Endocytic recycling pathways and the regulatory mechanisms
Long Lin, Anbing Shi1. Department of Biochemistry and Molecular Biology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China 2. Institute for Brain Research, Huazhong University of Science and Technology, Wuhan 430030, China 3. Key Laboratory of Neurological Disease of National Education Ministry, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China
Abstract Endocytic transport is imperative for the exchange of information between cells and the external environment. Specifically, the process of endocytic transport comprises precise regulation of uptake and sorting of extracellular macromolecules, phospholipids, and membrane proteins. In the endocytic transport system, the recycling pathways are responsible for delivering membrane proteins and phospholipids back to the plasma membrane. Thus, endocytic recycling plays critical roles in various biological processes, including nutrient absorption, cell polarity establishment, cell migration, cell division, synaptic plasticity, immune response, and growth factor receptor regulation. There are two essential types of recycling pathways in eukaryotic cells, recycling of clathrin-dependent endocytic cargos (CDE recycling) and recycling of clathrin-independent endocytic cargos (CIE recycling). The transferrin receptor TfR and the low-density lipoprotein receptor LDLR, which have essential physiological roles in vivo, are representative membrane proteins of the CDE recycling transport. In recent years, various membrane proteins governed by CIE recycling transport have been identified, including IL2 receptor α-subunit, major histocompatibility complex MHC Class I, and glucose transporter GLUT4. Therefore, the investigation of the regulatory mechanisms of CIE recycling has drawn notable attention in the field. Moreover, CIE recycling research presents fundamental significance in cell biology, which also provides scientific evidence and potential therapeutic clues for the diagnosis and treatment strategies of diseases such as type Ⅱ diabetes and cancer. Compared with the CDE recycling, the study on CIE recycling started later, and there is much to be learned of its regulatory mechanisms. To this end, this review summarizes the features of endocytic recycling pathways, focuses on the molecular basis of CIE recycling regulation and elaborates on the latest progress and newly developed research model systems in the field of CIE recycling. Keywords:endocytic recycling;clathrin-dependent endocytosis;clathrin-independent endocytosis;recycling endosome;Rab;Arf;phosphoinositide;F-actin;Caenorhabditis elegans
PDF (678KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 林珑, 史岸冰. 细胞内吞循环运输通路及其分子调控机制[J]. 遗传, 2019, 41(6): 451-468 doi:10.16288/j.yczz.19-124 Long Lin, Anbing Shi. Endocytic recycling pathways and the regulatory mechanisms[J]. Hereditas(Beijing), 2019, 41(6): 451-468 doi:10.16288/j.yczz.19-124
内吞运输过程受一系列Rab家族小G蛋白(Rab small GTPase)调控,在囊泡运动和囊泡栓系中发挥关键作用。Rab家族小G蛋白是高度保守的小型单体GTP酶,也是Ras小G蛋白超家族中最大的亚家族。Rab通过向特定的膜室(membrane compartment)表面募集其特异的效应因子(effector)来调控内膜系统的结构和功能[37]。类似其他Ras超家族小G蛋白的活性调控方式,Rab作为“分子开关”在GDP和GTP结合态之间循环。Rab自身具有较低的核苷酸交换和水解速率,需要其他蛋白因子来协助调节GTP-GDP循环,其中鸟嘌呤核苷酸交换因子(GEFs)和GTP酶激活蛋白(GAPs)分别催化GDP到GTP的交换和GTP的水解反应[38]。Rab的效应因子通过选择性结合Rab (GTP)活性形式,形成功能性的分子集合体,可以作为内膜区室的标志物参与调控囊泡形成、靶向定位和融合[37]。这种以Rab为中心的蛋白复合物的GTP-GDP结合态往复转换在空间和时间上受到精细调节,以实现内膜识别的动态性和货物运输的多样性[39],因而Rab活性在时空上的精确调节对细胞内吞运输至关重要。
活性态RAB-5(GTP)招募效应因子LET-413以促进DENN-4/GEF对RAB-10的激活,而活性态RAB-10(GTP)可以通过其效应因子TBC-2/GAP关闭上游RAB-5活性。RAB-10 (GTP)招募效应因子CNT-1/Arf-GAP对ARF-6活性进行负调控,下调内体上PI(4,5)P2水平。RAB-10的另外一个效应因子EHBP-1调控循环内体上微丝(F-actin)的成束捆绑,同时定位于循环内体的PTRN-1促进微丝的聚合,共同促进循环内体的膜出芽。ARF-6结合蛋白SAC-1以负反馈方式对ARF-6活性进行抑制以维持内体上PI(4,5)P2稳态。RME-1在RAB-10下游发挥功能,结合循环内体中的磷酸肌醇PI(4,5)P2,参与管状膜运输结构塑形。定位于循环内体的SNAREs和exocyst复合体进一步介导膜融合。黄色标示蛋白因子的早期内体定位,绿色标示蛋白因子的循环内体定位。 Fig. 2Regulation of recycling transport of clathrin-independent cargos
在秀丽线虫中的研究发现,小G蛋白RAC1 (Rac1 small GTPase)同源物CED-10与CED-12/ ELMO、CED-5/DOCK180、CED-2/CrkII协同作用,促进细胞骨架重组,吞噬凋亡细胞[91]。CED-12和CED-5形成CED-10二元GEF,促进CED-10(GDP)转化为活性CED-10(GTP)[92]。CED-2被认为是一种衔接因子,能够与CED-5结合将蛋白复合物与某些凋亡受体如MOM-5/Frizzled或Integrins连接起来[93,94]。此外,哺乳动物细胞中RAC1在内体上能够转化为GTP携载状态,依赖ARF6介导的循环运输将其定位于迁移细胞的前沿[95]。后续研究发现活性态CED-10能够定位在早期内体和循环内体,参与调控上皮肠细胞中的内吞循环运输。对其分子作用机制的研究发现CED-10能够募集Rab-GAP蛋白TBC-2,对早期内体关键调控因子RAB-5活性进行负调控以促进循环运输[96]。
CullenPJ, SteinbergF . To degrade or not to degrade: mechanisms and significance of endocytic recycling Nat Rev Mol Cell Biol, 2018,19(11):679-696. [本文引用: 1]
PrestonJE, Joan AbbottN, BegleyDJ . Transcytosis of macromolecules at the Blood-Brain barrier Adv Pharmacol, 2014,71:147-163. URL [本文引用: 1]
AntonescuCN, McgrawTE, KlipA . Reciprocal regulation of endocytosis and metabolism Cold Spring Harb Perspect Biol, 2014,6(7):a016964. URL [本文引用: 1]
MellmanI, YardenY . Endocytosis and cancer CSH Perspect Biol, 2013,5(12):a016949. [本文引用: 1]
IrannejadR, von ZastrowM . GPCR signaling along the endocytic pathway Curr Opin Cell Biol, 2014,27:109-116. URL [本文引用: 1]
SteinmanRM, MellmanIS, MullerWA, CohnZA . Endocytosis and the recycling of plasma membrane J Cell Biol, 1983,96(1):1-27. URL [本文引用: 2]
ConnerSD, SchmidSL . Regulated portals of entry into the cell Nature, 2003,422(6927):37-44. [本文引用: 2]
DohertyGJ , McMahon HT. Mechanisms of endocytosis Annu Rev Biochem, 2009,78:857-902. URL [本文引用: 2]
BonifacinoJS, RojasR . Retrograde transport from endosomes to the trans-Golgi network Nat Rev Mol Cell Bio, 2006,7(8):568-579. [本文引用: 1]
GruenbergJ, StenmarkH . The biogenesis of multivesicular endosomes Nat Rev Mol Cell Bio, 2004,5(4):317-323. [本文引用: 1]
CarpentierJL, GordenP, AndersonRG, GoldsteinJL, BrownMS, CohenS, OrciL . Co-localization of 125I-epidermal growth factor and ferritin-low density lipoprotein in coated pits: a quantitative electron microscopic study in normal and mutant human fibroblasts J Cell Biol, 1982,95(1):73-77. URL [本文引用: 1]
NeutraMR, CiechanoverA, OwenLS, LodishHF . Intracellular transport of transferrin- and asialoorosomucoid- colloidal gold conjugates to lysosomes after receptor- mediated endocytosis Chin J Histochem Cytochem, 1985,33(11):1134-1144. URL [本文引用: 1]
RobinsonMS . Forty years of Clathrin-coated vesicles Traffic, 2015,16(12):1210-1238. URL [本文引用: 1]
KirchhausenT . Adaptors for clathrin-mediated traffic Annu Rev Cell Dev Bi, 1999,15:705-732. URL [本文引用: 1]
McMahonHT, BoucrotE . Molecular mechanism and physiological functions of clathrin-mediated endocytosis Nat Rev Mol Cell Biol, 2011,12(8):517-533. [本文引用: 2]
KamikuraDM, CooperJA . Clathrin interaction and subcellular localization of Ce-DAB-1, an adaptor for protein secretion in Caenorhabditis elegans Traffic, 2006,7(3):324-336. URL [本文引用: 1]
CocucciE, AguetF, BoulantS, KirchhausenT . The first five seconds in the life of a clathrin-coated pit Cell, 2012,150(3):495-507. URL [本文引用: 1]
HenneWM, BoucrotE, MeineckeM, EvergrenE, VallisY, MittalR, McMahonHT . FCHo proteins are nucleators of clathrin-mediated endocytosis Science, 2010,328(5983):1281-1284. URL [本文引用: 1]
WoodwardMP, RothTF . Coated vesicles: characterization, selective dissociation, and reassembly Proc Natl Acad Sci USA, 1978,75(9):4394-4398. URL [本文引用: 1]
QualmannB, KochD, KesselsMM . Let’s go bananas: revisiting the endocytic BAR code EMBO J, 2011,30(17):3501-3515. URL [本文引用: 1]
AguetF, AntonescuCN, MettlenM, SchmidSL, DanuserG . Advances in analysis of low Signal-to-Noise images link dynamin and AP2 to the functions of an endocytic checkpoint Dev Cell, 2013,26(3):279-291. URL [本文引用: 1]
SchmidSL, FrolovVA . Dynamin: functional design of a membrane fission catalyst Annu Rev Cell Dev Biol, 2011,27:79-105. URL [本文引用: 2]
UngewickellE, UngewickellH, HolsteinSE, LindnerR, PrasadK, BarouchW, MartinB, GreeneLE, EisenbergE . Role of auxilin in uncoating clathrin-coated vesicles Nature, 1995,378(6557):632-635. [本文引用: 1]
DonaldsonJG, JacksonCL . ARF family G proteins and their regulators: roles in membrane transport, development and disease Nat Rev Mol Cell Biol, 2011,12(6):362-375. URL [本文引用: 1]
BucciC, PartonRG, MatherIH, StunnenbergH, SimonsK, HoflackB, ZerialM . The small GTPase rab5 functions as a regulatory factor in the early endocytic pathway Cell, 1992,70(5):715-728. URL [本文引用: 1]
WoodmanPG . Biogenesis of the sorting endosome: the role of Rab5 Traffic, 2000,1(9):695-701. URL [本文引用: 1]
CarneyDS, DaviesBA, HorazdovskyBF . Vps9 domain- containing proteins: activators of Rab5 GTPases from yeast to neurons Trends Cell Biol, 2005,16(1):27-35. [本文引用: 1]
ChotardL, SkorobogataO, SylvainMA, ShrivastavaS, RocheleauCE . TBC-2 Is required for embryonic yolk protein storage and larval survival during L1 diapause in Caenorhabditis elegans PLoS One, 2011,5(12):e15662. [本文引用: 1]
SatoM, SatoK, FonarevP, HuangCJ, LiouW, GrantBD . Caenorhabditis elegans RME-6 is a novel regulator of RAB-5 at the clathrin-coated pit Nat Cell Biol, 2005,7(6):559-569. [本文引用: 3]
SannSB, CraneMM, LuH, JinY . Rabx-5 regulates RAB-5 early endosomal compartments and synaptic vesicles in C. elegans PLoS One, 2012,7(6):e37930. URL [本文引用: 1]
PoteryaevD, DattaS, AckemaK, ZerialM, SpangA . Identification of the switch in Early-to-Late endosome transition Cell, 2010,141(3):497-508. URL [本文引用: 1]
DoiM, MinematsuH, KubotaY, NishiwakiK, MiyamotoM . The novel Rac effector RIN-1 regulates neuronal cell migration and axon pathfinding in C. elegans Development, 2013,140(16):3435-3444. URL [本文引用: 1]
FunderburkSF, WangQJ, YueZ . The Beclin 1-VPS34 complex-at the crossroads of autophagy and beyond Trends Cell Biol, 2010,20(6):355-362. URL [本文引用: 1]
LindmoK, StenmarkH . Regulation of membrane traffic by phosphoinositide 3-kinases J Cell Sci, 2006,119(4):605-614. URL [本文引用: 1]
DangH, LiZ, SkolnikEY, FaresH . Disease-related myotubularins function in endocytic traffic in Caenorhabditis elegans Mol Biol Cell, 2004,15(1):189-196. URL [本文引用: 1]
RuckA, AttonitoJ, GarcesKT, NúnezL, PalmisanoNJ, RubelZ, BaiZ, NguyenKCQ, SunL, GrantBD, HallDH, MeléndezA . The Atg6/Vps30/Beclin 1 ortholog BEC-1 mediates endocytic retrograde transport in addition to autophagy in C. elegans Autophagy, 2011,7(4):386-400. URL [本文引用: 1]
XueY, FaresH, GrantB, LiZ, RoseAM, ClarkSG, SkolnikEY . Genetic analysis of the myotubularin family of phosphatases in Caenorhabditis elegans J Biol Chem, 2003,278(36):34380-34386. URL [本文引用: 1]
ChenCC, SchweinsbergPJ, VashistS, MareinissDP, LambieEJ, GrantBD . RAB-10 is required for endocytic recycling in the Caenorhabditis elegans intestine Mol Biol Cell, 2006,17(3):1286-1297. URL [本文引用: 10]
ShiA, ChenCC, BanerjeeR, GlodowskiD, AudhyaA, RongoC, GrantBD . EHBP-1 functions with RAB-10 during endocytic recycling in Caenorhabditis elegans Mol Biol Cell, 2010,21(16):2930-2943. URL [本文引用: 9]
AngAL, TaguchiT, FrancisS, F?lschH, MurrellsLJ, PypaertM, WarrenG, MellmanI . Recycling endosomes can serve as intermediates during transport from the golgi to the plasma membrane of MDCK cells J Cell Biol, 2004,167(3):531-543. URL [本文引用: 1]
BabbeyCM, AhktarN, WangE, ChenCC, GrantBD, DunnKW . Rab10 regulates membrane transport through early endosomes of polarized Madin-Darby canine kidney cells Mol Biol Cell, 2006,17(7):3156-3175. URL [本文引用: 1]
SanoH, EguezL, TeruelMN, FukudaM, ChuangTD, ChavezJA, LienhardGE, McGrawTE . Rab10, a target of the AS160 Rab GAP, is required for insulin- stimulated translocation of GLUT4 to the adipocyte plasma membrane Cell Metab, 2007,5(4):293-303. URL [本文引用: 1]
GuilhermeA, SorianoNA, BoseS, HolikJ, BoseA, PomerleauDP, FurcinittiP, LeszykJ, CorveraS, CzechMP . EHD2 and the novel EH domain binding protein EHBP1 couple endocytosis to the actin cytoskeleton J Biol Chem, 2004,279(11):10593-10605. URL [本文引用: 2]
GuilhermeA, SorianoNA, FurcinittiPS, CzechMP . Role of EHD1 and EHBP1 in perinuclear sorting and insulin-regulated GLUT4 recycling in 3T3-L1 adipocytes J Biol Chem, 2004,279(38):40062-40075. URL [本文引用: 2]
SchuckS, GerlMJ, AngA, ManninenA, KellerP, MellmanI, SimonsK . Rab10 is involved in basolateral transport in polarized Madin-Darby canine kidney cells Traffic, 2007,8(1):47-60. URL [本文引用: 1]
HutagalungAH, NovickPJ . Role of Rab GTPases in membrane traffic and cell physiology Physiol Rev, 2011,91(1):119-149. URL [本文引用: 1]
LiuH, WangS, HangW, GaoJ, ZhangW, ChengZ, YangC, HeJ, ZhouJ, ChenJ, ShiA . LET-413/Erbin acts as a RAB-5 effector to promote RAB-10 activation during endocytic recycling J Cell Biol, 2018,217(1):299-314. URL [本文引用: 1]
GrantB, ZhangY, PaupardMC, LinSX, HallDH, HirshD . Evidence that RME-1, a conserved C. elegans EH-domain protein, functions in endocytic recycling Nat Cell Biol, 2001,3(6):573-579. [本文引用: 3]
ApodacaG, KatzLA, MostovKE . Receptor-mediated transcytosis of IgA in MDCK cells is via apical recycling endosomes J Cell Biol, 1994,125(1):67-86. URL [本文引用: 1]
LinSX, GrantB, HirshD, MaxfieldFR . Rme-1 regulates the distribution and function of the endocytic recycling compartment in mammalian cells Nat Cell Biol, 2001,3(6):567-572. [本文引用: 2]
SantoliniE, SalciniAE, KayBK, YamabhaiM, Di FiorePP . The EH network Exp Cell Res, 1999,253(1):186-209. URL [本文引用: 1]
de BeerT, HoofnagleAN, EnmonJL, BowersRC, YamabhaiM, KayBK, OverduinM . Molecular mechanism of NPF recognition by EH domains Nat Struct Biol, 2000,7(11):1018-1022. URL [本文引用: 1]
BraunA, PinyolR, DahlhausR, KochD, FonarevP, GrantBD, KesselsMM, QualmannB . EHD proteins associate with syndapin I and II and such interactions play a crucial role in endosomal recycling Mol Biol Cell, 2005,16(8):3642-3658. URL [本文引用: 1]
ShiA, PantS, BalklavaZ, ChenCC, FigueroaV, GrantBD . A novel requirement for C. elegans Alix/ALX-1 in RME-1-mediated membrane transport Curr Biol, 2007,17(22):1913-1924. URL [本文引用: 3]
LeeDW, ZhaoX, ScarsellettaS, SchweinsbergPJ, EisenbergE, GrantBD, GreeneLE . ATP binding regulates oligomerization and endosome association of RME-1 family proteins J Biol Chem, 2005,280(17):17213-17220. URL [本文引用: 1]
CaplanS, NaslavskyN, HartnellLM, LodgeR, PolishchukRS, DonaldsonJG, BonifacinoJS . A tubular EHD1-containing compartment involved in the recycling of major histocompatibility complex class I molecules to the plasma membrane EMBO J, 2002,21(11):2557-2567. URL [本文引用: 1]
DaumkeO, LundmarkR, VallisY, MartensS, ButlerPJ , McMahon HT. Architectural and mechanistic insights into an EHD ATPase involved in membrane remodelling Nature, 2007,449(7164):923-927. [本文引用: 4]
van DamEM, StoorvogelW . Dynamin-dependent transferrin receptor recycling by endosome-derived clathrin-coated vesicles Mol Biol Cell, 2002,13(1):169-182. URL [本文引用: 1]
HiroiT, SomeyaA, ThompsonW, MossJ, VaughanM . GEP100/BRAG2: activator of ADP-ribosylation factor 6 for regulation of cell adhesion and actin cytoskeleton via E-cadherin and alpha-catenin Proc Natl Acad Sci USA, 2006,103(28):10672-10677. URL [本文引用: 1]
BoshansRL, SzantoS, van AelstL, D'Souza-SchoreyC . ADP-ribosylation factor 6 regulates actin cytoskeleton remodeling in coordination with Rac1 and RhoA Mol Cell Biol, 2000,20(10):3685-3694. URL [本文引用: 1]
PalaciosF, PriceL, SchweitzerJ, CollardJG, D'Souza-SchoreyC . An essential role for ARF6-regulated membrane traffic in adherens junction turnover and epithelial cell migration EMBO J, 2001,20(17):4973-4986. URL [本文引用: 1]
YinHL, JanmeyPA . Phosphoinositide regulation of the actin cytoskeleton Annu Rev Physiol, 2002,65:761-789. [本文引用: 1]
Kinchen JM. RavichandranKS . Journey to the grave: signaling events regulating removal of apoptotic cells J Cell Sci, 2007,120(Pt 13):2143-2149. URL [本文引用: 1]
BrugneraE, HaneyL, GrimsleyC, LuM, WalkSF, Tosello-TrampontAC, MacaraIG, MadhaniH, FinkGR, RavichandranKS . Unconventional Rac-GEF activity is mediated through the Dock180-ELMO complex Nat Cell Biol, 2002,4(8):574-582. [本文引用: 1]
ReddienPW, HorvitzHR . CED-2/CrkII and CED-10/Rac control phagocytosis and cell migration in Caenorhabditis elegans Nat Cell Biol, 2000,2(3):131-136. [本文引用: 1]
CabelloJ, NeukommLJ, GünesdoganU, BurkartK, CharetteSJ, LochnitG, HengartnerMO, SchnabelR . The wnt pathway controls cell death engulfment, spindle orientation, and migration through CED-10/Rac PLoS Biol, 2010,8(2):e1000297. URL [本文引用: 1]
RadhakrishnaH, Al-AwarO, KhachikianZ, DonaldsonJG . ARF6 requirement for Rac ruffling suggests a role for membrane trafficking in cortical actin rearrangements J Cell Sci, 1999,112(Pt 6):855-866. [本文引用: 1]
SunL, LiuO, DesaiJ, KarbassiF, SylvainMA, ShiA, ZhouZ, RocheleauCE, GrantBD . CED-10/Rac1 regulates endocytic recycling through the RAB-5 GAP TBC-2 PLoS Genet, 2012,8(7):e1002785. URL [本文引用: 1]
TerBushDR, MauriceT, RothD, NovickP . The exocyst is a multiprotein complex required for exocytosis in saccharomyces cerevisiae EMBO J, 1996,15(23):6483-94. URL [本文引用: 2]
F?lschH, PypaertM, MadayS, PelletierL, MellmanI . The AP-1A and AP-1B clathrin adaptor complexes define biochemically and functionally distinct membrane domains J Cell Biol, 2003,163(2):351-362. URL [本文引用: 1]
WuS, MehtaSQ, PichaudF, BellenHJ, QuiochoFA . Sec15 interacts with Rab11 via a novel domain and affects Rab11 localization in vivo Nat Struct Mol Biol, 2005,12(10):879-885. [本文引用: 1]
OztanA, SilvisM, WeiszOA, BradburyNA, HsuSC, GoldenringJR, YeamanC, ApodacaG . Exocyst requirement for endocytic traffic directed toward the apical and basolateral poles of polarized MDCK cells Mol Biol Cell, 2007,18(10):3978-3992. URL [本文引用: 1]
ChenS, LiL, LiJ, LiuB, ZhuX, ZhengL, ZhangR, XuT . SEC-10 and RAB-10 coordinate basolateral recycling of clathrin-independent cargo through endosomal tubules in Caenorhabditis elegans Proc Natl Acad Sci USA, 2014,111(43):15432-15437. URL [本文引用: 3]
MallardF, TangBL, GalliT, TenzaD, Saint-PolA, YueX, AntonyC, HongW, GoudB, JohannesL . Early/recycling endosomes-to-TGN transport involves two SNARE complexes and a Rab6 isoform J Cell Biol, 2002,156(4):653-664. URL [本文引用: 1]
BonifacinoJS, HierroA . Transport according to GARP: receiving retrograde cargo at the trans-Golgi network Trends Cell Biol, 2011,21(3):159-167. URL [本文引用: 1]
SchindlerC, ChenY, PuJ, GuoX, BonifacinoJS . EARP is a multisubunit tethering complex involved in endocytic recycling Nat Cell Biol, 2015,17(5):639-650. [本文引用: 1]
Di PaoloG, De CamilliP . Phosphoinositides in cell regulation and membrane dynamics Nature, 2006,443(7112):651-657. [本文引用: 2]
GleasonAM, NguyenKC, HallDH, GrantBD . Syndapin/SDPN-1 is required for endocytic recycling and endosomal actin association in the C. elegans intestine Mol Biol Cell, 2016. [本文引用: 1]
DelevoyeC, HeiligensteinX, RipollL, Gilles-MarsensF, DennisMK, LinaresRA, DermanL, GokhaleA, MorelE, FaundezV, MarksMS, RaposoG . BLOC-1 brings together the actin and microtubule cytoskeletons to generate recycling endosomes Curr Biol, 2015,26(1):1-13. [本文引用: 1]
SchaferDA, D'Souza-SchoreyC, CooperJA . Actin assembly at membranes controlled by ARF6 Traffic, 2000,1(11):892-903. [本文引用: 2]
HendershottMC, ValeRD . Regulation of microtubule minus-end dynamics by CAMSAPs and patronin Proc Natl Acad Sci USA, 2014,111(16):5860-5865. URL [本文引用: 1]
RichardsonCE, SpilkerKA, CuevaJG, PerrinoJ, GoodmanMB, ShenK . PTRN-1, a microtubule minus end-binding CAMSAP homolog, promotes microtubule function in Caenorhabditis elegans neurons eLife, 2014,3:e01498. URL [本文引用: 1]
ChuangM, GoncharovA, WangS, OegemaK, JinY, ChisholmAD . The microtubule minus-end-binding protein patronin/PTRN-1 is required for axon regeneration in C. elegans Cell Rep, 2014,9(3):874-883. URL [本文引用: 2]
FergusonS, RaimondiA, ParadiseS, ShenH, MesakiK, FergusonA, DestaingO, KoG, TakasakiJ, CremonaO, O' TooleE, De CamilliP . Coordinated actions of actin and BAR proteins upstream of dynamin at endocytic Clathrin-Coated pits Dev Cell, 2009,17(6):811-822. URL [本文引用: 1]
MitsunariT, NakatsuF, ShiodaN, Love PE GrinbergA, BonifacinoJS, OhnoH . Clathrin adaptor AP-2 is essential for early embryonal development Mol Cell Biol, 2005,25(21):9318-9323. URL [本文引用: 1]
RoyleSJ . The cellular functions of clathrin Cell Mol Life Sci, 2006,63(16):1823-1832. URL [本文引用: 1]
ShiM, ZhangY, ZhouGQ . The critical roles of TBC proteins in human diseases Hereditas (Beijing), 2018,40(1):12-21. [本文引用: 1]
AdjeiIM, SharmaB, LabhasetwarV . Nanoparticles: cellular uptake and cytotoxicity,in nanomaterial: impacts on cell biology and medicine. D.G.apco and Y.Chen Editors 2014, Springer Netherlands:Dordrecht. 73-91. [本文引用: 1]
EspositoG, Ana ClaraF, VerstrekenP . Synaptic vesicle trafficking and parkinson's disease Dev Neurobiol, 2012,72(1):134-144. URL [本文引用: 1]
JiangS, LiY, ZhangX, BuG, XuH, ZhangYW . Trafficking regulation of proteins in Alzheimer’s disease Mol Neurodegener, 2014,9(1):6. URL [本文引用: 1]
GoldenringJR . A central role for vesicle trafficking in epithelial neoplasia: intracellular highways to carcinogenesis Nat Rev Cancer, 2013,13:813-820. [本文引用: 1]
TangFC, XueYF . RNA interference and gene silencing Hereditas (Beijing), 2001,23(2):167-266. URLMagsci [本文引用: 1] 双链RNA介导的遗传干涉的机制是1998年发现的。它通过双链RNA的介导特异性地降解相应序列的mRNA,从而导致转录后水平的基因沉默。到目前为止在真菌、拟南芥、线虫、锥虫、水螅、涡虫、果蝇、斑马鱼、小鼠等真核生物中都发现存在这一基因沉默机制。目前的研究表明,RNA干涉与植物中的共抑制(cosuppression)、真菌中的基因压制(quelling)很可能具有共同的基本分子机制。这也说明,很可能在进化的很早期阶段,生物就获得了这种机制。RNA干涉对于抵抗病毒入侵、抑制转座子活动等具有重要作用,对于生物体的发育和基因调控可能也有重要作用。<br>Abstract:The mechanism of a new phenomenon――genetic interference directed by double-stranded RNA was first discovered in 1998 by Andrew Fire and Craig Mello. It degrades mRNA specifically and potently through the mediation of corresponding double-stranded RNA and leads to post-transcriptional gene silencing. Consequently RNA interference was observed is eukaryotic organisms including fungi, Arabidopsis,C. elegans, trypanosomes, hydra, planaria, Drosophila, zabrafish, and mouse. More and more evidence supports that RNAi,co-suppression in plants, and quelling in fungi shares the same basic molecular mechanism. It indicates that this mechanism was acquired during early evolution. RNAi plays important roles in resistance to virus invasion, and inhibition of transportable elements. And it is very likely that RNAi is also important during the normal development and regulation of gene expression. 汤富酬, 薛友纺 . RNA干涉与基因沉默 遗传, 2001,23(2):167-266. URLMagsci [本文引用: 1] 双链RNA介导的遗传干涉的机制是1998年发现的。它通过双链RNA的介导特异性地降解相应序列的mRNA,从而导致转录后水平的基因沉默。到目前为止在真菌、拟南芥、线虫、锥虫、水螅、涡虫、果蝇、斑马鱼、小鼠等真核生物中都发现存在这一基因沉默机制。目前的研究表明,RNA干涉与植物中的共抑制(cosuppression)、真菌中的基因压制(quelling)很可能具有共同的基本分子机制。这也说明,很可能在进化的很早期阶段,生物就获得了这种机制。RNA干涉对于抵抗病毒入侵、抑制转座子活动等具有重要作用,对于生物体的发育和基因调控可能也有重要作用。<br>Abstract:The mechanism of a new phenomenon――genetic interference directed by double-stranded RNA was first discovered in 1998 by Andrew Fire and Craig Mello. It degrades mRNA specifically and potently through the mediation of corresponding double-stranded RNA and leads to post-transcriptional gene silencing. Consequently RNA interference was observed is eukaryotic organisms including fungi, Arabidopsis,C. elegans, trypanosomes, hydra, planaria, Drosophila, zabrafish, and mouse. More and more evidence supports that RNAi,co-suppression in plants, and quelling in fungi shares the same basic molecular mechanism. It indicates that this mechanism was acquired during early evolution. RNAi plays important roles in resistance to virus invasion, and inhibition of transportable elements. And it is very likely that RNAi is also important during the normal development and regulation of gene expression.
MaXY, ZhaoYL, JiaFX, SongYK, TseYC . Utilization of Caenorhabditis elegans in laboratory teaching of genetics Hereditas (Beijing), 2017,39(8):763-768. [本文引用: 1]
ZhangXM, GaoJ, ChenCH, TuHJ . Progress in the mechanisms of neural modulation of innate immunity in Caenorhabditis elegans Hereditas (Beijing), 2018,40(12):1066-1074. [本文引用: 1]
ShiA, SunL, BanerjeeR, TobinM, ZhangY, GrantBD . Regulation of endosomal clathrin and retromer- mediated endosome to golgi retrograde transport by the J-domain protein RME-8 EMBO J, 2009,28(21):3290-3302. URL [本文引用: 1]


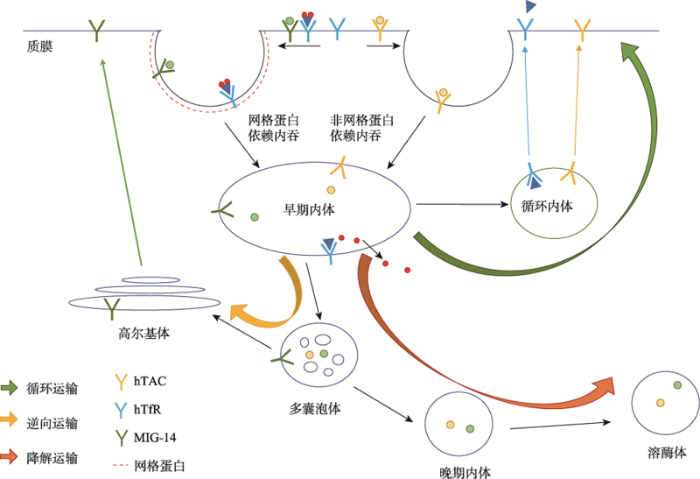 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT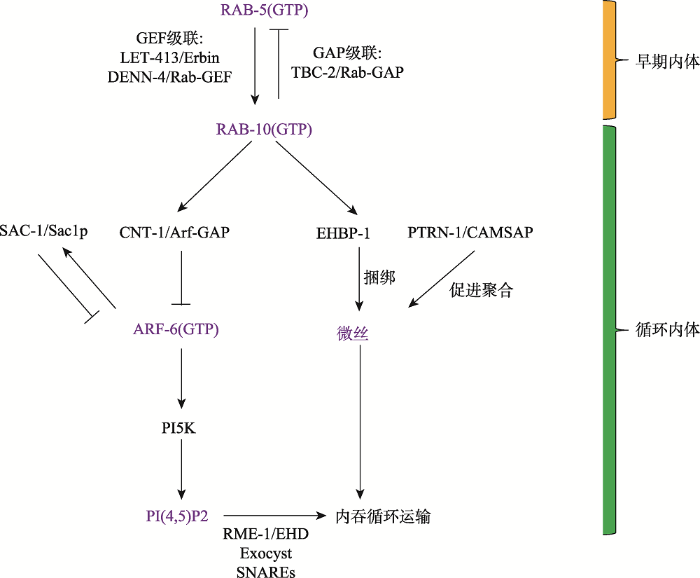 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT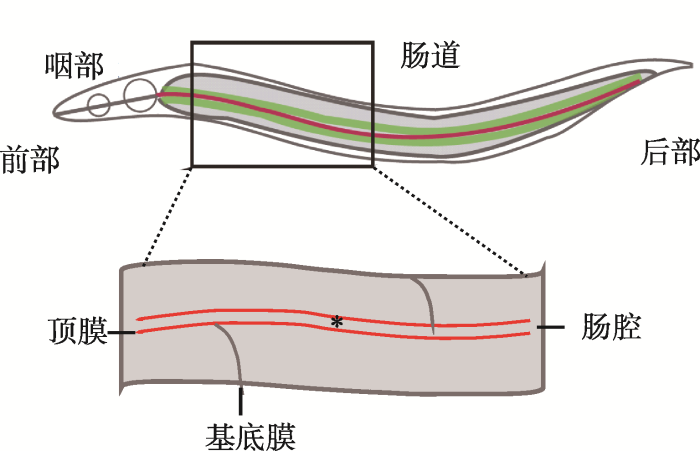 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}