Research Progress on Genetics and Breeding of Rice Roots
Yilan Zhang1, Xue Lin1, Yi Wu1, Mengjia Li1, Shengjie Zhang1, Mei Lu1, Yuchun Rao,1,*, Yuexing Wang,2,*1College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua 321004, China 2State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 310006, China
Abstract As an important part of rice (Oryza sativa), the root system plays multiple roles in rice growth, including plant fixation, water and nutrients acquisition, amino acid and hormone biosynthesis. Root morphological structure and physiological function are closely related to rice yield and quality, and resistance. So far, many QTLs and genes regulating rice root system have been identified through genetic and biochemical approaches. In this paper, we summarized current progress on the study of rice root system-related QTLs and genes and the future research direction, so as to provide a reference for further screening and cloning root related genes and improving the model of ideal plant architecture of rice. Keywords:rice;root system;QTL mapping;control gene
PDF (2977KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 章怡兰, 林雪, 吴仪, 李梦佳, 张晟婕, 路梅, 饶玉春, 王跃星. 水稻根系遗传育种研究进展. 植物学报, 2020, 55(3): 382-393 doi:10.11983/CBB20021 Zhang Yilan, Lin Xue, Wu Yi, Li Mengjia, Zhang Shengjie, Lu Mei, Rao Yuchun, Wang Yuexing. Research Progress on Genetics and Breeding of Rice Roots. Chinese Bulletin of Botany, 2020, 55(3): 382-393 doi:10.11983/CBB20021
AduMO, YawsonDO, ArmahFA, AsarePA, FrimponKA (2018). Meta-analysis of crop yields of full, deficit, and partial root-zone drying irrigation Agric Water Manag 197, 79-90. [本文引用: 1]
ChenG, LiuCL, GaoZY, ZhangY, ZhuL, HuJ, RenDY, XuGH, QianQ (2018a). Driving the expression of RAA1 with a drought-responsive promoter enhances root growth in rice, its accumulation of potassium and its tolerance to moisture stress Environ Exp Bot 147, 147-156. [本文引用: 1]
ChenH, MaB, ZhouY, HeSJ, TangSY, LuX, XieQ, ChenSY, ZhangJS (2018b). E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein Proc Natl Acad Sci USA 115, 4513-4518. [本文引用: 1]
ChoSH, KangKY, LeeSH, LeeIJ, PaekNC (2016). OsWOX3A is involved in negative feedback regulation of the gibberellic acid biosynthetic pathway in rice (Oryza sativa) J Exp Bot 67, 1677-1687. URLPMID:26767749 [本文引用: 1]
ChoSH, YooSC, ZhangHT, PandeyaD, KohHJ, HwangJY, KimGT, PeakNC (2013). The ricenarrow leaf 2 and narrow leaf 3 loci encode WUSCHEL-related homeobox 3a (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development New Phytol 198, 1071-1084. [本文引用: 1]
ClarkRT, FamosoAN, ZhaoK, ShaffJE, CraftEJ, BustamanteCD, McCouchSR, AneshansleyDJ, KochianLV (2013). High-throughput two-dimensional root system phenotyping platform facilitates genetic analysis of root growth and development Plant Cell Environ 36, 454-466. [本文引用: 2]
CoudertY, PérinC, CourtoisB, KhongNG, GantetP (2010). Genetic control of root development in rice, the model cereal Trends Plant Sci 15, 219-226. [本文引用: 1]
DaiXY, WangYY, ZhangWH (2016). OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice J Exp Bot 67, 947-960. URLPMID:26663563 [本文引用: 1]
DonaldCM (1968). The breeding of crop ideotypes Euphytica 17, 385-403. [本文引用: 1]
GaoSP, FangJ, XuF, WangW, SunXH, ChuJF, CaiBD, FengYQ, ChuCC (2014). CYTOKININ OXIDASE/DEHYDROGENASE 4 integrates cytokinin and auxin signaling to control rice crown root formation Plant Physiol 165, 1035-1046. DOI:10.1104/pp.114.238584URLPMID:24808099 [本文引用: 1] Crown roots constitute the majority of the rice (Oryza sativa) root system and play an important role in rice growth and development. However, the molecular mechanism of crown root formation in rice is not well understood. Here, we characterized a rice dominant mutant, root enhancer1 (ren1-D), which was observed to exhibit a more robust root system, increased crown root number, and reduced plant height. Molecular and genetic analyses revealed that these phenotypes are caused by the activation of a cytokinin oxidase/dehydrogenase (CKX) family gene, OsCKX4. Subcellular localization demonstrated that OsCKX4 is a cytosolic isoform of CKX. OsCKX4 is predominantly expressed in leaf blades and roots. It is the dominant CKX, preferentially expressed in the shoot base where crown root primordia are produced, underlining its role in root initiation. OsCKX4 is induced by exogenous auxin and cytokinin in the roots. Furthermore, one-hybrid assays revealed that OsCKX4 is a direct binding target of both the auxin response factor OsARF25 and the cytokinin response regulators OsRR2 and OsRR3. Overexpression and RNA interference of OsCKX4 confirmed that OsCKX4 plays a positive role in crown root formation. Moreover, expression analysis revealed a significant alteration in the expression of auxin-related genes in the ren1-D mutants, indicating that the OsCKX4 mediates crown root development by integrating the interaction between cytokinin and auxin. Transgenic plants harboring OsCKX4 under the control of the root-specific promoter RCc3 displayed enhanced root development without affecting their shoot parts, suggesting that this strategy could be a powerful tool in rice root engineering.
HuangSJ, ChenS, LiangZH, ZhangCM, YanM, ChenJG, XuGH, FanXR, ZhangYL (2015). Knockdown of the partner protein OsNAR2.1 for high-affinity nitrate transport represses lateral root formation in a nitrate-dependent manner Sci Rep 5, 18192. URLPMID:26644084 [本文引用: 2]
InukaiY, SakamotoT, Ueguchi-TanakaM, ShibataY, GomiK, UmemuraI, HasegawaY, AshikariM, KitanoH, MatsuokaM (2005). Crown rootless 1, which is essential for crown root formation in rice, is a target of an AUXIN RESPONSE FACTOR in auxin signaling Plant Cell 17, 1387-1396. URLPMID:15829602 [本文引用: 1]
JongdeeB, FukaiS, CooperM (2002). Leaf water potential and osmotic adjustment as physiological traits to improve drought tolerance in rice Field Crops Res 76, 153-163. [本文引用: 1]
KitomiY, ItoH, HoboT, AyaK, KitanoH, InukaiY (2011). The auxin responsive AP2/ERF transcription factor CROWN ROOTLESS5 is involved in crown root initiation in rice through the induction of OsRR1, a type-A response regulator of cytokinin signaling Plant J 67, 472-484. DOI:10.1111/j.1365-313X.2011.04610.xURL [本文引用: 1] Cytokinin is known to have negative effects on de novo auxin-induced root formation. However, the regulatory mechanisms of root initiation by both cytokinin and auxin are poorly understood. In this study, we characterized a rice mutant, termed crown rootless5 (crl5), which produced fewer crown roots and displayed impaired initiation of crown root primordia. The expression of CRL5, which encodes a member of the large AP2/ERF transcription factor family protein, was observed in the stem region where crown root initiation occurs. Exogenous auxin treatment induced CRL5 expression without de novo protein biosynthesis, which also required the degradation of AUX/IAA proteins. A putative auxin response element in the CRL5 promoter region specifically interacted with a rice ARF, demonstrating that CRL5 may be a direct target of an ARF, similar to CRL1/ADVENTITIOUS ROOTLESS1 (ARL1) that also regulates crown root initiation. A crl1 crl5 double mutant displayed an additive phenotype, indicating that these two genes function in different genetic pathways for crown root initiation. In addition, ProACT: CRL5/WT showed a cytokinin-resistant phenotype for crown root initiation, and also up-regulated the expression of two negative regulators of cytokinin signaling, OsRR1 and OsRR2, which were downregulated in crl5. Transgenic plants that over-expressed OsRR1 under the control of the CRL5 promoter in a crl5 mutant background produced a higher number of crown roots than the crl5 plant. Taken together, these results indicate that auxin-induced CRL5 promotes crown root initiation through repression of cytokinin signaling by positively regulating type-A RR, OsRR1.
KitomiY, OgawaA, KitanoH, InukaiY (2008). CRL4 regulates crown root formation through auxin transport in rice Plant Root 2, 19-28. DOI:10.3117/plantroot.2.19URL [本文引用: 1]
KongFL ( 2006). Summary of QTL mapping In: Quantitative Genetics in Plants. Beijing: China Agricultural University Press. pp. 358-359. [本文引用: 1]
KumariS, JoshiR, SinghK, RoyS, TripathiAK, SinghP, Singla-PareekSL, PareekA (2015). Expression of a cyclophilin OsCyp2-P isolated from a salt-tolerant landrace of rice in tobacco alleviates stress via ion homeostasis and limiting ROS accumulation Funct Integr Genomics 15, 395-412. URLPMID:25523384 [本文引用: 1]
LiJ, ZhuSH, SongXW, ShenY, ChenHM, YuJ, YiKK, LiuYF, KarplusVJ, WuP, DengXW (2006). A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem Plant Cell 18, 340-349. [本文引用: 1]
LiuHJ, WangSF, YuXB, YuJ, HeXW, ZhangSL, ShouHX, WuP (2005). ARL1, a LOB-domain protein required for adventitious root formation in rice Plant J 43, 47-56. DOI:10.1111/j.1365-313X.2005.02434.xURLPMID:15960615 [本文引用: 1] Adventitious roots constitute the bulk of the fibrous root system in cereals. Compared with the current understanding of shoot development, knowledge of the molecular mechanisms of development of the adventitious roots of cereals is limited. We have isolated and characterized a novel gene controlling the initiation of adventitious root primordia in rice (Oryza sativa L.). The gene, designated Adventitious rootless1 (ARL1), encodes a protein with a LATERAL ORGAN BOUNDARIES (LOB) domain. It is expressed in lateral and adventitious root primordia, tiller primordia, vascular tissues, scutellum, and young pedicels. ARL1 is a nuclear protein and can form homodimers. ARL1 is an auxin- and ethylene-responsive gene, and the expression pattern of ARL1 in roots parallels auxin distribution. Our findings suggest that ARL1 is an auxin-responsive factor involved in auxin-mediated cell dedifferentiation, and that it promotes the initial cell division in the pericycle cells adjacent to the peripheral vascular cylinder in the stem.
LiuSP, WangJR, WangL, WangXF, XueYH, WuP, ShouHX (2009). Adventitious root formation in rice requires OsGNOM1 and is mediated by the OsPINs family Cell Res 19, 1110-1119. DOI:10.1038/cr.2009.70URLPMID:19546891 [本文引用: 1] The fibrous root system in cereals comprises primarily adventitious roots (ARs), which play important roles in nutrient and water uptake. Current knowledge regarding the molecular mechanism underlying AR development is still limited. We report here the isolation of four rice (Oryza sativa L.) mutants, from different genetic backgrounds, all of which were defective in AR formation. These mutants exhibited reduced numbers of lateral roots (LRs) and partial loss of gravitropism. The mutants also displayed enhanced sensitivity to N-1-naphthylphthalamic acid, an inhibitor of polar auxin transport (PAT), indicating that the mutations affected auxin transport. Positional cloning using one of the four mutants revealed that it was caused by loss-of-function of a guanine nucleotide exchange factor for ADP-ribosylation factor (OsGNOM1). RT-PCR and analysis of promoter::GUS transgenic plants showed that OsGNOM1 is expressed in AR primordia, vascular tissues, LRs, root tips, leaves, anthers and lemma veins, with a distribution pattern similar to that of auxin. In addition, the expressions of OsPIN2, OsPIN5b and OsPIN9 were altered in the mutants. Taken together, these findings indicate that OsGNOM1 affects the formation of ARs through regulating PAT.
LiuW, XuZH, LuoD, XueHW (2003). Roles of OsCKI1, a rice casein kinase I, in root development and plant hormone sensitivity Plant J 36, 189-202. URLPMID:14535884 [本文引用: 1]
LuGW, ConevaV, CasarettoJA, YingS, MahmoodK, LiuF, NambaraE, BiYM, RothsteinSJ (2015). OsPIN5b modulates rice (Oryza sativa) plant architecture and yield by changing auxin homeostasis, transport and distribution Plant J 83, 913-925. DOI:10.1111/tpj.12939URLPMID:26213119 [本文引用: 2] Plant architecture attributes such as tillering, plant height and panicle size are important agronomic traits that determine rice (Oryza sativa) productivity. Here, we report that altered auxin content, transport and distribution affect these traits, and hence rice yield. Overexpression of the auxin efflux carrier-like gene OsPIN5b causes pleiotropic effects, mainly reducing plant height, leaf and tiller number, shoot and root biomass, seed-setting rate, panicle length and yield parameters. Conversely, reduced expression of OsPIN5b results in higher tiller number, more vigorous root system, longer panicles and increased yield. We show that OsPIN5b is an endoplasmic reticulum (ER) -localized protein that participates in auxin homeostasis, transport and distribution in vivo. This work describes an example of an auxin-related gene where modulating its expression can simultaneously improve plant architecture and yield potential in rice, and reveals an important effect of hormonal signaling on these traits.
McCouchSR, TeytelmanL, XuYB, LobosKB, ClareK, WaltonM, FuBY, MaghirangR, LiZK, XingYZ, ZhangQF, KonoI, YanoM, FjellstromR, DeClerckG, SchneiderD, CartinhourS, WareD, SteinL (2002). Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.) DNA Res 9, 199-207. [本文引用: 17]
MochizukiS, JikumaruY, NakamuraH, KoiwaiH, SasakiK, KamiyaY, IchikawaH, MinamiE, NishizawaY (2014). Ubiquitin ligase EL5 maintains the viability of root meristems by influencing cytokinin-mediated nitrogen effects in rice J Exp Bot 65, 2307-2318. [本文引用: 1]
NadaRM, Abo-HegazySE, BudranEG, AbogadallahGM (2019). The interaction of genes controlling root traits is required for the developmental acquisition of deep and thick root traits and improving root architecture in response to low water or nitrogen content in rice (Oryza sativa L.) cultivars Plant Physiol Biochem 141, 122-132. URLPMID:31151078 [本文引用: 1]
NakamuraA, UmemuraI, GomiK, HasegawaY, KitanoH, SazukaT, MatsuokaM (2006). Production and characterization of auxin-insensitive rice by overexpression of a mutagenized rice IAA protein Plant J 46, 297-306. [本文引用: 1]
ObaraM, FukutaY, YanagiharaS (2019). Genetic variation and QTLs related to root development in upland New Rice for Africa (NERICA) varieties Breeding Sci 69, 94-103. [本文引用: 3]
ShresthaR, Al-ShugeairyZ, Al-OgaidiF, MunasingheM, RadermacherM, VandenhirtzJ, PriceAH (2014). Comparing simple root phenotyping methods on a core set of rice genotypes Plant Biol 16, 632-642. URLPMID:24015692 [本文引用: 1]
SunLJ, ZhangQ, WuJX, ZhangLQ, JiaoXW, ZhangSW, ZhangZG, SunDY, LuTG, SunY (2014). Two rice authentic histidine phosphotransfer proteins, OsAHP1 and OsAHP2, mediate cytokinin signaling and stress responses in rice Plant Physiol 165, 335-345. URLPMID:24578505 [本文引用: 2]
TemnykhS, ParkWD, AyresN, CartinhourS, HauckN, LipovichL, ChoYG, IshiiT, McCouchSR (2000). Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.) Theor Appl Genet 100, 697-712. [本文引用: 13]
WangF, CoeRA, KarkiS, WanchanaS, ThakurV, HenryA, LinHC, HuangJL, PengSB, QuickWP (2016). Overexpression of OsSAP16 regulates photosynthesis and the expression of a broad range of stress response genes in rice (Oryza sativa L.) PLoS One 11, e0157244. URLPMID:27303811 [本文引用: 3]
WangXF, HeFF, MaXX, MaoCZ, HodgmanC, LuCG, WuP (2011). OsCAND1 is required for crown root emergence in rice Mol Plant 4, 289-299. URLPMID:20978084 [本文引用: 1]
WangYH, WangD, GanT, LiuLL, LongWH, WangYL, NiuM, LiXH, ZhengM, JiangL, WanJM (2016). CRL6, a member of the CHD protein family, is required for crown root development in rice Plant Physiol Biochem 105, 185-194. URLPMID:27108205
XuJ, WangL, ZhouMY, ZengDL, HuJ, ZhuL, RenDY, DongGJ, GaoZY, GuoLB, QianQ, ZhangWZ, ZhangGH (2017). Narrow albino leaf 1 is allelic to CHR729, regulates leaf morphogenesis and development by affecting auxin metabolism in rice Plant Growth Regul 82, 175-186. [本文引用: 1]
YangSQ, LiWQ, MiaoH, GanPF, QiaoL, ChangYL, ShiCH, ChenKM (2016). REL2, a gene encoding an unknown function protein which contains DUF630 and DUF632 domains controls leaf rolling in rice Rice 9, 37. DOI:10.1186/s12284-016-0105-6URLPMID:27473144 [本文引用: 2] BACKGROUND: Rice leaves are important energy source for the whole plant. An optimal structure will be beneficial for rice leaves to capture light energy and exchange gas, thus increasing the yield of rice. Moderate leaf rolling and relatively erect plant architecture may contribute to high yield of rice, but the relevant molecular mechanism remains unclear. RESULTS: In this study, we identified and characterized a rolling and erect leaf mutant in rice and named it as rel2. Histological analysis showed that the rel2 mutant has increased number of bulliform cells and reduced size of middle bulliform cells. We firstly mapped REL2 to a 35-kb physical region of chromosome 10 by map-based cloning strategy. Further analysis revealed that REL2 encodes a protein containing DUF630 and DUF632 domains. In rel2 mutant, the mutation of two nucleotide substitutions in DUF630 domain led to the loss-of-function of REL2 locus and the function of REL2 could be confirmed by complementary expression of REL2 in rel2 mutant. Further studies showed that REL2 protein is mainly distributed along the plasma membrane of cells and the REL2 gene is relatively higher expressed in younger leaves of rice. The results from quantitative RT-PCR analysis indicated that REL2 functioning in the leaf shape formation might have functional linkage with many genes associated with the bulliform cells development, auxin synthesis and transport, etc. CONCLUSIONS: REL2 is the DUF domains contained protein which involves in the control of leaf rolling in rice. It is the plasma membrane localization and its functions in the control of leaf morphology might involve in multiple biological processes such as bulliform cell development and auxin synthesis and transport.
ZhangHG, ZhangLJ, SiH, GeYS, LiangGH, GuMH, TangSZ (2016). Rf5 is able to partially restore fertility to Honglian-type cytoplasmic male sterile japonica rice (Oryza sativa) lines Mol Breeding 36, 102. DOI:10.1007/s11032-016-0511-xURL [本文引用: 2]
ZhaoY, ChengSF, SongYL, HuangYL, ZhouSL, LiuXY, ZhouDX (2015). The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling Plant Cell 27, 2469-2483. DOI:10.1105/tpc.15.00227URLPMID:26307379 [本文引用: 2] Crown roots are the main components of the fibrous root system in rice (Oryza sativa). WOX11, a WUSCHEL-related homeobox gene specifically expressed in the emerging crown root meristem, is a key regulator in crown root development. However, the nature of WOX11 function in crown root development has remained elusive. Here, we identified a rice AP2/ERF protein, ERF3, which interacts with WOX11 and was expressed in crown root initials and during crown root growth. Functional analysis revealed that ERF3 was essential for crown root development and acts in auxin- and cytokinin-responsive gene expression. Downregulation of ERF3 in wox11 mutants produced a more severe root phenotype. Also, increased expression of ERF3 could partially complement wox11, indicating that the two genes functioned cooperatively to regulate crown root development. ERF3 and WOX11 shared a common target, the cytokinin-responsive gene RR2. The expression of ERF3 and WOX11 only partially overlapped, underlining a spatio-temporal control of RR2 expression and crown root development. Furthermore, ERF3-regulated RR2 expression was involved in crown root initiation, while the ERF3/WOX11 interaction likely repressed RR2 during crown root elongation. These results define a mechanism regulating gene expression involved in cytokinin signaling during different stages of crown root development in rice. 水稻根系育种的意义与前景 1 2016
Driving the expression of RAA1 with a drought-responsive promoter enhances root growth in rice, its accumulation of potassium and its tolerance to moisture stress 1 2018
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein 1 2018
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
OsWOX3A is involved in negative feedback regulation of the gibberellic acid biosynthetic pathway in rice (Oryza sativa) 1 2016
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
The ricenarrow leaf 2 and narrow leaf 3 loci encode WUSCHEL-related homeobox 3a (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development 1 2013
... 已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
High-throughput two-dimensional root system phenotyping platform facilitates genetic analysis of root growth and development 2 2013
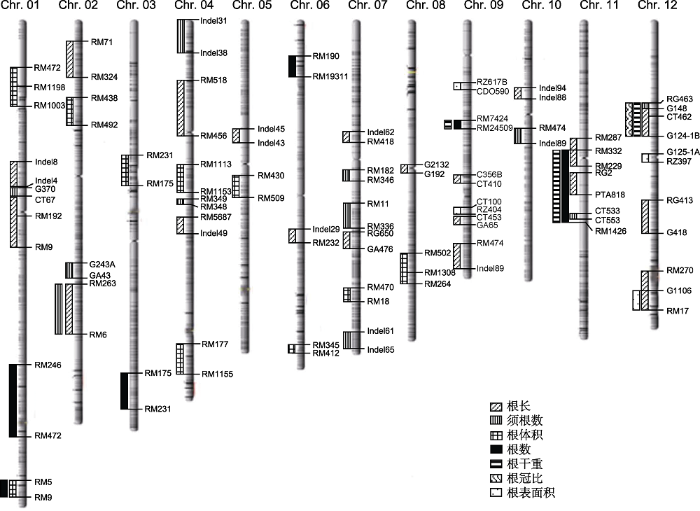
... 根据国家水稻数据中心网站(www.ricedata.cn)公布的数据, 我们统计了已克隆的水稻根系基因(表4).发现目前已经成功克隆的大量基因参与调控水稻主根、不定根、侧根以及根冠生长.在水稻根系性状的调控基因克隆进展中, 根长性状受到较多关注.GLR3.1是水稻中1个glu受体基因, 突变体的根分生组织活性降低, 并伴有程序性细胞死亡, 通常表现为初生根、不定根和侧根均变短, 初生根顶端直径变小, 该基因对维持苗期根尖分生组织的细胞分裂和个体细胞存活至关重要(Li et al., 2006).另1个与根长发育QTL相关的基因Dro1控制根的生长角度并增加根深(Clark et al., 2013).生长素通过AUXIN响应因子(ARF转录因子)调节Dro1的表达.Dro1可能通过调节表皮细胞伸长来调节根的重力反应, 使表皮根相对于重力而定向生长, 增加了根与水平轴之间的夹角, 导致根扎得更深.OsNAR2.1基因编码一种硝酸盐转运蛋白伴侣蛋白, 当N供应为NO3-时, 敲减OsNAR2.1会抑制根部生长和侧根的发生, 使主根变短, 侧根数目减少, 不定根变短(Huang et al., 2015). ...
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
Genetic control of root development in rice, the model cereal 1 2010
... 国际水稻研究所数据库中提供的3 024个水稻品种的62个性状中没有一个根系性状(Shrestha et al., 2014).这表明在水稻育种计划中缺乏可用的根系特征数据资源, 且根系性状并没有得到广泛应用.由于根属于地下部分且结构庞大复杂, 难以进行表型分析, 通过常规育种对根系进行遗传改良具有较大困难(Nada et al., 2019).数量性状定位(QTL)及其在标记辅助育种(MAB)中的应用成为根系研究中具有突破性的方法(Coudert et al., 2010).近年来, 利用分子标记辅助选择技术对水稻根性状QTL的遗传改良逐渐增多(Temnykh et al., 2000; McCouch et al., 2002; 胡兴明, 2004; 韩龙植, 2005; 吴伟明, 2006; 翟荣荣, 2012; 徐晓明等, 2016; 曲志恒, 2016; 索艺宁等, 2018).此外, 自1996年超级稻育种技术路线提出以来, 株型改良与杂种优势相结合的作物育种方式, 其科学性与正确性已经过充分的实践检验.中国的超级杂交水稻育种计划于2016年已实现第五期育种产量目标(周正平等, 2019), 以QTL定位为基础的水稻理想根系育种必将会不断助力水稻增产、稳产潜力的挖掘. ...
OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice 1 2016
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
The auxin responsive AP2/ERF transcription factor CROWN ROOTLESS5 is involved in crown root initiation in rice through the induction of OsRR1, a type-A response regulator of cytokinin signaling 1 2011
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
CRL4 regulates crown root formation through auxin transport in rice 1 2008
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
Expression of a cyclophilin OsCyp2-P isolated from a salt-tolerant landrace of rice in tobacco alleviates stress via ion homeostasis and limiting ROS accumulation 1 2015
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem 1 2006
... 根据国家水稻数据中心网站(www.ricedata.cn)公布的数据, 我们统计了已克隆的水稻根系基因(表4).发现目前已经成功克隆的大量基因参与调控水稻主根、不定根、侧根以及根冠生长.在水稻根系性状的调控基因克隆进展中, 根长性状受到较多关注.GLR3.1是水稻中1个glu受体基因, 突变体的根分生组织活性降低, 并伴有程序性细胞死亡, 通常表现为初生根、不定根和侧根均变短, 初生根顶端直径变小, 该基因对维持苗期根尖分生组织的细胞分裂和个体细胞存活至关重要(Li et al., 2006).另1个与根长发育QTL相关的基因Dro1控制根的生长角度并增加根深(Clark et al., 2013).生长素通过AUXIN响应因子(ARF转录因子)调节Dro1的表达.Dro1可能通过调节表皮细胞伸长来调节根的重力反应, 使表皮根相对于重力而定向生长, 增加了根与水平轴之间的夹角, 导致根扎得更深.OsNAR2.1基因编码一种硝酸盐转运蛋白伴侣蛋白, 当N供应为NO3-时, 敲减OsNAR2.1会抑制根部生长和侧根的发生, 使主根变短, 侧根数目减少, 不定根变短(Huang et al., 2015). ...
ARL1, a LOB-domain protein required for adventitious root formation in rice 1 2005
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
Adventitious root formation in rice requires OsGNOM1 and is mediated by the OsPINs family 1 2009
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
Roles of OsCKI1, a rice casein kinase I, in root development and plant hormone sensitivity 1 2003
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
OsPIN5b modulates rice (Oryza sativa) plant architecture and yield by changing auxin homeostasis, transport and distribution 2 2015
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
... 在众多已克隆的水稻根系基因中, 有一部分基因不仅调控根系发育, 还调控其它性状, 即一因多效基因.例如, Rf1a/Rf-1/Rf5转基因植物表现为根系发达, 主根变长, 侧根数增加(Zhang et al., 2016).研究表明, 该基因还可以提高自身的抗逆性, 使水稻的耐旱性和耐盐性增强.此外, 携带有Rf-1基因的胞质雄性不育近等基因系花粉可育, 而无Rf-1基因的胞质雄性不育近等基因系花粉则不育, 表明该基因在一定程度上可以综合调控水稻的生长发育, 最终实现水稻产量与品质的提高.基因OsPIN5b编码一种生长素输出载体蛋白, 该基因通过调节生长素稳态、运输和分布来实现对水稻株型和产量的综合调控(Lu et al., 2015), 调控机制表现为负向调控, 即基因过表达不仅会导致根的生物量明显降低, 而且导致株高、叶片和分蘖数、茎生物量、结实率、穗长以及产量参数等均降低, 减少表达则有相反的效果.CRL6基因在水稻根、茎和叶等多个组织中表达, 尤其是在控制根冠生长的区域表达水平最高.该基因突变体表现为冠状根减少、株高变矮、旗叶变短变窄、穗变小和结实率降低(Wang et al., 2016). ...
Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.) 17 2002
... 国际水稻研究所数据库中提供的3 024个水稻品种的62个性状中没有一个根系性状(Shrestha et al., 2014).这表明在水稻育种计划中缺乏可用的根系特征数据资源, 且根系性状并没有得到广泛应用.由于根属于地下部分且结构庞大复杂, 难以进行表型分析, 通过常规育种对根系进行遗传改良具有较大困难(Nada et al., 2019).数量性状定位(QTL)及其在标记辅助育种(MAB)中的应用成为根系研究中具有突破性的方法(Coudert et al., 2010).近年来, 利用分子标记辅助选择技术对水稻根性状QTL的遗传改良逐渐增多(Temnykh et al., 2000; McCouch et al., 2002; 胡兴明, 2004; 韩龙植, 2005; 吴伟明, 2006; 翟荣荣, 2012; 徐晓明等, 2016; 曲志恒, 2016; 索艺宁等, 2018).此外, 自1996年超级稻育种技术路线提出以来, 株型改良与杂种优势相结合的作物育种方式, 其科学性与正确性已经过充分的实践检验.中国的超级杂交水稻育种计划于2016年已实现第五期育种产量目标(周正平等, 2019), 以QTL定位为基础的水稻理想根系育种必将会不断助力水稻增产、稳产潜力的挖掘. ...
Ubiquitin ligase EL5 maintains the viability of root meristems by influencing cytokinin-mediated nitrogen effects in rice 1 2014
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
The interaction of genes controlling root traits is required for the developmental acquisition of deep and thick root traits and improving root architecture in response to low water or nitrogen content in rice (Oryza sativa L.) cultivars 1 2019
... 国际水稻研究所数据库中提供的3 024个水稻品种的62个性状中没有一个根系性状(Shrestha et al., 2014).这表明在水稻育种计划中缺乏可用的根系特征数据资源, 且根系性状并没有得到广泛应用.由于根属于地下部分且结构庞大复杂, 难以进行表型分析, 通过常规育种对根系进行遗传改良具有较大困难(Nada et al., 2019).数量性状定位(QTL)及其在标记辅助育种(MAB)中的应用成为根系研究中具有突破性的方法(Coudert et al., 2010).近年来, 利用分子标记辅助选择技术对水稻根性状QTL的遗传改良逐渐增多(Temnykh et al., 2000; McCouch et al., 2002; 胡兴明, 2004; 韩龙植, 2005; 吴伟明, 2006; 翟荣荣, 2012; 徐晓明等, 2016; 曲志恒, 2016; 索艺宁等, 2018).此外, 自1996年超级稻育种技术路线提出以来, 株型改良与杂种优势相结合的作物育种方式, 其科学性与正确性已经过充分的实践检验.中国的超级杂交水稻育种计划于2016年已实现第五期育种产量目标(周正平等, 2019), 以QTL定位为基础的水稻理想根系育种必将会不断助力水稻增产、稳产潜力的挖掘. ...
Production and characterization of auxin-insensitive rice by overexpression of a mutagenized rice IAA protein 1 2006
... The cloned root-related genes in rice Table 4
基因
对应基因编码的蛋白
对应的突变体表型
参考文献
Dro1
Dro1
深根
Clark et al., 2013
Rf1a/Rf-1/Rf5
三角状五肽重复区蛋白
主根变长, 侧根数增加, 育性恢复
Zhang et al., 2016
CRL1
1个冠状根和侧根形成的正向调节因子
冠状根的发生
Inukai et al., 2005
ARL1
1个含有LOB结构域的蛋白
不定根的形成
Liu et al., 2005
OsCKI1
I型酪蛋白激酶
较短的初生根、侧根及不定根数少
Liu et al., 2003
WOX11
1个由262氨基酸组成的蛋白产物
冠状根数增加, 产生不定根
Zhao et al., 2015
Crl4
1个拟南芥GNOM的同源蛋白
冠状根的形成
Kitomi et al., 2008
OsGNOM1
ADP核糖基化因子的鸟苷酸交换因子
不定根的形成
Liu et al., 2009
OsRAA1
1个12 kDa的小G蛋白
主根生长被抑制, 不定根增加
Chen et al., 2018a
OsPIN1
生长素输出载体
抑制不定根的发生
Xu et al., 2014
OsCAND1
1个拟南芥CAND1的同源基因
抑制冠状根形成
Wang et al., 2011
OsAHP1
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsAHP2
组氨酸磷酸转移蛋白
侧根生长, 育性降低
Sun et al., 2014
OsCYP2
肽基脯氨酰顺反异构酶
侧根丧失, 育性降低
Kumari et al., 2015
LRT2
亲环素蛋白
侧根减少
Jing et al., 2015
OsPIN5b
生长素输出载体
降低根生物量、结实率以及产量
Lu et al., 2015
OsSAP16
1个胁迫相关的蛋白
根生物量降低, 根系结构变小
Wang et al., 2016
SOR1
1个E3泛素连接酶
形成地表根, 抑制根正常生长
Chen et al., 2018b
OsIAA3
水稻Aux/IAA蛋白
冠状根数目减少
Nakamura et al., 2006
EL5
泛素连接酶
无根, 冠状根变短, 侧根坏死
Mochizuki et al., 2014
Crl-5
1个AP2/ERF转录因子家族成员
不定根发生
Kitomi et al., 2011
CHR729
染色质域解旋酶DNA结合蛋白
抑制根生长
Xu et al., 2017
CRL6
SNF2家族蛋白
冠状根减少, 结实率降低
Wang et al., 2016
OsIAA11
水稻Aux/IAA蛋白
抑制侧根发育
Jing et al., 2015
OsERF3
乙烯应答因子
促进冠状根发育
Zhao et al., 2015
OsWRKY74
WRKY转录因子
根伸长, 不定根增加, 产量增加
Dai et al., 2016
OsWOX3A
WUSCHEL相关的同源框蛋白
侧根数目减少, 结实率降低
Cho et al., 2016
OsNAR2.1
硝酸盐转运蛋白伴侣蛋白
根长/不定根变短, 侧根数目减少
Huang et al., 2015
OsCKX4
细胞分裂素氧化酶/脱氢酶
冠状根生长
Gao et al., 2014
REL2
1个包含DUF结构域的蛋白
不定根减少且变短
Yang et al., 2016
已克隆基因中也有许多与根数目相关.已经检测到如OsWOX3A基因在器官发育中起重要作用, 包括叶片侧向生长和叶脉形成、叶柄和外稃的形态发生以及分蘖和侧根数量(Cho et al., 2013).rel2突变体表现为不定根数目减少且变短, 水分和营养物质的运输受到抑制, 该突变体地上部分表现为叶片中叶绿素a和b含量显著升高, 光合效率轻微升高, 植株分蘖数减少, 圆粒, 每穗粒数减少(Yang et al., 2016).由于水稻根直径、根表面积和根体积等性状表现值考察难度较大, 尚未实现更多相关性状调控基因的克隆. ...
Genetic variation and QTLs related to root development in upland New Rice for Africa (NERICA) varieties 3 2019
 ,1,*, 王跃星
,1,*, 王跃星
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}