合酶,Bing在线工具分析CG8005基因参与细胞周期进程、信使RNA衰减等过程,可催化亚精胺的NAD依赖性氧化裂解,与真核翻译起始因子5A(elf-5A)的相互作用蛋白,它可能参与应激反应并维持细胞壁的完整性,充当细胞凋亡的调节剂。然而,CG8005基因在果蝇睾丸生殖细胞中的研究报道十分少见。
本研究结果提示,缺失CG8005基因雄性果蝇尽管可以存活,但其生育能力明显下降。更重要的是,nos-gal4主要在生殖干细胞和精原细胞中表达,并在生精过程中逐渐下降,CG8005 RNAi组中睾丸形态缩小,生殖细胞及融合体消失,包囊细胞及CySCs代偿性增加,尽管这些细胞具有一定的增殖能力,最终依然导致生殖细胞死亡。而tj-gal4主要在体细胞的包囊细胞中表达,有研究报道GSCs正常分化过程受到CySCs的调控,CySCs缺陷可以介导生殖细胞分化阻滞最终发展为生殖细胞肿瘤[1]。本研究运用tj-gal4介导CG8005 RNAi使用Vasa抗体观察这些过度增殖细胞的表达模式,结果发现异常的细胞团块被生殖细胞信号捕捉,通过FasIII的表达模式中心细胞缺失,连接生殖细胞之间的融合体逐渐由树杈状变为点状,CySCs和包囊细胞消失更进一步的说明细胞不受调控的增长,形成生殖细胞肿瘤。S2细胞是果蝇中经典的细胞模型,在果蝇S2细胞中敲减CG8005基因导致细胞数目减少,抑制细胞增殖并加重细胞凋亡。最后,CG8005基因可能竞争性地影响剪接体亚基的表达,已有证据表明剪接体复合物可能在无精症和生殖细胞肿瘤中起关键作用[16],使用种系干细胞模型将在未来的研究中进一步评估。
综上所述,本研究初步揭示了CG8005基因在果蝇睾丸生殖细胞中的生物学功能,CG8005基因是一个果蝇睾丸优势表达的生殖细胞作用因子,在生殖干细胞微环境中发挥重要的作用,主要影响生殖细胞的自我更新和分化过程。干涉效率验证筛选到CG8005 siRNA片段,为制备CG8005 RNAi果蝇提供靶向序列。而CG8005基因与剪接体亚基之间的竞争性关系为未来的研究提供方向,为理解男性不育及睾丸生殖细胞肿瘤的发病机理提供实验基础。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
DOI:10.1126/science.1140861URLPMID:17446390 [本文引用: 1]

Drosophila male and female germline stem cells (GSCs) are sustained by niches and regulatory pathways whose common principles serve as models for understanding mammalian stem cells. Despite striking cellular and genetic similarities that suggest a common evolutionary origin, however, male and female GSCs also display important differences. Comparing these two stem cells and their niches in detail is likely to reveal how a common heritage has been adapted to the differing requirements of male and female gamete production.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLPMID:27471256 [本文引用: 4]
[本文引用: 1]
[本文引用: 2]
URLPMID:31160709 [本文引用: 1]
URLPMID:31295251 [本文引用: 1]
[本文引用: 1]
URLPMID:11752574 [本文引用: 1]
[本文引用: 1]
URLPMID:23419515 [本文引用: 1]
[本文引用: 1]
URLPMID:31292463 [本文引用: 2]
URLPMID:26651223 [本文引用: 2]
[本文引用: 1]
URLPMID:25503408 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}