Current advance on molecular genetic regulation of rice fertility
Yongyao Xie1, Jintao Tang1, Bowen Yang1, Jun Hu2, Yao-Guang Liu1, Letian Chen,11. State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, College of Life Sciences, South China Agricultural University, Guangzhou 510642, China 2. State Key Laboratory of Hybrid Rice, College of Life Sciences, Wuhan University, Wuhan 430072, China
Supported by the National Natural Science Foundation of China.31701499 The Key Research Program of Guangzhou Science Technology and Innovation Commission.201707020016 The China Postdoctoral Science Foundation.2018M630955 The State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources.SKLCUSA-b201717
作者简介 About authors 谢勇尧,博士,博士后,研究方向:作物育性分子机制与杂种优势利用E-mail:xieyongyao2005@163.com。
Abstract Hybrid rice has contributed greatly to global food security. Cytoplasmic male sterility (CMS) and photo/ thermo sensitive genic male sterility (P/TGMS) are genetic bases for three-line and two-line hybrid rice production, respectively. In contrast, (sub-) specific hybrid sterility (HS) is a major barrier for utilization of hybrid vigor of distant hybrid rice. Therefore, understanding the molecular regulatory mechanism of rice fertility is a key technical issue for hybrid rice industry, and a long-standing basic scientific issue for nuclear-cytoplasmic interaction and reproductive isolation. Chinese geneticists of plant sciences have made tremendous contributions on the molecular genetic basis of rice fertility related to hybrid rice production. Here, we review the development of hybrid rice production systems in China and summarize current advance on genetic basis and molecular mechanism of CMS, P/TGMS, and HS involved in hybrid rice. We also discuss problems of hybrid rice production in China and point out new direction for future utilization of heterosis in rice. Keywords:hybrid rice;cytoplasmic male sterility;hybrid sterility;photo/thermo sensitive genic male sterility;fertility restoration
PDF (497KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 谢勇尧, 汤金涛, 杨博文, 胡骏, 刘耀光, 陈乐天. 水稻育性调控的分子遗传研究进展[J]. 遗传, 2019, 41(8): 703-715 doi:10.16288/j.yczz.19-162 Yongyao Xie, Jintao Tang, Bowen Yang, Jun Hu, Yao-Guang Liu, Letian Chen. Current advance on molecular genetic regulation of rice fertility[J]. Hereditas(Beijing), 2019, 41(8): 703-715 doi:10.16288/j.yczz.19-162
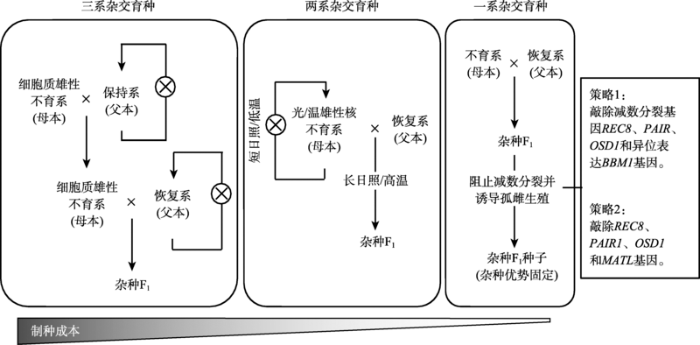
杂交水稻育种体系分为三系、两系、一系3个发展阶段。三系杂交育种体系由保持系、细胞质雄性不育系和恢复系组成。保持系和恢复系能够通过自交进行扩繁,而细胞质雄性不育系通过与保持系杂交进行繁种,不育系与恢复系杂交获得杂种F1。两系杂交育种体系由光/温敏不育系和恢复系组成。光/温敏不育系在短日照或低温条件下可育,能自交繁殖;在长日照或高温条件下不育,与恢复系杂交获得杂种F1。一系杂交育种体系则是通过杂种F1的无融合生殖,实现持续繁殖杂交种,固定杂种优势。 Fig. 1The development of hybrid rice breeding systems
三系育种中,水稻细胞质雄性不育系的发现与利用是三系杂交稻的杂种优势应用的基础,对粮食安全具有划时代的意义[9]。目前,野败型细胞质雄性不育(wild-abortive type cytoplasmic male sterility, CMS-WA)、包台型细胞质雄性不育(BoroⅡtype CMS, CMS-BT)和红莲型细胞质雄性不育(Hong-Lian type CMS, CMS-HL)是应用于三系杂交稻的3大细胞质雄性不育系。其中,CMS-WA和CMS-HL是由我国科学家袁隆平院士团队和朱英国院士团队独立发掘的种质资源[3]。这些不育系已经推广应用了几十年,但长期以来关于它们的育性控制分子机制仍不清楚。直到近10多年,我国植物遗传学家才一一揭开了谜底,为阐明控制三大不育系的不育与恢复分子机制做出了突出贡献。
A:智能型雄性不育系繁种及杂交育种原理;B:智能型雌性不育系繁种及杂交育种原理。它们的核心都是利用核雄性不育突变体(msms)或核雌性不育突变体(fsfs)作为受体,转入一个含有雄性或雌性功能型基因MS或FS、特异雄配子致死基因(gamete killer, GK)和红色荧光素酶基因(RFP)的转基因元件(MS-GK-RFPt或FS-GK-RFPt),形成转基因保持系(msms/MS-GK-RFPt—或fsfs/FS-GK-RFPt—)。在其分离后代中,MS或FS可以回补孢子体核不育表型,GK能杀死含有转基因元件的雄配子,RFP能作为选择标记筛选出带转基因的保持系用于智能不育系的繁种。不带转基因的雄性不育系或雌性不育恢复系则用于杂交制种。虚线指示的基因型个体是致死的,理论上不出现。 Fig. 2The application and fertility regulation of intelligent genic male or female sterility lines
LiN, ZhangDS, LiuHS, YinCS, LiXX, LiangWQ, YuanZ, XuB, ChuHW, WangJ, WenTQ, HuangH, LuoD, MaH, ZhangDB . The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development Plant Cell, 2006,18(11):2999-3014. [本文引用: 1]
BalkJ, LeaverCJ . The PET1-CMS mitochondrial mutation in sunflower is associated with premature programmed cell death and cytochrome c release Plant Cell, 2001,13(8):1803-1818. [本文引用: 1]
NotsuY, MasoodS, NishikawaT, KuboN, AkidukiG, NakazonoM, HiraiA, KadowakiK . The complete sequence of the rice (Oryza sativa L.) mitochondrial genome: frequent DNA sequence acquisition and loss during the evolution of flowering plants Mol Genet Genomics, 2002,268(4):434-445. [本文引用: 1]
LiuZL, XuH, GuoJX, LiuYG . Structural and expressional variations of the mitochondrial genome conferring the wild abortive type of cytoplasmic male sterility in rice J Integr Plant Biol, 2010,49(6):908-914. [本文引用: 1]
DeweyRE, SiedowJN, TimothyDH , Levings CS 3rd. A 13-kilodalton maize mitochondrial protein in E. coli confers sensitivity to Bipolaris maydis toxin Science, 1988,239(4837):293-295. [本文引用: 1]
WangZ, ZouY, LiX, ZhangQ, ChenL, WuH, SuD, ChenY, GuoJ, LuoD, LongY, ZhongY, LiuYG . Cytoplasmic male sterility of rice with boro II cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing Plant Cell, 2006,18(3):676-687. [本文引用: 4]
ShinjyoC . Cytoplasmic genetic male sterility in cultivated rice,Oryza sativa L. II The inheritance of male sterility .. Jpn J Genet, 1969,44(3):149-156. [本文引用: 1]
SabarM, GagliardiD, BalkJ, LeaverCJ . ORFB is a subunit of F1F(O)-ATP synthase: insight into the basis of cytoplasmic male sterility in sunflower EMBO Rep, 2003,4(4):381-386. [本文引用: 1]
WangK, GaoF, JiY, LiuY, DanZ, YangP, ZhuY, LiS . ORFH79 impairs mitochondrial function via interaction with a subunit of electron transport chain complex III in Honglian cytoplasmic male sterile rice New Phytol, 2013,198(2):408-418. [本文引用: 2]
YiP, WangL, SunQP, ZhuYG . Identification of mitochondrially chimeric gene associated with Honglian type of cytoplasmic male sterility of rice Chin Sci Bull, 2002(2):130-133. [本文引用: 1]
PengX, WangK, HuC, ZhuY, WangT, YangJ, TongJ, LiS, ZhuY . The mitochondrial gene orfH79 plays a critical role in impairing both male gametophyte development and root growth in CMS-Honglian rice BMC Plant Biol, 2010,10:125. [本文引用: 1]
TangH, LuoD, ZhouD, ZhangQ, TianD, ZhengX, ChenL, LiuYG . The rice restorer Rf4 for Wild-Abortive cytoplasmic male sterility encodes a mitochondrial-localized PPR protein that functions in reduction of WA352 transcripts Mol Plant, 2014,7(9):1497-1500. [本文引用: 1]
HuJ, WangK, HuangW, LiuG, GaoY, WangJ, HuangQ, JiY, QinX, WanL, ZhuR, LiS, YangD, ZhuY . The rice pentatricopeptide repeat protein RF5 restores fertility in Hong-Lian cytoplasmic male-sterile lines via a complex with the glycine-rich protein GRP162 Plant Cell, 2012,24(1):109-122. [本文引用: 1]
HuangW, YuC, HuJ, WangL, DanZ, ZhouW, HeC, ZengY, YaoG, QiJ, ZhangZ, ZhuR, ChenX, ZhuY . Pentatricopeptide-repeat family protein RF6 functions with hexokinase 6 to rescue rice cytoplasmic male sterility Proc Natl Acad Sci USA, 2015,112(48):14984-14989. [本文引用: 1]
ShiMS . The discovery and preliminary studies of the photoperiod-sensitive recessive male sterile rice (Oryza sativa L. subsp. japonica) Sci Agric Sin, 1985,18(2):44-48. Magsci [本文引用: 1]
MeiM, DaiX, XuC, ZhangQ . Mapping and genetic analysis of the genes for photoperiod-sensitive genic male sterility in rice using the original mutant Nongken 58S Crop Sci, 1999,39(6):1711-1715. [本文引用: 1]
DingJ, LuQ, OuyangY, MaoH, ZhangP, YaoJ, XuC, LiX, XiaoJ, ZhangQ . A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice Proc Natl Acad Sci USA, 2012,109(7):2654-2659. [本文引用: 2]
ZhouH, LiuQ, LiJ, JiangD, ZhouL, WuP, LuS, LiF, ZhuL, LiuZ, ChenL, LiuYG, ZhuangC . Photoperiod- and thermo-sensitive genic male sterility in rice are caused by a point mutation in a novel noncoding RNA that produces a small RNA Cell Res, 2012,22(4):649-660. [本文引用: 2]
ZhouH, ZhouM, YangY, LiJ, ZhuL, JiangD, DongJ, LiuQ, GuL, ZhouL, FengM, QinP, HuX, SongC, ShiJ, SongX, NiE, WuX, DengQ, LiuZ, ChenM, LiuYG, CaoX, ZhuangC . RNase Z S1processes UbL40 mRNAs and controls thermosensitive genic male sterility in rice Nat Commun, 2014,5:4884. [本文引用: 2]
ZhangH, XuC, HeY, ZongJ, YangX, SiH, SunZ, HuJ, LiangW, ZhangD . Mutation in CSA creates a new photoperiod-sensitive genic male sterile line applicable for hybrid rice seed production Proc Natl Acad Sci USA, 2013,110(1):76-81. [本文引用: 2]
YuJ, HanJ, KimYJ, SongM, YangZ, HeY, FuR, LuoZ, HuJ, LiangW, ZhangD . Two rice receptor-like kinases maintain male fertility under changing temperatures Proc Natl Acad Sci USA, 2017,114(46):12327-12332. [本文引用: 2]
OuyangY, ZhangQ . Understanding reproductive isolation based on the rice model Annu Rev Plant Biol, 2013,64:111-135. [本文引用: 1]
OuyangYD . Progress of indica-japonica hybrid sterility and wide-compatibility in rice Chin Sci Bull, 2016,61(35):3833-3841. [本文引用: 1]
ChenJ, DingJ, OuyangY, DuH, YangJ, ChengK, ZhaoJ, QiuS, ZhangX, YaoJ, LiuK, WangL, XuC, LiX, XueY, XiaM, JiQ, LuJ, XuM, ZhangQ . A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of indica-japonica hybrids in rice Proc Natl Acad Sci USA, 2008,105(32):11436-11441. [本文引用: 1]
HouJ, CaoC, RuanY, DengY, LiuY, ZhangK, TanL, ZhuZ, CaiH, LiuF, SunH, GuP, SunC, FuY . ESA1 is involved in embryo sac abortion in interspecific hybrid progeny of rice Plant Physiol, 2019,180(1):356-366. [本文引用: 1]
KoideY, OginoA, YoshikawaT, KitashimaY, SaitoN, KanaokaY, OnishiK, YoshitakeY, TsukiyamaT, SaitoH, TeraishiM, YamagataY, UemuraA, TakagiH, HayashiY, AbeT, FukutaY, OkumotoY, KanazawaA . Lineage- specific gene acquisition or loss is involved in interspecific hybrid sterility in rice Proc Natl Acad Sci USA, 2018,115(9):E1955-E1962. [本文引用: 2]
KuboT, TakashiT, AshikariM, YoshimuraA, KurataN . Two tightly linked genes at the hsa1 locus cause both F1 and F2 hybrid sterility in rice Mol Plant, 2016,9(2):221-232. [本文引用: 1]
LongY, ZhaoL, NiuB, SuJ, WuH, ChenY, ZhangQ, GuoJ, ZhuangC, MeiM, XiaJ, WangL, WuH, LiuYG . Hybrid male sterility in rice controlled by interaction between divergent alleles of two adjacent genes Proc Natl Acad Sci USA, 2008,105(48):18871-18876. [本文引用: 3]
MizutaY, HarushimaY, KurataN . Rice pollen hybrid incompatibility caused by reciprocal gene loss of duplicated genes Proc Natl Acad Sci USA, 2010,107(47):20417-20422. [本文引用: 4]
NguyenGN, YamagataY, ShigematsuY, WatanabeM, MiyazakiY, DoiK, TashiroK, KuharaS, KanamoriH, WuJ, MatsumotoT, YasuiH, YoshimuraA . Duplication and loss of function of genes encoding RNA polymerase III subunit C4 causes hybrid incompatibility in rice G3, 2017, 7(8): 2565-2575. [本文引用: 2]
XieY, XuP, HuangJ, MaS, XieX, TaoD, ChenL, LiuYG . Interspecific hybrid sterility in rice is mediated by OgTPR1 at the S1 locus encoding a peptidase-like protein Mol Plant, 2017,10(8):1137-1140. [本文引用: 4]
YamagataY, YamamotoE, AyaK, WinKT, DoiK, Sobrizal, ItoT, KanamoriH, WuJ, MatsumotoT, MatsuokaM, AshikariM, YoshimuraA . Mitochondrial gene in the nuclear genome induces reproductive barrier in rice Proc Natl Acad Sci USA, 2010,107(4):1494-1499. [本文引用: 2]
YangJ, ZhaoX, ChengK, DuH, OuyangY, ChenJ, QiuS, HuangJ, JiangY, JiangL, DingJ, WangJ, XuC, LiX, ZhangQ . A killer-protector system regulates both hybrid sterility and segregation distortion in rice Science, 2012,337(6100):1336-1340. [本文引用: 2]
YuY, ZhaoZ, ShiY, TianH, LiuL, BianX, XuY, ZhengX, GanL, ShenY, WangC, YuX, WangC, ZhangX, GuoX, WangJ, IkehashiH, JiangL, WanJ . Hybrid sterility in rice (Oryza sativa L.) involves the tetratricopeptide repeat domain containing protein Genetics, 2016,203(3):1439-1451. [本文引用: 1]
XieY, ShenR, ChenL, LiuYG . Molecular mechanisms of hybrid sterility in rice Sci China Life Sci, 2019 62(6), 737-743. [本文引用: 1]
KitamuraE . Genetics studies on sterility observed in hybrids between distantly related varieties of rice (Oryza sativa L.) Bull Chgoku Agri Exp Sta Ser A, 1962,8:141-205. [本文引用: 2]
ZhangGQ, LuYG . Development of the indica-compatibilitic japonica lines for the breeding of super-rice J Shenyang Agric Univ, 2007,38(5):676-680. [本文引用: 1]
MaRR, WangXY, LuYF, ZhouHC, CaiKF, LiXN, ZhangZY . Breeding and application of late japonica CMS line Yongjing 2A and its late indica-japonica hybrid rice combinations Hybrid Rice, 2010,25(S1):185-189. [本文引用: 1]
XieY, NiuB, LongY, LiG, TangJ, ZhangY, RenD, LiuYG, ChenL . Suppression or knockout of SaF/SaM overcomes the Sa-mediated hybrid male sterility in rice J Integr Plant Biol, 2017,59(9):669-679. [本文引用: 1]
ChangZ, ChenZ, WangN, XieG, LuJ, YanW, ZhouJ, TangX, DengXW . Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene Proc Natl Acad Sci USA, 2016,113(49):14145-14150. [本文引用: 2]
XiaY, TangN, HuY, LiD, LiS, BuX, YuM, QiS, YangY, ZhuH, CaoC, LiP, YuanL, CaoM . A method for mechanized hybrid rice seed production using female sterile rice Rice, 2019,12(1):39. [本文引用: 1]
LiQ, ZhangD, ChenM, LiangW, WeiJ, QiY, YuanZ . Development of japonica photo-sensitive genic male sterile rice lines by editing Carbon Starved Anther using CRISPR/Cas9 J Genet Genomics, 2016,43(6):415-419. [本文引用: 1]
ZhouH, HeM, LiJ, ChenL, HuangZ, ZhengS, ZhuL, NiE, JiangD, ZhaoB, ZhuangC . Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediated TMS5 editing system Sci Rep, 2016,6:37395. [本文引用: 1]
GaravitoA, GuyotR, LozanoJ, GavoryF, SamainS, PanaudO, TohmeJ, GhesquièreA, LorieuxM . A genetic model for the female sterility barrier between asian and african cultivated rice species Genetics, 2010,185(4):1425-1440. [本文引用: 1]
 ,11.
,11. 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT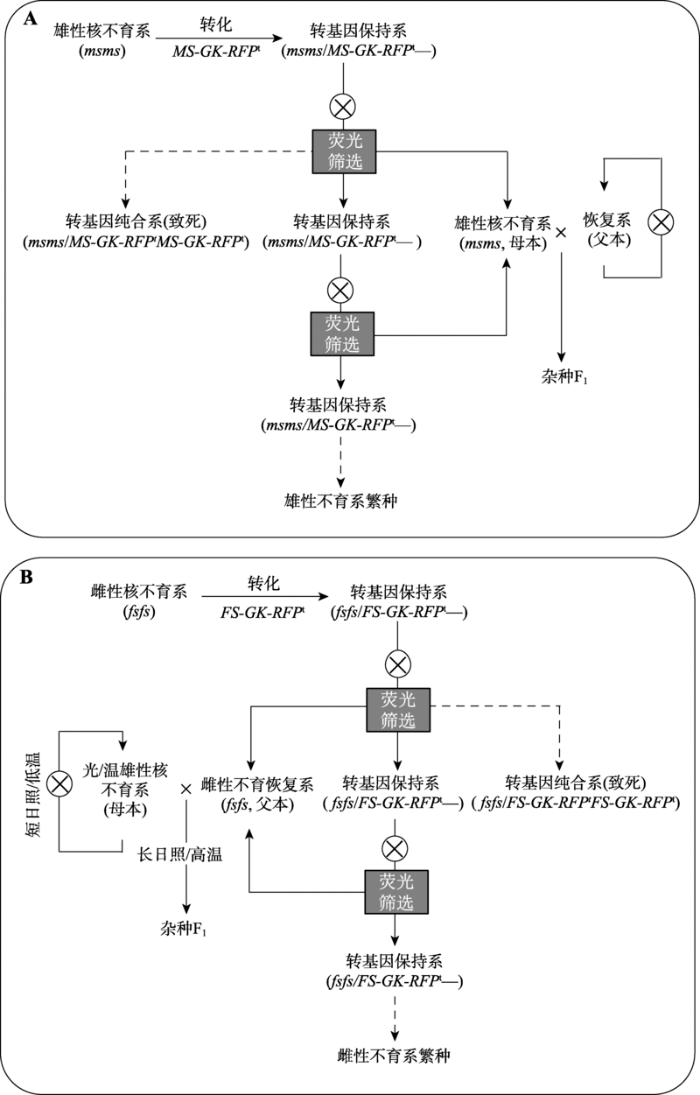 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}