罗虹, 温小蕙, 周圆圆, 戴思兰
, * 北京林业大学园林学院, 国家花卉工程技术研究中心, 花卉种质创新与分子育种北京市重点实验室, 城乡生态环境北京实验室, 北京 100083
Establishment of In Vitro Regeneration System of Helenium aromaticum Hong Luo, Xiaohui Wen, Yuanyuan Zhou, Silan Dai
, * Laboratory of Urban and Rural Ecological Environment, Beijing Key Laboratory of Flower Germplasm Innovation and Molecular Breeding, National Flower Engineering Technology Research Center, College of Landscape Architecture, Beijing Forestry University, Beijing 100083, China
通讯作者: *E-mail: silandai@sina.com 责任编辑: 白羽红
收稿日期: 2019-10-28
接受日期: 2020-02-26
网络出版日期: 2020-05-01
基金资助: 国家自然科学基金 (31530064 )国家自然科学基金 (314719072 )国家重点研发计划 (2018YFD1000405 )北京市科技计划 (Z191100008519002 )
Corresponding authors: E-mail: silandai@sina.com Received: 2019-10-28
Accepted: 2020-02-26
Online: 2020-05-01
摘要 芳香堆心菊(Helenium aromaticum )全株具芳香气味, 且头状花序仅含管状花, 是研究菊科植物花香和花型的良好材料, 但目前尚缺乏对其转基因技术体系的研究。为建立高效的芳香堆心菊离体再生体系, 以叶片、茎段和下胚轴为外植体, 进行25组不同激素及不同浓度配比的不定芽诱导研究。结果表明, 以芳香堆心菊叶片为外植体, 培养基为MS+ 0.2 mg·L-1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ, 培养20天后愈伤组织诱导率高达100%, 丛生芽的诱导率为62.10%; 将不定芽接种于1/2MS培养基中进行生根培养, 16天即可生根, 且生根率为63.33%; 生根后继续培养14天现蕾, 开花率达93.33%。此外, 研究表明芳香堆心菊的再生受外植体来源、激素种类和浓度的影响。2,4-D不利于芳香堆心菊不定芽的诱导, 适宜浓度的6-BA和TDZ组合能有效促进芳香堆心菊不定芽的形成。研究初步建立了芳香堆心菊组织培养条件下的离体再生体系, 为建立其遗传转化体系奠定了坚实的基础。研究结果还可用于后续有关菊科植物花香和花型的研究。 关键词: 芳香堆心菊 ;
再生体系 ;
组织培养 Abstract Helenium aromaticum has a fragrant odor and its capitulum contains only disk flowers. H . aromaticum is a good model for studying development of floral patterns in Compositae. However, methods for genetic transformation of H . aromaticum is currently not available. In this study, we report the establishment of an efficient in vitro regeneration system of the H. aromaticum. The induction of adventitious buds was tested with various combinations and concentrations of 25 different phytohormones by using leaves, stem segments, and hypocotyls as explants. We found that when leaf explants were cultured on MS medium supplemented with 0.2 mg·L-1 NAA, 1 mg·L-1 6-BA, and 0.2 mg·L-1 TDZ for 20 days, the induction of callus was 100% and the induction rate of adventitious buds was 62.10%. The newly-formed adventitious buds were then cultured on 1/2MS medium for 16 days and the rooting rate reached 63.33%. After culturing for an additional 14 days, floral buds initiated and eventually flowering with a rate of 93.33%. In addition, we also found that the regeneration of H. aromaticum was affected by the nature of explants, the types and concentrations of phytohormones. Whereas 2,4-D is incapable of inducing adventitious buds, the combination of appropriate concentrations of 6-BA and TDZ effectively promotes the formation of adventitious buds of H. aromaticum . This study has established an in vitro regeneration system of H. aromaticum , which is a key perquisite for the subsequent establishment of its genetic transformation system. Moreover, this method will also be an important reference for studies on the floral development patterns in Compositae. Keywords: Helenium aromaticum ;
regeneration system ;
tissue culture PDF (5700KB) 摘要页面 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 罗虹, 温小蕙, 周圆圆, 戴思兰 . 芳香堆心菊离体再生体系的建立. 植物学报, 2020,
55 (3): 318-328 doi:10.11983/CBB19211
Luo Hong, Wen Xiaohui, Zhou Yuanyuan, Dai Silan .
Establishment of In Vitro Regeneration System of Helenium aromaticum .
Chinese Bulletin of Botany , 2020,
55 (3): 318-328 doi:10.11983/CBB19211
芳香堆心菊(
Helenium aromaticum )为菊科(Compositae)堆心菊属草本花卉, 原产于智利, 自然条件下12月份成花, 耐寒性较差。芳香堆心菊花型独特, 头状花序上仅着生管状花, 且全株具芳香气味(
Gómez-González et al., 2016 )。此外, 芳香堆心菊含有大量对人体健康有益的药用化学物质, 具有极高的药用及观赏价值(
Bloszyk et al., 1975 ;
Bierner, 1978 ;
Alamgir, 2017 )。 迄今为止, 芳香堆心菊的再生体系尚未见报道。因此, 建立芳香堆心菊的离体再生体系对菊科植物花型、花香等重要性状的分子机理研究及提高其药用价值具有十分重要的意义。
菊科植物是开花植物中进化最成功的家族之一。其头状花序由众多小花排列组合在同一压缩的花序上组成(
Chen et al., 2018 )。
中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科。依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (
Barkley et al., 2006 )。近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(
Chrysanthemum lavandulifolium )、菊花金不凋(
C .
morifolium cv. ‘Jinbudiao’)、药蒲公英(
Taraxacum officinale )和天山雪莲(
Saussurea involucrate ) (
陈华等, 2005 ;
荆甜蕊等, 2016 ;
刘萌萌, 2017 ;
吴志苹等, 2020 )。研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(
付建新等, 2012 ;
程密密, 2015 ;
Naing et al., 2016 ;
李金童等, 2016 ;
徐术菁, 2016 ;
刘萌萌, 2017 )。虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异。影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类。不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同。已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(
吴志苹等, 2020 )。但部分菊科植物的诱导再生培养基中需要添加适量的其它激素。例如, 在以北苍木(
Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(
荆甜蕊等, 2016 ;
刘萌萌, 2017 ;
左静静等, 2017 )。目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强。尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求。而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花。基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值。
在植物的再生过程中, 不同植物的再生体系差异较大。影响再生体系建立的主要因素有外植体、生长激素、遗传背景、培养环境及培养基成分(
程越, 2014 ;
张旭红等, 2018 )。培养基中添加的激素种类和浓度配比是影响植物再生的重要因素。研究表明, TDZ具有生长素和细胞分裂素的双重作用(
徐晓峰和黄学林, 2003 ;
杨怡帆等, 2019 )。使用TDZ可以有效诱导愈伤组织产生和不定芽分化(
王洪霞和郭尚敬, 2012 ;
包晗等, 2018 )。
陈雪等(2011) 以月季(
Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用。在以甜椒(
Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(
张俊华等, 2019 )。同样, 加入2,4-D可提高济麦系列小麦(
Triticum aestivum )的再生效率(
王树芸等, 2012 )。在培养基中加入KT可有效诱导珍珠半夏(
Pinellia ternate )球茎快速产生不定芽(
高亚曼等, 2016 )。在培养基中加入ZT (zeatin)和TDZ可使蓝莓(
Vaccinium spp.)叶片不定芽诱导率达100%。综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道。
本研究以芳香堆心菊叶片、茎段及下胚轴为外植体, 探讨NAA、6-BA、TDZ和2,4-D四种植物生长调节剂的不同浓度配比组合对芳香堆心菊再生的影响, 以筛选出适用于芳香堆心菊离体再生的培养基, 进而初步建立芳香堆心菊的再生体系, 以期为其遗传转化体系的建立及花型、花香和特殊药用价值的研究奠定基础。
1 植物材料 实验材料为2018年6月在北京花乡花木集团有限公司购入的芳香堆心菊(
Helenium aromaticum Hook Bailey)种子, 置于4°C保存。
2 培养基成分与培养条件 2.1 无菌播种及培养条件 芳香堆心菊无菌播种参照 罗虹等(2018) 的方法。将灭菌后的种子接种于普通MS培养基上, 置于北京林业大学组织培养室中培养。培养条件: 光强54 μmol·m
-2 ·s
-1 , 温度为(25±1)°C, 光周期为16小时光照/8小时黑暗。
2.2 愈伤组织诱导及不定芽分化 以MS为基本培养基, 添加不同种类和浓度的激素进行愈伤组织诱导及不定芽分化培养(
表1 )。激素种类包括NAA、6-BA、2,4-D和TDZ。共设计了25种不同激素浓度配比的培养基。取芳香堆心菊无菌叶片、茎段以及下胚轴作为外植体, 将叶片剪成0.5 cm×0.5 cm的小块, 并将其近轴面接触培养基。每个培养皿接种12-16个外植体。实验重复3次。统计出现愈伤组织的时间以及不定芽的天数、出愈率和再生率。
Table 1 表1 表1 芳香堆心菊愈伤组织诱导及不定芽分化培养基
Table 1
Medium for callus induction and adventitious buds differentiation of Helenium aromaticum Treatments Plant growth regulators (mg·L-1 ) NAA 6-BA 2,4-D TDZ A1 0.5 1 0 0 A2 0.5 2 1 0 A3 0.5 3 2 0 A4 1 1 1 0 A5 1 2 2 0 A6 1 3 0 0 A7 2 1 2 0 A8 2 2 0 0 A9 2 3 1 0 B1 0.2 1 0 0 B2 1 1 0 0 B3 0.2 2 0 0 B4 0.5 2 0 0 B5 1 2 0 0 B6 0.2 3 0 0 B7 0.5 3 0 0 C1 0.1 0.5 0 0.2 C2 0.2 1 0 0.2 C3 0.5 1.5 0 0.2 C4 0.2 0.5 0 0.5 C5 0.5 1 0 0.5 C6 0.1 1.5 0 0.5 C7 0.5 0.5 0 1 C8 0.1 1 0 1 C9 0.2 1.5 0 1
MS medium was used as basal medium in each treatment. 各处理均以MS为基本培养基。 新窗口打开 |
下载CSV 2.3 生根培养 待不定芽伸长至2 cm时, 将其接种于以1/2MS培养基为基本培养基, 添加不同浓度NAA的培养基中(
表2 )。培养条件同2.1节。生根培养15天后统计根数、生根率、根系状况以及开花率, 筛选出最佳生根培养基配方。
Table 2 表2 表2 芳香堆心菊生根培养基
Table 2
Medium for root culture of Helenium aromaticum Treatments Plant growth regulators (mg·L-1 ) NAA R0 0 R1 0.1 R2 0.2 R3 0.3
1/2MS medium was used as basal medium in each treatment. 各处理均以1/2MS为基本培养基。 新窗口打开 |
下载CSV 2.4 数据统计 使用Excel软件进行数据处理。利用以下公式进行计算。
出愈率=(诱导出愈伤组织的外植体数/同一处理中未污染的外植体总数)×100%;
再生率=(分化出不定芽的外植体数/同一处理中未污染的外植体总数)×100%;
生根率=(生根株数/同一处理中未污染的总株数)×100%;
开花率=(开花株数/同一处理中未污染的总株数)×100%。
3 结果与讨论 3.1 花部形态特征 在适宜的培养条件下, 芳香堆心菊可在试管内开花。其头状花序上仅具有管状花, 且全株(
图1A )具有特殊的芳香气味。管状花的花冠呈现闭合状态, 雌蕊和花药伸出花冠筒, 雄蕊完全被包裹在花冠筒内, 花瓣呈明显的绿色(
图1B , C), 且花冠表面被覆腺毛(
图1C )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1芳香堆心菊整株及花器官形态 (A) 整株(Bar=2 cm); (B) 头状花序(Bar=2 mm); (C) 单个小花(Bar=1 mm)。PE: 花瓣; OV: 子房; PI: 雌蕊; GT: 腺毛
Figure 1The whole plant and flower morphology of Helenium aromaticum (A) The whole plant (Bar=2 cm); (B) The capitulum (Bar=2 mm); (C) Single flower (Bar=1 mm). PE: Petal; OV: Ovary; PI: Pistil; GT: Glandular trichomes
3.2 离体再生体系的建立 3.2.1 不同外植体离体再生率
芳香堆心菊的叶片、茎段及下胚轴3种外植体在相同配方的培养基中其出愈时间、出愈率及愈伤组织的生长情况均有差异。在添加6-BA、NAA、2,4-D及TDZ不同组合种类的培养基中, 芳香堆心菊不同外植体均能诱导出愈伤组织。在添加TDZ的组合中, 3种外植体的出愈时间较短(在10-20天之间), 且出愈率均较高(平均在93.78%以上), 其中以下胚轴和茎段的出愈时间最短, 一般为10天左右。
以叶片为外植体时(
图2A ), 在未添加2,4-D处理下的叶片出愈时间早, 出愈率高, 且愈伤组织状态好; 在添加一定量2,4-D的处理下叶片出愈时间较长, 出愈率较低。同时, 当NAA的浓度高于6-BA时, 不利于愈伤组织的生长, 愈伤组织质地较软且易褐化, 出愈率仅41.67%。因此, 2,4-D不利于芳香堆心菊叶片愈伤组织的诱导, 且NAA的浓度不能高于6-BA。当6-BA的浓度达3 mg·L
-1 时, 芳香堆心菊的愈伤组织较软、生长不紧密, 说明较高浓度的6-BA不适合芳香堆心菊叶片愈伤组织的诱导。芳香堆心菊叶片在6-BA、2,4-D和NAA组合的培养基中均未分化出小芽。在添加TDZ的培养基中, 芳香堆心菊平均出愈时间为22天(
图2B )。TDZ浓度较高时, 愈伤组织较软, 生长状态较差, 表明较高浓度(1 mg·L
-1 )的TDZ不利于芳香堆心菊叶片愈伤组织的诱导及不定芽分化。在添加TDZ的培养基中, 芳香堆心菊叶片愈伤组织均能分化出芽点, 但只有在MS+0.2 mg·L
-1 NAA+1 mg·L
-1 6-BA+0.2 mg·L
-1 TDZ的培养基中愈伤组织才能成功分化出不定芽, 且再生率可达62.10%, 说明芳香堆心菊叶片在加入一定量TDZ的培养基中可以成功诱导出小芽(
图2C )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2芳香堆心菊叶片愈伤组织诱导及不定芽分化的阶段(C2) (A) 叶片外植体; (B) 愈伤组织(20天, 箭头所示); (C) 不定芽(30天, 箭头所示)。Bars=1 cm。C2同
表1。Figure 2Several stages of callus induction and adventitious bud differentiation of Helenium aromaticum leaves (C2) Table 1.图3A ), 2,4-D和6-BA对其愈伤组织的诱导能力与以叶片为外植体时相同。在不添加2,4-D的培养基中, 所有处理出愈时间均提前至20天左右(图3B ), 且出愈率均达100%; 高浓度(3 mg·L-1 ) 6-BA不利于愈伤组织的生长。在6-BA+NAA的基础上添加TDZ的培养基中, 茎段的出愈时间缩短至10天, 出愈率平均达98.41%。但随着TDZ浓度的增加, 茎段的出愈率有所降低, 愈伤组织质地较坚硬且颜色发黄。茎段在添加TDZ的培养基中并未诱导出不定芽, 即TDZ对茎段不定芽的诱导无促进作用。茎段在MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA培养基中, 愈伤组织在培养57天后出现芽点, 在65天分化出小芽(图3C ), 但分化率极低, 不定芽生长缓慢, 再生率仅为9.75%。图3 新窗口打开 |下载原图ZIP |生成PPT 图3芳香堆心菊茎段愈伤组织诱导及不定芽分化的阶段(B1) 表1。Figure 3Several stages of callus induction and adventitious bud differentiation of Helenium aromaticum stem fragments (B1) Table 1.图4A ), 在添加2,4-D的培养基中出愈时间相比叶片和茎段有所缩短(22天), 但出愈率较低; 在未添加2,4-D的培养基中, 所有处理均在20天内出现愈伤组织, 且出愈率较高(93.69%以上) (图4B )。由此表明, 2,4-D对以下胚轴为外植体的愈伤组织生长影响较小。高浓度6-BA不利于下胚轴愈伤组织的诱导, 这一结果与以叶片和茎段为外植体相同。在添加TDZ的培养基中, 出愈时间整体缩短, 平均在11天左右, 出愈率达80%以上, 但均未分化出不定芽。芳香堆心菊下胚轴在所有培养基中均未分化出不定芽, 但在MS+0.2 mg·L-1 NAA+0.5 mg·L-1 6-BA+ 0.5 mg·L-1 TDZ (C4)培养基中, 下胚轴的出愈时间为10天, 出愈率为100%, 且愈伤组织的生长状况良好。图4 新窗口打开 |下载原图ZIP |生成PPT 图4芳香堆心菊下胚轴愈伤组织诱导的阶段(C4) 表1。Figure 4Several stages of callus induction of Helenium aromaticum hypocotyls (C4) Table 1.表3 )。在外植体相同的情况下, 添加TDZ的培养基在愈伤组织诱导时间及出愈率方面均表现最佳。3种外植体在添加2,4-D的培养基中愈伤组织诱导时间及出愈率均表现较差, 说明2,4-D相较于TDZ并不利于芳香堆心菊愈伤组织的诱导; 在添加高浓度6-BA的培养基中愈伤组织质地较软, 没有分化小芽的迹象, 说明高浓度6-BA不能有效促进芳香堆心菊诱导分化出芽。在培养基相同的情况下, 下胚轴的诱导时间明显比茎段和叶片长, 但是并不能有效诱导不定芽。而叶片愈伤组织在MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ培养基上, 培养23天即开始分化出不定芽, 再生率可达62.10%。茎段愈伤组织在MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA培养基中能分化出不定芽, 但分化时间较长, 再生率仅为9.75%。由此可见, 以叶片为外植体时, 芳香堆心菊离体再生体系最优培养基配方为MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ。Table 3 表3 表3 芳香堆心菊不同外植体再生体系Different explant regeneration systems of Helenium aromaticum Explant types Callus induction medium Adventitious bud Regeneration Rooting medium Leaf MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ 62.1 1/2MS Stem segment MS+0.2 mg·L-1 NAA+1 mg·L-1 6-BA 9.75 1/2MS Hypocotyl MS+0.2 mg·L-1 NAA+0.5 mg·L-1 6-BA+0.5 mg·L-1 TDZ - - -
新窗口打开 |下载CSV -1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ培养基为诱导不定芽的最佳培养基。将芳香堆心菊不定芽接种于以1/2MS为基本培养基且添加了不同浓度NAA的生根培养基中(表2 )。生根培养10天时R1处理开始陆续生根, 根系细长繁密, 基本为白色, 部分为黄色, 生根率为45.46%; R2处理下, 不定芽13天开始生根, 但根系偏黄, 生根率较低, 为12.50%; R0处理下的不定芽16天开始生根, 根系细长、呈白色, 少数为浅黄色, 生根率达63.33%; 不定芽在R3处理下生根最慢(需18天), 且根系状态不佳, 生长缓慢, 生根率为14.71%。由此表明, 芳香堆心菊不定芽在4种培养基中均能生根, 但不同培养基对芳香堆心菊生根的影响不同。随着NAA浓度的增加, 生根率和生根数降低, 根系稀疏且植株处于低矮状态, 叶片偏黄(表4 )。Table 4 表4 表4 芳香堆心菊不定芽生根情况Rooting results of adventitious buds of Helenium aromaticum Treatments Time of The situation of root and plant Rate of Budding time (d) Rate of R0 16 Root system is sparse and slender; the plant is robust with normal stems and leaves, flowering normally 63.33 30 93.33 R1 10 The root system is dense and crude, some of which are deformed roots; the plants are dwarf with large leaves 45.46 34 86.36 R2 13 The root system is few and short, most of which are deformed roots; the plants dwarf can’t grow with normal leaves 12.50 - 0 R3 18 Root deformity and plants hardly grow taller and leaves weaker 14.71 - 0
新窗口打开 |下载CSV 表4 ; 图5A 1-B2)。图5 新窗口打开 |下载原图ZIP |生成PPT 图5芳香堆心菊不定芽生根及植株状态(从左至右分别为R0、R1、R2和R3) 表2。Figure 5Rooting and plant status of Helenium aromaticum adventitious buds (from left to right are R0, R1, R2, and R3, respectively) Table 2.-1 NAA+1 mg·L-1 6-BA+0.2 mg·L-1 TDZ)中(图6A )。叶片逐渐由淡绿色转变为深绿色, 约10天后开始逐渐膨大(图6B )。膨大的叶片再经过10天, 在周围的伤口处逐渐形成深绿色的愈伤组织(图6C )。 继续培养约23天诱导分化不定芽(图6D )。待不定芽继续生长2周, 长至2 cm时, 切取芳香堆心菊的不定芽接入1/2MS生根培养基中, 不定芽的生根率为63.33% (图6E )。继续培养15天, 芳香堆心菊可在试管内开花, 开花率达93.33% (图6F )。图6 新窗口打开 |下载原图ZIP |生成PPT 图6芳香堆心菊叶片离体再生体系的建立 Figure 6Establishment of in vitro regeneration system of Helenium aromaticum leaves 3.3 讨论 开展芳香堆心菊再生体系研究对于其遗传转化体系的建立具有重要意义。目前虽已有大量菊科植物再生体系的研究(赵喜亭等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ), 但多数研究是以头状花序类型为舌管兼备的菊科植物为材料, 少有利用头状花序上仅具有管状花的材料进行研究。本研究以头状花序上仅含管状花且全株具有芳香气味的芳香堆心菊为实验材料, 以叶片、分化状态较低的下胚轴及茎段作为外植体, 初步建立了芳香堆心菊叶片再生体系, 为建立芳香堆心菊遗传转化体系奠定了基础。丁晓霞, 2018 )。目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 )。在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 )。以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 )。在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽。Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似。以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%。综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致。以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%。李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 )。已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 )。然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响。本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好。以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽。冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率。在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 )。张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率。同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 )。Dewir等(2018) 认为TDZ的最适浓度具有物种特异性。本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生。不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别。添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 )。本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果。结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 )。2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 )。而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 )。-1 NAA、1 mg·L-1 6-BA和0.2 mg·L-1 TDZ的MS培养基中, 芳香堆心菊不定芽的诱导率达62.1%。后在1/2MS生根培养基中培养即可获得完整的再生植株。组织培养技术是研究菊科植物花香和花型的重要工具, 目前已经初步建立芳香堆心菊以叶片为外植体的再生体系, 后续将对此再生体系进一步优化, 提高其稳定性和分化效率, 为建立其遗传转化体系及基因工程育种奠定重要基础。[1] 包晗 , 张芮 , 张美玲 , 张虹 , 陈任 (2018 ). 甜叶菊叶片外植体再生体系的建立北方园艺 42(6 ), 16 -22 . [本文引用: 2] [2] 陈华 , 李平 , 刘晶 , 李银心 (2005 ). 药蒲公英再生体系的建立和优化生物工程学报 21, 244 -249 . [本文引用: 1] [3] 陈雪 , 张金柱 , 潘兵兵 , 桑成瑾 , 马雪 , 杨涛 , 车代弟 (2011 ). 月季愈伤组织的诱导及植株再生植物学报 46, 569 -574 . [本文引用: 1] [4] 程密密 (2015 ). 翠菊(Callistephus chinensis (L.) Nees)高频再生体系的建立及多倍体诱导硕士论文. 重庆: 西南大学 . pp. 17 . [本文引用: 2] [5] 程越 (2014 ). 菊花脑再生体系建立的研究硕士论文. 南京: 南京农业大学 . pp. 4 . [本文引用: 1] [6] 丁晓霞 (2018 ). 菊花组培再生体系研究进展辽宁林业科技 (4 ), 64 -65 , 68. [本文引用: 2] [7] 冯欢 , 易姝利 , 谢佳恒 , 雷梦琦 , 黄萱 (2014 ). 微型月季愈伤组织诱导及植株再生植物学报 49, 595 -602 . [本文引用: 1] [8] 付建新 , 张超 , 王翊 , 戴思兰 (2012 ). 甘菊下胚轴离体再生体系的建立北京林业大学学报 34(3 ), 91 -96 . [本文引用: 3] [9] 高亚曼 , 李妹芳 , 冀芦沙 (2016 ). 珍珠半夏(Pinellia ternate )高效再生体系的建立及其遗传稳定性检测分子植物育种 14, 980 -985 . [本文引用: 1] [10] 郝志华 , 顾偌铖 , 黄洁兰 , 武彦博 , 田铃 (2017 ). 常见多肉植物繁育技术的研究进展广东蚕业 51(4 ), 13 -16 . [本文引用: 1] [11] 荆甜蕊 , 徐思雅 , 倪亦飞 , 张勇娟 , 郭斌 , 尉亚辉 (2016 ). 天山雪莲生物技术研究进展基因组学与应用生物学 35, 2201 -2210 . [本文引用: 2] [12] 李海萍 , 张鲁刚 , 张静 , 茹磊 , 刘学成 , 孙希禄 (2011 ). 萝卜带柄子叶高频再生体系的建立植物学报 46, 331 -337 . [本文引用: 1] [13] 李金童 , 吴雅妮 , 丁兵 , 齐学军 , 张旸 , 解莉楠 (2016 ). 四种露地菊再生体系的建立北方园艺 40(19 ), 119 -124 . [本文引用: 3] [14] 刘晨旭 , 马欣 , 董凤丽 , 周蕴薇 (2015 ). 地被菊‘紫妍’和‘纽9722’的再生体系建立草业科学 32, 188 -195 . [本文引用: 1] [15] 刘娟旭 , 刘玲 , 王静 , 余义勋 (2008 ). 大叶相思下胚轴离体培养再生植株的研究林业科学研究 21, 403 -406 . [本文引用: 1] [16] 刘军 , 赵兰勇 , 丰震 , 张美蓉 , 吴银凤 (2004 ). 菊花叶盘片转基因再生体系的优化选择植物学通报 21, 556 -558 . [本文引用: 1] [17] 刘萌萌 (2017 ). 盆栽小菊高频再生体系建立与试管开花研究硕士论文. 银川: 宁夏大学 . pp. 1 . [本文引用: 4] [18] 罗虹 , 温小蕙 , 刘影 , 蒲娅 , 戴思兰 (2018 ). 芳香堆心菊试管开花及释香机制初探中国观赏园艺研究进展 (2018 ). 北京: 中国林业出版社. pp. 448 -460 . [本文引用: 2] [19] 孙永莲 , 戴晓港 , 李小平 , 陈赢男 (2019 ). 簸箕柳组培再生体系的建立南京林业大学学报(自然科学版) 43(2 ), 31 -37 . [本文引用: 1] [20] 王碧玉 (2017 ). 菊花再生及遗传转化体系的研究硕士论文. 沈阳: 沈阳农业大学 . pp. 8 . [本文引用: 1] [21] 王洪霞 , 郭尚敬 (2012 ). TDZ对甜椒不定芽分化的影响北方园艺 36(3 ), 104 -106 . [本文引用: 1] [22] 王欢 , 刘洋 , 苑禹 , 李旭 , 杜凤国 (2015 ). 蚂蚱腿子离体快繁技术研究植物生理学报 51, 2275 -2279 . [本文引用: 1] [23] 王树芸 , 吕波 , 李坤 , 吴佳洁 , 付道林 (2012 ). “济麦”系列品种成熟胚再生体系的优化研究分子植物育种 10, 476 -484 . [本文引用: 1] [24] 王自布 , 莫国秀 , 罗会兰 , 张德英 (2015 ). 菊花不同外植体组培快繁及其再生体系的研究北方园艺 39(18 ), 106 -109 . [本文引用: 1] [25] 吴志苹 , 高亦珂 , 范敏 , 高耀辉 (2020 ). 菊花‘金不凋’再生及遗传转化体系的构建分子植物育种 18, 150 -158 . [本文引用: 4] [26] 咸宏康 , 支秋娟 , 李卉 , 王长泉 (2019 ). 金樱子(Rosa laevigata Michx.)叶片直接再生不定芽体系的建立北方园艺 43(13 ), 93 -100 . [本文引用: 1] [27] 徐术菁 (2016 ). 东北蒲公英(Taraxacum ohwianum )叶片再生体系建立与优化硕士论文. 沈阳: 沈阳农业大学 . pp. 24 . [本文引用: 2] [28] 徐晓峰 , 黄学林 (2003 ). TDZ: 一种有效的植物生长调节剂植物学通报 20, 227 -237 . [本文引用: 1] [29] 杨怡帆 , 杨清江 , 潘峰 , 张新利 (2019 ). 基于主成分分析的不同浓度TDZ处理下‘红地球’葡萄膨大效果的综合评价农业科技通讯 (7 ), 165 -168 . [本文引用: 1] [30] 张俊华 , 苏振华 , 张泽鑫 , 李妹芳 , 郭尚敬 , 曹雪松 , 冀芦沙 (2019 ). 甜椒离体再生体系的构建分子植物育种 17, 8208 -8214 . [本文引用: 2] [31] 张旭红 , 王頔 , 梁振旭 , 孙美玉 , 张金政 , 石雷 (2018 ). 欧洲百合愈伤组织诱导及植株再生体系的建立植物学报 53, 840 -847 . [本文引用: 2] [32] 张媛 , 王康才 , 汤兴利 (2008 ). 杭白菊子叶和下胚轴组织培养技术的研究中草药 39, 1721 -1723 . [本文引用: 1] [33] 赵喜亭 , 蒋丽微 , 王苗 , 朱玉婷 , 张文芳 , 李明军 (2016 ). 怀黄菊间接体胚受体再生体系的建立及CmTGA1 的遗传转化植物学报 51, 525 -532 . [本文引用: 1] [34] 中国科学院中国植物志编辑委员会 (1984 ). 中国植物志, Vol. 74 . 北京: 科学出版社. pp. 1 . [本文引用: 1] [35] 左静静 , 闫贵云 , 安晓宁 , 霍利光 , 刘少翔 (2017 ). 正交设计优化北苍术组织培养研究中国农学通报 33(11 ), 21 -24 . [本文引用: 1] [36] Alamgir ANM 2017 ). Medicinal, non-medicinal, biopestici- des, color- and dye-yielding plants; secondary metabolites and drug principles; significance of medicinal plants; use of medicinal plants in the systems of traditional and complementary and alternative medicines (CAMs)In: Alamgir ANM, ed. Therapeutic Use of Medicinal Plants and Their Extracts. Cham Heidelberg: Springer . pp. 61 -104 . [本文引用: 1] [37] Barkley TM Brouillet L Strother JL 2006 ). Asteraceae . In: Flora of North America, Vol. 19. New York: Oxford Uni- versity Press. pp. 3 -69 . [本文引用: 1] [38] Bierner MW 1978 ). The taxonomy of Helenium sect. Cephalophora (Asteraceae)Syst Bot 277 -298 . [本文引用: 1] [39] Bloszyk E Samek Z Toman J Holub M 1975 ). Linifolin a and helenalin from Helenium aromaticum Phytochemistry 1444 -1445 . [本文引用: 1] [40] Bush SR Earle ED Langhans RW 1976 ). Plantlets from petal segments, petal epidermis, and shoot tips of the periclinal chimera, Chrysanthemum morifolium ‘Indianapolis’Am J Bot 729 -737 . [本文引用: 1] [41] Chen J Shen CZ Guo YP Rao GY 2018 ). Patterning the Asteraceae capitulum: duplications and differential expression of the flower symmetry CYC2-like genesFront Plant Sci 551 . [本文引用: 1] [42] Dewir YH Nurmansyah , Naidoo Y Teixeira da Silva JA 2018 ). Thidiazuron-induced abnormalities in plant tissue culturesPlant Cell Rep 1451 -1470 . [本文引用: 1] [43] Gómez-González S Ojeda F Torres-Morales P Palma JE 2016 ). Seed pubescence and shape modulate adaptive responses to fire cuesPLoS One e0159655 . [本文引用: 1] [44] Naing AH Park KI Mi YC Lim KB Kim CK 2016 ). Optimization of factors affecting efficient shoot regeneration in Chrysanthemum cv. ‘Shinma’Braz J Bot 975 -984 . [本文引用: 1] [45] Renou JP Brochard P Jalouzot R 1993 ). Recovery of transgenic chrysanthemum (Dendranthema grandiflora Tzvelev) after hygromycin resistance selectionPlant Sci 185 -197 . [本文引用: 1] [46] Tanaka K Kanno Y Kudo S Suzuki M 2000 ). Somatic embryogenesis and plant regeneration in chrysanthemum (Dendranthema grandiflorum ( Ramat.) Kitamura)Plant Cell Rep 946 -953 . URL PMID:30754837 [本文引用: 1] 程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...Callistephus chinensis (L.) Nees)高频再生体系的建立及多倍体诱导Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Pinellia ternate )高效再生体系的建立及其遗传稳定性检测程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...赵喜亭等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ), 但多数研究是以头状花序类型为舌管兼备的菊科植物为材料, 少有利用头状花序上仅具有管状花的材料进行研究.本研究以头状花序上仅含管状花且全株具有芳香气味的芳香堆心菊为实验材料, 以叶片、分化状态较低的下胚轴及茎段作为外植体, 初步建立了芳香堆心菊叶片再生体系, 为建立芳香堆心菊遗传转化体系奠定了基础. ...芳香堆心菊无菌播种参照罗虹等(2018) 的方法.将灭菌后的种子接种于普通MS培养基上, 置于北京林业大学组织培养室中培养.培养条件: 光强54 μmol·m-2 ·s-1 , 温度为(25±1)°C, 光周期为16小时光照/8小时黑暗. ...罗虹等(2018) 的方法.将灭菌后的种子接种于普通MS培养基上, 置于北京林业大学组织培养室中培养.培养条件: 光强54 μmol·m-2 ·s-1 , 温度为(25±1)°C, 光周期为16小时光照/8小时黑暗. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...赵喜亭等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ), 但多数研究是以头状花序类型为舌管兼备的菊科植物为材料, 少有利用头状花序上仅具有管状花的材料进行研究.本研究以头状花序上仅含管状花且全株具有芳香气味的芳香堆心菊为实验材料, 以叶片、分化状态较低的下胚轴及茎段作为外植体, 初步建立了芳香堆心菊叶片再生体系, 为建立芳香堆心菊遗传转化体系奠定了基础. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Rosa laevigata Michx.)叶片直接再生不定芽体系的建立李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Taraxacum ohwianum )叶片再生体系建立与优化Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...程越, 2014 ; 张旭红等, 2018 ).培养基中添加的激素种类和浓度配比是影响植物再生的重要因素.研究表明, TDZ具有生长素和细胞分裂素的双重作用(徐晓峰和黄学林, 2003 ; 杨怡帆等, 2019 ).使用TDZ可以有效诱导愈伤组织产生和不定芽分化(王洪霞和郭尚敬, 2012 ; 包晗等, 2018 ).陈雪等(2011) 以月季(Rosa chinensis )叶片为外植体, 在NAA+6-BA的基础上添加TDZ, 成功获得完整的再生植株, 说明TDZ对月季不定芽的诱导有一定的促进作用.在以甜椒(Capsicum annuum )叶片为外植体, 诱导不定芽时加入一定量的TDZ可增加每个愈伤组织产生的不定芽数目(张俊华等, 2019 ).同样, 加入2,4-D可提高济麦系列小麦(Triticum aestivum )的再生效率(王树芸等, 2012 ).在培养基中加入KT可有效诱导珍珠半夏(Pinellia ternate )球茎快速产生不定芽(高亚曼等, 2016 ).在培养基中加入ZT (zeatin)和TDZ可使蓝莓(Vaccinium spp.)叶片不定芽诱导率达100%.综上, 激素种类对不同物种的再生效率影响极大, 而目前大多数菊科植物的再生体系均以NAA+6-BA为主要激素, 少有添加其它激素(如2,4-D和TDZ)的相关报道. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...CmTGA1 的遗传转化赵喜亭等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ), 但多数研究是以头状花序类型为舌管兼备的菊科植物为材料, 少有利用头状花序上仅具有管状花的材料进行研究.本研究以头状花序上仅含管状花且全株具有芳香气味的芳香堆心菊为实验材料, 以叶片、分化状态较低的下胚轴及茎段作为外植体, 初步建立了芳香堆心菊叶片再生体系, 为建立芳香堆心菊遗传转化体系奠定了基础. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...Helenium aromaticum )为菊科(Compositae)堆心菊属草本花卉, 原产于智利, 自然条件下12月份成花, 耐寒性较差.芳香堆心菊花型独特, 头状花序上仅着生管状花, 且全株具芳香气味(Gómez-González et al., 2016 ).此外, 芳香堆心菊含有大量对人体健康有益的药用化学物质, 具有极高的药用及观赏价值(Bloszyk et al., 1975 ; Bierner, 1978 ; Alamgir, 2017 ). 迄今为止, 芳香堆心菊的再生体系尚未见报道.因此, 建立芳香堆心菊的离体再生体系对菊科植物花型、花香等重要性状的分子机理研究及提高其药用价值具有十分重要的意义. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...Helenium sect. Cephalophora (Asteraceae)Helenium aromaticum )为菊科(Compositae)堆心菊属草本花卉, 原产于智利, 自然条件下12月份成花, 耐寒性较差.芳香堆心菊花型独特, 头状花序上仅着生管状花, 且全株具芳香气味(Gómez-González et al., 2016 ).此外, 芳香堆心菊含有大量对人体健康有益的药用化学物质, 具有极高的药用及观赏价值(Bloszyk et al., 1975 ; Bierner, 1978 ; Alamgir, 2017 ). 迄今为止, 芳香堆心菊的再生体系尚未见报道.因此, 建立芳香堆心菊的离体再生体系对菊科植物花型、花香等重要性状的分子机理研究及提高其药用价值具有十分重要的意义. ...Helenium aromaticum Helenium aromaticum )为菊科(Compositae)堆心菊属草本花卉, 原产于智利, 自然条件下12月份成花, 耐寒性较差.芳香堆心菊花型独特, 头状花序上仅着生管状花, 且全株具芳香气味(Gómez-González et al., 2016 ).此外, 芳香堆心菊含有大量对人体健康有益的药用化学物质, 具有极高的药用及观赏价值(Bloszyk et al., 1975 ; Bierner, 1978 ; Alamgir, 2017 ). 迄今为止, 芳香堆心菊的再生体系尚未见报道.因此, 建立芳香堆心菊的离体再生体系对菊科植物花型、花香等重要性状的分子机理研究及提高其药用价值具有十分重要的意义. ...Chrysanthemum morifolium ‘Indianapolis’丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Helenium aromaticum )为菊科(Compositae)堆心菊属草本花卉, 原产于智利, 自然条件下12月份成花, 耐寒性较差.芳香堆心菊花型独特, 头状花序上仅着生管状花, 且全株具芳香气味(Gómez-González et al., 2016 ).此外, 芳香堆心菊含有大量对人体健康有益的药用化学物质, 具有极高的药用及观赏价值(Bloszyk et al., 1975 ; Bierner, 1978 ; Alamgir, 2017 ). 迄今为止, 芳香堆心菊的再生体系尚未见报道.因此, 建立芳香堆心菊的离体再生体系对菊科植物花型、花香等重要性状的分子机理研究及提高其药用价值具有十分重要的意义. ...Chrysanthemum cv. ‘Shinma’Chen et al., 2018 ).中国科学院中国植物志编辑委员会(1984) 将菊科植物分为管状花亚科和舌状花亚科.依据其头状花序上小花性别分化的不同, 可将菊科植物分为舌状花和管状花兼具的异型头状花序(heterogamous capitula)和仅具有管状花或仅具有舌状花的同型头状花序(homogamous capitula) (Barkley et al., 2006 ).近年来, 已有很多关于菊科植物再生体系的研究, 如盆栽小菊品种Chrystal Pink、Rainbow Worth、Chrystal Bronze和Little Rock, 甘菊(Chrysanthemum lavandulifolium )、菊花金不凋(C . morifolium cv. ‘Jinbudiao’)、药蒲公英(Taraxacum officinale )和天山雪莲(Saussurea involucrate ) (陈华等, 2005 ; 荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 吴志苹等, 2020 ).研究表明, 大多数菊科植物的再生体系以叶片、茎段、下胚轴及花瓣等为外植体, 添加一定配比的细胞分裂素和生长素, 即可诱导愈伤组织的产生和不定芽的分化(付建新等, 2012 ; 程密密, 2015 ; Naing et al., 2016 ; 李金童等, 2016 ; 徐术菁, 2016 ; 刘萌萌, 2017 ).虽然目前已有很多关于菊科植物再生体系的建立方法, 但是并不具有通用性, 且不同品种的诱导时间、诱导效率及再生芽数均有较大差异.影响菊科植物再生的主要因素包括外植体类型、培养基中激素种类与配比、培养条件、苗龄、叶片着生部位及基本培养基的种类.不同菊科植物由于其遗传背景的不同, 其离体再生体系也各不相同.已有研究表明, 菊科再生体系建立过程中, 在培养基中添加NAA和6-BA可以快速诱导不定芽的产生(吴志苹等, 2020 ).但部分菊科植物的诱导再生培养基中需要添加适量的其它激素.例如, 在以北苍木(Atractylodes lancea )胚轴为外植体时, 添加2,4-D和KT (kinetin)可有效诱导其长成完整植株; 在天山雪莲中添加TDZ (thidiazuron), 其不定芽的诱导率高达69%, 且每个外植体上不定芽的数量达8.5个, 其不定芽数量比利用NAA进行诱导时高1倍(荆甜蕊等, 2016 ; 刘萌萌, 2017 ; 左静静等, 2017 ).目前, 大多数菊科植物的再生体系均以舌管兼备的异型头状花序为材料, 且以叶片为外植体的愈伤组织诱导率较高且再生能力较强.尚缺乏对仅含管状花且具备芳香气味的物种进行再生体系研究, 故不能满足菊科植物头状花序多样性研究的目的和要求.而芳香堆心菊不仅全株具有芳香气味, 还具有极高的药用价值, 且头状花序上仅含管状花.基于芳香堆心菊具有多个优良性状的特点, 建立其再生体系有助于解析菊科植物花型、花香和药用价值相关的关键基因功能, 因此具有重要的生物学研究价值. ...Dendranthema grandiflora Tzvelev) after hygromycin resistance selection李海萍等, 2011 ; 王自布等, 2015 ; 李金童等, 2016 ).已有研究建立的菊科植物再生体系大多利用6-BA添加NAA诱导再生不定芽(Renou et al., 1993 ; 刘晨旭等, 2015 ; 徐术菁, 2016 ; 吴志苹等, 2020 ).然而, 不同外植体及培养基配方对芳香堆心菊再生体系均有影响.本研究表明, 同一外植体在添加TDZ的培养基中生长状况良好.以叶片为外植体, 加入适量的TDZ可诱导芳香堆心菊愈伤组织产生不定芽.冯欢等(2014) 以微型月季(Rosa hybrida )叶片为外植体, 在含6-BA和NAA培养基的基础上添加适量的TDZ, 可提高不定芽的诱导效率.在以甜叶菊(Stevia rebaudianum )的茎段和叶片为外植体时, 其最佳分化培养基为MS+0.5 mg·L-1 KT+1 mg·L-1 6-BA+3 mg·L-1 TDZ (包晗等, 2019 ).张俊华等(2019) 在建立较难再生的甜椒再生体系时, 使用TDZ代替NAA, 明显提高了甜椒的再生效率.同样, 加入TDZ有助于簸箕柳(Salix suchowensis )幼嫩茎段的再生(孙永莲等, 2019 ).Dewir等(2018) 认为TDZ的最适浓度具有物种特异性.本研究结果与以上研究相符, 在以叶片为外植体的培养基中加入TDZ可有效诱导芳香堆心菊的再生.不同物种再生体系的建立需要的外植体类型、植物生长调节剂的种类组合及浓度配比均有很大差别.添加TDZ和NAA可有效诱导欧洲百合(Lilium martagon )愈伤组织, 但不利于增殖培养(张旭红等, 2018 ).本研究选取6-BA、TDZ和2,4-D三种植物生长调节剂, 探讨其对芳香堆心菊叶片、茎段及下胚轴的愈伤组织及不定芽诱导的作用效果.结果表明, 高浓度6-BA不利于不定芽的诱导, 这与高浓度6-BA会抑制地被菊(C. morifolium )不定芽的分化研究结果一致(李金童等, 2016 ).2,4-D对叶片和下胚轴愈伤组织的诱导效果不佳, 且在后期容易产生褐化, 说明其不利于芳香堆心菊不定芽的诱导, 这与前人在甘菊、大叶相思(Acacia auriculaeformis )和杭白菊(C. morifolium )中的研究结果相似(刘娟旭等, 2008 ; 张媛等, 2008 ; 付建新等, 2012 ).而加入一定量的TDZ不仅提高了下胚轴及叶片愈伤组织的诱导效率, 还可以诱导芳香堆心菊叶片产生不定芽, 这与前人在小美旱杨(Populus popularis )及金樱子(R. laevigata )中的研究结果相似(咸宏康等, 2019 ). ...Dendranthema grandiflorum ( Ramat.) Kitamura)丁晓霞, 2018 ).目前在菊科植物再生体系的研究中大多选择叶片、茎段及下胚轴作为外植体, 也有使用花瓣、叶柄和腋芽作为外植体, 并获得较好的再生效率(程密密, 2015 ; 王欢等, 2015 ; 郝志华等, 2017 ; 王碧玉, 2017 ; 丁晓霞, 2018 ).在以甘菊下胚轴为外植体时, 虽然成功诱导出不定芽, 但分化率仅为25.5% (付建新等, 2012 ).以菊花叶片为外植体时, 其不定芽的分化率可达96% (刘军等, 2004 ).在以翠菊(Callistephus chinensis )无菌苗的茎段为外植体时可获得完整的再生植株, 且再生率为82.2%, 而以叶柄为外植体时仅能诱导出愈伤组织, 不能分化出不定芽.Tanaka等(2000) 利用菊花叶片和茎段作为外植体, 其再生效率明显优于其它组织和器官, 这与Bush等(1976) 的结果相似.以菊花金不换的叶片为外植体, 其在MS+1.5 mg·L-1 NAA+ 1 mg·L-1 6-BA培养基中分化率可达100%.综上可知, 在菊科植物再生体系的建立过程中, 不同的菊科植物最适宜诱导再生的外植体不同, 但大多数菊科植物以叶片为外植体建立再生体系的诱导效率最高, 这与本研究的结果一致.以芳香堆心菊叶片为外植体时, 愈伤组织诱导率为100%, 不定芽分化率可达62.10%. ...京ICP备16067583号-21 北京玛格泰克科技发展有限公司 设计开发  ,*北京林业大学园林学院, 国家花卉工程技术研究中心, 花卉种质创新与分子育种北京市重点实验室, 城乡生态环境北京实验室, 北京 100083
,*北京林业大学园林学院, 国家花卉工程技术研究中心, 花卉种质创新与分子育种北京市重点实验室, 城乡生态环境北京实验室, 北京 100083
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT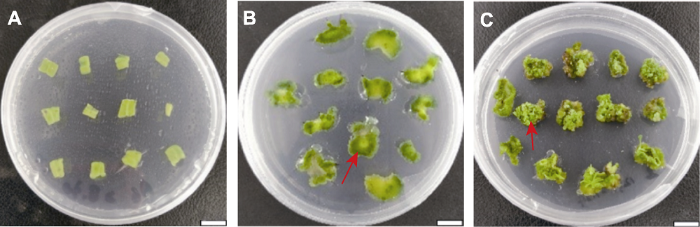 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT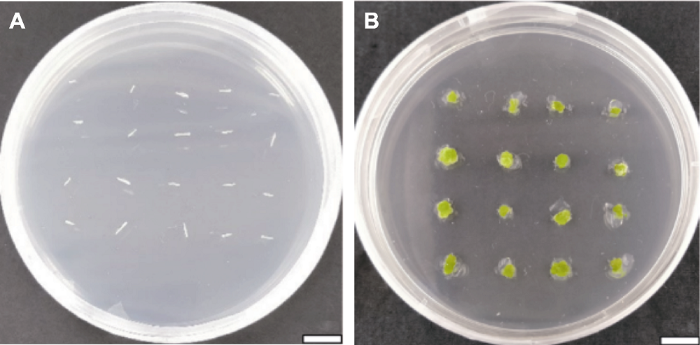 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT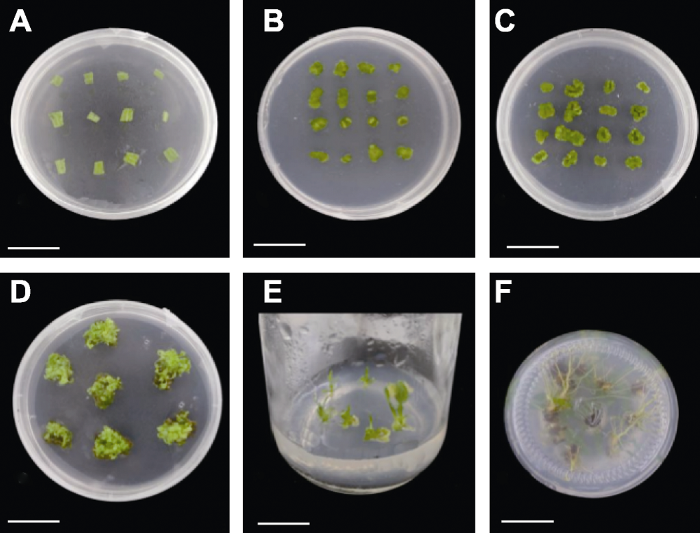 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}