何俊1, Fernando B. Lopes2, 吴晓林,1,2,31. 湖南农业大学动物科技学院,长沙410128 2. 美国威斯康星大学动物科学系,威斯康星州麦迪逊市53706 3. 美国纽勤公司生物信息与生物统计部,内布拉斯加州林肯市68504
Methods and applications of animal genomic mating
Jun He1, Fernando B. Lopes2, Xiao-Lin Wu,1,2,31. CollegeofAnimalScienceand Technology, HunanAgricultural University, Changsha 410128, China 2. Department of Animal Science, University of Wisconsin, Madison WI 53706, USA 3. Biostatisticsand Bioinformatics, NeogenGeneSeek, LincolnNE 68504, USA
Supported by Key Project of Scientific Research Plan of Hunan Province.2018NK2081 Key Project of Scientific Research Plan of Changsha city.kq1801014 Hundred-Talent Project of Hunan Province and Hunan Innovation Center of Animal Safety Production
作者简介 About authors 何俊,博士,副教授,研究方向:动物遗传育种E-mail:hejun@hunau.edu.cn。
Abstract Genomic selection (GS) is a powerful tool which can be used to estimate the breeding value of individual animals by using the molecular markers of the animal’s entire genome. GS improves the accuracy and intensity of selection, reduces the interval of generation, and realizes the effects of early accuracy selection contributing to a significant evolution in animal breeding. In the past decade, GS was successfully applied in the genetic improvement of dairy animals with improved selection accuracy and genetic gain of breeding animals. However, GS focuses on the genetic gain of target traits while it ignores the genetic relationship between mating pairs such that it ignores long term genetic merits such as an increase in inbreeding coefficient of offspring population, a decrease of genetic diversity and the homozygous presentation of harmful genes. In 2016, genomic mating (GM) was proposed as a sustainable genetic selection method using genomic information of the breeding candidate individuals to optimize selection and mating with resultant control of the growth rate of population inbreeding coefficient and achieving long-term and sustainable genetic progress. Therefore, GM is more suitable for modern animal breeding than GS, especially for the genetic improvement of indigenous species. In this review, we summarize the basic concepts, methods, and applications of GM, and then present examples comparing the effects of six simulated mating schemes. This review serves as a valuable reference for the applications of animal breeding methods. Keywords:genomic selection;genomic mating;optimal contribution selection
PDF (339KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 何俊, Fernando B. Lopes, 吴晓林. 动物基因组选配方法与应用[J]. 遗传, 2019, 41(6): 486-493 doi:10.16288/j.yczz.19-053 Jun He, Fernando B. Lopes, Xiao-Lin Wu. Methods and applications of animal genomic mating[J]. Hereditas(Beijing), 2019, 41(6): 486-493 doi:10.16288/j.yczz.19-053
许多研究表明,维持中长期选择的遗传进展就必须控制群体近交程度的快速提升和遗传变异的丧失[16,27,43,44]。因为不管是表型选择还是GS,都会因为近交程度的提升和遗传变异的降低,长期的选择反应都会快速降低。特别是GS导致快速遗传进展和高近交增量的相关,使近交控制在长期选择中变得越来越重要。Pryce等[29]通过分析选配后代的预期遗传进展和近交以及隐性有害等位基因纯合子的变化情况,比较了配种方案,利用系谱、基因组和ROH信息的3种控制近交程度的配种策略的效果。结果表明配种方案中使用基因组信息是一种有效的方法,比系谱信息更能降低后代期望近交系数而对遗传进展影响最小。在获得相同遗传进展的情况下,相比于利用系谱信息,利用基因组信息可使后代的期望近交程度降低几乎2倍。Liu等[45]利用系谱和基因组信息,使用随机模拟来比较最小共祖选配(minimum-coancestry mating, MC)和最小化祖先间遗传贡献的协方差(minimizing the covariance between ancestral genetic contributions, MCAC)两种配种策略在5种育种方案中实现的近交增量和遗传进展,同时模拟了离散世代的随机交配作为参照。模拟了2000个QTL控制的单性状,对动物进行截断选择,选择之前,对所有后备个体进行表型测定。选择依据是岭回归模型预测的GEBV。研究结果表明,利用基因组信息时,MC和MCAC选配策略估计的近交增量比利用系谱信息的低6%~22%,而不影响遗传进展,且两种策略的近交增量和遗传进展差别不大。然而,与随机交配方案相比,在利用基因组信息的MC和MCAC选配策略下,估计的近交增量比随机交配方案低28%~44%,遗传进展提高14%。因此,利用基因组信息进行选配,可以有效控制近交增量,同时保持较高的遗传进展。
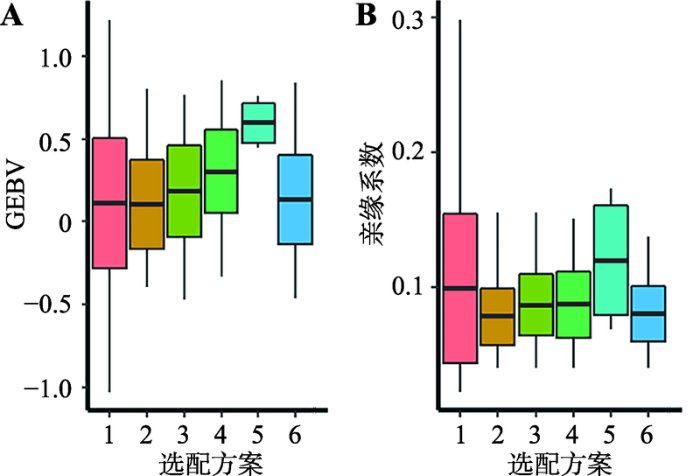
A:基因组育种值;B:亲缘系数。1~6代表6种不同的选配方案。 Fig. 1Comparison of genomic estimation breeding value and kinship coefficient between parents and offspring in six mating schemes
Table1 表1 表1 模拟群体基因组信息的参数 Table1 The parameters of genomic information in simulation population
GianolaD, RosaGJ . One hundred years of statistical developments in animal breeding Annu Rev AnimBiosci, 2014,3:19-56. [本文引用: 1]
Henderson CR . Best linear unbiased estimation and prediction under a selection model Biometrics, 1975,31(2):423-447. URL [本文引用: 1]
VergaraOD, ElzoMA, Cerón-Mu?ozMF . Genetic parameters and genetic trends for age at first calving and calving interval in an angus- blancoorejinegro-zebu multibreed cattle population in colombia LivestSci, 2009,126(1):318-322. [本文引用: 1]
AguilarI, MisztalI, JohnsonDL, LegarraA, TsurutaS, LawlorTJ . Hot topic: a unified approach to utilize phenotypic, full pedigree, and genomic information for genetic evaluation of Holstein final score J dairy sci, 2010,93(2):743-752. URL [本文引用: 1]
CaetanoSL, SavegnagoRP, BoligonAA, RamosSB, ChudTCS, L?boRB, MunariDP . Estimates of genetic parameters for carcass, growth and reproductive traits in nellore cattle LivestSci, 2013,155(1):1-7. [本文引用: 1]
LopesFB, da SilvaMC, MagnaboscoCU, GoncalvesNarcisoMG, SainzRD . Selection indices and multivariate analysis show similar results in the evaluation of growth and carcass traits in beef cattle PLoS One, 2016,11(1):e0147180. URL [本文引用: 1]
MeuwissenTH, HayesBJ, GoddardME . Prediction of total genetic value using genome-wide dense marker maps Genetics, 2001,157(4):1819-1829. [本文引用: 1]
MeuwissenT. Genomic selection: the future of marker assisted selection and animal breeding. In: Proceedings of Electronic forum on biotechnology in food and agriculture. MAS: a fast track to increase genetic gain in plant and animal breeding, Session II: MAS in animals. FAO, Conference 10. University of Turin via L. da Vinci 44, Grugliasco (TO) Italy. 2003, 54-59. [本文引用: 1]
WiggansGR, ColeJB, HubbardSM, SonstegardTS . Genomic selection in dairy cattle: The USDA experience Annu Rev AnimBiosci, 2017,5:309-327. [本文引用: 1]
MeuwissenT, HayesB, GoddardM . Accelerating improvement of livestock with genomic selection Annu Rev AnimBiosci, 2013,1:221-237. [本文引用: 1]
de los CamposG, HickeyJM, Pong-WongR, DaetwylerHD, CalusMP . Whole-genome regression and prediction methods applied to plant and animal breeding Genetics, 2013,193(2):327-345. URL [本文引用: 1]
LourencoDA, TsurutaS, FragomeniBO, MasudaY, AguilarI, LegarraA, BertrandJK, AmenTS, WangL, MoserDW, MisztalI . Genetic evaluation using single-step genomic best linear unbiased predictor in american angus, J AnimSci, 2015,93(6):2653-2662. [本文引用: 1]
MeuwissenTH . Maximizing the response of selection with a predetermined rate of inbreeding J AnimSci, 1997,75(4):934-940. [本文引用: 5]
GrundyB, VillanuevaB, WoolliamsJA . Dynamic selection procedures for constrained inbreeding and their consequences for pedigree development Genet Res, 1998,72(2):159-168. URL [本文引用: 4]
GrundyB, VillanuevaB, WoolliamsJA . Dynamic selection for maximizing response with constrained inbreeding in schemes with overlapping generations AnimSci, 2000,70(3), 373-382. [本文引用: 1]
WoolliamsJA, ThompsonR. A theory of genetic contributions. In: Proceedings of 5th World Congress of Genetics Applied to Livestock Production, University of Guelph, Guelph, Ontario, Canada. 1994,25:127-134. [本文引用: 1]
Avenda?oS, WoolliamsJA, VillanuevaB . Mendelian sampling terms as a selective advantage in optimum breeding schemes with restrictions on the rate of inbreeding Genet Res, 2004,83(1), 55-64. URL [本文引用: 1]
HenryonM, BergP, S?rensenAC . Animal-breeding schemes using genomic information need breeding plans designed to maximise long-term genetic gains LivestSci, 2014,166:38-47. [本文引用: 1]
SonessonAK, WoolliamsJA, MeuwissenTHE . Genomic selection requires genomic control of inbreeding Genet SelEvol, 2012,44:27. [本文引用: 4]
AkdemirD, SánchezJI . Efficient breeding by genomic mating Front Genet, 2016,7:210. [本文引用: 8]
PryceJE, HayesBJ, GoddardME . Novel strategies to minimize progeny inbreeding while maximizing genetic gain using genomic information J Dairy Sci, 2012,95(1):377-388. URL [本文引用: 5]
JansenGB, WiltonJW . Selecting mating pairs with linear programming techniques J Dairy Sci, 1985,68(5):1302-1305. URL [本文引用: 1]
SchierenbeckS, PimentelECG, TietzeM, K?rteJ, ReentsR, ReinhardtF, SimianerH, K?nigS . Controlling inbreeding and maximizing genetic gain using semi-definite programming with pedigree-based and genomic relationships J Dairy Sci, 2011,94(12):6143-6152. URL [本文引用: 1]
HillWG, WeirBS . Variation in actual relationship as a consequence of mendelian sampling and linkage Genet Res, 2011,93(1):47-64. URL [本文引用: 1]
ClarkSA, KinghornBP, HickeyJM, van der WerfJH . The effect of genomic information on optimal contribution selection in livestock breeding programs Genet SelEvol, 2013,45:44. doi: 10.1186/1297-9686-45-44. [本文引用: 2]
WeigelKA, LinSW . Use of computerized mate selection programs to control inbreeding of holstein and jersey cattle in the next generation J Dairy Sci, 2000,83(4):822-828. URL [本文引用: 2]
MeuwissenTHE. GENCONT: An operational tool for controlling inbreeding in selection and conservation schemes. In: Proceedings of 7th World Congress on Genetics Applied to Livestock Production Montpellier, France. 2002, CD-ROM communication no 28-20. [本文引用: 2]
WoolliamsJA, BergP, DagnachewBS, MeuwissenTH . Genetic contributions and their optimization J Anim Breed Genet, 2015,132(2):89-99. URL [本文引用: 2]
Pong-WongR, WoolliamsJA . Optimisation of contribution of candidate parents to maximise genetic gain and restricting inbreeding using semidefinite programming Genet SelEvol, 2007,39(1):3-25. [本文引用: 1]
AhlinderJ, MullinTJ, YamashitaM . Using semidefinite programming to optimize unequal deployment of genotypes to a clonal seed orchard Tree Genet Genomes, 2014,10(1):27-34. URL [本文引用: 1]
CarvalheiroR, QueirozSAD, KinghornB . Optimum contribution selection using differential evolution R Bras Zootec, 2010,39(7):1429-1436. URL [本文引用: 1]
StornR, PriceK . Differential evolution a simple and efficient heuristic for global optimization over continuous spaces J Global Optim, 1997,11(4):341-359. URL [本文引用: 1]
KinghornBP . An algorithm for efficient constrained mate selection Genet SelEvol, 2011,43(1):4. [本文引用: 1]
MullinTJ, BelottiP . Using branch-and-bound algorithms to optimize selection of a fixed-size breeding population under a relatedness constraint Tree Genet Genomes, 2016,12(1):4. URL [本文引用: 1]
GoddardM . Genomic selection: prediction of accuracy and maximisation of long term response Genetica, 2009,136(2):245-257. URL [本文引用: 2]
LiuH, HenryonM, S?rensenAC . Mating strategies with genomic information reduce rates of inbreeding in animal breeding schemes without compromising genetic gain Animal, 2017,11(4):547-555. URL [本文引用: 2]
LiB, LealSM . Methods for detecting associations with rarevariants for common diseases: application to analysis of sequence data Am J Hum Genet, 2008,83(3):311-321. URL [本文引用: 1]
SargolzaeiM, SchenkelFS . QMSim: a large-scale genome simulator for livestock Bioinformatics, 2009,25(5):680-681. URL [本文引用: 1]
BritoFV, NetoJB, SargolzaeiM, CobuciJA, SchenkelFS . Accuracy of genomic selection in simulated populations mimicking the extent of linkage disequilibrium in beef cattle BMC Genet, 2011,12(1):80. [本文引用: 1]
WellmannR . Optimum contribution selection for animal breeding and conservation: the R package optiSel BMC Bioinformatics, 2019,20(1):25. [本文引用: 1]
 ,1,2,31.
,1,2,31. 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}