Epigenetics in the genesis and development of cancers
Jingwen Zhang, Qian Xu, Guoliang Li,College of Informatics, National Key Laboratory of Crop Genetic Improvement, Agricultural Bioinformatics Key Laboratory of Hubei Province, Hubei Engineering Technology Research Center of Agricultural Big Data, Huazhong Agricultural University, Wuhan 430070, China
Abstract Cancer is a complex disease caused by the malignant cellular proliferation and metastasis. Elucidating its pathogenic mechanism is one of the major challenges that we face currently. Epigenetic mechanisms are essential for maintaining specific patterns of gene expression and normal development and growth of living individuals. Disorders of epigenetic markers, such as histone modification, DNA/RNA methylation, and changes in the three-dimensional conformation of chromatin, can interfere with gene expression to some extent, and result in cancers. This review provides a brief overview of epigenetics, focusing on their association with the genesis of cancers, and we look forward to the application of epigenetics in cancer clinical diagnosis and treatment. Keywords:cancers;epigenetics;DNA/RNA methylation;histone modification;nucleosome positioning;non-coding RNA;chromatin three-dimensional structure
PDF (653KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张競文, 续倩, 李国亮. 癌症发生发展中的表观遗传学研究[J]. 遗传, 2019, 41(7): 567-581 doi:10.16288/j.yczz.19-077 Jingwen Zhang, Qian Xu, Guoliang Li. Epigenetics in the genesis and development of cancers[J]. Hereditas(Beijing), 2019, 41(7): 567-581 doi:10.16288/j.yczz.19-077
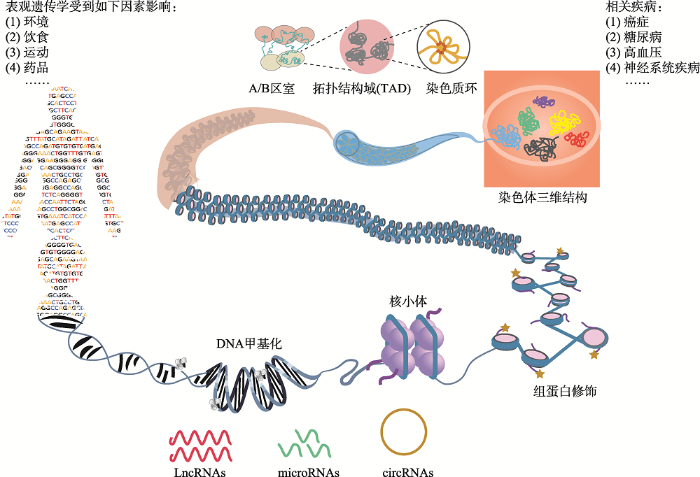
表观遗传学研究DNA序列之外可遗传的变化特征。生命个体的所有细胞具有基本相同的DNA,但不同的器官和组织具有不同的功能,而且随着多次细胞分裂仍能维持其特定的细胞身份,这在很大程度上被认为是由表观遗传信息介导的。表观遗传信息诸如组蛋白修饰、DNA甲基化和染色质三维构象等受基因组序列、环境暴露、饮食习惯和其他随机因素等的共同调控,代表着基因和环境等因素之间的相互作用。表观遗传信息对维持基因特定的表达模式和生命个体的正常发育至关重要,表观遗传修饰紊乱可能改变基因表达,导致疾病发生。癌症是由细胞恶性增殖和扩散引发的常见疾病,已经成为最主要的疾病死亡原因。深入研究并解析其致病机制是当今人类社会面临的重大挑战之一。近年来,癌症基因组图谱(the cancer genome atlas, TCGA)揭示了许多癌症的基因组突变,包括单碱基突变、拷贝数变异和染色体重排等。另外,许多研究也表明表观遗传修饰在癌症中发生了改变。表观遗传修饰的动态变化性和可逆性也为癌症的治疗提供了思路。因此,表观遗传学研究在癌症中的应用将有助于人们解析病人基因组、环境暴露和生活方式等因素之间的关系,进而更好地预防、诊治癌症,促进人类健康。本文将从DNA/RNA甲基化、组蛋白修饰、核小体定位、非编码RNA和染色质三维结构等方面概述表观遗传学的研究内容,并聚焦其与癌症发生发展之间的关联,展望表观遗传学在癌症临床诊治中的应用。
非编码RNA (non-coding RNA, ncRNA)被认为是基因组中不编码蛋白质的“暗物质”,占人类基因组的80%左右。DNA元件百科全书(encyclopedia of DNA elements, ENCODE)和表观基因组学图谱(roadmap epigenomics project)等项目通过生化方法和生物信息学手段发现了数百万个非编码调控区域。功能性非编码RNA主要包括miRNA、lncRNA、circRNA和siRNA等。
LiZ, WengH, SuR, WengX, ZuoZ, LiC, HuangH, NachtergaeleS, DongL, HuC, QinX, TangL, WangY, HongGM, HuangH, WangX, ChenP, GurbuxaniS, ArnovitzS, LiY, LiS, StrongJ, NeillyMB, LarsonRA, JiangX, ZhangP, JinJ, HeC, ChenJ . FTO plays an oncogenic role in acute myeloid leukemia as a N 6-methyladenosine RNA demethylase Cancer Cell, 2016,31(1):127-141. [本文引用: 2]
CuiQ, ShiH, YeP, LiL, QuQ, SunG, SunG, LuZ, HuangY, YangCG, D. RiggsAD, HeC, ShiY . m 6A RNA methylation regulates the self-renewal and tumorigenesis of glioblastoma stem cells Cell Rep, 2017,18(11):2622-2634. URL [本文引用: 3]
ZhangS, ZhaoBS, ZhouA, LinK, ZhengS, LuZ, ChenY, SulmanEP, XieK, BöglerO, MajumderS, HeC, HuangS . m 6A demethylase ALKBH5 maintains tumorigenicity of glioblastoma stem-like cells by sustaining FOXM1 expression and cell proliferation program Cancer Cell, 2017, 31(4): 591-606. e6. URL [本文引用: 2]
ZhangC, SamantaD, LuH, BullenJW, ZhangH, ChenI, HeX, SemenzaGL . Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5- mediated m 6A-demethylation of NANOG mRNA Proc Natl Acad Sci USA, 2016,113(14):E2047-E2056. URL [本文引用: 2]
ChenK, ChenZ, WuD, ZhangL, LinX, SuJ, RodriguezB, XiY, XiaZ, ChenX, ShiX, WangQ, LiW . Broad H3K4me3 is associated with increased transcription elongation and enhancer activity at tumor-suppressor genes Nat Genet, 2015,47(10):1149-1157. [本文引用: 2]
LaiWKM, PughBF . Understanding nucleosome dynamics and their links to gene expression and DNA replication Nat Rev Mol Cell Bio, 2017,18(9):548-562. [本文引用: 1]
NairSS, KumarR . Chromatin remodeling in cancer: a gateway to regulate gene transcription Mol Oncol, 2012,6(6):611-619. URL [本文引用: 2]
KadochC, CrabtreeGR . Mammalian SWI/SNF chromatin remodeling complexes and cancer: mechanistic insights gained from human genomics Science Advances, 2015,1(5):e1500447. URL [本文引用: 2]
AndersonDM, ChangCL, MakarewichCA, NelsonBR, McAnallyJR, KasaragodP, SheltonJM, LiouJ, Bassel-DubyR, OlsonEN . A micropeptide encoded by a putative long noncoding RNA regulates muscle performance Cell, 2015,160(4):595-606. URL [本文引用: 1]
NelsonB, MakarewichCA, AndersonDM, WindersBR, TroupesCD, WuF, ReeseAL, McAnallyJR, ChenX, KavalaliET, CannonSC, HouserSR, Bassel-DubyR, OlsonEN . A peptide encoded by a transcript annotated as long noncoding RNA enhances SERCA activity in muscle Science, 2016, 271-275. [本文引用: 1]
LegniniI, Di TimoteoG, RossiF, MorlandoM, BrigantiF, SthandierO, FaticaA, SantiniT, AndronacheA, WadeM, LaneveP, RajewskyN, BozzoniI . Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis Mol Cell, 2017, 66(1): 22-37. e9. URL [本文引用: 1]
HuangJZ, ChenM, ChenD, GaoXC, ZhuS, HuangH, HuM, ZhuH, YanGR . A peptide encoded by a putative lncRNA HOXB-AS3 suppresses colon cancer growth Mol Cell, 2017, 68(1): 171-3184. e6. URL [本文引用: 2]
YangY, GaoX, ZhangM, YanS, SunC, XiaoF, HuangN, YangX, ZhaoK, ZhouH, HuangS, XieB, ZhangN . Novel role of FBXW7 circular RNA in repressing glioma tumorigenesis J Natl Cancer Inst Monographs, 2018,110(3). [本文引用: 1]
ZhangM, HuangN, YangX, LuoJ, YanS, XiaoF, ChenW, GaoX, ZhaoK, ZhouH, LiZ, MingL, XieB, ZhangN . A novel protein encoded by the circular form of the SHPRH gene suppresses glioma tumorigenesis Oncogene, 2018,37(13):1805-1814. [本文引用: 2]
LiR, LiuY, HouY, GanJ, WuP, LiC . 3D genome and its disorganization in diseases Cell Biol Toxicol, 2018,34(5):351-365. [本文引用: 1]
DixonJ, GorkinD, RenB . Chromatin domains: the unit of chromosome organization Mol Cell, 2016,62(5):668-680. URL [本文引用: 1]
RandoOJ . Intergenerational transfer of epigenetic information in sperm CSH Perspect Med, 2016,6(5):a022988. [本文引用: 1]
BiswasS, RaoCM . Epigenetics in cancer: fundamentals and beyond Pharmacol Ther, 2017,173:118-134. URL [本文引用: 3]
TimpW, BravoHC, McDonaldOG, GogginsM, UmbrichtC, ZeigerM, FeinbergAP, IrizarryRA . Large hypomethylated blocks as a universal defining epigenetic alteration in human solid tumors Genome Med, 2014,6(8):61. URL [本文引用: 2]
ChalitchagornK, ShuangshotiS, HourpaiN, KongruttanachokN, TangkijvanichP, Thong-ngamD, VoravudN, SriuranpongV, MutiranguraA . Distinctive pattern of LINE-1 methylation level in normal tissues and the association with carcinogenesis Oncogene, 2004,23(54):8841-8846. [本文引用: 1]
VisvanathanA, PatilV, AroraA, HegdeAS, ArivazhaganA, SantoshV, SomasundaramK . Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance Oncogene, 2017,37(4):522-533. [本文引用: 2]
QiuZ, WangJ, WuY . The landscape of histone modification in cancer metastasis. 2018. [本文引用: 3]
WangP, LinC, SmithER, GuoH, SandersonBW, WuM, GogolM, AlexanderT, SeidelC, WiedemannLM, GeK, KrumlaufR, ShilatifardA . Global analysis of H3K4 methylation defines mll family member targets and points to a role for MLL1-Mediated H3K4 methylation in the regulation of transcriptional initiation by RNA polymerase Ⅱ Mol Cell Biol, 2009,29(22):6074-6085. URL [本文引用: 1]
SunJM, GuoCC, WangCQ, CaoK, LiuH, HanWC, ZhengMJ . Expression of BRG1 in colorectal cancer: correlation with prognosis and MMP-2 expression World Chin J Digestol, 2016,24(35):4691-4699. [本文引用: 1]
TordellaL, KhanS, HohmeyerA, BanitoA, KlotzS, RaguzS, MartinN, DhamarlingamG, CarrollT, González MeljemJM, DeswalS, Martínez-BarberaJP, García- EscuderoR, ZuberJ, ZenderL, GilJ . SWI/SNF regulates a transcriptional program that induces senescence to prevent liver cancer Genes Dev, 2016,30(19):2187-2198. URL [本文引用: 1]
LiX, YaoX, WangY, HuF, WangF, JiangL, LiuY, WangD, SunG, ZhaoY . MLH1 promoter methylation frequency in colorectal cancer patients and related clinicopathological and molecular features PLoS One, 2013,8(3):e59064. URL [本文引用: 1]
LanF, ShiY . Histone H3.3 and cancer: a potential reader connection Proc Natl Acad Sci USA, 2014,112(22):6814-6819. [本文引用: 1]
LinCP, HeL . Noncoding RNAs in cancer development Annu Rev Cancer Biol, 2017,1(1):163-184. URL [本文引用: 1]
Ivo D'UrsoP, Fernando D’UrsoO, Damiano GianfredaC, MezzollaV, StorelliC, MarsiglianteS . MiR-15b and miR-21 as Circulating biomarkers for diagnosis of glioma Curr Genomics, 2015,16(5):304-311. URL [本文引用: 1]
PardoOE, CastellanoL, E MunroC, HuY, MauriF, KrellJ, LaraR, G PinhoF, ChoudhuryT, FramptonAE, PellegrinoL, PshezhetskiyD, WangY, WaxmanJ, SecklMJ, StebbingJ . MIR-515-5p controls cancer cell migration through MARK4 regulation EMBO Rep, 2016,17(4):570-584. URL [本文引用: 1]
MeiQ, LiX, ZhangK, WuZ, LiX, MengY, GuoM, LuoG, FuX, HanW . Genetic and methylation-induced loss of miR-181a2/181b2 within chr9q33.3 facilitates tumor growth of cervical cancer through PIK3R3/Akt/FoxO signaling pathway Clin Cancer Res, 2017,23(2):575-586. URL [本文引用: 1]
ZhaoJL, HuangF, HeF, GaoCC, LiangSQ, MaPF, DongGY, HanH, QinHY . Forced activation of notch in macrophages represses tumor growth by upregulating miR-125a and disabling Tumor-Associated macrophages Cancer Res, 2016,76(6):1403-1415. URL [本文引用: 1]
ZhangY, HuJ, WangH, CuiJ, GaoS, HoffmanAR, LiW . CRISPR Cas9-guided chromatin immunoprecipitation identifies miR483 as an epigenetic modulator of IGF2 imprinting in tumors Oncotarget, 2016,8(21):34177-34190. [本文引用: 1]
YangL, MaY, HanW, LiW, CuiL, ZhaoX, TianY, ZhouZ, WangW, WangH . Proteinase-activated receptor 2 promotes cancer cell migration through RNA methylation- mediated repression of MiR-125b J Biol Chem, 2015,290(44):26627-26637. URL [本文引用: 1]
GuoJ, LvJ, LiuM, TangH . MiR-346 Up-regulates argonaute 2 (AGO2) protein expression to augment the activity of other (MiRNAs) and contributes to cervical cancer cell malignancy J Biol Chem, 2015,290(51):30342-30350. URL [本文引用: 1]
Celià-TerrassaT, LiuDD, ChoudhuryA, HangX, WeiY, ZamalloaJ, Alfaro-AcoR, ChakrabartiR, JiangYZ, KohBI, SmithHA, DecosteC, LiJJ, ZhaoN, KangY . Normal and cancerous mammary stem cells evade interferon- induced constraint through the MIR-199a-LCOR axis Nat Cell Biol, 2017,19(6):711-723. [本文引用: 1]
YangMH, YuJ, ChenN, WangXY, LiuXY, WangS, DingYQ . Elevated MicroRNA-31 expression regulates colorectal cancer progression by repressing its target gene SATB2 PLoS One, 2013,8(12):e85353. URL [本文引用: 1]
FangJ, MorsalinS, RaoVN, ShyamP. RE . Decoding of non-coding DNA and non-coding RNA: pri-micro RNA- encoded novel peptides regulate migration of cancer cells J Pharm Sci Pharmacol2017,3(1):23-27. URL [本文引用: 1]
ShiJ, LiY, FangXD . The mechanism and clinical significance of long noncoding RNA-mediated gene expression via nuclear architecture Hereditas (Beijing), 2017,39(3):189-199. [本文引用: 1]
YangH, LiuP, ZhangJ, PengX, LuZ, YuS, MengY, TongWM, ChenJ . Long noncoding RNA MIR31HG exhibits oncogenic property in pancreatic ductal adenocarcinoma and is negatively regulated by miR-193b Oncogene, 2015,35(28):3647-3657. [本文引用: 1]
GeY, YanX, JinY, YangX, YuX, ZhouL, HanS, YuanQ, YangM . MiRNA-192 and miRNA-204 directly suppress lncRNA HOTTIP and Interrupt GLS1-mediated glutamineolysis in hepatocellular carcinoma PLoS Genet, 2015,11(12):e1005726. URL [本文引用: 1]
GuarnerioJ, BezziM, JeongJC, PaffenholzSV, BerryK, NaldiniMM, Lo-CocoF, TayY, BeckAH, PandolfiPP . Oncogenic role of Fusion-circRNAs derived from cancer- associated chromosomal translocations Cell, 2016,165(2):289-302. URL [本文引用: 1]
YangQ, DuWW, WuN, YangW, AwanFM, FangL, MaJ, LiX, ZengY, YangZ, DongJ, KhorshidiA, YangBB . A circular RNA promotes tumorigenesis by inducing c-myc nuclear translocation Cell Death Differ, 2017,24(9):1609-1620. [本文引用: 1]
LiF, ZhangL, LiW, DengJ, ZhengJ, AnM, LuJ, ZhouY . Circular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathway Oncotarget, 2015,6(8):6001-6013. [本文引用: 1]
WanL, ZhangL, FanK, ChengZX, SunQC, WangJJ . Circular RNA-ITCH suppresses lung cancer proliferation via inhibiting the Wnt/β-catenin pathway Biomed Res Int, 2016,2016 1579490. [本文引用: 1]
HuangG, ZhuH, ShiY, WuW, CaiH, ChenX . Cir-ITCH plays an inhibitory role in colorectal cancer by regulating the Wnt/β-catenin pathway PLoS One, 2015,10(6):e0131225. URL [本文引用: 1]
GuoW, ZhangJ, ZhangD, CaoS, LiG, ZhangS, WangZ, WenP, YangH, ShiX, PanJ, YeH . Polymorphisms and expression pattern of circular RNA circ-ITCH contributes to the carcinogenesis of hepatocellular carcinoma Oncotarget, 2017,8(29):48169-48177. [本文引用: 1]
LiCL, NieH, WangM, SuLP, LiJF, YuYY, YanM, QuQL, ZhuZG, LiuBY . MicroRNA-155 is downregulated in gastric cancer cells and involved in cell metastasis Oncol Rep, 2012,27(6):1960-1966. [本文引用: 1]
BarutcuAR, LajoieBR, MccordRP, TyeCE, HongD, MessierTL, BrowneG, van WijnenAJ, LianJB, SteinJL, DekkerJ, ImbalzanoAN, SteinGS . Chromatin interaction analysis reveals changes in small chromosome and telomere clustering between epithelial and breast cancer cells Genome Biol, 2015,16(1):214. URL [本文引用: 2]
TaberlayPC, Achinger-kaweckaJ, LunAT, BuskeFA, SabirK, GouldCM, ZotenkoE, BertSA, GilesKA, BauerDC, SmythGK, StirzakerC . Three-dimensional disorganization of the cancer genome occurs coincident with long-range genetic and epigenetic alterations Genome Res, 2016,26(6):719-731. URL [本文引用: 3]
WuP, LiT, LiR, JiaL, ZhuP, LiuY, ChenQ, TangD, YuY, LiC . 3D genome of multiple myeloma reveals spatial genome disorganization associated with copy number variations Nat Commun, 2017,8(1):1937. URL [本文引用: 2]
 ,华中农业大学信息学院,作物遗传改良国家重点实验室,农业生物信息学湖北省重点实验室,湖北省农业大数据工程技术研究中心,武汉 430070
,华中农业大学信息学院,作物遗传改良国家重点实验室,农业生物信息学湖北省重点实验室,湖北省农业大数据工程技术研究中心,武汉 430070
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}