Mechanism on Drought Tolerance Enhanced by Exogenous Trehalose in C4-PEPC Rice
Jiaxin Li1,2, Xia Li,1,2,3,*, Yinfeng Xie11College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China 2Nanjing Branch of China National Center for Rice Improvement/Jiangsu High Quality Rice Engineering Technology Research Center/Institute of Food Crops, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China 3Collaborative Innovation Center for Modern Production Technology of Grain Crops of Jiangsu Province, Agricultural College, Yangzhou University, Yangzhou 225009, China
Abstract In order to reveal the mechanism of trehalose (Tre) involved in the drought tolerance of transgenic rice (Oryza sativa) (PC) expressing the C4-PEPC gene of maize (Zea mays), we analyzed the physiological and biochemistry characteristics of PC and wild-type rice (WT) treated hydroponically with Tre and 12% (m/v) PEG alone or in combination. The results showed that Tre treatment promoted the growth of PC and WT seedlings, and alleviated the inhibition of plant growth caused by drought stress (DS), with the effect being more significant for PC. Compared with DS treatment, Tre+DS treatment maintained functional leaves a higher relative water content, photochemical efficiency and antioxidant enzyme activity. Under DS, compared with WT, PC showed significant increase in the content of Tre and sucrose and decrease in the content of glucose, and up-regulated expression of genes associated with Tre metabolism and SnRK1s after Tre application. Tre application also improved PC the ABA synthesis, and expression of genes related to signal transduction and drought response, and maintained PC a relatively stable photosynthetic capacity, thus possibly conferring stronger drought tolerance of PC. Keywords:C4-type PEPC transgenic rice;drought stress;phosphate phosphoenolpyruvate carboxylase;trehalose;rice (Oryza sativa)
PDF (7620KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李佳馨, 李霞, 谢寅峰. 外源海藻糖增强高表达转玉米C4型PEPC水稻耐旱性的机制. 植物学报, 2021, 56(3): 296-314 doi:10.11983/CBB20168 Li Jiaxin, Li Xia, Xie Yinfeng. Mechanism on Drought Tolerance Enhanced by Exogenous Trehalose in C4-PEPC Rice. Chinese Bulletin of Botany, 2021, 56(3): 296-314 doi:10.11983/CBB20168
ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020)。值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016)。前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017)。海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题。本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据。
采用TaKaRa Mini BEST Plant RNA Extraction Kit (TaKaRa, Cat No.9769)试剂盒提取总RNA。采用TaKaRa Prime Script RT Master Mix Perfect Real Time (TaKaRa, Cat No.RR036A)试剂盒进行反转录。采用TB Green Premix Ex Taq Tli RNaseH Plus (TaKaRa, Cat No.RR82LR)试剂盒进行Real Time PCR。用Applied Biosystems Step One实时PCR系统(Applied Biosystems, Foster City, CA, USA)进行分析。qRT-PCR反应程序为95°C10分钟; 94°C30秒, 55°C40秒, 60°C1分钟, 共32次循环。重复3次。采用Primer 3设计引物, 以水稻组成型表达的Act基因为内参, 引物序列见表1。
Table 1 表1 表1qRT-PCR的基因和引物 Table 1Genes and primers for qRT-PCR
模拟干旱处理2小时后C4-PEPC基因的相对表达量(A)、磷酸烯醇式丙酮酸羧化酶(PEPC)活性(B)和Osppc2a基因的相对表达量(C)。缩写同图1。图中数值代表来自3个生物学重复的平均值±标准差(n=6)。不同小写字母的柱值表示差异显著(P<0.05) (Student-Neuman-Keuls)。 Figure 2Trehalose treatments increase PEPC transcription and translation in rice leaves under drought stress
C4-PEPC expression (A), phosphoenolpyruvate carboxylase (PEPC) activity (B) and Osppc2a expression (C) after 2 hours of simulated drought treatment. Abbreviations are the same as those given in Figure 1. Values represent means±SD (n=6) from three biological replicates. Bars labeled with different lowercase letters indicate significant differences (P<0.05) (Student-Neuman-Keuls).
模拟干旱处理2小时后PC和WT的最大光化学效率(Fv/Fm) (A)、有效光化学效率(Fv′/Fm′) (B)、实际光化学效率(ΦPSII) (C)、光化学淬灭系数(qP) (D)和非光化学淬灭系数(NPQ) (E)。缩写同图1。图中数值代表来自3个生物学重复的平均值±标准差(n=6)。不同小写字母的柱值表示差异显著(P<0.05) (Student-Neuman-Keuls)。 Figure 3Trehalose treatment maintain the photosynthetic performance of rice leaves under drought stress Fv/Fm
(A), Fv′/Fm′ (B), ΦPSII (C), qP (D) and NPQ (E) of PC and WT after 2 hours of simulated drought treatment. Abbreviations are the same as those given in Figure 1. Values represent means±SD (n=6) from three biological replicates. Bars labeled with different lowercase letters indicate significant differences (P<0.05) (Student-Neuman-Keuls).
2.6 海藻糖处理调节干旱胁迫下水稻叶片内海藻糖代谢相关基因的表达
本研究表明, 在DS处理后, PC内源海藻糖含量与CK相比显著上升; 而DS+Tre处理进一步上调PC的海藻糖含量, 且PC显著高于WT (表3)。是否外施海藻糖调节水稻内源海藻糖代谢? 而且为什么PC的含量高于WT? 已有研究表明, 海藻糖代谢相关基因在调控能量平衡中发挥作用(Ge et al., 2008; Zang et al., 2011; Islam et al., 2019)。OsTre1是目前唯一已知参与调控海藻糖酶合成的基因, 海藻糖酶通过分解海藻糖降低植物中海藻糖的含量(Islam et al., 2019)。因此, 本研究进一步分析了海藻糖合成相关基因(OsTPP1、OsTPP2、OsTPP3和OsTPP7)、海藻糖分解相关基因(OsTre1)以及海藻糖合成前体T6P合成相关基因(OsTPP1、OsTPS2和OsTPS8)的表达(图4A)。与CK相比, OsTPP2、OsTPP3和OsTPP7在DS处理下表达量均显著上升, 且PC内的基因表达量均显著高于WT; 与DS相比, DS+Tre处理下两供试材料的OsTPP1和OsTPP3表达量进一步上调, 而OsTPP2和OsTPP7表达量下调, 但PC始终显著高于WT (附表1)。在WT中, DS处理下OsTre1的表达量较CK处理显著上调; 与DS相比, DS+Tre处理在PC和WT中OsTre1的表达量均下调, 且在DS和DS+Tre处理下, PC始终低于WT且差异显著(附表1; 图4A)。此外, 在DS处理下, PC中与海藻糖上游合成前体T6P相关的OsTPS2和OsTPS8表达量较CK均显著上升, DS+Tre处理使其表达量进一步上调, 且PC显著高于WT (附表1)。但WT中上述基因在各处理下变幅不大。可见, 在干旱条件下, 海藻糖处理通过上调海藻糖合成以及下调其分解相关基因的转录水平来提高PC的内源海藻糖含量, 从而表现出较高的海藻糖水平。
Table 3 表3 表3海藻糖处理改变干旱胁迫下水稻叶片内可溶性糖的积累与分配 Table 3Trehalose treatments change the accumulation and distribution of soluble sugars in rice leaves under drought stress
Index (mg·g-1 FW)
WT
PC
CK
CK+Tre
DS
DS+Tre
CK
CK+Tre
DS
DS+Tre
Soluble sugar content
16.10 e
16.50 e
29.01 c
34.32 b
14.97 f
19.01 d
35.31 b
41.57 a
Sucrose content
9.40 e
10.78 d
10.34 d
12.36 b
8.51 f
11.18 c
11.61 c
13.81 a
Fructose content
4.01 d
4.09 d
4.95 c
5.81 b
3.86 e
3.88 e
5.49 b
7.66 a
Glucose content
1.64 b
1.28 c
2.35 a
1.62 b
0.88 d
0.78 e
1.32 c
0.91 d
Trehalose content
8.22 f
9.46 e
10.84 d
13.60 b
8.55 f
10.40 d
11.23 c
15.80 a
Abbreviations are the same as those given in Figure 1. Values represent means±SD (n=6) from three biological replicates. Different lowercase letters indicate significant differences (P<0.05) (Student-Neuman-Keuls). 缩写同图1。表中数值代表来自3个生物学重复的平均值±标准差(n=6)。不同小写字母表示差异显著(P<0.05) (Student-Neuman- Keuls)。
(A) 海藻糖合成与代谢相关基因的表达; (B) SnRKs相关基因的表达; (C) ABA代谢相关基因的表达。缩写同图1。热图显示基因相对表达量对数转换均一化值, 显著性分析表示基因相对表达量之间的差异(附表1)。图中不同小写字母表示差异显著(P<0.05) (Student-Neuman-Keuls)。 Figure 4Trehalose treatments affect the expression of trehalose, SnRKs and ABA-related genes in rice leaves under drought stress
(A) The expression of trehalose biosynthesis and metabolism related genes; (B) The expression of SnRKSs-related genes; (C) The expression of ABA metabolism-related genes. Abbreviations are the same as those given in Figure 1. The heat map shows the log-transformed normalized values of relative gene expression, and the significance analysis showed the difference between relative gene expressions (Appendix Table 1). Different lowercase letters indicate significant differences (P<0.05) (Student-Neuman-Keuls).
已有研究表明, 水稻OsbZIP23和OsMYB2是响应ABA耐旱的转录因子(Yang et al., 2012; Dey et al., 2016), 而OsLEA3、OsNAC6、OsRab16b和OsRab21等作为靶基因(Hong et al., 2009; Duan and Cai, 2012; Hong et al., 2016)在下游发挥作用。为了进一步明确海藻糖与ABA在PC干旱调节中的机制, 本研究测定了ABA耐旱相关转录因子和靶基因的表达量(图4C)。结果表明, 在PC中, DS处理上调了OsbZIP23的表达量; 相比DS, DS+Tre处理进一步上调了OsbZIP23和OsMYB2的转录水平, 且PC显著高于WT。同时, OsRab16b、OsRab21、OsLEA3和OsNAC6在PC叶内CK、DS和DS+Tre处理下的表达模式与OsbZIP23和OsMYB2基本保持一致, 且上述基因的表达水平在DS+Tre处理下均表现为PC显著高于WT (图4C; 附表1)。综上, 与WT相比, 海藻糖可能通过诱导ABA信号转导途径上调与ABA相关的耐旱转录因子及其靶基因的表达水平, 进而增强PC的耐旱性。
2.9 海藻糖处理诱导干旱胁迫下水稻叶片内Ca2+和NO的产生以及NR和HXK的酶活性
Ca2+和NO是调节植物耐旱性的重要第二信使分子(Corpas and Barroso, 2018; Liu et al., 2020)。本研究表明, 两信号分子的含量在WT和PC间存在差异, 其中Ca2+水平在WT的各处理间基本保持稳定(图5A, B)。相比CK, PC中DS处理下的Ca2+含量显著升高, 但PC显著低于WT (图5A)。而DS+Tre处理进一步显著增加了Ca2+含量, 且PC显著高于WT。有意思的是, 与CK相比, DS处理显著下调了两试材的NO含量, 而DS+Tre处理仅提升了PC叶内的NO水平, 且PC中NO含量在CK+Tre、DS及DS+Tre处理下均显著高于WT (图5B)。以上表明干旱条件下PC中的NO可能作为信号, 低水平时促进ABA的信号转导, 高水平时则起到反馈抑制作用。可见, 与DS处理相比, 海藻糖处理有利于启动信号分子参与干旱响应, 且较WT, PC的促进效应更显著。
(A) 钙离子含量; (B) 一氧化氮含量; (C) 硝酸还原酶活性; (D) 己糖激酶活性。缩写同图1。图中数值代表来自3个生物学重复的平均值±标准差(n=6)。不同小写字母的柱值表示差异显著(P<0.05) (Student-Neuman-Keuls)。 Figure 5Trehalose treatments affect the content of signal molecules and the activity of related enzymes in rice leaves under drought stress
(A) Ca2+ content; (B) NO content; (C) Nitate reductase (NR) activity; (D) Hexokinase (HXK) activity. Abbreviations are the same as those given in Figure 1. Values represent means±SD (n=6) from three biological replicates. Bars labeled with different lowercase letters indicate significant differences (P<0.05) (Student-Neuman-Keuls).
AkramNA, WaseemM, AmeenR, AshrafM (2016). Trehalose pretreatment induces drought tolerance in radish (Raphanus sativus L.) plants: some key physio-biochemical traits Acta Physiol Plant 38, 3. DOI:10.1007/s11738-015-2018-1URL [本文引用: 2]
AlamMM, NaharK, HasanuzzamanM, FujitaM (2014). Trehalose-induced drought stress tolerance: a comparative study among different brassica species Plant Omics J 7, 271-283. [本文引用: 1]
AmbavaramMMR, BasuS, KrishnanA, RamegowdaV, BatlangU, RahmanL, BaisakhN, PereiraA (2014). Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress Nat Commun 5, 5302. DOI:10.1038/ncomms6302PMID:25358745 [本文引用: 2] Plants capture solar energy and atmospheric carbon dioxide (CO2) through photosynthesis, which is the primary component of crop yield, and needs to be increased considerably to meet the growing global demand for food. Environmental stresses, which are increasing with climate change, adversely affect photosynthetic carbon metabolism (PCM) and limit yield of cereals such as rice (Oryza sativa) that feeds half the world. To study the regulation of photosynthesis, we developed a rice gene regulatory network and identified a transcription factor HYR (HIGHER YIELD RICE) associated with PCM, which on expression in rice enhances photosynthesis under multiple environmental conditions, determining a morpho-physiological programme leading to higher grain yield under normal, drought and high-temperature stress conditions. We show HYR is a master regulator, directly activating photosynthesis genes, cascades of transcription factors and other downstream genes involved in PCM and yield stability under drought and high-temperature environmental stress conditions.
AsamiP, RupasingheT, MoghaddamL, NjaciI, RoessnerU, MundreeS, WilliamsB (2019). Roots of the resurrection plant tripogon loliiformis survive desiccation without the activation of autophagy pathways by maintaining energy reserves Front Plant Sci 10, 459. DOI:10.3389/fpls.2019.00459URL [本文引用: 1]
Baena-GonzálezE, LunnJE (2020). SnRK1 and trehalose 6-phosphate: two ancient pathways converge to regulate plant metabolism and growth Curr Opin Plant Biol 55, 52-59. DOI:10.1016/j.pbi.2020.01.010URL [本文引用: 1]
BlázquezMA, LagunasR, GancedoC, GancedoJM (1993). Trehalose-6-phosphate, a new regulator of yeast glycolysis that inhibits hexokinases FEBS Lett 329, 51-54. PMID:8354408 [本文引用: 1] Trehalose-6-phosphate (P) competitively inhibited the hexokinases from Saccharomyces cerevisiae. The strongest inhibition was observed upon hexokinase II, with a Ki of 40 microM, while in the case of hexokinase I the Ki was 200 microM. Glucokinase was not inhibited by trehalose-6-P up to 5 mM. This inhibition appears to have physiological significance, since the intracellular levels of trehalose-6-P were about 0.2 mM. Hexokinases from other organisms were also inhibited, while glucokinases were unaffected. The hexokinase from the yeast, Yarrowia lipolytica, was particularly sensitive to the inhibition by trehalose-6-P: when assayed with 2 mM fructose an apparent Ki of 5 microM was calculated. Two S. cerevisiae mutants with abnormal levels of trehalose-6-P exhibited defects in glucose metabolism. It is concluded that trehalose-6-P plays an important role in the regulation of the first steps of yeast glycolysis, mainly through the inhibition of hexokinase II.
ChenPB, LiX, HuoK, WeiXD, DaiCC, LuCG (2014). Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors J Plant Physiol 171, 458-466. DOI:10.1016/j.jplph.2013.11.006URL [本文引用: 2]
ClaeysH, ViSL, XuXS, Satoh-NagasawaN, EvelandAL, GoldshmidtA, FeilR, BeggsGA, SakaiH, BrennanRG, LunnJE, JacksonD (2019). Control of meristem determinacy by trehalose 6-phosphate phosphatases is uncoupled from enzymatic activity Nat Plants 5, 352-357. DOI:10.1038/s41477-019-0394-zURL [本文引用: 1]
CorpasFJ, BarrosoJB (2018). Peroxisomal plant metabolism: an update on nitric oxide, Ca 2+ and the NADPH recycling network J Cell Sci 131, jcs202978. [本文引用: 2]
DelorgeI, JaniakM, CarpentierS, van DijckF (2014). Fine tuning of trehalose biosynthesis and hydrolysis as novel tools for the generation of abiotic stress tolerant plants Front Plant Sci 5, 147. DOI:10.3389/fpls.2014.00147PMID:24782885 [本文引用: 2] The impact of abiotic stress on plant growth and development has been and still is a major research topic. An important pathway that has been linked to abiotic stress tolerance is the trehalose biosynthetic pathway. Recent findings showed that trehalose metabolism is also important for normal plant growth and development. The intermediate compound trehalose-6-phosphate (T6P) is now confirmed to act as a sensor for available sucrose, hereby directly influencing the type of response to the changing environmental conditions. This is possible because T6P and/or trehalose or their biosynthetic enzymes are part of complex interaction networks with other crucial hormone and sugar-induced signaling pathways, which may function at different developmental stages. Because of its effect on plant growth and development, modification of trehalose biosynthesis, either at the level of T6P synthesis, T6P hydrolysis, or trehalose hydrolysis, has been utilized to try to improve crop yield and biomass. It was shown that alteration of the amounts of either T6P and/or trehalose did result in increased stress tolerance, but also resulted in many unexpected phenotypic alterations. A main challenge is to characterize the part of the signaling pathway resulting in improved stress tolerance, without affecting the pathways resulting in the unwanted phenotypes. One such specific pathway where modification of trehalose metabolism improved stress tolerance, without any side effects, was recently obtained by overexpression of trehalase, which results in a more sensitive reaction of the stomatal guard cells and closing of the stomata under drought stress conditions. We have used the data that have been obtained from different studies to generate the optimal plant that can be constructed based on modifications of trehalose metabolism.
DeyA, SamantaMK, GayenS, SenSK, MaitiMK (2016). Enhanced gene expression rather than natural polymorphism in coding sequence of the OsbZIP23 determines drought tolerance and yield improvement in rice genotypes PLoS One 11, e0150763. DOI:10.1371/journal.pone.0150763URL [本文引用: 1]
DingZS, HuangSH, ZhouBY, SunXF, ZhaoM (2013). Over-expression of phosphoenolpyruvate carboxylase cDNA from C4 millet ( Seteria italica) increase rice photosynthesis and yield under upland condition but not in wetland fields Plant Biotechnol Rep 7, 155-163. DOI:10.1007/s11816-012-0244-1URL [本文引用: 1]
DingZS, ZhouBY, SunXF, ZhaoM (2012). High light tolerance is enhanced by overexpressed PEPC in rice under drought stress Acta Agron Sin 38, 285-292. [本文引用: 1]
DuanJL, CaiWM (2012). OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance PLoS One 7, e45117. DOI:10.1371/journal.pone.0045117URL [本文引用: 1]
DurandM, MainsonD, PorcheronB, MauroussetL, LemoineR, PourtauN (2018). Carbon source-sink relationship in Arabidopsis thaliana: the role of sucrose transporters Planta 247, 587-611. DOI:10.1007/s00425-017-2807-4URL [本文引用: 1]
ErmakovaM, DanilaFR, FurbankRT, CaemmererS (2020). On the road to C4 rice: advances and perspectives Plant J 101, 940-950. DOI:10.1111/tpj.14562 [本文引用: 1] The international C-4 rice consortium aims to introduce into rice a high capacity photosynthetic mechanism, the C-4 pathway, to increase yield. The C-4 pathway is characterised by a complex combination of biochemical and anatomical specialisation that ensures high CO2 partial pressure at RuBisCO sites in bundle sheath (BS) cells. Here we report an update of the progress of the C-4 rice project. Since its inception in 2008 there has been an exponential growth in synthetic biology and molecular tools. Golden Gate cloning and synthetic promoter systems have facilitated gene building block approaches allowing multiple enzymes and metabolite transporters to be assembled and expressed from single gene constructs. Photosynthetic functionalisation of the BS in rice remains an important step and there has been some success overexpressing transcription factors in the cytokinin signalling network which influence chloroplast volume. The C-4 rice project has rejuvenated the research interest in C-4 photosynthesis. Comparative anatomical studies now point to critical features essential for the design. So far little attention has been paid to the energetics. C-4 photosynthesis has a greater ATP requirement, which is met by increased cyclic electron transport in BS cells. We hypothesise that changes in energy statues may drive this increased capacity for cyclic electron flow without the need for further modification. Although increasing vein density will ultimately be necessary for high efficiency C-4 rice, our modelling shows that small amounts of C-4 photosynthesis introduced around existing veins could already provide benefits of increased photosynthesis on the road to C-4 rice.
FengYC, ChenXY, HeYL, KouXH, XueZH (2019). Effects of exogenous trehalose on the metabolism of sugar and abscisic acid in tomato seedlings under salt stress Trans Tianjin Univ 25, 451-471. DOI:10.1007/s12209-019-00214-xURL [本文引用: 3]
FichtnerF, LunnJE (2021). The role of trehalose 6-phosphate (Tre6P) in plant metabolism and development Annu Rev Plant Biol 72, 187-220. [本文引用: 1]
FigueroaCM, LunnJE (2016). A tale of two sugars: trehalose 6-phosphate and sucrose Plant Physiol 172, 7-27. [本文引用: 2]
FoyerCH, HalliwellB (1976). The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism Planta 133, 21-25. DOI:10.1007/BF00386001PMID:24425174 [本文引用: 1] Both glutathione and an NADPH-dependent glutathione reductase are present in spinach (Spinacia oleracea L.) chloroplasts. It is proposed that glutathione functions to stabilise enzymes of the Calvin cycle, and it may also act to keep ascorbic acid in chloroplasts in the reduced form.
GeLF, ChaoDY, ShiM, ZhuMZ, GaoJP, LinHX (2008). Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes Planta 228, 191-201. DOI:10.1007/s00425-008-0729-xURL [本文引用: 3]
GiannopolitisCN, RiesSK (1977). Superoxide dismutases I: occurrence in higher plants Plant Physiol 59, 309-314. PMID:16659839 [本文引用: 1] Shoots, roots, and seeds of corn (Zea mays L., cv. Michigan 500), oats (Avena sativa L., cv. Au Sable), and peas (Pisum sativum L., cv. Wando) were analyzed for their superoxide dismutase content using a photochemical assay system consisting of methionine, riboflavin, and p-nitro blue tetrazolium. The enzyme is present in the shoots, roots, and seeds of the three species. On a dry weight basis, shoots contain more enzyme than roots. In seeds, the enzyme is present in both the embryo and the storage tissue. Electrophoresis indicated a total of 10 distinct forms of the enzyme. Corn contained seven of these forms and oats three. Peas contained one of the corn and two of the oat enzymes. Nine of the enzyme activities were eliminated with cyanide treatment suggesting that they may be cupro-zinc enzymes, whereas one was cyanide-resistant and may be a manganese enzyme. Some of the leaf superoxide dismutases were found primarily in mitochondria or chloroplasts. Peroxidases at high concentrations interfere with the assay. In test tube assays of crude extracts from seedlings, the interference was negligible. On gels, however, peroxidases may account for two of the 10 superoxide dismutase forms.
Giglioli-Guivarc'hG, PierreJN, BrownS, CholletR, VidalJ, GadalP (1996). The light-dependent transduction pathway controlling the regulatory phosphorylation of C4 phosphoenolpyruvate carboxylase in protoplasts from digitaria sanguinalis Plant Cell 8, 573-586. DOI:10.2307/3870336URL [本文引用: 1]
GómezLD, GildayA, FeilR, LunnJE, GrahamIA (2010). AtTPS1-mediated trehalose 6-phosphate synthesis is essential for embryogenic and vegetative growth and responsiveness to ABA in germinating seeds and stomatal guard cells Plant J 64, 1-13. [本文引用: 1]
HanBY, FuLL, ZhangD, HeXQ, ChenQ, PengM, ZhangJM (2016). Interspecies and intraspecies analysis of trehalose contents and the biosynthesis pathway gene family reveals crucial roles of trehalose in osmotic-stress tolerance in cassava Int J Mol Sci 17, 1077. DOI:10.3390/ijms17071077URL [本文引用: 1]
HeYF, XieYF, LiX, YangJ (2020). Drought tolerance of transgenic rice overexpressing maize C4-PEPC gene related to increased anthocyanin synthesis regulated by sucrose and calcium Biol Plantarum 64, 136-149. DOI:10.32615/bp.2020.031URL [本文引用: 3]
HongCY, ChaoYY, YangMY, ChengSY, ChoSC, KaoCH (2009). NaCl-induced expression of glutathione reductase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid Plant Soil 320, 103-115. DOI:10.1007/s11104-008-9874-zURL [本文引用: 1]
HongYB, ZhangHJ, HuangL, LiDY, SongFM (2016). Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice Front Plant Sci 7, 4. [本文引用: 1]
HuoK, LiX, HeYF, WeiXD, LuW, ZhaoCF, WangCL (2017). Exogenous ATP enhance signal response of suspension cells of transgenic rice ( Oryza sativa L.) expressing maize C4-pepc encoded phosphoenolpyruvate carboxylase under PEG treatment Plant Growth Regul 82, 55-67. DOI:10.1007/s10725-016-0238-zURL [本文引用: 2]
IlhanS, OzdemirF, BorM (2015). Contribution of trehalose biosynthetic pathway to drought stress tolerance of Capparis ovata desf Plant Biol 17, 402-407. DOI:10.1111/plb.12227URL [本文引用: 1]
IslamMO, KatoH, ShimaS, TezukaD, MatsuiH, ImaiR (2019). Functional identification of a rice trehalose gene involved in salt stress tolerance Gene 685, 42-49. DOI:10.1016/j.gene.2018.10.071URL [本文引用: 2]
JiangDG, ChenWT, GaoJ, YangF, ZhuangCX (2019). Overexpression of the trehalose-6-phosphate phosphatase OsTPP3 increases drought tolerance in rice Plant Biotechnol Rep 13, 285-292. DOI:10.1007/s11816-019-00541-4URL [本文引用: 2]
JiaoDM, JiBH, LiX (2003). Characteristics of chlorophyll fluorescence and membrane-lipid peroxidation during senescence of flag leaf in different cultivars of rice Photosynthetica 41, 33-41. DOI:10.1023/A:1025848110029URL [本文引用: 1]
JohnR, RajaV, AhmadM, JanN, MajeedU, AhmadS, YaqoobU, KaulT (2017). Trehalose: metabolism and role in stress signaling in plants. In: Sarwat M, Ahmad A, Abdin M, Ibrahim M, eds. Stress Signaling in Plants: Genomics and Proteomics Perspective, Volume 2. Cham: Springer International Publishing. pp. 261-275. [本文引用: 2]
KarkiS, RizalG, QuickWP (2013). Improvement of photosynthesis in rice (Oryza sativa L.) by inserting the C4 pathway Rice 6, 28. DOI:10.1186/1939-8433-6-28URL [本文引用: 1]
KimYM, HeinzelN, GieseJO, KoeberJ, MelzerM, RuttenT, von WirénN, SonnewaldU, HajirezaeiMR (2013). A dual role of tobacco hexokinase 1 in primary metabolism and sugar sensing Plant Cell Environ 36, 1311-1327. DOI:10.1111/pce.2013.36.issue-7URL [本文引用: 1]
KretzschmarT, PelayoMAF, TrijatmikoKR, GabunadaLFM, AlamR, JimenezR, MendioroMS, Slamet- LoedinIH, SreenivasuluN, Bailey-SerresJ, IsmailAM, MackillDJ, SeptiningsihEM (2015). A trehalose-6- phosphate phosphatase enhances anaerobic germination tolerance in rice Nat Plants 1, 15124. DOI:10.1038/nplants.2015.124PMID:27250677 [本文引用: 3] Global socioeconomic developments create strong incentives for farmers to shift from transplanted to direct-seeded rice (DSR) as a means of intensification and economization(1). Rice production must increase to ensure food security(2) and the bulk of this increase will have to be achieved through intensification of cultivation, because expansion of cultivated areas is reaching sustainable limits(3). Anaerobic germination tolerance, which enables uniform germination and seedling establishment under submergence(4), is a key trait for the development of tropical DSR varieties(5,6). Here, we identify a trehalose-6-phosphate phosphatase gene, OsTPP7, as the genetic determinant in qAG-9-2, a major quantitative trait locus (QTL) for anaerobic germination tolerance(7). OsTPP7 is involved in trehalose-6-phosphate (T6P) metabolism, central to an energy sensor that determines anabolism or catabolism depending on local sucrose availability(8,9). OsTPP7 activity may increase sink strength in proliferating heterotrophic tissues by indicating low sugar availability through increased T6P turnover, thus enhancing starch mobilization to drive growth kinetics of the germinating embryo and elongating coleoptile, which consequently enhances anaerobic germination tolerance.
KuMSB, AgarieS, NomuraM, FukayamaH, TsuchidaH, OnoK, HiroseS, TokiS, MiyaoM, MatsuokaM (1999). High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants Nat Biotechnol 17, 76-80. PMID:9920274 [本文引用: 2] Using an Agrobacterium-mediated transformation system, we have introduced the intact gene of maize phosphoenolpyruvate carboxylase (PEPC), which catalyzes the initial fixation of atmospheric CO2 in C4 plants into the C3 crop rice. Most transgenic rice plants showed high-level expression of the maize gene; the activities of PEPC in leaves of some transgenic plants were two- to threefold higher than those in maize, and the enzyme accounted for up to 12% of the total leaf soluble protein. RNA gel blot and Southern blot analyses showed that the level of expression of the maize PEPC in transgenic rice plants correlated with the amount of transcript and the copy number of the inserted maize gene. Physiologically, the transgenic plants exhibited reduced O2 inhibition of photosynthesis and photosynthetic rates comparable to those of untransformed plants. The results demonstrate a successful strategy for installing the key biochemical component of the C4 pathway of photosynthesis in C3 plants.
LatzkoE, KellyGJ (1983). The many-faceted function of phosphoenolpyruvate carboxylase in C3 plants Physiol Végét 21, 805-815. [本文引用: 1]
LiCX, ShenHY, WangT, WangXL (2015). ABA regulates subcellular redistribution of OsABI-LIKE2, a negative regulator in ABA signaling, to control root architecture and drought resistance in Oryza sativa Plant Cell Physiol 56, 2396-2408. DOI:10.1093/pcp/pcv154URL [本文引用: 1]
LiX, WangC, RenCG (2011). Effects of 1-butanol, neomycin and calcium on the photosynthetic characteristics of pepc transgenic rice Afr J Biotechnol 10, 17466-17476. [本文引用: 2]
LiZW, ZhaoQ, ChengFM (2020). Sugar starvation enhances leaf senescence and genes involved in sugar signaling pathways regulate early leaf senescence in mutant rice Rice Sci 27, 201-204. DOI:10.1016/j.rsci.2019.11.001URL [本文引用: 1]
LiuT, YeXL, LiM, LiJM, QiHY, HuXH (2020). H2O2 and NO are involved in trehalose-regulated oxidative stress tolerance in cold-stressed tomato plants Environ Exp Bot 171, 103961. DOI:10.1016/j.envexpbot.2019.103961URL [本文引用: 2]
MamedovTG, MoelleringER, CholletR (2005). Identification and expression analysis of two inorganic C- and N- responsive genes encoding novel and distinct molecular forms of eukaryotic phosphoenolpyruvate carboxylase in the green microalga Chlamydomonas reinhardtii Plant J 42, 832-843. PMID:15941397 [本文引用: 1] Phosphoenolpyruvate carboxylase (PEPC [Ppc]) has been previously purified and characterized in biochemical and immunological terms from two green microalgae, Chlamydomonas reinhardtii and Selenastrum minutum. The findings indicate that these algae possess at least two distinct PEPC enzyme-forms, homotetrameric Class-1 and heteromeric Class-2, that differ significantly from each other and their plant and prokaryotic counterparts. Surprisingly, however, green-algal PEPC has been unexplored to date in molecular terms. This study reports the molecular cloning of the two Ppc genes in C. reinhardtii (CrPpc1, CrPpc2), each of which is transcribed in vivo and encodes a fully active, recombinant PEPC that lacks the regulatory, N-terminal seryl-phosphorylation domain that typifies the vascular-plant enzyme. These distinct catalytic subunit-types differ with respect to their (i) predicted molecular mass ( approximately 108.9 [CrPpc1] versus approximately 131.2 kDa [CrPpc2]) and critical C-terminal tetrapeptide; and (ii) immunoreactivity with antisera against the p102 and p130 polypeptides of S. minutum PEPC1/PEPC2 and PEPC2, respectively. Only the Ppc1 transcript encodes the p102 catalytic subunits common to both Class-1 and Class-2 enzyme-forms in C. reinhardtii. The steady-state transcript levels of both CrPpc1/2 are coordinately up-/down-regulated by changes in [CO2] or [NH] during growth, and generally mirror the response of cytoplasmic glutamine synthetase (Gs1) transcript abundance to changes in inorganic [N] at 5% CO2. These collective findings provide key molecular insight into the Ppc genes and corresponding PEPC catalytic subunits in the eukaryotic algae.
MatsuokaM, MinamiEI (1989). Complete structure of the gene for phosphoenolpyruvate carboxylase from maize Eur J Biochem 181, 593-598. PMID:2731539 [本文引用: 1] Phosphoenolpyruvate carboxylase is a key enzyme in photosynthesis in some plants that exploit the C4 photosynthetic pathway for the fixation of CO2. We cloned the gene for this enzyme from maize genomic libraries and analyzed its complete primary structure. The sequence of the cloned gene spans 6781 bp and consists of 10 exons and 9 introns. The site of initiation of transcription is located 84 nucleotides upstream from the first nucleotide of the initiation codon (position -84), as determined by the method of primer-extension analysis. The analysis suggests that there is another initiation site located at position -81. The 5'-flanking region of the gene lacks typical TATA and CCAAT elements in the anticipated regions, but there is a TATA-similar sequence (TATTT) around the -30 regions as well as sequence homologous to the Sp-1 protein-binding site (CCGCCC). Six long, direct repeated sequences and a light-responsive element are also present in the 5'-flanking region. The results of Southern blot analysis indicated that the phosphoenolpyruvate carboxylase gene exists as a small multi-gene family, but the enzyme that is expressed at high levels in green leaves and is involved in C4 photosynthesis is encoded by a single-copy gene in the maize genome.
MoralesF, AncínM, FakhetD, González-TorralbaJ, GámezAL, SeminarioA, SobaD, Ben MariemS, GarrigaM, AranjueloI (2020). Photosynthetic metabolism under stressful growth conditions as a bases for crop breeding and yield improvement Plants 9, 88. DOI:10.3390/plants9010088URL [本文引用: 1]
NakanoY, AsadaK (1981). Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts Plant Cell Physiol 22, 867-880. [本文引用: 1]
NoctorG, ReichheldJP, FoyerCH (2018). ROS-related redox regulation and signaling in plants Semin Cell Dev Biol 80, 3-12. DOI:10.1016/j.semcdb.2017.07.013URL [本文引用: 1]
OladosuY, RafiiMY, SamuelC, FataiA, MagajiU, KareemI, KamarudinZS, MuhammadI, KolapoK (2019). Drought resistance in rice from conventional to molecular breeding: a review Intl J Mol Sci 20, 3519. DOI:10.3390/ijms20143519URL [本文引用: 1]
PramanikMHR, ImaiR (2005). Functional identification of a trehalose 6-phosphate phosphatase gene that is involved in transient induction of trehalose biosynthesis during chilling stress in rice Plant Mol Biol 58, 751-762. DOI:10.1007/s11103-005-7404-4PMID:16240171 [本文引用: 1] Trehalose serves as a stress protectant and/or reserve carbohydrate in a variety of organisms including bacteria, yeast, and invertebrates. Recently, trace amounts of trehalose have been detected in higher plants, although the function of trehalose in plants remains unknown. A cDNA clone (OsTPP1) encoding a putative trehalose-6-phosphate phosphatase (TPP) for trehalose biosynthesis was isolated from rice. Functionality of the clone was demonstrated by complementation of a yeast mutant and enzymatic activity of the recombinant protein. Northern blots revealed that the OsTPP1 transcript levels were fairly low or under detectable limits in most of the tissues under ambient conditions but were highly induced within 1-2 h of chilling stress (12 degrees C) in both root and shoot tissues of seedlings. This induction was transient and disappeared after 6 h of the chilling stress. Transient expression of OsTPP1 was also induced under severe chilling stress (4 degrees C) as well as salinity and drought stresses at ambient temperatures. Application of exogenous ABA (50 microM) resulted in a transient increase of OsTPP1 expression within 20 min of the treatment, thereby suggesting involvement of ABA in OsTPP1 gene regulation. Measurements of total cellular TPP activity and trehalose content in roots indicated that both TPP activity and trehalose levels were transiently increased after chilling (12 degrees C) stress. Collectively, the data indicate that transient activation of trehalose biosynthesis is involved in early chilling stress response in rice. Possible functions of trehalose in the early stages of chilling stress response are discussed.
QianBY, LiX, LiuXL, ChenPB, RenCG, DaiCC (2015a). Enhanced drought tolerance in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene via NO and Ca 2+ J Plant Physiol 175, 9-20. DOI:10.1016/j.jplph.2014.09.019URL [本文引用: 2]
QianBY, LiX, LiuXL, WangM (2015b). Improved oxidative tolerance in suspension-cultured cells of C4-pepc transgenic rice by H2O2 and Ca 2+ under PEG-6000 J Integr Plant Biol 57, 534-539. DOI:10.1111/jipb.12283URL [本文引用: 2]
SadakMS (2016). Mitigation of drought stress on fenugreek plant by foliar application of trehalose Int J ChemTech Res 9, 147-155. [本文引用: 1]
SamadiS, HabibiG, VaziriA (2019). Exogenous trehalose alleviates the inhibitory effects of salt stress in strawberry plants Acta Physiol Plant 41, 112. DOI:10.1007/s11738-019-2905-yURL [本文引用: 1]
SchafferAA, PetreikovM (1997). Sucrose-to-starch metabolism in tomato fruit undergoing transient starch accumulation Plant Physiol 113, 739-746. DOI:10.1104/pp.113.3.739URL [本文引用: 1]
ShahbazM, AbidA, MasoodA, WaraichEA (2017). Foliar-applied trehalose modulates growth, mineral nutrition, photosynthetic ability, and oxidative defense system of rice (Oryza sativa L.) under saline stress J Plant Nutr 40, 584-599. DOI:10.1080/01904167.2016.1263319URL [本文引用: 1]
ShiJH, YiKK, LiuY, XieL, ZhouZJ, ChenY, HuZH, ZhengT, LiuRH, ChenYL, ChenJQ (2015). Phospho enol pyruvate carboxylase in Arabidopsis leaves plays a crucial role in carbon and nitrogen metabolism Plant Physiol 167, 671-681. DOI:10.1104/pp.114.254474URL [本文引用: 1]
SimonNML, KusakinaJ, Fernández-Lópezá, ChembathA, BelbinFE, DoddAN (2018). The energy-signaling hub SnRK1 is important for sucrose-induced hypocotyl elongation Plant Physiol 176, 1299-1310. DOI:10.1104/pp.17.01395URL [本文引用: 2]
SmartRE, BinghamGE (1974). Rapid estimates of relative water content Plant Physiol 53, 258-260. PMID:16658686 [本文引用: 1] Relative water content may be accurately estimated using the ratio of tissue fresh weight to tissue turgid weight, termed here relative tissue weight. That relative water content and relative tissue weight are linearly related is demonstrated algebraically. The mean value of r(2) for grapevine (Vitis vinifera L. cv. Shiraz) leaf tissue over eight separate sampling occasions was 0.993. Similarly high values were obtained for maize (Zea mays cv. Cornell M-3) (0.998) and apple (Malus sylvestris cv. Northern Spy) (0.997) using a range of leaf ages. The proposal by Downey and Miller (1971. Rapid measurements of relative turgidity in maize (Zea mays L.). New Phytol. 70: 555-560) that relative water content in maize may be estimated from water uptake was also investigated for grapevine leaves; this was found to be a less reliable estimate than that obtained with relative tissue weight. With either method, there is a need for calibration, although this could be achieved for relative tissue weight at least with only a few subsamples.
SunSJ, QiGN, GaoQF, WangHQ, YaoFY, HussainJ, WangYF (2016). Protein kinase OsSAPK8 functions as an essential activator of S-type anion channel OsSLAC1, which is nitrate-selective in rice Planta 243, 489-500. DOI:10.1007/s00425-015-2418-xURL [本文引用: 1]
TachibaanaS, KonishiN, KandaH (1991). Diurnal variation of in vivo and in vitro nitrate reductase activity in cucumber plants J Jap Soc Hortic Sci 60, 593-599. DOI:10.2503/jjshs.60.593URL [本文引用: 1]
TheerakulpP, PhongngarmS (2013). Alleviation of adverse effects of salt stress on rice seedlings by exogenous trehalose Asian J Crop Sci 5, 405-415. DOI:10.3923/ajcs.2013.405.415URL [本文引用: 1]
TianLF, XieZJ, LuCQ, HaoXH, WuS, HuangY, LiDP, ChenLB (2019). The trehalose-6-phosphate synthase TPS5 negatively regulates ABA signaling in Arabidopsis thaliana Plant Cell Rep 38, 869-882. DOI:10.1007/s00299-019-02408-yURL [本文引用: 2]
UzildayB, TurkanI, OzgurR, SekmenAH (2014). Strategies of ROS regulation and antioxidant defense during transition from C3 to C4 photosynthesis in the genus Flaveria under PEG-induced osmotic stress J Plant Physiol 171, 65-75. DOI:10.1016/j.jplph.2013.06.016URL [本文引用: 3]
VelikovaV, YordanovI, EdrevaA (2000). Oxidative stress and some antioxidant systems in acid rain-treated bean plants Plant Sci 151, 59-66. DOI:10.1016/S0168-9452(99)00197-1URL [本文引用: 1]
VishalB, KrishnamurthyP, RamamoorthyR, KumarPP (2019). OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition New Phytol 221, 1369-1386. DOI:10.1111/nph.2019.221.issue-3URL [本文引用: 1]
WangPC, ZhuJK, LangZB (2015). Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins Plant Signal Behav 10, e1031939. DOI:10.1080/15592324.2015.1031939URL [本文引用: 2]
WangWJ, ChenQB, XuSM, LiuWC, ZhuXH, SongCP (2020a). Trehalose-6-phosphate phosphatase E modulates ABA-controlled root growth and stomatal movement inArabidopsis J Integr Plant Biol 62, 1518-1534. DOI:10.1111/jipb.v62.10URL [本文引用: 1]
WangY, WangLP, MicallefBJ, TetlowIJ, MullenRT, FeilR, LunnJE, EmesMJ (2020b). AKINβ1, a subunit of SnRK1, regulates organic acid metabolism and acts as a global modulator of genes involved in carbon, lipid, and nitrogen metabolism J Exp Bot 71, 1010-1028. [本文引用: 1]
WinglerA (2018). Transitioning to the next phase: the role of sugar signaling throughout the plant life cycle Plant Physiol 176, 1075-1084. DOI:10.1104/pp.17.01229URL [本文引用: 2]
YangA, DaiXY, ZhangWH (2012). A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice J Exp Bot 63, 2541-2556. DOI:10.1093/jxb/err431URL [本文引用: 1]
YoshidaS, FornoDA, CockJ (1971). Laboratory Manual for Physiological Studies of Rice Los Banos: International Rice Research Institute. pp. 62. [本文引用: 1]
YuWQ, ZhaoRR, WangL, ZhangSJ, LiR, ShengJP, ShenL (2019). ABA signaling rather than ABA metabolism is involved in trehalose-induced drought tolerance in tomato plants Planta 250, 643-655. DOI:10.1007/s00425-019-03195-2URL [本文引用: 3]
ZangBS, LiHW, LiWJ, DengXW, WangXP (2011). Analysis of trehalose-6-phosphate synthase (TPS) gene family suggests the formation of TPS complexes in rice Plant Mol Biol 76, 507-522. DOI:10.1007/s11103-011-9781-1URL [本文引用: 1]
ZhangC, LiX, HeYF, ZhangJF, YanT, LiuXL (2017). Physiological investigation of C4-phosphoenolpyruvate-carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance Plant Physiol Biochem 115, 328-342. DOI:10.1016/j.plaphy.2017.03.019URL [本文引用: 5]
ZhangC, PengX, GuoXF, TangGJ, SunFL, LiuSD, XiYJ (2018). Transcriptional and physiological data reveal the dehydration memory behavior in switchgrass ( Panicum virgatum L.) Biotechnol Biofuels 11, 91. DOI:10.1186/s13068-018-1088-xPMID:29619087 [本文引用: 1] Background: Switchgrass (Panicum virgatum L.) is a model biofuel plant because of its high biomass, cellulose-richness, easy degradation to ethanol, and the availability of extensive genomic information. However, a little is currently known about the molecular responses of switchgrass plants to dehydration stress, especially multiple dehydration stresses.& para;& para;Results: Studies on the transcriptional profiles of 35-day-old tissue culture plants revealed 741 dehydration memory genes. Gene Ontology and pathway analysis showed that these genes were enriched in phenylpropanoid biosynthesis, starch and sucrose metabolism, and plant hormone signal transduction. Further analysis of specific pathways combined with physiological data suggested that switchgrass improved its dehydration resistance by changing various aspects of its responses to secondary dehydration stress (D2), including the regulation of abscisic acid (ABA) and jasmonic acid (JA) biosynthesis and signal transduction, the biosynthesis of osmolytes (L-proline, stachyose and trehalose), energy metabolism (i.e., metabolic process relating to photosynthetic systems, glycolysis, and the TCA cycle), and lignin biosynthesis. The transcriptional data and chemical substance assays showed that ABA was significantly accumulated during both primary (D1) and secondary (D2) dehydration stresses, whereas JA accumulated during D1 but became significantly less abundant during D2. This suggests the existence of a complicated signaling network of plant hormones in response to repeated dehydration stresses. A homology analysis focusing on switchgrass, maize, and Arabidopsis revealed the conservation and species-specific distribution of dehydration memory genes.& para;& para;Conclusions: The molecular responses of switchgrass plants to successive dehydration stresses have been systematically characterized, revealing a previously unknown transcriptional memory behavior. These results provide new insights into the mechanisms of dehydration stress responses in plants. The genes and pathways identified in this study will be useful for the genetic improvement of switchgrass and other crops.
ZhaoRQ (2019). Expression, purification and characterization of the plant Snf1-related protein kinase 1 from Escherichia coli Protein Expr Purif 162, 24-31. DOI:10.1016/j.pep.2019.05.007URL [本文引用: 1]
ZhongRL, WangYX, GaiRN, XiDD, MaoCJ, FengM (2020). Rice SnRK protein kinase OsSAPK8 acts as a positive regulator in abiotic stress responses Plant Sci 292, 110373. DOI:10.1016/j.plantsci.2019.110373URL [本文引用: 1]
ZhuG, YeN, ZhangJ (2009). Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis Plant Cell Physiol 50, 644-651. DOI:10.1093/pcp/pcp022URL [本文引用: 1]
干旱条件下DCMU对高表达转C4-pepc水稻的花青素合成基因及其相关信号的影响 2 2018
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
转PEPC基因水稻对光氧化逆境的响应 1 2005
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
外源Ca 2+对PEG处理下转C4型PEPC基因水稻光合生理的调节 2 2015
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
脱落酸和己糖激酶抑制剂对高表达C4-PEPC转基因稻苗耐旱性的影响 4 2017
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
Roots of the resurrection plant tripogon loliiformis survive desiccation without the activation of autophagy pathways by maintaining energy reserves 1 2019
SnRK1 and trehalose 6-phosphate: two ancient pathways converge to regulate plant metabolism and growth 1 2020
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
Enhanced photosynthesis rate in genetically engineered indica rice expressing pepc gene cloned from maize 1 2007
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
Trehalose-6-phosphate, a new regulator of yeast glycolysis that inhibits hexokinases 1 1993
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors 2 2014
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Control of meristem determinacy by trehalose 6-phosphate phosphatases is uncoupled from enzymatic activity 1 2019
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
Peroxisomal plant metabolism: an update on nitric oxide, Ca 2+ and the NADPH recycling network 2 2018
... Ca2+和NO是调节植物耐旱性的重要第二信使分子(Corpas and Barroso, 2018; Liu et al., 2020).本研究表明, 两信号分子的含量在WT和PC间存在差异, 其中Ca2+水平在WT的各处理间基本保持稳定(图5A, B).相比CK, PC中DS处理下的Ca2+含量显著升高, 但PC显著低于WT (图5A).而DS+Tre处理进一步显著增加了Ca2+含量, 且PC显著高于WT.有意思的是, 与CK相比, DS处理显著下调了两试材的NO含量, 而DS+Tre处理仅提升了PC叶内的NO水平, 且PC中NO含量在CK+Tre、DS及DS+Tre处理下均显著高于WT (图5B).以上表明干旱条件下PC中的NO可能作为信号, 低水平时促进ABA的信号转导, 高水平时则起到反馈抑制作用.可见, 与DS处理相比, 海藻糖处理有利于启动信号分子参与干旱响应, 且较WT, PC的促进效应更显著. ...
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
Fine tuning of trehalose biosynthesis and hydrolysis as novel tools for the generation of abiotic stress tolerant plants 2 2014
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
Enhanced gene expression rather than natural polymorphism in coding sequence of the OsbZIP23 determines drought tolerance and yield improvement in rice genotypes 1 2016
... 已有研究表明, 水稻OsbZIP23和OsMYB2是响应ABA耐旱的转录因子(Yang et al., 2012; Dey et al., 2016), 而OsLEA3、OsNAC6、OsRab16b和OsRab21等作为靶基因(Hong et al., 2009; Duan and Cai, 2012; Hong et al., 2016)在下游发挥作用.为了进一步明确海藻糖与ABA在PC干旱调节中的机制, 本研究测定了ABA耐旱相关转录因子和靶基因的表达量(图4C).结果表明, 在PC中, DS处理上调了OsbZIP23的表达量; 相比DS, DS+Tre处理进一步上调了OsbZIP23和OsMYB2的转录水平, 且PC显著高于WT.同时, OsRab16b、OsRab21、OsLEA3和OsNAC6在PC叶内CK、DS和DS+Tre处理下的表达模式与OsbZIP23和OsMYB2基本保持一致, 且上述基因的表达水平在DS+Tre处理下均表现为PC显著高于WT (图4C; 附表1).综上, 与WT相比, 海藻糖可能通过诱导ABA信号转导途径上调与ABA相关的耐旱转录因子及其靶基因的表达水平, 进而增强PC的耐旱性. ...
Over-expression of phosphoenolpyruvate carboxylase cDNA from C4 millet ( Seteria italica) increase rice photosynthesis and yield under upland condition but not in wetland fields 1 2013
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
High light tolerance is enhanced by overexpressed PEPC in rice under drought stress 1 2012
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance 1 2012
... 已有研究表明, 水稻OsbZIP23和OsMYB2是响应ABA耐旱的转录因子(Yang et al., 2012; Dey et al., 2016), 而OsLEA3、OsNAC6、OsRab16b和OsRab21等作为靶基因(Hong et al., 2009; Duan and Cai, 2012; Hong et al., 2016)在下游发挥作用.为了进一步明确海藻糖与ABA在PC干旱调节中的机制, 本研究测定了ABA耐旱相关转录因子和靶基因的表达量(图4C).结果表明, 在PC中, DS处理上调了OsbZIP23的表达量; 相比DS, DS+Tre处理进一步上调了OsbZIP23和OsMYB2的转录水平, 且PC显著高于WT.同时, OsRab16b、OsRab21、OsLEA3和OsNAC6在PC叶内CK、DS和DS+Tre处理下的表达模式与OsbZIP23和OsMYB2基本保持一致, 且上述基因的表达水平在DS+Tre处理下均表现为PC显著高于WT (图4C; 附表1).综上, 与WT相比, 海藻糖可能通过诱导ABA信号转导途径上调与ABA相关的耐旱转录因子及其靶基因的表达水平, 进而增强PC的耐旱性. ...
Carbon source-sink relationship in Arabidopsis thaliana: the role of sucrose transporters 1 2018
... 植物中SnRK1可能在生长发育的不同阶段和不同组织中都需要被抑制和激活, 以适应其生长和发育进程(Zhao, 2019).已有研究表明, T6P和SnRK1的相互作用是调控细胞响应内源能量及碳状态的中枢信号途径(John et al., 2017).然而, 禾本科植物中T6P在调节SnRK1中具有双重功能, 并且二者在调节能量平衡时还涉及更多未知的潜在机制(Zhai et al., 2018).在水稻种子萌发和幼苗生长过程中, SnRK1是养分再动员和供应到生长库所必需的元件(Kretzschmar et al., 2015).代谢组学研究结果也表明, T6P是维持蔗糖水平(光合作用产物)的信号因子, 特别是在植物体内糖代谢和糖流入过程中起重要作用, 且蔗糖水平与T6P浓度的变化呈正相关(Figueroa et al., 2016).还有研究推测, T6P或海藻糖或海藻糖生物合成酶可能与植物激素以及糖诱导的信号通路相互作用(Durand et al., 2018).本研究也发现, 海藻糖联合干旱处理, PC中T6P合成相关基因的表达与WT差异明显, 而且T6P基因表达水平的提高伴随着与蔗糖含量和SnRK1相关基因的表达上调, 表明其可能通过蔗糖分子加速PC叶内T6P的清除, 进而动员海藻糖代谢途径响应胁迫.可见, 上述研究结果均可归因于蔗糖-T6P-SnRK1关系的高度动态性质, 以及每个成分参与独立的或相互关联的信号通路(Zhai et al., 2018). ...
On the road to C4 rice: advances and perspectives 1 2020
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
Effects of exogenous trehalose on the metabolism of sugar and abscisic acid in tomato seedlings under salt stress 3 2019
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
... ABA主要通过关闭叶片的气孔、减少蒸腾和保持植物体内的水分来提高植物的抗逆性(Feng et al., 2019).通过外源施加或过量表达基因来增加内源ABA含量, 可以上调ABA信号相关基因的表达, 从而对植物的抗逆性产生积极影响(Yu et al., 2019).在逆境条件下, 植物ABA生物合成和信号转导基因的普遍上调与海藻糖合成相关基因的上调协同变化(Zhang et al., 2018).SnRK2基因是ABA信号转导的关键成分, 并通过转录因子AREBs/ABFs的磷酸化, 对ABA诱导的ABA反应基因的激活起重要作用(Zhong et al., 2020), 其中OsSAPK8已被证明是干旱胁迫相关基因OsSLAC1的激活剂(Sun et al., 2016).海藻糖处理也显著上调了干旱胁迫下番茄(Solanum lycopersicum) SISnRK2.3和SISnRK2.4的转录水平, 减少失水, 增强耐旱能力(Yu et al., 2019).本研究表明, 与单独干旱胁迫相比, 海藻糖联合干旱处理显著上调了供试材料中与ABA相关的3个SnRK2基因(OsSAPK8、OsSAPK9和OsSAPK10)的表达, 并且选择性地促进ABA合成代谢相关基因的上调和ABA分解代谢相关基因的下调, 这些表现与其耐旱指标(叶片的RWC)表现一致, 而且PC的表达水平显著高于WT, 表明外施海藻糖处理可能增强了供试材料ABA依赖的SnRK2的干旱响应能力, 即增强ABA合成代谢相关基因的表达, 有利于正向激活ABA信号通路, 诱导bZIP和MYB转录因子, 进一步上调相关靶基因Rab、LEA和NAC的表达, 从而有利于增强PC的耐旱性. ...
The role of trehalose 6-phosphate (Tre6P) in plant metabolism and development 1 2021
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Trehalose 6-phosphate coordinates organic and amino acid metabolism with carbon availability 2 2016
... 植物中SnRK1可能在生长发育的不同阶段和不同组织中都需要被抑制和激活, 以适应其生长和发育进程(Zhao, 2019).已有研究表明, T6P和SnRK1的相互作用是调控细胞响应内源能量及碳状态的中枢信号途径(John et al., 2017).然而, 禾本科植物中T6P在调节SnRK1中具有双重功能, 并且二者在调节能量平衡时还涉及更多未知的潜在机制(Zhai et al., 2018).在水稻种子萌发和幼苗生长过程中, SnRK1是养分再动员和供应到生长库所必需的元件(Kretzschmar et al., 2015).代谢组学研究结果也表明, T6P是维持蔗糖水平(光合作用产物)的信号因子, 特别是在植物体内糖代谢和糖流入过程中起重要作用, 且蔗糖水平与T6P浓度的变化呈正相关(Figueroa et al., 2016).还有研究推测, T6P或海藻糖或海藻糖生物合成酶可能与植物激素以及糖诱导的信号通路相互作用(Durand et al., 2018).本研究也发现, 海藻糖联合干旱处理, PC中T6P合成相关基因的表达与WT差异明显, 而且T6P基因表达水平的提高伴随着与蔗糖含量和SnRK1相关基因的表达上调, 表明其可能通过蔗糖分子加速PC叶内T6P的清除, 进而动员海藻糖代谢途径响应胁迫.可见, 上述研究结果均可归因于蔗糖-T6P-SnRK1关系的高度动态性质, 以及每个成分参与独立的或相互关联的信号通路(Zhai et al., 2018). ...
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
A tale of two sugars: trehalose 6-phosphate and sucrose 2 2016
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes 3 2008
... 本研究表明, 在DS处理后, PC内源海藻糖含量与CK相比显著上升; 而DS+Tre处理进一步上调PC的海藻糖含量, 且PC显著高于WT (表3).是否外施海藻糖调节水稻内源海藻糖代谢? 而且为什么PC的含量高于WT? 已有研究表明, 海藻糖代谢相关基因在调控能量平衡中发挥作用(Ge et al., 2008; Zang et al., 2011; Islam et al., 2019).OsTre1是目前唯一已知参与调控海藻糖酶合成的基因, 海藻糖酶通过分解海藻糖降低植物中海藻糖的含量(Islam et al., 2019).因此, 本研究进一步分析了海藻糖合成相关基因(OsTPP1、OsTPP2、OsTPP3和OsTPP7)、海藻糖分解相关基因(OsTre1)以及海藻糖合成前体T6P合成相关基因(OsTPP1、OsTPS2和OsTPS8)的表达(图4A).与CK相比, OsTPP2、OsTPP3和OsTPP7在DS处理下表达量均显著上升, 且PC内的基因表达量均显著高于WT; 与DS相比, DS+Tre处理下两供试材料的OsTPP1和OsTPP3表达量进一步上调, 而OsTPP2和OsTPP7表达量下调, 但PC始终显著高于WT (附表1).在WT中, DS处理下OsTre1的表达量较CK处理显著上调; 与DS相比, DS+Tre处理在PC和WT中OsTre1的表达量均下调, 且在DS和DS+Tre处理下, PC始终低于WT且差异显著(附表1; 图4A).此外, 在DS处理下, PC中与海藻糖上游合成前体T6P相关的OsTPS2和OsTPS8表达量较CK均显著上升, DS+Tre处理使其表达量进一步上调, 且PC显著高于WT (附表1).但WT中上述基因在各处理下变幅不大.可见, 在干旱条件下, 海藻糖处理通过上调海藻糖合成以及下调其分解相关基因的转录水平来提高PC的内源海藻糖含量, 从而表现出较高的海藻糖水平. ...
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
... )的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Superoxide dismutases I: occurrence in higher plants 1 1977
The light-dependent transduction pathway controlling the regulatory phosphorylation of C4 phosphoenolpyruvate carboxylase in protoplasts from digitaria sanguinalis 1 1996
AtTPS1-mediated trehalose 6-phosphate synthesis is essential for embryogenic and vegetative growth and responsiveness to ABA in germinating seeds and stomatal guard cells 1 2010
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Interspecies and intraspecies analysis of trehalose contents and the biosynthesis pathway gene family reveals crucial roles of trehalose in osmotic-stress tolerance in cassava 1 2016
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Drought tolerance of transgenic rice overexpressing maize C4-PEPC gene related to increased anthocyanin synthesis regulated by sucrose and calcium 3 2020
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
NaCl-induced expression of glutathione reductase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid 1 2009
... 已有研究表明, 水稻OsbZIP23和OsMYB2是响应ABA耐旱的转录因子(Yang et al., 2012; Dey et al., 2016), 而OsLEA3、OsNAC6、OsRab16b和OsRab21等作为靶基因(Hong et al., 2009; Duan and Cai, 2012; Hong et al., 2016)在下游发挥作用.为了进一步明确海藻糖与ABA在PC干旱调节中的机制, 本研究测定了ABA耐旱相关转录因子和靶基因的表达量(图4C).结果表明, 在PC中, DS处理上调了OsbZIP23的表达量; 相比DS, DS+Tre处理进一步上调了OsbZIP23和OsMYB2的转录水平, 且PC显著高于WT.同时, OsRab16b、OsRab21、OsLEA3和OsNAC6在PC叶内CK、DS和DS+Tre处理下的表达模式与OsbZIP23和OsMYB2基本保持一致, 且上述基因的表达水平在DS+Tre处理下均表现为PC显著高于WT (图4C; 附表1).综上, 与WT相比, 海藻糖可能通过诱导ABA信号转导途径上调与ABA相关的耐旱转录因子及其靶基因的表达水平, 进而增强PC的耐旱性. ...
Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice 1 2016
... 已有研究表明, 水稻OsbZIP23和OsMYB2是响应ABA耐旱的转录因子(Yang et al., 2012; Dey et al., 2016), 而OsLEA3、OsNAC6、OsRab16b和OsRab21等作为靶基因(Hong et al., 2009; Duan and Cai, 2012; Hong et al., 2016)在下游发挥作用.为了进一步明确海藻糖与ABA在PC干旱调节中的机制, 本研究测定了ABA耐旱相关转录因子和靶基因的表达量(图4C).结果表明, 在PC中, DS处理上调了OsbZIP23的表达量; 相比DS, DS+Tre处理进一步上调了OsbZIP23和OsMYB2的转录水平, 且PC显著高于WT.同时, OsRab16b、OsRab21、OsLEA3和OsNAC6在PC叶内CK、DS和DS+Tre处理下的表达模式与OsbZIP23和OsMYB2基本保持一致, 且上述基因的表达水平在DS+Tre处理下均表现为PC显著高于WT (图4C; 附表1).综上, 与WT相比, 海藻糖可能通过诱导ABA信号转导途径上调与ABA相关的耐旱转录因子及其靶基因的表达水平, 进而增强PC的耐旱性. ...
Exogenous ATP enhance signal response of suspension cells of transgenic rice ( Oryza sativa L.) expressing maize C4-pepc encoded phosphoenolpyruvate carboxylase under PEG treatment 2 2017
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Contribution of trehalose biosynthetic pathway to drought stress tolerance of Capparis ovata desf 1 2015
The many-faceted function of phosphoenolpyruvate carboxylase in C3 plants 1 1983
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
ABA regulates subcellular redistribution of OsABI-LIKE2, a negative regulator in ABA signaling, to control root architecture and drought resistance in Oryza sativa 1 2015
... 已有研究表明, ABA分解代谢相关基因(OsABA8ox2和OsABA8ox3)和ABA合成代谢相关基因(OsNCED3 和OsNCED4)直接参与调控植株ABA含量(Zhu et al., 2009; Li et al., 2015).本研究发现, ABA分解代谢相关基因(OsABA8ox2和OsABA8ox3)在CK、DS和DS+Tre处理间均表现为PC显著低于WT (图4C; 附表1).在DS处理下, PC中OsABA8ox2和OsABA8ox3基因表达量较CK处理显著下降, DS+Tre处理后进一步下调; 而WT中OsABA8ox3在DS处理下上调, 在DS+Tre处理下虽然较DS显著下调, 但仍高于CK的表达水平.ABA合成代谢相关基因(OsNCED3和OsNCED4)在各处理间均表现为PC高于WT.在PC中, 相比CK, OsNCED3和OsNCED4的表达水平在DS处理下上调, DS+Tre处理后进一步上调; 而WT内各处理中其表达水平变幅不大(图4C).可见, 海藻糖处理选择性促进了干旱条件下PC内ABA合成相关基因的上调, 表明ABA代谢途径可能参与海藻糖增强PC的耐旱性机制. ...
Effects of 1-butanol, neomycin and calcium on the photosynthetic characteristics of pepc transgenic rice 2 2011
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Sugar starvation enhances leaf senescence and genes involved in sugar signaling pathways regulate early leaf senescence in mutant rice 1 2020
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
H2O2 and NO are involved in trehalose-regulated oxidative stress tolerance in cold-stressed tomato plants 2 2020
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
... Ca2+和NO是调节植物耐旱性的重要第二信使分子(Corpas and Barroso, 2018; Liu et al., 2020).本研究表明, 两信号分子的含量在WT和PC间存在差异, 其中Ca2+水平在WT的各处理间基本保持稳定(图5A, B).相比CK, PC中DS处理下的Ca2+含量显著升高, 但PC显著低于WT (图5A).而DS+Tre处理进一步显著增加了Ca2+含量, 且PC显著高于WT.有意思的是, 与CK相比, DS处理显著下调了两试材的NO含量, 而DS+Tre处理仅提升了PC叶内的NO水平, 且PC中NO含量在CK+Tre、DS及DS+Tre处理下均显著高于WT (图5B).以上表明干旱条件下PC中的NO可能作为信号, 低水平时促进ABA的信号转导, 高水平时则起到反馈抑制作用.可见, 与DS处理相比, 海藻糖处理有利于启动信号分子参与干旱响应, 且较WT, PC的促进效应更显著. ...
Improved short-term drought response of transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase via calcium signal cascade 3 2017a
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Phosphoenolpyruvate carboxylase regulation in C4-PEPC expressing transgenic rice during early responses to drought stress 2 2017b
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Analysis of relative gene expression data using real-time quantitative PCR 1 2002
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Identification and expression analysis of two inorganic C- and N- responsive genes encoding novel and distinct molecular forms of eukaryotic phosphoenolpyruvate carboxylase in the green microalga Chlamydomonas reinhardtii 1 2005
ROS-related redox regulation and signaling in plants 1 2018
... 干旱抑制植物光合作用过程可能是由于氧化胁迫引起的光氧化, 破坏了光合器官, 导致光合碳同化降低(Theerakulp and Phongngarm, 2013; Samadi et al., 2019).已有研究表明, 植物膜脂质过氧化是由暴露在胁迫下的活性氧过度积累所造成的次级伤害(Uzilday et al., 2014).然而, 活性氧也是双刃剑, 即在适度的活性氧水平下可将胁迫信号传递到下游信号通路, 并与蛋白激酶和转录因子协同作用, 最终调控植物的新陈代谢、发育和抗逆相关基因的表达(Noctor et al., 2018).有研究显示, 海藻糖诱导的抗氧化酶活性增加可以抑制H2O2的积累, 从而降低对氧化胁迫的敏感性, 保护光合结构和细胞膜免受氧化损伤(Yu et al., 2019).与C3植物相比, 类C4和C4植物对PEG诱导的渗透胁迫具有更强的耐受性, 且更能防止氧化胁迫造成的伤害(Uzilday et al., 2014).本研究表明, 海藻糖处理可以降低PC的膜脂过氧化程度, 提高光合能力, 增强耐旱性, 暗示高表达转玉米C4型PEPC基因水稻赋予了比非转基因的WT更强的PEG处理耐受性. ...
Drought resistance in rice from conventional to molecular breeding: a review 1 2019
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
Functional identification of a trehalose 6-phosphate phosphatase gene that is involved in transient induction of trehalose biosynthesis during chilling stress in rice 1 2005
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Enhanced drought tolerance in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene via NO and Ca 2+ 2 2015
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Improved oxidative tolerance in suspension-cultured cells of C4-pepc transgenic rice by H2O2 and Ca 2+ under PEG-6000 2 2015
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Hydrogen peroxide regulated photosynthesis in C4-pepc transgenic rice 2 2014
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Mitigation of drought stress on fenugreek plant by foliar application of trehalose 1 2016
Foliar-applied trehalose modulates growth, mineral nutrition, photosynthetic ability, and oxidative defense system of rice (Oryza sativa L.) under saline stress 1 2017
... 糖水平与细胞的能量状态直接相关, 糖既是中间新陈代谢的底物, 也是将碳代谢与植物生长发育联系起来的信号分子(Li et al., 2020).植物中海藻糖(trehalose, Tre)是参与糖信号传递的3种主要糖之一, 由海藻糖-6-磷酸磷酸酯酶(trehalose-6-phosphate phosphatase, TPP)磷酸化海藻糖-6-磷酸(trehalose-6-phosphate, T6P)生成, 其含量极低(Lunn et al., 2014).已发现外源施加海藻糖能提高多种植物对环境胁迫的耐受性, 且主要通过增强光合能力和抗氧化防护能力以及诱导信号分子(如H2O2和NO)等调节抗逆相关基因的表达来实现(Alam et al., 2014; Shahbaz et al., 2017; Feng et al., 2019; Kosar et al., 2019).此外, 海藻糖代谢还在调控葡萄糖信号转导、蔗糖代谢和淀粉合成中发挥重要作用(Akram et al., 2016).SnRKs是植物中的一类糖信号相关蛋白激酶, 包括3个亚族(SnRK1s、SnRK2s和SnRK3s), 是联系糖信号与胁迫响应之间的枢纽(张金飞等, 2017).海藻糖可通过修饰蛋白激酶与14-3-3蛋白互作(Delorge et al., 2014), 参与T6P/SnRK1介导的糖信号级联放大(John et al., 2017), 控制源库能量平衡.然而, 禾本科植物中T6P与SnRK1之间涉及更多额外的信号转导, 如己糖激酶(hexokinase, HXK)途径(Baena- González and John, 2020)、脱落酸(abscisic acid, ...
Phospho enol pyruvate carboxylase in Arabidopsis leaves plays a crucial role in carbon and nitrogen metabolism 1 2015
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
The energy-signaling hub SnRK1 is important for sucrose-induced hypocotyl elongation 2 2018
Enhanced photorespiration in transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase gene contributes to alleviating low nitrogen stress 1 2018
... 水稻(Oryza sativa)是世界上重要的粮食作物, 也是较易受干旱影响的植物之一(Oladosu et al., 2019).干旱逆境是全球性普遍存在且不断加剧的环境问题, 干旱胁迫导致水稻减产, 造成严重的粮食生产损失, 威胁世界粮食安全.目前, 在充分利用传统作物育种技术的基础上, 要想实现产量潜力的显著提升, 改善光合作用成为今后最值得研究的方向之一(Morales et al., 2020).C4植物较水稻等C3植物更耐旱, 在干旱条件下具有明显的生长及产量优势, 主要表现为光合能力、能量及水分利用率较高(Ermakova et al., 2020).磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是参与C4植物光合作用的关键酶, 可为三羧酸循环(tricarboxylic acid cycle, TCA)提供草酰乙酸(oxaloacetic acid, OAA), 并参与调节植物的碳、氮代谢和气孔运动, 尤其在耐旱机制中发挥重要作用(Karki et al., 2013; Shi et al., 2015).现已通过基因工程技术获得了高表达转玉米(Zea mays) C4型PEPC基因水稻(Ku et al., 1999) (以下简称PC).已有研究表明, PC水稻不仅光合能力增强、产量提高, 且表现出耐高光强、耐光氧化、耐旱及耐低氮等特性(Jiao et al., 2003; 李霞等, 2005; Bandyopadhyay et al., 2007; Ding et al., 2012, 2013; Liu et al., 2017a; Tang et al., 2018), 深入研究该材料的耐旱机制, 将为抵御水稻干旱胁迫提供有效手段. ...
Alleviation of adverse effects of salt stress on rice seedlings by exogenous trehalose 1 2013
... 干旱抑制植物光合作用过程可能是由于氧化胁迫引起的光氧化, 破坏了光合器官, 导致光合碳同化降低(Theerakulp and Phongngarm, 2013; Samadi et al., 2019).已有研究表明, 植物膜脂质过氧化是由暴露在胁迫下的活性氧过度积累所造成的次级伤害(Uzilday et al., 2014).然而, 活性氧也是双刃剑, 即在适度的活性氧水平下可将胁迫信号传递到下游信号通路, 并与蛋白激酶和转录因子协同作用, 最终调控植物的新陈代谢、发育和抗逆相关基因的表达(Noctor et al., 2018).有研究显示, 海藻糖诱导的抗氧化酶活性增加可以抑制H2O2的积累, 从而降低对氧化胁迫的敏感性, 保护光合结构和细胞膜免受氧化损伤(Yu et al., 2019).与C3植物相比, 类C4和C4植物对PEG诱导的渗透胁迫具有更强的耐受性, 且更能防止氧化胁迫造成的伤害(Uzilday et al., 2014).本研究表明, 海藻糖处理可以降低PC的膜脂过氧化程度, 提高光合能力, 增强耐旱性, 暗示高表达转玉米C4型PEPC基因水稻赋予了比非转基因的WT更强的PEG处理耐受性. ...
The trehalose-6-phosphate synthase TPS5 negatively regulates ABA signaling in Arabidopsis thaliana 2 2019
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
Strategies of ROS regulation and antioxidant defense during transition from C3 to C4 photosynthesis in the genus Flaveria under PEG-induced osmotic stress 3 2014
OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition 1 2019
... 光合作用的改善与内源糖信号网络的相互作用有关(Feng et al., 2019).蔗糖是参与光合作用的主要糖类, 己糖是海藻糖的主要合成底物(Wingler, 2018), 而海藻糖对于维持植物的生长以及对逆境的响应至关重要(Lunn et al., 2014).本研究表明, 海藻糖联合干旱处理使水稻叶片内的蔗糖和海藻糖含量进一步增加, 葡萄糖含量降低(表3), 其中PC内蔗糖和海藻糖含量显著高于WT, 而葡萄糖含量显著低于WT, 这暗示PC具有较WT更强的光合能力与内源糖组分的差异有关.有研究表明, 植物中具有C4型PEPC基因有利于更多光合产物的积累, 同时由于较强的源库活动而对蔗糖积累可能出现的反馈抑制表现出较低的敏感性(Kim et al., 2013).海藻糖-6-磷酸合酶(trehalose-6-phosphate synthase, TPS)催化葡萄糖从UDP-葡萄糖到葡萄糖6磷酸酯的转移以形成T6P, 随后T6P被TPP磷酸化为海藻糖(Fichtner and Lunn, 2021).本研究表明, 相比WT, PC一方面在更大程度上维持了其功能叶片光合性能的稳定, 同时也因此积累了相对较多的可溶性糖(蔗糖), 进而增强其耐旱性; 另一方面, PC可通过消耗更多的葡萄糖作为底物进而合成海藻糖, 也有利于植株提高抵御胁迫的能力.已有研究表明, 己糖积累可导致光合作用相关基因表达下调(Wingler, 2018).植物细胞中HXK是葡萄糖信号的感受器, HXK可被T6P抑制(Blázquez et al., 1993).HXK作为关键信号元件在PC中维持很低的水平, 尤其是外施海藻糖后其在干旱胁迫下的活性伴随葡萄糖含量而降低, 暗示海藻糖也可能通过下调HXK途径参与干旱响应.已有研究表明, 转化海藻糖生物合成基因的转基因植物其内源海藻糖含量显著增加, 并表现出更好的抗逆性(Ge et al., 2008; Han et al., 2016; Kosar et al., 2019).需要特别指出的是, OsTPP1和OsTPP2可以编码功能性TPP, 与海藻糖含量直接相关, 且OsTPP1与OsTPP2在胁迫下协同调控TPP酶的合成(Pramanik and Imai, 2005), 而OsTPP3和OsTPP7作为II类TPP基因对海藻糖含量起主要调节作用(Kretzschmar et al., 2015; Jiang et al., 2019).水稻OsTPP1、OsTPP2和OsTPP3通过调控ABA合成(OsNCED3和OsNCED4)和信号转导基因(OsRab16)的表达来响应胁迫, 增强抗性(Ge et al., 2008; Jiang et al., 2019).拟南芥(Arabidopsis thaliana) TPS1基因产物在调节营养以及胚发生组织的生长中发挥至关重要的作用, 其机制涉及ABA和糖代谢(Gómez et al., 2010).OsTPS8可能通过ABA信号转导调节水稻中的木质素沉积和海藻糖积累, 从而赋予水稻耐盐性(Vishal et al., 2019).本研究通过外施海藻糖联合干旱处理, 发现PC叶片中OsTPP1和OsTPP2的总表达量显著升高, 表明外施海藻糖可通过增强内源海藻糖代谢途径促进PC内源海藻糖积累(表3), 同时发现OsTPP3和OsTPS8基因的表达显著增强(图4A), 推测海藻糖可能通过调控ABA信号通路抵御胁迫.而OsTPP7在海藻糖代谢中对海藻糖含量起调节作用, 它与T6P/蔗糖和海藻糖含量呈负相关, 即当能量较少时OsTPP7的上调可以促进淀粉酶活化, 调控源库能量平衡(Kretzschmar et al., 2015).本研究还发现OsTPP7的表达下降(图4A), 这与其内源海藻糖含量的增加相反(表3), 推测其在PC的糖代谢过程中可能具有关键的平衡调节功能, 相关推测还需要深入研究加以验证. ...
Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins 2 2015
... ABA)介导的气孔导度调节(Wang et al., 2015)以及与NO相关的硝酸还原酶(nitrate reductase, NR)和SnRK2s的共价修饰(Tian et al., 2019), 且依赖于第二信使分子H2O2和NO (Claeys et al., 2019; Liu et al., 2020).值得注意的是, T6P途径也是激活PEPC活性的潜在机制之一(Figueroa and Lunn, 2016).前期研究发现, HXK和ABA均参与PC的干旱响应(吴敏怡等, 2017).海藻糖是否在PC的耐旱机制中发挥作用是一个值得深入研究的科学问题.本研究通过外源引入海藻糖, 解析光合、抗氧化酶系统以及相关的信号转导途径参与PC的干旱响应过程, 探明海藻糖代谢与糖信号相关基因SnRKs、ABA信号以及下游干旱响应基因的变化特点, 旨在从海藻糖代谢和信号转导的角度解析PC响应干旱的作用机制, 丰富海藻糖在水稻干旱响应中的相关信息, 并为“C4稻”在干旱胁迫中的应用提供理论依据. ...
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
Trehalose-6-phosphate phosphatase E modulates ABA-controlled root growth and stomatal movement inArabidopsis 1 2020
... 植物抗逆调节机制中, 海藻糖与信号分子Ca2+和NO以及激素等密切相关(Delorge et al., 2014; Corpas and Barroso, 2018; Wang et al., 2020a).ABA可快速上调SnRK2基因来磷酸化下游效应蛋白, 响应胁迫(Wang et al., 2015); 同时激活的SnRK2可促进T6P相关基因的表达, 通过NR亚硝基化积累NO, 但当NO达到较高水平时将会对ABA信号途径起负反馈抑制效应(Tian et al., 2019).本研究也发现, 外源海藻糖联合干旱处理可在一定程度上促进Ca2+和NO的产生, 而且NO含量的增加与其主要产生途径硝酸还原酶活性的增强同步, 表明PC中第二信使分子(NO和Ca2+)可能参与了海藻糖代谢途径对糖信号及ABA信号响应干旱的调控.在C3植物中, PEPC对光合作用几乎不起作用, 为非光合型PEPC, 且相比C4植物中的光合型PEPC, 其酶活性非常低(Latzko and, 1983).大多数PEPC是变构酶, 其活性受多个新陈代谢效应子的调控(Izui et al., 2004).本研究表明, PC和WT在海藻糖处理后可以增强其干旱条件下C3型PEPC (Osppc2a)基因的转录水平.但相比PC, WT的PEPC活性变幅很小, 说明在PC中PEPC活性的增强主要归因于C4型PEPC基因的高表达, 且这部分PEPC为光合型PEPC, 是PSII光合性能增强的关键.有研究表明, T6P可部分通过NR激活PEPC翻译水平来调节光同化物的分配(Figueroa et al., 2016), 且HXK可负调节PEPC酶活(吴敏怡等, 2017).与之相对应, 海藻糖联合干旱处理后PC也表现PEPC活性的增强伴随NR的上调和HXK的下调, 表明PC也可能通过下调HXK途径调节PEPC活性, 但具体机制尚待深入研究. ...
AKINβ1, a subunit of SnRK1, regulates organic acid metabolism and acts as a global modulator of genes involved in carbon, lipid, and nitrogen metabolism 1 2020
Physiological investigation of C4-phosphoenolpyruvate-carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance 5 2017
... 近年来, 关于PEPC在植物逆境耐性中的分子机制已成为新的研究热点, 而对PC水稻耐旱机制的研究, 也为深入了解该酶的生物学功能提供了新线索.已有研究表明, 干旱使PC触发第二信使, 如过氧化氢(hydrogen peroxide, H2O2) (Ren et al., 2014)、一氧化氮(nitric oxide, NO) (Chen et al., 2014)、钙离子(calcium ion, Ca2+) (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)和磷脂酸(Li et al., 2011), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9以及蔗糖非发酵1 (sucrose nonfermenting-1, SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinase, SnRKs)亚家族中SnRK1s基因(OsSnRK1a、OsSnRK24和OsSnRK35)激活级联反应, 诱导靶基因C4-PEPC的转录与翻译, 积累渗透调节物质并维持稳定的光合作用, 进而抵御胁迫(刘小龙等, 2015; Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... ).此外, PC响应胁迫的调节过程都与其内源糖水平的差异密切相关(Zhang et al., 2017).PC内源的高蔗糖含量通过调节花青素及合成相关基因的表达, 增强花青素代谢而表现耐旱(何亚飞等, 2018).外源葡萄糖可通过调控内源糖代谢激活内源SnRK3s基因(OsSnRK3.1和OsSnRK3.4)与钙调蛋白基因(CBL)在PC耐旱机制中发挥作用(张金飞等, 2018).PC中内源蔗糖和葡萄糖与SnRK2s基因表达密切相关, 参与花青素代谢的调节(He et al., 2020), 表明糖信号在PC干旱响应中发挥重要作用.然而, 内源糖组分是多元的, 且彼此间涉及复杂的信号网络, 需要更多深入的研究. ...
... 磷酸烯醇式丙酮酸羧化酶不仅是植物碳固定和碳回补的主导物, 而且在保卫细胞中也作为一种胞质酶, 主要通过提供苹果酸作为反离子和渗透调节器参与植物的耐逆调节(Akram et al., 2016).作为具有耐旱特性的高表达转玉米C4型PEPC基因水稻, PC前期已有系列研究(严婷等, 2019), 表明干旱会使PC触发第二信使, 如H2O2 (Ren et al., 2014)、NO (Chen et al., 2014)、Ca2+ (Qian et al., 2015a, 2015b; Liu et al., 2017a, 2017b)、磷脂酸(Li et al., 2011)和糖(Zhang et al., 2017), 通过调控钙依赖和糖信号激酶相关基因, 如CPK4、CPK9 (刘小龙等, 2015)以及SnRKs基因激活级联反应, 诱导外源C4-PEPC的转录与翻译, 促进渗透调节物质的积累, 并维持稳定的光合作用, 进而表现耐旱(Huo et al., 2017; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
... ; Zhang et al., 2017; 张金飞等, 2018; He et al., 2020).本实验室前期的研究表明, 糖是水稻生长发育和基因表达的重要调节因子, 它不仅影响PC代谢途径, 而且在信号转导中具有初级信使作用(吴敏怡等, 2017; 何亚飞等, 2018; 张金飞等, 2018; He et al., 2020).本研究进一步表明, 在干旱条件下, 海藻糖处理使PC一方面通过蔗糖含量增加激活SnRK1, 加速动员海藻糖代谢, 促进内源海藻糖合成, 从而提高抗氧化酶活性, 降低膜脂过氧化作用, 缓解对光系统的伤害; 另一方面提高蔗糖含量, 还有利于上调NR活性并下调HXK活性, 进而提高PEPC的转录与翻译水平, 缓解PSII的光合性能损伤.此外, PC还可能通过诱导NO和钙离子, 提高ABA依赖的SnRK2基因的转录水平, 增强ABA代谢及其信号转导, 激活下游干旱相关靶基因的表达, 维持植物正常的生长发育, 增强PC的耐旱性. ...
Transcriptional and physiological data reveal the dehydration memory behavior in switchgrass ( Panicum virgatum L.) 1 2018
... ABA主要通过关闭叶片的气孔、减少蒸腾和保持植物体内的水分来提高植物的抗逆性(Feng et al., 2019).通过外源施加或过量表达基因来增加内源ABA含量, 可以上调ABA信号相关基因的表达, 从而对植物的抗逆性产生积极影响(Yu et al., 2019).在逆境条件下, 植物ABA生物合成和信号转导基因的普遍上调与海藻糖合成相关基因的上调协同变化(Zhang et al., 2018).SnRK2基因是ABA信号转导的关键成分, 并通过转录因子AREBs/ABFs的磷酸化, 对ABA诱导的ABA反应基因的激活起重要作用(Zhong et al., 2020), 其中OsSAPK8已被证明是干旱胁迫相关基因OsSLAC1的激活剂(Sun et al., 2016).海藻糖处理也显著上调了干旱胁迫下番茄(Solanum lycopersicum) SISnRK2.3和SISnRK2.4的转录水平, 减少失水, 增强耐旱能力(Yu et al., 2019).本研究表明, 与单独干旱胁迫相比, 海藻糖联合干旱处理显著上调了供试材料中与ABA相关的3个SnRK2基因(OsSAPK8、OsSAPK9和OsSAPK10)的表达, 并且选择性地促进ABA合成代谢相关基因的上调和ABA分解代谢相关基因的下调, 这些表现与其耐旱指标(叶片的RWC)表现一致, 而且PC的表达水平显著高于WT, 表明外施海藻糖处理可能增强了供试材料ABA依赖的SnRK2的干旱响应能力, 即增强ABA合成代谢相关基因的表达, 有利于正向激活ABA信号通路, 诱导bZIP和MYB转录因子, 进一步上调相关靶基因Rab、LEA和NAC的表达, 从而有利于增强PC的耐旱性. ...
Expression, purification and characterization of the plant Snf1-related protein kinase 1 from Escherichia coli 1 2019
... 植物中SnRK1可能在生长发育的不同阶段和不同组织中都需要被抑制和激活, 以适应其生长和发育进程(Zhao, 2019).已有研究表明, T6P和SnRK1的相互作用是调控细胞响应内源能量及碳状态的中枢信号途径(John et al., 2017).然而, 禾本科植物中T6P在调节SnRK1中具有双重功能, 并且二者在调节能量平衡时还涉及更多未知的潜在机制(Zhai et al., 2018).在水稻种子萌发和幼苗生长过程中, SnRK1是养分再动员和供应到生长库所必需的元件(Kretzschmar et al., 2015).代谢组学研究结果也表明, T6P是维持蔗糖水平(光合作用产物)的信号因子, 特别是在植物体内糖代谢和糖流入过程中起重要作用, 且蔗糖水平与T6P浓度的变化呈正相关(Figueroa et al., 2016).还有研究推测, T6P或海藻糖或海藻糖生物合成酶可能与植物激素以及糖诱导的信号通路相互作用(Durand et al., 2018).本研究也发现, 海藻糖联合干旱处理, PC中T6P合成相关基因的表达与WT差异明显, 而且T6P基因表达水平的提高伴随着与蔗糖含量和SnRK1相关基因的表达上调, 表明其可能通过蔗糖分子加速PC叶内T6P的清除, 进而动员海藻糖代谢途径响应胁迫.可见, 上述研究结果均可归因于蔗糖-T6P-SnRK1关系的高度动态性质, 以及每个成分参与独立的或相互关联的信号通路(Zhai et al., 2018). ...
Rice SnRK protein kinase OsSAPK8 acts as a positive regulator in abiotic stress responses 1 2020
... ABA主要通过关闭叶片的气孔、减少蒸腾和保持植物体内的水分来提高植物的抗逆性(Feng et al., 2019).通过外源施加或过量表达基因来增加内源ABA含量, 可以上调ABA信号相关基因的表达, 从而对植物的抗逆性产生积极影响(Yu et al., 2019).在逆境条件下, 植物ABA生物合成和信号转导基因的普遍上调与海藻糖合成相关基因的上调协同变化(Zhang et al., 2018).SnRK2基因是ABA信号转导的关键成分, 并通过转录因子AREBs/ABFs的磷酸化, 对ABA诱导的ABA反应基因的激活起重要作用(Zhong et al., 2020), 其中OsSAPK8已被证明是干旱胁迫相关基因OsSLAC1的激活剂(Sun et al., 2016).海藻糖处理也显著上调了干旱胁迫下番茄(Solanum lycopersicum) SISnRK2.3和SISnRK2.4的转录水平, 减少失水, 增强耐旱能力(Yu et al., 2019).本研究表明, 与单独干旱胁迫相比, 海藻糖联合干旱处理显著上调了供试材料中与ABA相关的3个SnRK2基因(OsSAPK8、OsSAPK9和OsSAPK10)的表达, 并且选择性地促进ABA合成代谢相关基因的上调和ABA分解代谢相关基因的下调, 这些表现与其耐旱指标(叶片的RWC)表现一致, 而且PC的表达水平显著高于WT, 表明外施海藻糖处理可能增强了供试材料ABA依赖的SnRK2的干旱响应能力, 即增强ABA合成代谢相关基因的表达, 有利于正向激活ABA信号通路, 诱导bZIP和MYB转录因子, 进一步上调相关靶基因Rab、LEA和NAC的表达, 从而有利于增强PC的耐旱性. ...
Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis 1 2009
... 已有研究表明, ABA分解代谢相关基因(OsABA8ox2和OsABA8ox3)和ABA合成代谢相关基因(OsNCED3 和OsNCED4)直接参与调控植株ABA含量(Zhu et al., 2009; Li et al., 2015).本研究发现, ABA分解代谢相关基因(OsABA8ox2和OsABA8ox3)在CK、DS和DS+Tre处理间均表现为PC显著低于WT (图4C; 附表1).在DS处理下, PC中OsABA8ox2和OsABA8ox3基因表达量较CK处理显著下降, DS+Tre处理后进一步下调; 而WT中OsABA8ox3在DS处理下上调, 在DS+Tre处理下虽然较DS显著下调, 但仍高于CK的表达水平.ABA合成代谢相关基因(OsNCED3和OsNCED4)在各处理间均表现为PC高于WT.在PC中, 相比CK, OsNCED3和OsNCED4的表达水平在DS处理下上调, DS+Tre处理后进一步上调; 而WT内各处理中其表达水平变幅不大(图4C).可见, 海藻糖处理选择性促进了干旱条件下PC内ABA合成相关基因的上调, 表明ABA代谢途径可能参与海藻糖增强PC的耐旱性机制. ...
 ,1,2,3,*, 谢寅峰1
,1,2,3,*, 谢寅峰1
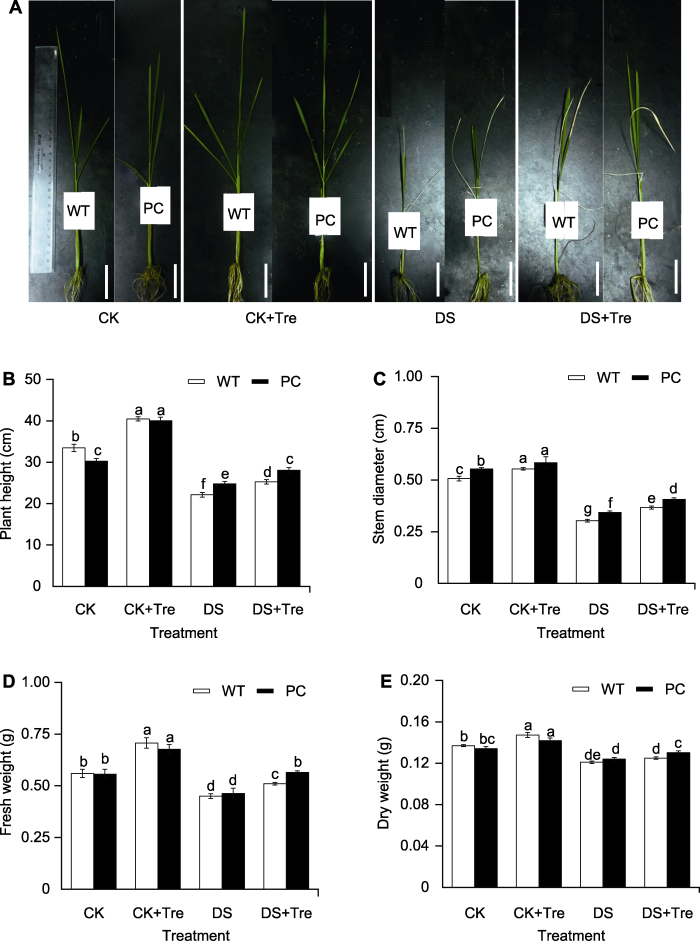 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT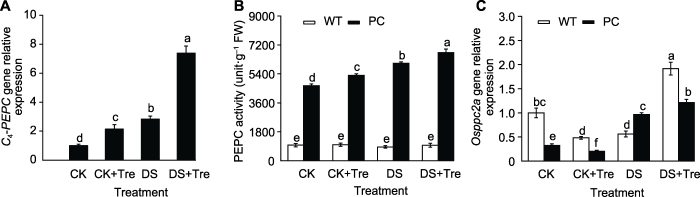 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT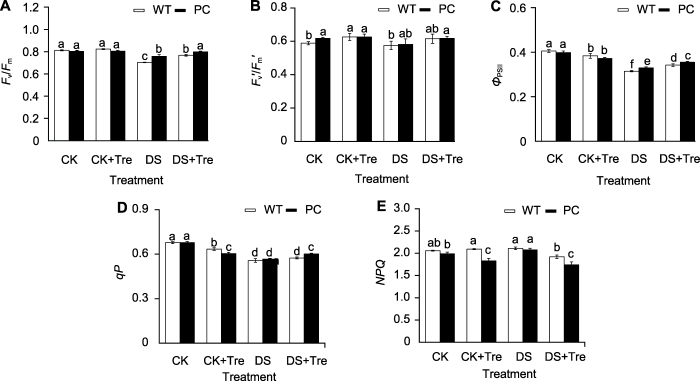 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT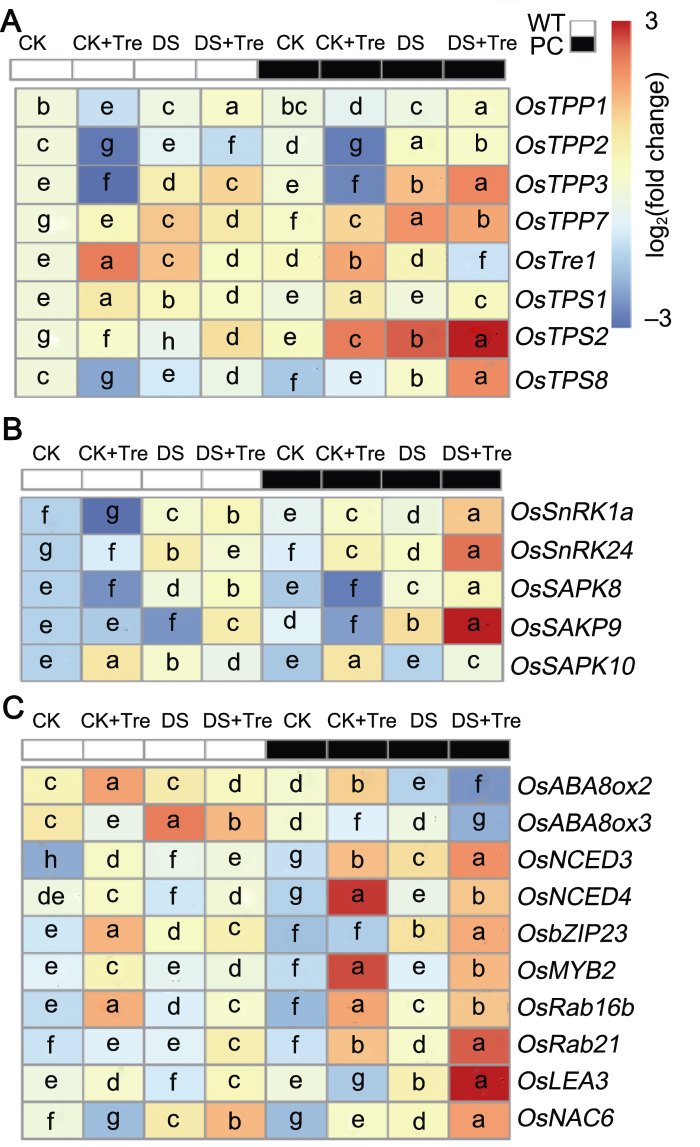 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT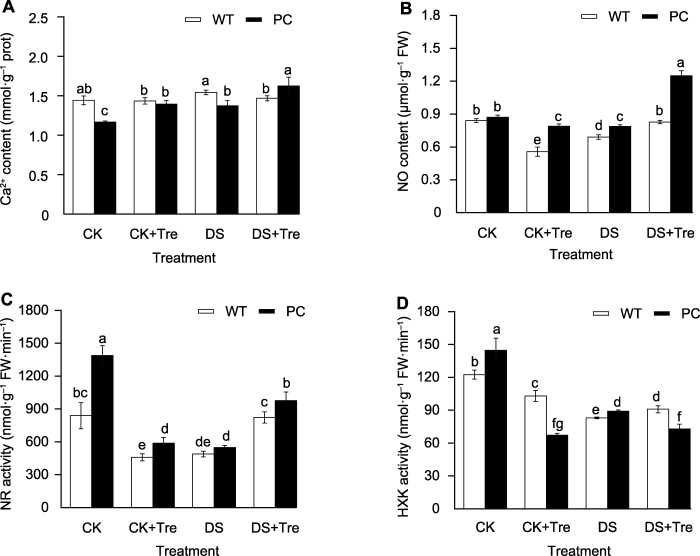 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}