A Role of Arabidopsis Phosphoinositide Kinase, FAB1, in Root Hair Growth
Yuting Yao, Jiaqi Ma, Xiaoli Feng, Jianwei Pan, Chao Wang,*Ministry of Education Key Laboratory of Cell Activities and Stress Adaptations, School of Life Sciences, Lanzhou University, Lanzhou 730000, China
Abstract Lipid kinase FORMATION of APLOID and BINUCLEATE CELLS 1 (FAB1/PIKfyve) is a key enzyme that generates PI(3,5)P2. PI(3,5)P2 plays an important role in regulating membrane trafficking in yeast and animal cells, but its function in plants remains poorly understood. Here we study the functional role of PI(3,5)P2 via analysis of phenotypes of Arabidopsis FAB1 T-DNA knockout lines. The Arabidopsis FAB1 family contains four orthologous genes: FAB1A, FAB1B, FAB1C, and FAB1D. The fab1a/b double mutant showed a complete male gametophyte lethal phenotype. fab1b/c/d mutant was successfully isolated. Genetic analyses showed that the loss of FAB1B, FAB1C and FAB1D function disrupts root hairs elongation. Further pharmacological analysis showed that the FAB1-specific inhibitor YM201636 inhibited the root hairs growth. In addition, the transcription level of DR5-GFP, an indicator of auxin expression and distribution, was downregulated in single mutation of FAB1B, FAB1C and FAB1D. Moreover, triple mutant phenotypes (short root hairs) were partially rescued by exogenous application of auxin analog 2,4-D and NAA. However, the mutant’s sensitivity to 1-NOA (auxin influx inhibitor) and TIBA (auxin efflux inhibitor) in root hair elongation assay is identical to that of the wild-type. Furthermore, loss of FAB1B, FAB1C and FAB1D function reduces the level of reactive oxygen species (ROS) and affects actin expression in root hairs. Taken together, the FAB1B/C/D affects Arabidopsis root hair elongation by regulating auxin distribution, levels of ROS, and actin expression. Keywords:FAB1;root hair;auxin;ROS;actin;Arabidopsis thaliana
PDF (2793KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 姚玉婷, 马家琦, 冯晓莉, 潘建伟, 王超. 磷酸肌醇激酶FAB1调控拟南芥根毛伸长. 植物学报, 2020, 55(2): 126-136 doi:10.11983/CBB19242 Yao Yuting, Ma Jiaqi, Feng Xiaoli, Pan Jianwei, Wang Chao. A Role of Arabidopsis Phosphoinositide Kinase, FAB1, in Root Hair Growth. Chinese Bulletin of Botany, 2020, 55(2): 126-136 doi:10.11983/CBB19242
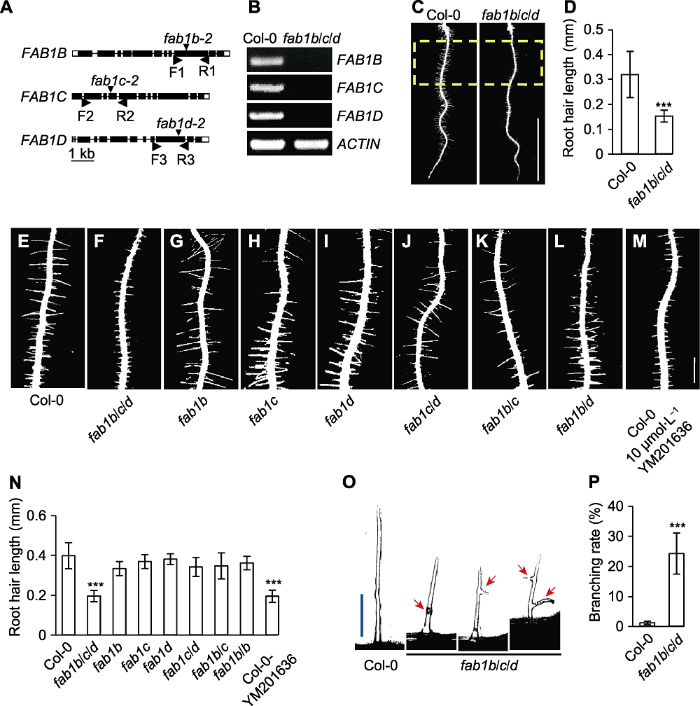
(A) FAB1 gene structure and T-DNA insertion sites; (B) Analysis of gene expression of FAB1B, FAB1C and FAB1D by RT-PCR; (C) Assay of root hair length in 5-day-old seedlings (the yellow dotted box represents quantitative area) (Bar=5 mm); (D) Quantification of root hair length; (E)-(M) The root hair phenotype of the FAB1 single mutants, double mutants and YM201636 (FAB1-specific inhibitor) treatment of Col-0 seedling (Bar=0.5 mm); (N) Quantification of (E)-(M) mutant root hair length; (O) Images of the typical root hair morphologies, the root hair morphologies were categorized as swollen or branched (red arrow indicates) (Bar=75 μm); (P) Quantification of frequency of branched root hairs (%). *** P<0.001 (Student’s t-test)
(A) Growth dynamics of individual Col-0 and fab1b/c/d root hairs (fluorescence microscopy was used to assess root hair elongation, showing consecutive frames of growing root hairs for a period of 50 minutes, pictures were taken every 5 minutes) (Bar=50 µm); (B) Root hair length per unit time; (C) Root hairs growth speed (*** P<0.001, Student’s t-test); (D) Average root hairs number.
2.3 FAB1影响生长素的分布
前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019)。为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天。经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C)。有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D)。上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成。DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布。由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B)。这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导。
(A) 用DCF-DA (ROS染液)检测Col-0、fab1b/c/d和Col-0- YM201636根毛中的ROS含量(Bar=25 µm); (B) 定量分析DCF-DA荧光强度; (C) Col-0、fab1b/c/d和Col-0-YM201636的根毛肌动蛋白细胞骨架标记ABD2的分布(Bar=25 µm); (D) 定量分析ABD2-GFP荧光强度。*** P<0.001 (Student’s t-test) Figure 5Reactive oxygen species (ROS) intensity and actin stability were altered in root hairs of Arabidopsis fab1b/c/d seedlings
(A) Total ROS generated by oxidation of DCF-DA in wild type, fab1b/c/d and Col-0-YM201636 root (Bar=25 µm); (B) Relative intensity of the GFP signals; (C) Distribution of actin cytoskeleton marker ABD2 in root hair of Col-0, fab1b/c/d and Col-0-YM201636 treatment (Bar=25 µm); (D) Average relative intensity of the ABD2-GFP signals. *** P<0.001 (Student’s t-test)
CuiSK, SuzakiT, Tominaga-WadaR, YoshidaS ( 2018). Regulation and functional diversification of root hairs Semin Cell Dev Biol 83, 115-122. [本文引用: 2]
De CraeneJO, BertazziD, B?rS, FriantS ( 2017). Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways Int J Mol Sci 18, 634. [本文引用: 1]
DharmasiriN, DharmasiriS, WeijersD, LechnerE, YamadaM, HobbieL, EhrismannJS, JürgensG, EstelleM ( 2005). Plant development is regulated by a family of auxin receptor F box proteins Dev Cell 9, 109-119. [本文引用: 1]
DoveSK, DongKZ, KobayashiT, WilliamsFK, MichellRH ( 2009). Phosphatidylinositol 3,5-bisphosphate and Fab1p/PIKfyve under PPIn endo-lysosome function Biochem J 419, 1-13. [本文引用: 2]
GangulyA, LeeSH, ChoM, LeeOR, YooH, ChoHT ( 2010). Differential auxin-transporting activities of PIN- FORMED proteins in Arabidopsis root hair cells Plant Physiol 153, 1046-1061. [本文引用: 2]
GaryJD, WurmserAE, BonangelinoCJ, WeismanLS, EmrSD ( 1998). Fab1p is essential for PtdIns(3)P5-kinase activity and the maintenance of vacuolar size and membrane homeostasis J Cell Biol 143, 65-79. [本文引用: 1]
HiranoT, SteckerK, MunnikT, XuHX, SatoMH ( 2017). Visualization of phosphatidylinositol 3,5-bisphosphate dynamics by a tandem ML1N-based fluorescent protein probe in Arabidopsis Plant Cell Physiol 58, 1185-1195. [本文引用: 1]
IshidaT, KurataT, OkadaK, WadaT ( 2008). A genetic regulatory network in the development of trichomes and root hairs Annu Rev Plant Biol 59, 365-386. [本文引用: 2]
JinN, LangMJ, WeismanLS ( 2016). Phosphatidylinositol 3,5-bisphosphate: regulation of cellular events in space and time Biochem Soc Trans 44, 177-184. [本文引用: 1]
JonesAR, KramerEM, KnoxK, SwarupR, BennettMJ, LazarusCM, LeyserHMO, GriersonCS ( 2009). Auxin transport through non-hair cells sustains root-hair development Nat Cell Biol 11, 78-84. [本文引用: 2]
JonesMA, RaymondMJ, YangZB, SmirnoffN ( 2007). NADPH oxidase-dependent reactive oxygen species formation required for root hair growth depends on ROP GTPase J Exp Bot 58, 1261-1270. [本文引用: 1]
KetelaarT, de RuijterNCA, EmonsAMC ( 2003). Unstable F-Actin specifies the area and microtubule direction of cell expansion in Arabidopsis root hairs Plant Cell 15, 285-292. [本文引用: 2]
KirschSA, KugemannA, CarpanetoA, B?ckmannRA, DietrichP ( 2018). Phosphatidylinositol-3,5-bisphosphate lipid-binding-induced activation of the human two-pore channel 2 Cell Mol Life Sci 75, 3803-3815. [本文引用: 1]
KusanoH, TesterinkC, VermeerJEM, TsugeT, ShimadaH, OkaA, MunnikT, AoyamaT ( 2008). The Arabidopsis phosphatidylinositol phosphate 5-Kinase PIP5K3 is a key regulator of root hair tip growth Plant Cell 20, 367-380. [本文引用: 1]
LeeRDW, ChoHT ( 2013). Auxin, the organizer of the hormonal/environmental signals for root hair growth Front Plant Sci 4, 448. [本文引用: 1]
MendrinnaA, PerssonS ( 2015). Root hair growth: it's a one way street F1000Prime Rep 7, 23. [本文引用: 1]
MonshausenGB, BibikovaTN, MesserliMA, ShiC, GilroyS ( 2007). Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs Proc Natl Acad Sci USA 104, 20996-21001. [本文引用: 1]
Mueller-RoeberB, PicalC ( 2002). Inositol phospholipid metabolism in Arabidopsis Characterized and putative isoforms of inositol phospholipid kinase and phosphoinositide-specific phospholipase C. Plant Physiol 130, 22-46. [本文引用: 1]
NakamuraM, ClaesAR, GrebeT, HermkesR, ViottiC, IkedaY, GrebeM ( 2018). Auxin and ROP GTPase signaling of polar nuclear migration in root epidermal hair cells Plant Physiol 176, 378-391. [本文引用: 2]
NishimuraT, HayashiK, SuzukiH, GyohdaA, TakaokaC, SakaguchiY, MatsumotoS, KasaharaH, SakaiT, KatoJ, KamiyaY, KoshibaT ( 2014). Yucasin is a potent inhibitor of YUCCA, a key enzyme in auxin biosynthesis Plant J 77, 352-366. [本文引用: 1]
NiuYF, JinCW, JinGL, ZhouQY, LinXY, TangCX, ZhangYS ( 2011). Auxin modulates the enhanced development of root hairs in Arabidopsis thaliana(L.) Heynh. under elevated CO2 Plant Cell Environ 34, 1304-1317. [本文引用: 1]
PayrastreB, MissyK, GiuriatoS, BosinS, PlantavidM, GratacapMP ( 2001). Phosphoinositides: key players in cell signaling, in time and space Cell Signal 13, 377-387. [本文引用: 1]
PeiWK, DuF, ZhangY, HeT, RenHY ( 2012). Control of the actin cytoskeleton in root hair development Plant Sci 187, 10-18. [本文引用: 1]
QinH, HuangRF ( 2018). Auxin controlled by ethylene steers root development Int J Mol Sci 19, 3656. [本文引用: 2]
RahmanA, HosokawaS, OonoY, AmakawaT, GotoN, TsurumiS ( 2002). Auxin and ethylene response interactions during Arabidopsis root hair development dissected by auxin influx modulators Plant Physiol 130, 1908-1917. [本文引用: 1]
SchoenaersS, BalcerowiczD, BreenG, HillK, ZdanioM, MouilleG, HolmanTJ, OhJ, WilsonMH, NikonorovaN, VuLD, De SmetI, SwarupR, De VosWH, PintelonI, AdriaensenD, GriersonC, BennettMJ, VissenbergK ( 2018). The auxin-regulated CrRLK1L kinase ERULUS controls cell wall composition during root hair tip growth Curr Biol 28, 722-732. [本文引用: 2]
SerrazinaS, DiasFV, MalhóR ( 2014). Characterization of FAB1 phosphatidylinositol kinases in Arabidopsis pollen tube growth and fertilization New Phytol 203, 784-793. [本文引用: 3]
ShibataM, SugimotoK ( 2019). A gene regulatory network for root hair development J Plant Res 132, 301-309. [本文引用: 2]
StenzelI, IschebeckT, K?nigS, HolubowskaA, SporyszM, HauseB, HeilmannI ( 2008). The type B phosphatidylinositol-4-phosphate 5-kinase 3 is essential for root hair formation in Arabidopsis thaliana Plant Cell 20, 124-141. [本文引用: 1]
TakasugaS, HorieY, SasakiJ, Sun-WadaGH, KawamuraN, IizukaR, MizunoK, EquchiS, KofujiS, KimuraH, YamazakiM, HorieC, OdanagaE, SatoY, ChidaS, KontaniK, HaradaA, KatadaT, SuzukiA, WadaY, OhnishiH, SasakiT ( 2013). Critical roles of type III phosphatidylinositol phosphate kinase in murine embryonic visceral endoderm and adult intestine Proc Natl Acad Sci USA 110, 1726-1731. [本文引用: 2]
VijayakumarP, DattaS, DolanL ( 2016). ROOT HAIR DEFECTIVE SIX-LIKE 4 (RSL4) promotes root hair elongation by transcriptionally regulating the expression of genes required for cell growth New Physiol 212, 944-953. [本文引用: 1]
WhitleyP, HinzS, DoughtyJ ( 2009). Arabidopsis FAB1/PIKfyve proteins are essential for development of viable pollen Plant Physiol 151, 1812-1822. [本文引用: 7]
ZhangYL, LiE, FengQN, ZhaoXY, GeFR, ZhangY, LiS ( 2015). Protein palmitoylation is critical for the polar growth of root hairs in Arabidopsis BMC Plant Biol 15, 50. [本文引用: 1]
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
Rapid structural changes and acidification of guard cell vacuoles during stomatal closure require phosphatidylinositol 3,5-bisphosphate 3 2013
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... ; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Phosphoinositides: tiny lipids with giant impact on cell regulation 1 2013
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
The role of reactive oxygen species in cell growth: lessons from root hairs 1 2006
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
Dissecting Arabidopsis lateral root development 1 2003
Regulation and functional diversification of root hairs 2 2018
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019).为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天.经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C).有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D).上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成.DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布.由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B).这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导. ...
Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways 1 2017
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Plant development is regulated by a family of auxin receptor F box proteins 1 2005
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Phosphatidylinositol 3,5-bisphosphate and Fab1p/PIKfyve under PPIn endo-lysosome function 2 2009
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... , 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Differential auxin-transporting activities of PIN- FORMED proteins in Arabidopsis root hair cells 2 2010
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
... ; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
Fab1p is essential for PtdIns(3)P5-kinase activity and the maintenance of vacuolar size and membrane homeostasis 1 1998
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Root hairs 3 2014
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
PI5P and PI(3,5)P2: minor, but essential phosphoinositides 1 2017
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
PtdIns(3,5)P2 mediates root hair shank hardening in Arabidopsis 2 2018
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Loss-of-function and gain-of-function mutations in FAB1A/B impair endomembrane homeostasis, conferring pleiotropic developmental abnormalities in Arabidopsis 2 2011
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... FAB1是催化PI(3)P合成PI(3,5)P2的磷脂激酶, 几乎存在于所有真核生物中(Jin et al., 2016).研究表明, 拟南芥中有4个FAB1蛋白(FAB1A、FAB1B、FAB1C和FAB1D)与酵母和人类(Homo sapiens)中的FAB1蛋白序列同源(Whitley et al., 2009).为深入解析FAB1在拟南芥生长发育过程中的生物学功能, 我们分离鉴定了fab1 T-DNA插入突变体, 并分别获得各个单、双、三突变体(图1A).其中fab1a/b双突变体表现雄配子体致死(Whitley et al., 2009), fab1b-2、fab1c-2和fab1d-2的单突变体和双突变体的根毛表型与野生型相比无显著差异(图1E-L, N), 而fab1b- 2/fab1c-2/fab1d-2 (fab1b/c/d)三突变体呈现非常明显的短根毛表型(图1C), 根毛长度约为野生型的50% (图1D).RT-PCR结果表明, 在纯合的fab1b/c/d三突变体株系中未检测到FAB1B、FAB1C和FAB1D的表达, 表明这3个基因均完全缺失(图1B), 暗示FAB1B、FAB1C和FAB1D之间存在功能冗余, 这与前人的研究(拟南芥FAB1基因功能冗余)相一致(Whitley et al., 2009; Hirano et al., 2011).为进一步验证fab1b/c/d三突变体根毛变短是否由FAB1的功能缺陷造成, 我们使用FAB1特异性抑制剂YM201636进行药理学实验, 发现含有YM201636平板上的拟南芥根毛明显短于对照组(图1M).我们还发现fab1b/c/d三突变体的根毛形态也发生变化, 出现了分叉和局部膨大的根毛(图1O).在fab1b/c/d背景下, 具有分叉表型的根毛约占25% (野生型中仅占不到1%) (图1P).综上表明, FAB1B、FAB1C和FAB1D以功能冗余的方式参与根毛的生长发育. ...
Phosphatidylinositol 3-phosphate 5-kinase, FAB1/PIKfyve kinase mediates endosome maturation to establish endosome-cortical microtubule interaction in Arabidopsis 2 2015
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
... 功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Visualization of phosphatidylinositol 3,5-bisphosphate dynamics by a tandem ML1N-based fluorescent protein probe in Arabidopsis 1 2017
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
A genetic regulatory network in the development of trichomes and root hairs 2 2008
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
Phosphatidylinositol 3,5-bisphosphate: regulation of cellular events in space and time 1 2016
... FAB1是催化PI(3)P合成PI(3,5)P2的磷脂激酶, 几乎存在于所有真核生物中(Jin et al., 2016).研究表明, 拟南芥中有4个FAB1蛋白(FAB1A、FAB1B、FAB1C和FAB1D)与酵母和人类(Homo sapiens)中的FAB1蛋白序列同源(Whitley et al., 2009).为深入解析FAB1在拟南芥生长发育过程中的生物学功能, 我们分离鉴定了fab1 T-DNA插入突变体, 并分别获得各个单、双、三突变体(图1A).其中fab1a/b双突变体表现雄配子体致死(Whitley et al., 2009), fab1b-2、fab1c-2和fab1d-2的单突变体和双突变体的根毛表型与野生型相比无显著差异(图1E-L, N), 而fab1b- 2/fab1c-2/fab1d-2 (fab1b/c/d)三突变体呈现非常明显的短根毛表型(图1C), 根毛长度约为野生型的50% (图1D).RT-PCR结果表明, 在纯合的fab1b/c/d三突变体株系中未检测到FAB1B、FAB1C和FAB1D的表达, 表明这3个基因均完全缺失(图1B), 暗示FAB1B、FAB1C和FAB1D之间存在功能冗余, 这与前人的研究(拟南芥FAB1基因功能冗余)相一致(Whitley et al., 2009; Hirano et al., 2011).为进一步验证fab1b/c/d三突变体根毛变短是否由FAB1的功能缺陷造成, 我们使用FAB1特异性抑制剂YM201636进行药理学实验, 发现含有YM201636平板上的拟南芥根毛明显短于对照组(图1M).我们还发现fab1b/c/d三突变体的根毛形态也发生变化, 出现了分叉和局部膨大的根毛(图1O).在fab1b/c/d背景下, 具有分叉表型的根毛约占25% (野生型中仅占不到1%) (图1P).综上表明, FAB1B、FAB1C和FAB1D以功能冗余的方式参与根毛的生长发育. ...
Auxin transport through non-hair cells sustains root-hair development 2 2009
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
NADPH oxidase-dependent reactive oxygen species formation required for root hair growth depends on ROP GTPase 1 2007
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
Unstable F-Actin specifies the area and microtubule direction of cell expansion in Arabidopsis root hairs 2 2003
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
Phosphatidylinositol-3,5-bisphosphate lipid-binding-induced activation of the human two-pore channel 2 1 2018
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
The Arabidopsis phosphatidylinositol phosphate 5-Kinase PIP5K3 is a key regulator of root hair tip growth 1 2008
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
Auxin, the organizer of the hormonal/environmental signals for root hair growth 1 2013
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
PINOID positively regulates auxin efflux in Arabidopsis root hair cells and tobacco cells 1 2006
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
Roles of phosphatidylinositol 3-kinase in root hair growth 2 2008
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
Molecular link between auxin and ROS-mediated polar growth 1 2017
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
Phosphatidylinositol 3,5-bisphosphate: low abundance, high significance 3 2014
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... ; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Root hair growth: it's a one way street 1 2015
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs 1 2007
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
Inositol phospholipid metabolism in Arabidopsis 1 2002
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Auxin and ROP GTPase signaling of polar nuclear migration in root epidermal hair cells 2 2018
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019).为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天.经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C).有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D).上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成.DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布.由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B).这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导. ...
Yucasin is a potent inhibitor of YUCCA, a key enzyme in auxin biosynthesis 1 2014
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
Auxin modulates the enhanced development of root hairs in Arabidopsis thaliana(L.) Heynh. under elevated CO2 1 2011
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
Phosphoinositides: key players in cell signaling, in time and space 1 2001
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
Control of the actin cytoskeleton in root hair development 1 2012
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
Auxin controlled by ethylene steers root development 2 2018
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019).为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天.经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C).有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D).上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成.DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布.由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B).这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导. ...
Auxin and ethylene response interactions during Arabidopsis root hair development dissected by auxin influx modulators 1 2002
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
The auxin-regulated CrRLK1L kinase ERULUS controls cell wall composition during root hair tip growth 2 2018
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
... 前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019).为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天.经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C).有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D).上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成.DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布.由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B).这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导. ...
Characterization of FAB1 phosphatidylinositol kinases in Arabidopsis pollen tube growth and fertilization 3 2014
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
... ).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
A gene regulatory network for root hair development 2 2019
... 前人的研究表明, 生长素参与根毛生长发育的多个过程(Cui et al., 2018; Nakamura et al., 2018; Qin and Huang, 2018; Schoenaers et al., 2018; Shibata and Sugimoto, 2019).为了探究FAB1在生长素调控根毛发育过程中的作用, 我们将萌发后3天的野生型和fab1b/c/d突变体幼苗转移到含有NAA或2,4-D的1/2MS培养基上培养3天.经过生长素类似物(NAA和2,4-D)处理后, 野生型和fab1b/c/d突变体的根毛相比对照组均明显伸长(图3A-C).有趣的是, 2,4-D处理后突变体的根毛长度与野生型之间无显著差异(图3B, D), 而NAA处理后三突变体与野生型之间的差异缩小(图3C, D).上述结果表明, fab1b/c/d短根毛表型可能是生长素不足造成.DR5是生长素反应的强启动子, 通过分析DR5-GFP和DR5-GUS (DR5-GFP/GUS)的表达可间接反映生长素的水平与分布.由于直接检测根毛细胞中的DR5-GFP/GUS比较困难, 因此为了测定FAB1在生长素分布中的作用, 我们观察了拟南芥根尖DR5-GFP/GUS的分布, 结果发现fab1b/c/d突变体中DR5-GFP/GUS表达水平下降(图4A, B).这一结果同样暗示FAB1可能调控拟南芥根毛伸长过程中生长素的分布或影响生长素信号转导. ...
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
The type B phosphatidylinositol-4-phosphate 5-kinase 3 is essential for root hair formation in Arabidopsis thaliana 1 2008
... 肌动蛋白细胞骨架参与根毛发育的所有阶段(根毛起始、伸长和成熟), 并在不同阶段呈现不同的排布(Ketelaar et al., 2003; Griersona et al., 2014).此外, 肌动蛋白结合蛋白介导磷酸肌醇调控细胞骨架(Pei et al., 2012).抑制PI(4,5)P2合成(抑制剂mastoparan处理)和敲除PI(4,5)P2合成酶(pip5k3突变体)都造成根毛显著变短(Kusano et al., 2008; Stenzel et al., 2008), 其它磷脂(如PA、PI3P和PI4P)也参与根毛尖端伸长(Ishida et al., 2008).在fab1b/c/d突变体和YM201636处理的拟南芥中, 肌动蛋白Marker (ABD2- GFP)表达量显著下降(图5C, D), 结合肌动蛋白在根毛发育中的作用, 暗示FAB1可能调控微丝的动态解聚和聚合, 进而介导根毛伸长. ...
Critical roles of type III phosphatidylinositol phosphate kinase in murine embryonic visceral endoderm and adult intestine 2 2013
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Auxin and cellular elongation 2 2016
... 对生长素信号元件和多种生长素转运体的研究表明, 根毛细胞的生长素信号转导和稳态对根毛伸长至关重要(Velasquez et al., 2016).生长素信号受体(Transport inhibitor response 1)突变体tir1及其afb1、afb2和afb3 (auxin signaling f-box, AFB)突变体均表现出根毛伸长缺陷(Dharmasiri et al., 2005), 而根毛特异性过表达TIR1可提高根毛伸长速率(Ganguly et al., 2010).生长素抗性突变体iaa7、iaa17和iaa28 (indole-3-acid protein, IAA)都表现出根毛伸长受抑制(Lee and Cho, 2013).而且, 生长素信号突变体arf7/arf19也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
... 也表现出短根毛表型(Velasquez et al., 2016).此外, 生长素生物合成抑制剂yucasin处理会抑制拟南芥根毛伸长(Nishimura et al., 2014).在fab1b/c/d三突变体中我们观察到根尖DR5的表达量降低, 说明fab1b/c/d三突变体中生长素分布受到影响(图4A, B).外源施加生长素类似物(2,4-D和NAA)能部分恢复fab1b/c/d突变体根毛伸长的缺陷(图3B, C), 暗示FAB1可能通过影响根毛生长素分布或生长素信号转导途径调控根毛伸长.同时, 生长素只能部分恢复突变体的表型, 这增加了生长素运输缺陷的可能性.已有研究表明, AUX1介导非生毛细胞生长素运输维持根毛发育(Jones et al., 2009); 在生毛细胞中过表达6种PIN (Pin-formed)蛋白(PIN1-PIN4、PIN7和PIN8)和3种PGP (Phosphoglycoprotein)蛋白(PGP1、PGP4和PGP19)都极大地抑制根毛细胞的伸长, 这可能是由于它们的高生长素输出活性降低了生毛细胞的生长素水平(Lee and Cho, 2006; Ganguly et al., 2010); 此外, 生长素输出抑制剂TIBA和NPA通过抑制生长素的输出增加内源IAA的积累, 进而促进根毛伸长(Niu et al., 2011).反之, 生长素输入抑制剂1-NOA则抑制根毛伸长(Rahman et al., 2002).药理学实验结果表明, 用TIBA和1-NOA分别处理对野生型和fab1b/c/d根毛伸长的影响相似(图3E, F).上述结果表明, FAB1调控根毛发育并不是通过影响生长素运输, 而是干扰了生长素的分布.同样, fab1b/c/d突变体生长素分布的变化(图4A, B)也可能是由于FAB1的缺失打破了生长素合成和降解的平衡, 但还需要通过实验证实. ...
ROOT HAIR DEFECTIVE SIX-LIKE 4 (RSL4) promotes root hair elongation by transcriptionally regulating the expression of genes required for cell growth 1 2016
... 根毛伸长除了与生长素有关外, 也与ROS密切相关(Shibata and Sugimoto, 2019).fab1b/c/d突变体的ROS水平下降(图5A, B), 暗示FAB1对维持根毛细胞ROS水平至关重要.NADPH氧化酶RHD2 (ROOT HAIR DEFECTIVE 2)是根毛ROS产生的主要来源(Lee et al., 2008), 敲除RHD2或突变NADPH氧化酶的转录因子RSL4 (ROOT HAIR DEFFECTIVE SIX-LIKE 4)都表现短根毛表型(Jones et al., 2007; Vijayakumar et al., 2016; Mangano et al., 2017).同时, 外源施加ROS可以部分恢复rhd2突变体的根毛表型(Carol and Dolan, 2006).有研究表明, 在拟南芥中ROS调控PI(3,5)P2后期在体内的积累(Hirano et al., 2017).PI(3,5)P2合成抑制剂处理拟南芥野生型产生短根毛表型(图1M)并降低根毛ROS水平(图5A, B), 这暗示PI(3,5)P2和ROS之间的相互影响是根毛正常发育所必需的. ...
Arabidopsis FAB1/PIKfyve proteins are essential for development of viable pollen 7 2009
... 磷酸肌醇(phosphoinositides, PIs)在20世纪70年代被发现后, 即引起人们极大的研究兴趣(Balla, 2013).虽然磷酸肌醇只占细胞磷脂的一小部分(Payrastre et al., 2001; Mueller-Roeber and Pical, 2002), 但它们几乎控制包括信号转导、细胞骨架重组、膜动力学和囊泡运输在内的各种细胞过程(McCartney et al., 2014).PI(3,5)P2在酵母(Saccharomyces cerevisiae)和哺乳动物中分别占总磷酸肌醇的0.1%和0.04%, 占总磷脂的0.05%-0.1% (Dove et al., 2009; McCartney et al., 2014; Hasegawa et al., 2017).PI(3,5)P2自1997年被发现以来, 虽作为丰度最低的磷酸肌醇, 但可控制内体到溶酶体过程中多个点的囊泡转运, 进而调节溶酶体内腔的大小、形状和酸度(Dove et al., 1997, 2009; De Craene et al., 2017).在酵母和哺乳动物中, PI3P被唯一的FAB1/PIKfyve激酶磷酸化产生PI(3,5)P2 (Gary et al., 1998).在酵母中敲除FAB1导致PI(3,5)P2完全缺失, 造成液泡急剧增大(Kirsch et al., 2018).小鼠(Mus musculus)中PIKfyve功能缺失造成早期胚胎致死(Takasuga et al., 2013).不同于酵母和哺乳动物细胞, 拟南芥(Arabidopsis thaliana)基因组中包含4个FAB1/PIKfyve激酶: FAB1A、FAB1B、FAB1C和FAB1D (Whitley et al., 2009).反向遗传学研究表明, 同时敲除FAB1B和FAB1D影响花粉管伸长并使质膜内吞速度减慢(Serrazina et al., 2014); 而同时敲除FAB1B和FAB1C造成气孔关闭缺陷(Bak et al., 2013); FAB1A和FAB1B功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... 功能缺失则引起花粉致死(Whitley et al., 2009); 而下调FAB1A和FAB1B的表达造成多效性的表型缺陷, 如液泡变大、液泡酸性改变和根毛形态异常(Hirano et al., 2011, 2018; Bak et al., 2013).上述结果充分表明, 在植物细胞中, FAB1同样具有重要的生物学功能.本研究利用遗传学、细胞生物学和药理学等分析方法, 首次证明了FAB1参与调控拟南芥根毛伸长. ...
... FAB1是催化PI(3)P合成PI(3,5)P2的磷脂激酶, 几乎存在于所有真核生物中(Jin et al., 2016).研究表明, 拟南芥中有4个FAB1蛋白(FAB1A、FAB1B、FAB1C和FAB1D)与酵母和人类(Homo sapiens)中的FAB1蛋白序列同源(Whitley et al., 2009).为深入解析FAB1在拟南芥生长发育过程中的生物学功能, 我们分离鉴定了fab1 T-DNA插入突变体, 并分别获得各个单、双、三突变体(图1A).其中fab1a/b双突变体表现雄配子体致死(Whitley et al., 2009), fab1b-2、fab1c-2和fab1d-2的单突变体和双突变体的根毛表型与野生型相比无显著差异(图1E-L, N), 而fab1b- 2/fab1c-2/fab1d-2 (fab1b/c/d)三突变体呈现非常明显的短根毛表型(图1C), 根毛长度约为野生型的50% (图1D).RT-PCR结果表明, 在纯合的fab1b/c/d三突变体株系中未检测到FAB1B、FAB1C和FAB1D的表达, 表明这3个基因均完全缺失(图1B), 暗示FAB1B、FAB1C和FAB1D之间存在功能冗余, 这与前人的研究(拟南芥FAB1基因功能冗余)相一致(Whitley et al., 2009; Hirano et al., 2011).为进一步验证fab1b/c/d三突变体根毛变短是否由FAB1的功能缺陷造成, 我们使用FAB1特异性抑制剂YM201636进行药理学实验, 发现含有YM201636平板上的拟南芥根毛明显短于对照组(图1M).我们还发现fab1b/c/d三突变体的根毛形态也发生变化, 出现了分叉和局部膨大的根毛(图1O).在fab1b/c/d背景下, 具有分叉表型的根毛约占25% (野生型中仅占不到1%) (图1P).综上表明, FAB1B、FAB1C和FAB1D以功能冗余的方式参与根毛的生长发育. ...
... 基因功能冗余)相一致(Whitley et al., 2009; Hirano et al., 2011).为进一步验证fab1b/c/d三突变体根毛变短是否由FAB1的功能缺陷造成, 我们使用FAB1特异性抑制剂YM201636进行药理学实验, 发现含有YM201636平板上的拟南芥根毛明显短于对照组(图1M).我们还发现fab1b/c/d三突变体的根毛形态也发生变化, 出现了分叉和局部膨大的根毛(图1O).在fab1b/c/d背景下, 具有分叉表型的根毛约占25% (野生型中仅占不到1%) (图1P).综上表明, FAB1B、FAB1C和FAB1D以功能冗余的方式参与根毛的生长发育. ...
... 研究表明, 在酵母和动植物中PI(3,5)P2作为FAB1的产物参与调控细胞内膜运输、细胞自噬和逆境响应信号转导(McCartney et al., 2014).大多数真核生物 (如小鼠、果蝇(Drosophila melanogaster)和线虫(Caenorhabditis elegans))中敲除唯一的FAB1基因造成胚胎致死(Takasuga et al., 2013).然而, 拟南芥中却存在4个FAB1基因(FAB1A-D), FAB1的多拷贝暗示FAB1和PI(3,5)P2在拟南芥中的功能可能更加丰富, 遗传学证据也支撑了这种推断.在拟南芥fab1单突变体中并未观察到明显可见的发育异常(图1G-I), 而FAB1A和FAB1B功能缺失都会导致雄配子体致死, FAB1A/FAB1B基因表达下调的突变体则表现生长缓慢、向地性反应异常、生长素敏感性改变以及花器官发育和根毛形态异常(Serrazina et al., 2014; Hirano et al., 2015, 2018); 敲除FAB1B和FAB1C导致气孔关闭障碍(Bak et al., 2013); fab1b/d呈现花粉管形态异常等表型(Whitley et al., 2009).但是, 在拟南芥FAB1的单突变体中并未观察到明显的缺陷表型(图1G-I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
... -I), 说明FAB1之间存在功能冗余(Whitley et al., 2009).本研究成功获得了fab1b/c/d三突变体纯合体、FAB1的单突变体(fab1b、fab1c和fab1d)和双突变体(fab1b/c、fab1b/d和fab1c/d) (图1F-L), 对5天龄幼苗的单、双突变体进行表型分析未见异常发育, 但在三突变体中发现FAB1调控根毛的伸长.在fab1b/c/d三突变体中, 根毛长度约为野生型的50% (图1D).在FAB1特异性抑制剂YM201636处理下, 野生型幼苗的根毛长度受到抑制(图1M).在单根根毛水平上, 利用显微镜成像技术, 我们发现fab1b/c/d植株中根毛的起始是正常的, 但不能完成随后的快速伸长(图2A).已有研究表明, FAB1在真核生物中参与调控内膜运输及在植物中调控花粉管囊泡运输(Serrazina et al., 2014).我们推测FAB1B、FAB1C和FAB1D功能缺失可能干扰了根毛细胞中的内膜系统转运途径, 进而调控根毛的尖端生长.虽然拟南芥FAB1缺失影响花粉管的膜泡运输和根尖PIN2的转运(Hirano et al., 2015), 但是根毛中的囊泡运输情况并没有直接被检测, 还有待深入研究证实. ...
Protein palmitoylation is critical for the polar growth of root hairs in Arabidopsis 1 2015
... 根毛是植物根系的重要组成部分, 具有吸收水分、矿物质营养以及与土壤微生物互作等功能(王立德等, 2004).根毛的快速顶端伸长为研究细胞的极性生长机制提供了很好的模型(Griersona et al., 2014).生长素是根毛顶端伸长的重要因素, 在根毛顶端伸长发育过程中必不可少.生长素信号转导缺陷双突变体arf7/arf9 (auxin response factor, ARF)的根毛相比野生型更短、更少(Qin and Huang, 2018).rhd6 (root hair defective 6)突变体短根毛表型可以通过外源施加生长素IAA得以恢复(Cui et al., 2018).生长素通过调节CrRLK1L激酶ERULUS来控制根尖伸长过程中细胞壁组分的合成(Schoenaers et al., 2018).根毛伸长过程伴随着细胞核向根毛顶端的移动(Zhang et al., 2015), 而根毛中的核迁移依赖F-肌动蛋白(Nakamura et al., 2018).微丝系统如果遭到破坏, 就会抑制根毛伸长(Ketelaar et al., 2003).钙离子(Ca2+)和活性氧(ROS)是根毛伸长过程中2个主要的信号传感器(Mendrinna and Persson, 2015).Ca2+在细胞质中以连续的浓度梯度分布(Ishida et al., 2008), 其浓度变化需要ROS的产生并且需要增加细胞表面的pH来调节, 细胞表面ROS的产生又激活Ca2+流入细胞, 维持胞内Ca2+浓度梯度(Monshausen et al., 2007).因此, 根毛顶端伸长受生长素、Ca2+水平、微丝、pH和ROS等多种因素调控. ...
 ,*兰州大学生命科学学院, 细胞活动与逆境适应教育部重点实验室, 兰州 730000
,*兰州大学生命科学学院, 细胞活动与逆境适应教育部重点实验室, 兰州 730000
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT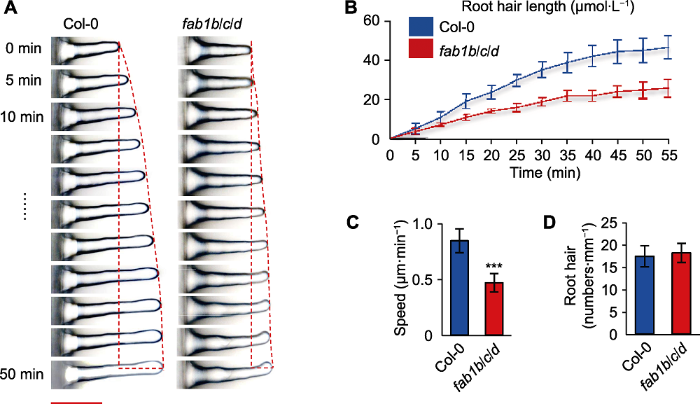 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT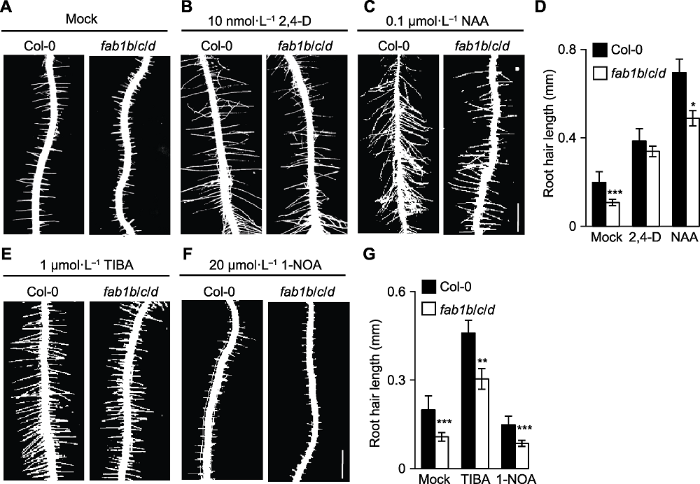 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT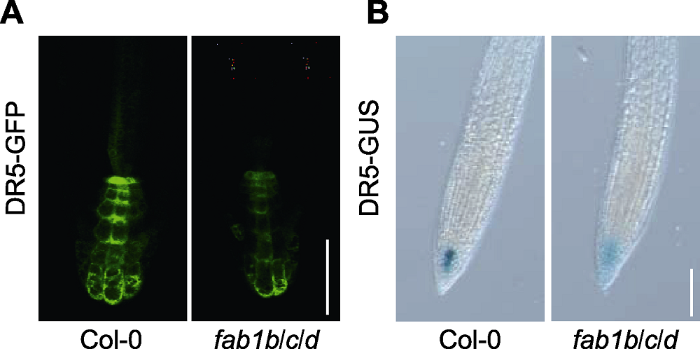 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT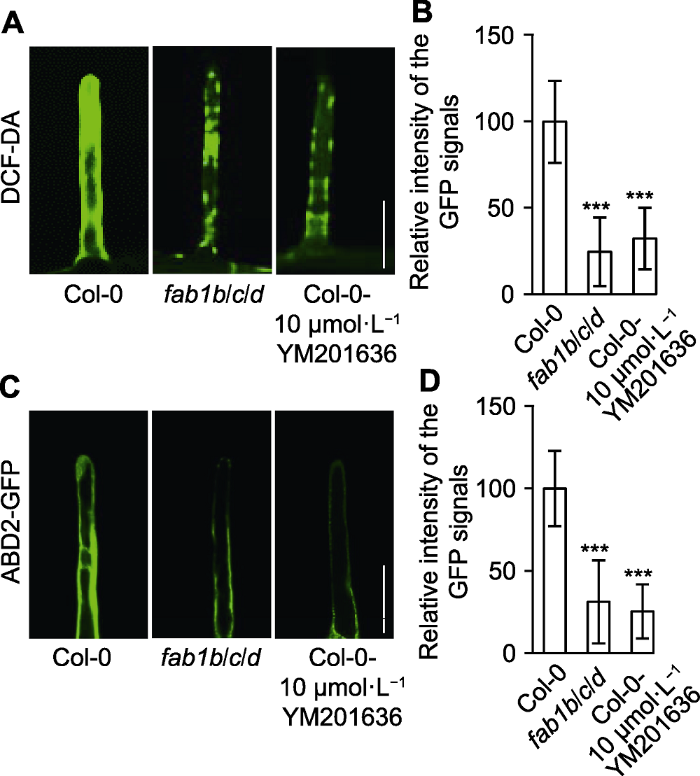 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}