Advances in Studies on the COPT Proteins in Arabidopsis thaliana
Hui Wang1, Jinjin Li2, Jinyu Xu1, Peng Liu1, Haiyan Zhang1,*1 Tianjin Key Laboratory of Animal and Plant Resistance, College of Life Sciences, Tianjin Normal University, Tianjin 300387, China 2 Key Laboratory of Plant Resources, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
Abstract Copper (Cu) is an essential trace element in plants and is involved in many physiological and biochemical reactions as a cofactor of various enzymes. Cu deficiency and excess can affect the normal growth and development, so plants have developed sophisticated regulatory networks to strictly control Cu content. The copper transporter COPT, having high affinity with Cu can regulate the absorption and transport of Cu, and plays an important role in maintaining Cu homeostasis. COPT proteins are involved in different processes of Cu transport, such as uptake of Cu from the external environment, export of Cu from organelles, transport of Cu over long distances, and mobilization and redistribution of Cu between different organs. In addition, COPT proteins play an important role in maintaining homeostasis of other ions, regulating the circadian clock, involving in synthesis of plant hormones and perception of hormone signals. Here we summarize the recent advances in the expression and localization, regulatory mechanisms, and biological functions of COPT proteins in the model plant Arabidopsis thaliana. Keywords:copper transporter;COPT;Cu homeostasis;Cu absorption;redistribution
PDF (1707KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王惠, 李谨谨, 徐金玉, 刘鹏, 张海燕. 拟南芥COPT家族蛋白研究进展. 植物学报, 2019, 54(6): 786-796 doi:10.11983/CBB19045 Wang Hui, Li Jinjin, Xu Jinyu, Liu Peng, Zhang Haiyan. Advances in Studies on the COPT Proteins in Arabidopsis thaliana. Chinese Bulletin of Botany, 2019, 54(6): 786-796 doi:10.11983/CBB19045
Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质。在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中。推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中。HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质。HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中。HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应。HMA5定位于细胞膜, 介导Cu+的外排。 Figure 1Subcellular localization of COPT and copper traffic in Arabidopsis (modified from Rodríguez et al., 1999; Balandin and Castresana, 2002; Wintz et al., 2003; Yruela, 2009; Garcia et al., 2014)
Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+.
3 COPT基因的调控机制
3.1 转录水平的调控
大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响。研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达。用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003)。用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012)。Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异。分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013)。尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018)。
Abdel-GhanySE, Müller-MouléP, NiyogiKK, PilonM, ShikanaiT ( 2005). Two P-type ATPases are required for copper delivery in Arabidopsis thaliana chloroplasts Plant Cell 17, 1233-1251. [本文引用: 1]
Andrés-ColásN, Carrió-SeguíA, Abdel-GhanySE, PilonM, Pe?arrubiaL ( 2018). Expression of the intracellular COPT3-mediated Cu transport is temporally regulated by the TCP16 transcription factor Front Plant Sci 9, 910. [本文引用: 9]
Andrés-ColásN, Perea-GarcíaA, PuigS, Pe?arrubiaL ( 2010). Deregulated copper transport affects Arabidopsis development especially in the absence of environmental cycles Plant Physiol 153, 170-184. [本文引用: 5]
BalandinT, CastresanaC ( 2002). AtCOX17, an Arabidop- sis homolog of the yeast copper chaperone COX17 Plant Physiol 129, 1852-1857. [本文引用: 2]
Bell-PedersenD, ShinoharaML, LorosJJ, DunlapJC ( 1996). Circadian clock-controlled genes isolated from Neurospora crassa are late night-to early morning specific Proc Natl Acad Sci USA 93, 13096-13101. [本文引用: 1]
BernalM, CaseroD, SinghV, WilsonGT, GrandeA, YangHJ, DodaniSC, PellegriniM, HuijserP, ConnollyEL, MerchantSS, Kr?merU ( 2012). Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis Plant Cell 24, 738-761. [本文引用: 1]
BockKW, HonysD, WardJM, PadmanabanS, NawrockiEP, HirschiKD, TwellD, SzeH ( 2006). Integrating mem- brane transport with male gametophyte development and function through transcriptomics Plant Physiol 140, 1151-1168. [本文引用: 1]
BorjiginJ, PayneAS, DengJ, LiXD, WangMM, OvodenkoB, GitlinJD, SnyderSH ( 1999). A novel pineal night- specific ATPase encoded by the Wilson disease gene J Neurosci 19, 1018-1026. [本文引用: 1]
CardonG, H?hmannS, KleinJ, NettesheimK, SaedlerH, HuijserP ( 1999). Molecular characterisation of the Arabidopsis SBP-box genes Gene 237, 91-104. [本文引用: 1]
Carrió-SeguíA, Garcia-MolinaA, SanzA, Pe?arrubiaL ( 2015). Defective copper transport in the copt5 mutant affects cadmium tolerance Plant Cell Physiol 56, 442-454. [本文引用: 1]
Carrió-Seguíà, RomeroP, SanzA, Pe?arrubiaL ( 2016). Interaction between ABA signaling and copper homeostasis in Arabidopsis thaliana Plant Cell Physiol 57, 1568-1582. [本文引用: 1]
CoegoA, BrizuelaE, CastillejoP, RuízS, KonczC, del PozoJC, Pi?eiroM, JarilloJA, Paz-AresJ, LeónJ, The TRANSPLANTAConsortium ( 2014). The TRANSPLANTA collection of Arabidopsis lines: a resource for functional analysis of transcription factors based on their conditional overexpression Plant J 77, 944-953. [本文引用: 1]
GarciaL, WelchenE, GonzalezDH ( 2014). Mitochondria and copper homeostasis in plants Mitochondrion 19, 269-274. [本文引用: 2]
Garcia-MolinaA, Andrés-ColásN, Perea-GarcíaA, Del Valle-TascónS, Pe?arrubiaL, PuigS ( 2011). The intracellular Arabidopsis COPT5 transport protein is required for photosynthetic electron transport under severe copper deficiency Plant J 65, 848-860. [本文引用: 5]
Garcia-MolinaA, Andrés-ColásN, Perea-GarcíaA, NeumannU, DodaniSC, HuijserP, Pe?arrubiaL, PuigS ( 2013). The Arabidopsis COPT6 transport protein functions in copper distribution under copper-deficient conditions Plant Cell Physiol 54, 1378-1390. [本文引用: 7]
GavnholtB, LarsenK ( 2002). Molecular biology of plant laccases in relation to lignin formation Physiol Plant 116, 273-280. [本文引用: 1]
GayombaSR, JungHI, YanJP, DankuJ, RutzkeMA, BernalM, Kr?merU, KochianLV, SaltDE, VatamaniukOK ( 2013). The CTR/COPT-dependent copper uptake and SPL7-dependent copper deficiency responses are required for basal cadmium tolerance in A. thaliana Metallomics 5, 1262-1275. [本文引用: 1]
GoreckaK, CvikrováM, KowalskaU, EderJ, SzafrańskaK, GóreckiR, JanasKM ( 2007). The impact of Cu treatment on phenolic and polyamine levels in plant material regenerated from embryos obtained in anther culture of carrot Plant Physiol Biochem 45, 54-61. [本文引用: 1]
Grat?oPL, PolleA, LeaPJ, AzevedoRA ( 2005). Making the life of heavy metal-stressed plants a little easier Funct Plant Biol 32, 481-494. [本文引用: 1]
JungHI, GayombaSR, RutzkeMA, CraftE, KochianLV, VatamaniukOK ( 2012). COPT6 is a plasma membrane transporter that functions in copper homeostasis in Arabidopsis and is a novel target of SQUAMOSA promoter- binding protein-like 7 J Biol Chem 287, 33252-33267. [本文引用: 10]
KampfenkelK, KushnirS, BabiychukE, InzéD, Van MontaguM ( 1995). Molecular characterization of a putative Arabidopsis thaliana copper transporter and its yeast homologue J Biol Chem 270, 28479-28486. [本文引用: 2]
KlaumannS, NickolausSD, FürstSH, StarckS, SchneiderS, Ekkehard NeuhausH, TrentmannO ( 2011). The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana New Phytol 192, 393-404. [本文引用: 3]
KomoriH, HiguchiY ( 2010). Structure and molecular evolution of multicopper blue proteins Biomol Concepts 1, 31-40. [本文引用: 1]
KuperJ, LlamasA, HechtHJ, MendelRR, SchwarzG ( 2004). Structure of the molybdopterin-bound Cnx1G domain links molybdenum and copper metabolism Nature 430, 803-806. [本文引用: 1]
LiHX, FanRC, LiLB, WeiB, LiGL, GuLQ, WangXP, ZhangXQ ( 2014). Identification and characterization of a novel copper transporter gene family TaCT1 in common wheat Plant Cell Environ 37, 1561-1573. [本文引用: 1]
LvQD, ZhongYJ, WangYG, WangZY, ZhangL, ShiJ, WuZC, LiuY, MaoCZ, YiKK, WuP ( 2014). SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice Plant Cell 26, 1586-1597. [本文引用: 1]
Martín-TrilloM, CubasP ( 2010). TCP genes: a family snapshot ten years later Trends Plant Sci 15, 31-39. [本文引用: 1]
MishraP, PanigrahiKC ( 2015). GIGANTEA—an emerging story Front Plant Sci 6, 8. [本文引用: 1]
MocklerTC, MichaelTP, PriestHD, ShenR, SullivanCM, GivanSA, McEnteeC, KaySA, ChoryJ ( 2007). The DIURNAL project: DIURNAL and circadian expression profiling, model-based pattern matching, and promoter analysis Cold Spring Harb Symp Quant Biol 72, 353-363. [本文引用: 1]
NagaeM, NakataM, TakahashiY ( 2008). Identification of negative cis-acting elements in response to copper in the chloroplastic iron superoxide dismutase gene of the moss Barbula unguiculata Plant Physiol 146, 1687-1696. [本文引用: 1]
PageMD, JanetteK, HamelPP, MerchantSS ( 2009). Two Chlamydomonas CTR copper transporters with a novel cys-met motif are localized to the plasma membrane and function in copper assimilation Plant Cell 21, 928-943. [本文引用: 1]
Pe?arrubiaL, Andrés-ColásN, MorenoJ, PuigS ( 2010). Regulation of copper transport in Arabidopsis thaliana: a biochemical oscillator? J Biol Inorg Chem 15, 29-36. [本文引用: 1]
Pe?arrubiaL, RomeroP, Carrió-SeguíA, Andrés-BorderíaA, MorenoJ, SanzA ( 2015). Temporal aspects of copper homeostasis and its crosstalk with hormones Front Plant Sci 6, 255. [本文引用: 2]
Perea-GarcíaA, Andrés-BorderíaA, Mayo de AndrésS, SanzA, DavisAM, DavisSJ, HuijserP, Pe?arrubiaL ( 2016a). Modulation of copper deficiency responses by diurnal and circadian rhythms in Arabidopsis thaliana J Exp Bot 67, 391-403. [本文引用: 2]
Perea-GarcíaA, Garcia-MolinaA, Andrés-ColásN, Vera-SireraF, Pérez-AmadorMA, PuigS, Pe?arrubiaL ( 2013). Arabidopsis copper transport protein COPT2 participates in the cross talk between iron deficiency responses and low-phosphate signaling Plant Physiol 162, 180-194. [本文引用: 5]
Perea-GarcíaA, SanzA, MorenoJ, Andrés-BorderíaA, De AndrésSM, DavisAM, HuijserP, DavisSJ, Pe?arrubiaL ( 2016b). Daily rhythmicity of high affinity copper transport Plant Signal Behav 11, e1140291. [本文引用: 4]
PugaMI, MateosI, CharukesiR, WangZY, Franco-orrillaJM, De LorenzoL, IrigoyenML, MasieroS, BustosR, RodríguezJ, LeyvaA, RubioV, SommerH, Paz-AresJ ( 2014). Spx1 is a phosphate-dependent inhibitor of PHOSPHATE STARVATION RESPONSE 1 in Arabidopsis Proc Natl Acad Sci USA 111, 14947-14952. [本文引用: 1]
RavenJA, EvansMCW, KorbRE ( 1999). The role of trace metals in photosynthetic electron transport in O2-evolving organisms Photosynth Res 60, 111-150. [本文引用: 1]
RodríguezFI, EschJJ, HallAE, BinderBM, SchallerGE, BleeckerAB ( 1999). A copper cofactor for the ethylene receptor ETR1 from Arabidopsis Science 283, 996-998. [本文引用: 3]
SancenónV, PuigS, Mateu-AndrésI, DorceyE, ThieleDJ, Pe?arrubiaL ( 2004). The Arabidopsis copper transporter COPT1 functions in root elongation and pollen development J Biol Chem 279, 15348-15355. [本文引用: 3]
SancenónV, PuigS, MiraH, ThieleDJ, Pe?arrubiaL ( 2003). Identi?cation of a copper transporter family in Arabidopsis thaliana Plant Mol Biol 51, 577-587. [本文引用: 9]
SeoPJ, MasP ( 2015). STRESSing the role of the plant circadian clock Trends Plant Sci 20, 230-237. [本文引用: 1]
SommerF, KropatJ, MalasarnD, GrossoehmeNE, ChenX, GiedrocDP, MerchantSS ( 2010). The CRR1 nutritIonal copper sensor in Chlamydomonas contains two distinct metal-responsive domains Plant Cell 22, 4098-4113. [本文引用: 1]
TiwariM, VenkatachalamP, PenarrubiaL, SahiSV ( 2017). COPT2, a plasma membrane located copper transporter, is involved in the uptake of Au in Arabidopsis Sci Rep 7, 11430. [本文引用: 2]
WangHL, DuHM, LiHY, HuangY, DingJZ, LiuC, WangN, LanH, ZhangSZ ( 2018). Identification and functional characterization of the ZmCOPT copper transporter family in maize PLoS One 13, e0199081. [本文引用: 1]
WangXH, LiHY, DuXB, HarrisJ, GuoZJ, SunHZ ( 2012). Activation of carboplatin and nedaplatin by the N-terminus of human copper transporter 1 (hCTR1) Chem Sci 3, 3206-3215. [本文引用: 1]
WatersBM, McInturfSA, SteinRJ ( 2012). Rosette iron deficiency transcript and microRNA profiling reveals links between copper and iron homeostasis in Arabidopsis thaliana J Exp Bot 63, 5903-5918. [本文引用: 1]
WintzH, FoxT, WuYY, FengV, ChenWQ, ChangHS, ZhuT, VulpeC ( 2003). Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis J Biol Chem 278, 47644-47653. [本文引用: 2]
WuXB, SinaniD, KimH, LeeJ ( 2009). Copper transport activity of yeast Ctr1 is down-regulated via its C terminus in response to excess copper J Biol Chem 284, 4112-4122. [本文引用: 1]
WuY, ZhangD, ChuJY, BoyleP, WangY, BrindleID, De LucaV, DesprésC ( 2012). The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid Cell Rep 1, 639-647. [本文引用: 1]
YamasakiH, HayashiM, FukazawaM, KobayashiY, ShikanaiT ( 2009). SQUAMOSA promoter binding protein-like7 is a central regulator for copper homeostasis in Arabidopsis Plant Cell 21, 347-361. [本文引用: 4]
YanSP, DongXN ( 2014). Perception of the plant immune signal salicylic acid Curr Opin Plant Biol 20, 64-68. [本文引用: 1]
YruelaI ( 2009). Copper in plants: acquisition, transport and interactions Funct Plant Biol 36, 409-430. [本文引用: 3]
YuPL, YuanJH, DengX, MaM, ZhangHY ( 2014). Subcellular targeting of bacterial CusF enhances Cu accumulation and alters root to shoot Cu translocation in Arabidopsis Plant Cell Physiol 55, 1568-1581. [本文引用: 1]
YuPL, YuanJH, ZhangH, DengX, MaM, ZhangHY ( 2016). Engineering metal-binding sites of bacterial CusF to enhance Zn/Cd accumulation and resistance by subcellular targeting J Hazard Mater 302, 275-285. [本文引用: 1]
YuZL, ZhangJG, WangXC, ChenJ ( 2008). Excessive copper induces the production of reactive oxygen species, which is mediated by phospholipase D, nicotinamide adenine dinucleotide phosphate oxidase and antioxidant systems J Integr Plant Biol 50, 157-167. [本文引用: 1]
YuanM, LiXH, XiaoJH, WangSP ( 2011). Molecular and functional analyses of COPT/Ctr-type copper transporter-like gene family in rice BMC Plant Biol 11, 69. [本文引用: 1]
ZhangHY, ZhaoX, LiJG, CaiHQ, DengXW, LiL ( 2014). MicroRNA408 is critical for the HY5-SPL7 gene network that mediates the coordinated response to light and copper Plant Cell 26, 4933-4953. [本文引用: 1]
酵母和植物中铜的转运系统及其调控 1 2007
... Cu是有机体正常发育不可缺少的微量元素之一, 在生理条件下, Cu以还原态Cu+和氧化态Cu2+两种形式存在(房茜等, 2007; 赵雪芹等, 2012).通过在两种状态之间相互转变, 使得Cu在多种植物蛋白中起氧化还原活性辅因子的作用, 影响质体蓝素蛋白、细胞色素c、Cu/Zn超氧化物歧化酶(Cu/Zn-SOD)、漆酶以及乙烯受体ETR1 (ethylene responsive 1)等的生物学活性, 参与植物体内光合作用、呼吸作用、活性氧的消除、细胞壁合成及激素信号转导等多种重要生物学过程(Raven et al., 1999; Gavnholt and Larsen, 2002; Gratão et al., 2005; Yruela, 2009; Komori and Higuchi, 2010; Yuan et al., 2010; 张红晓和张芬琴, 2011; Peñarrubia et al., 2015; 张美琪等, 2018). ...
Expression of the intracellular COPT3-mediated Cu transport is temporally regulated by the TCP16 transcription factor 9 2018
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... 的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
... 大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响.研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达.用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003).用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012).Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异.分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013).尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018). ...
... Cu缺乏诱导表达的基因启动子区大多包含GTAC基序, 该基序被称为Cu响应元件(Cu responsive element, CuRE).COPT1、COPT2和COPT6启动子区的600 bp范围内分别包含4、4和2个紧密相连的GTAC基序.COPT3的启动子区500 bp范围内共有3个GTAC基序.然而COPT5的启动子区缺少GTAC基序(Yamasaki et al., 2009; Jung et al., 2012; Andrés-Colás et al., 2018).GTAC基序是转录因子SPL7 (SQUAMOSA promoter-binding protein-like 7)的结合位点.SPL7是拟南芥SBP家族中的一员, 含有SBP结构域, 可以通过该结构域与靶基因启动子中的GTAC基序特异性结合, 进而调控靶基因的表达(Cardon et al., 1999; Yamasaki et al., 2009; Bernal et al., 2012).在Cu缺乏条件下, SPL7通过上述方式上调COPT1、COPT2和COPT6的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
... ; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
Deregulated copper transport affects Arabidopsis development especially in the absence of environmental cycles 5 2010
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
AtCOX17, an Arabidop- sis homolog of the yeast copper chaperone COX17 2 2002
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道.
拟南芥COPT的亚细胞定位与Cu的转运(<xref ref-type="bibr" rid="b53">改自Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质.在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中.推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中.HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质.HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中.HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应.HMA5定位于细胞膜, 介导Cu+的外排. ...
Subcellular localization of COPT and copper traffic in <i>Arabidopsis</i> (modified from <xref ref-type="bibr" rid="b53">Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+. ...
Circadian clock-controlled genes isolated from Neurospora crassa are late night-to early morning specific 1 1996
Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis 1 2012
... Cu缺乏诱导表达的基因启动子区大多包含GTAC基序, 该基序被称为Cu响应元件(Cu responsive element, CuRE).COPT1、COPT2和COPT6启动子区的600 bp范围内分别包含4、4和2个紧密相连的GTAC基序.COPT3的启动子区500 bp范围内共有3个GTAC基序.然而COPT5的启动子区缺少GTAC基序(Yamasaki et al., 2009; Jung et al., 2012; Andrés-Colás et al., 2018).GTAC基序是转录因子SPL7 (SQUAMOSA promoter-binding protein-like 7)的结合位点.SPL7是拟南芥SBP家族中的一员, 含有SBP结构域, 可以通过该结构域与靶基因启动子中的GTAC基序特异性结合, 进而调控靶基因的表达(Cardon et al., 1999; Yamasaki et al., 2009; Bernal et al., 2012).在Cu缺乏条件下, SPL7通过上述方式上调COPT1、COPT2和COPT6的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
Integrating mem- brane transport with male gametophyte development and function through transcriptomics 1 2006
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
A novel pineal night- specific ATPase encoded by the Wilson disease gene 1 1999
Interaction between ABA signaling and copper homeostasis in Arabidopsis thaliana 1 2016
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
The TRANSPLANTA collection of Arabidopsis lines: a resource for functional analysis of transcription factors based on their conditional overexpression 1 2014
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
Mitochondria and copper homeostasis in plants 2 2014
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道.
拟南芥COPT的亚细胞定位与Cu的转运(<xref ref-type="bibr" rid="b53">改自Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质.在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中.推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中.HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质.HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中.HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应.HMA5定位于细胞膜, 介导Cu+的外排. ...
Subcellular localization of COPT and copper traffic in <i>Arabidopsis</i> (modified from <xref ref-type="bibr" rid="b53">Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+. ...
The intracellular Arabidopsis COPT5 transport protein is required for photosynthetic electron transport under severe copper deficiency 5 2011
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
... 大多数离子转运蛋白的表达不仅受到转录水平的调控, 而且也受到蛋白水平的调控.例如磷转运蛋白, 某些磷转运蛋白从内质网的输出取决于其C末端丝氨酸残基的磷酸化状态, 该蛋白的磷酸化可以阻止其从内质网到细胞膜的运输(Lv et al., 2014; Puga et al., 2014).然而, 关于Cu水平变化对COPT蛋白丰度和稳定性影响的报道很少.目前比较清楚的是COPT5的蛋白水平不受外界Cu水平变化的影响(Sancenón et al., 2003; Garcia-Molina et al., 2011).但我们对COPT1的蛋白序列进行分析后发现, 其C末端的赖氨酸残基可能被泛素化.然而外界Cu水平变化是否能够真正影响COPT1蛋白水平, 及其是否能够被泛素化, 且其泛素化程度是否与外界Cu水平相关等需要进一步实验证明. ...
The Arabidopsis COPT6 transport protein functions in copper distribution under copper-deficient conditions 7 2013
... 拟南芥COPT1、COPT2和COPT6的序列相似性(可达75%-79%)最高, 均含有3个富含Met和His的基序(Jung et al., 2012).COPT1和COPT2的C末端具有CXC基序, 而COPT6则无.将这些基因分别在酵母突变体ctr1∆/ctr3∆ (因缺失高亲和Cu转运蛋白基因, CTR1和CTR3无法正常吸收Cu, 因而表现出生长缺陷)中表达, 它们能够完全互补酵母突变体ctr1∆/ctr3∆的缺陷表型(Sancenón et al., 2003; Garcia-Molina et al., 2013).与COPT1、COPT2和COPT6不同, COPT3的N端仅含有2个富含Met和His的基序, COPT5的N端仅含有1个, COPT4不仅N端缺乏这样的基序, 而且其TM2中也无MXXXM基序.在酵母ctr1∆/ctr3∆中表达后, COPT3和COPT5仅能部分互补突变体的表型; 而COPT4不能互补突变体的表型, 推测COPT4本身不具备转运Cu的能力, 可能通过与其它COPT蛋白相互作用而行使功能(Sancenón et al., 2003).由此可见, 拟南芥COPT家族各成员在Cu稳态平衡中可能具有不同的作用. ...
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
... 大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响.研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达.用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003).用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012).Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异.分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013).尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018). ...
COPT6 is a plasma membrane transporter that functions in copper homeostasis in Arabidopsis and is a novel target of SQUAMOSA promoter- binding protein-like 7 10 2012
... 通过序列比对及酵母Cu吸收缺陷突变体的功能互补实验, 已在多种植物中发现COPT家族成员.例如, 拟南芥(Arabidopsis thaliana)中共有6个成员, COPT1-6 (Sancenón et al., 2003; Jung et al., 2012); 水稻(Oryza sativa)中有7个, OsCOPT1-7 (Yuan et al., 2010, 2011); 小麦(Triticum aestivum)中有1个, TaCT1 (Li et al., 2014); 绿藻(Chlamydomonas reinhardtii)中发现了4个, 分别为CrCTR1、CrCTR2、CrCTR3和CrCOPT1 (Page et al., 2009); 玉米(Zea mays)中有3个, ZmCOPT1-3 (Wang et al., 2018).其中, 对模式植物拟南芥中的COPT研究最多, 其功能也最清楚. ...
... 拟南芥COPT1、COPT2和COPT6的序列相似性(可达75%-79%)最高, 均含有3个富含Met和His的基序(Jung et al., 2012).COPT1和COPT2的C末端具有CXC基序, 而COPT6则无.将这些基因分别在酵母突变体ctr1∆/ctr3∆ (因缺失高亲和Cu转运蛋白基因, CTR1和CTR3无法正常吸收Cu, 因而表现出生长缺陷)中表达, 它们能够完全互补酵母突变体ctr1∆/ctr3∆的缺陷表型(Sancenón et al., 2003; Garcia-Molina et al., 2013).与COPT1、COPT2和COPT6不同, COPT3的N端仅含有2个富含Met和His的基序, COPT5的N端仅含有1个, COPT4不仅N端缺乏这样的基序, 而且其TM2中也无MXXXM基序.在酵母ctr1∆/ctr3∆中表达后, COPT3和COPT5仅能部分互补突变体的表型; 而COPT4不能互补突变体的表型, 推测COPT4本身不具备转运Cu的能力, 可能通过与其它COPT蛋白相互作用而行使功能(Sancenón et al., 2003).由此可见, 拟南芥COPT家族各成员在Cu稳态平衡中可能具有不同的作用. ...
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
... 大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响.研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达.用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003).用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012).Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异.分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013).尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018). ...
... Cu缺乏诱导表达的基因启动子区大多包含GTAC基序, 该基序被称为Cu响应元件(Cu responsive element, CuRE).COPT1、COPT2和COPT6启动子区的600 bp范围内分别包含4、4和2个紧密相连的GTAC基序.COPT3的启动子区500 bp范围内共有3个GTAC基序.然而COPT5的启动子区缺少GTAC基序(Yamasaki et al., 2009; Jung et al., 2012; Andrés-Colás et al., 2018).GTAC基序是转录因子SPL7 (SQUAMOSA promoter-binding protein-like 7)的结合位点.SPL7是拟南芥SBP家族中的一员, 含有SBP结构域, 可以通过该结构域与靶基因启动子中的GTAC基序特异性结合, 进而调控靶基因的表达(Cardon et al., 1999; Yamasaki et al., 2009; Bernal et al., 2012).在Cu缺乏条件下, SPL7通过上述方式上调COPT1、COPT2和COPT6的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
... ; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
Molecular characterization of a putative Arabidopsis thaliana copper transporter and its yeast homologue 2 1995
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana 3 2011
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
Structure and molecular evolution of multicopper blue proteins 1 2010
... Cu是有机体正常发育不可缺少的微量元素之一, 在生理条件下, Cu以还原态Cu+和氧化态Cu2+两种形式存在(房茜等, 2007; 赵雪芹等, 2012).通过在两种状态之间相互转变, 使得Cu在多种植物蛋白中起氧化还原活性辅因子的作用, 影响质体蓝素蛋白、细胞色素c、Cu/Zn超氧化物歧化酶(Cu/Zn-SOD)、漆酶以及乙烯受体ETR1 (ethylene responsive 1)等的生物学活性, 参与植物体内光合作用、呼吸作用、活性氧的消除、细胞壁合成及激素信号转导等多种重要生物学过程(Raven et al., 1999; Gavnholt and Larsen, 2002; Gratão et al., 2005; Yruela, 2009; Komori and Higuchi, 2010; Yuan et al., 2010; 张红晓和张芬琴, 2011; Peñarrubia et al., 2015; 张美琪等, 2018). ...
Structure of the molybdopterin-bound Cnx1G domain links molybdenum and copper metabolism 1 2004
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
Identification and characterization of a novel copper transporter gene family TaCT1 in common wheat 1 2014
... 通过序列比对及酵母Cu吸收缺陷突变体的功能互补实验, 已在多种植物中发现COPT家族成员.例如, 拟南芥(Arabidopsis thaliana)中共有6个成员, COPT1-6 (Sancenón et al., 2003; Jung et al., 2012); 水稻(Oryza sativa)中有7个, OsCOPT1-7 (Yuan et al., 2010, 2011); 小麦(Triticum aestivum)中有1个, TaCT1 (Li et al., 2014); 绿藻(Chlamydomonas reinhardtii)中发现了4个, 分别为CrCTR1、CrCTR2、CrCTR3和CrCOPT1 (Page et al., 2009); 玉米(Zea mays)中有3个, ZmCOPT1-3 (Wang et al., 2018).其中, 对模式植物拟南芥中的COPT研究最多, 其功能也最清楚. ...
SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice 1 2014
... 大多数离子转运蛋白的表达不仅受到转录水平的调控, 而且也受到蛋白水平的调控.例如磷转运蛋白, 某些磷转运蛋白从内质网的输出取决于其C末端丝氨酸残基的磷酸化状态, 该蛋白的磷酸化可以阻止其从内质网到细胞膜的运输(Lv et al., 2014; Puga et al., 2014).然而, 关于Cu水平变化对COPT蛋白丰度和稳定性影响的报道很少.目前比较清楚的是COPT5的蛋白水平不受外界Cu水平变化的影响(Sancenón et al., 2003; Garcia-Molina et al., 2011).但我们对COPT1的蛋白序列进行分析后发现, 其C末端的赖氨酸残基可能被泛素化.然而外界Cu水平变化是否能够真正影响COPT1蛋白水平, 及其是否能够被泛素化, 且其泛素化程度是否与外界Cu水平相关等需要进一步实验证明. ...
TCP genes: a family snapshot ten years later 1 2010
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
GIGANTEA—an emerging story 1 2015
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
The DIURNAL project: DIURNAL and circadian expression profiling, model-based pattern matching, and promoter analysis 1 2007
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
Identification of negative cis-acting elements in response to copper in the chloroplastic iron superoxide dismutase gene of the moss Barbula unguiculata 1 2008
Two Chlamydomonas CTR copper transporters with a novel cys-met motif are localized to the plasma membrane and function in copper assimilation 1 2009
... 通过序列比对及酵母Cu吸收缺陷突变体的功能互补实验, 已在多种植物中发现COPT家族成员.例如, 拟南芥(Arabidopsis thaliana)中共有6个成员, COPT1-6 (Sancenón et al., 2003; Jung et al., 2012); 水稻(Oryza sativa)中有7个, OsCOPT1-7 (Yuan et al., 2010, 2011); 小麦(Triticum aestivum)中有1个, TaCT1 (Li et al., 2014); 绿藻(Chlamydomonas reinhardtii)中发现了4个, 分别为CrCTR1、CrCTR2、CrCTR3和CrCOPT1 (Page et al., 2009); 玉米(Zea mays)中有3个, ZmCOPT1-3 (Wang et al., 2018).其中, 对模式植物拟南芥中的COPT研究最多, 其功能也最清楚. ...
Regulation of copper transport in Arabidopsis thaliana: a biochemical oscillator? 1 2010
... COPT蛋白属于Cu转运蛋白Ctr (copper transporter)家族, 在哺乳动物和酵母(Saccharomyces cerevisiae)中也有其同源蛋白.生物化学、遗传学和拓扑结构学研究表明, 该家族蛋白包含3个保守的跨膜结构域(TMs).其中TM2中存在1个MXXXM基序(M代表甲硫氨酸(Met); X代表任意氨基酸), 在TM3中存在1个GXXXG基序(G代表甘氨酸(Gly); X代表任意氨基酸), 这些结构和Cu离子的结合与转运有关; 其N端具有富含Met的基序, 且游离在细胞外间隙, 能够与胞外Cu离子结合, 介导Cu离子跨膜运输; C端具有富含半胱氨酸(Cys)的基序(CXC基序), 存在于细胞质内, 能够与胞内Cu离子结合, 并将Cu离子传递给胞内的Cu伴侣蛋白, 在Cu过量的情况下能阻断Cu的转运(Wu et al., 2009; Peñarrubia et al., 2010; Wang et al., 2012; 袁金红等, 2016). ...
Temporal aspects of copper homeostasis and its crosstalk with hormones 2 2015
... Cu是有机体正常发育不可缺少的微量元素之一, 在生理条件下, Cu以还原态Cu+和氧化态Cu2+两种形式存在(房茜等, 2007; 赵雪芹等, 2012).通过在两种状态之间相互转变, 使得Cu在多种植物蛋白中起氧化还原活性辅因子的作用, 影响质体蓝素蛋白、细胞色素c、Cu/Zn超氧化物歧化酶(Cu/Zn-SOD)、漆酶以及乙烯受体ETR1 (ethylene responsive 1)等的生物学活性, 参与植物体内光合作用、呼吸作用、活性氧的消除、细胞壁合成及激素信号转导等多种重要生物学过程(Raven et al., 1999; Gavnholt and Larsen, 2002; Gratão et al., 2005; Yruela, 2009; Komori and Higuchi, 2010; Yuan et al., 2010; 张红晓和张芬琴, 2011; Peñarrubia et al., 2015; 张美琪等, 2018). ...
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
Modulation of copper deficiency responses by diurnal and circadian rhythms in Arabidopsis thaliana 2 2016
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
... 的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
Arabidopsis copper transport protein COPT2 participates in the cross talk between iron deficiency responses and low-phosphate signaling 5 2013
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道. ...
... 大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响.研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达.用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003).用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012).Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异.分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013).尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018). ...
... ; Perea-García et al., 2016b), COPT3在ZT12的表达(光期结束)高于ZT0点(光期开始) (Andrés-Colás et al., 2018). ...
... COPT1、COPT2和COPT6的启动子序列中存在多个光响应调控元件和昼夜节律相关的顺式调控元件, 如C-box (GACGTC)和Hybrid C/A-box (GACGTA), 二者均为光形态建成调控因子HY5 (elongated hypoctyl 5)结合元件.其中一些元件参与响应光敏色素(phytochrome)对基因表达的调控过程, 这与phy突变体中COPT2表达水平较低相符(Perea-García et al., 2016b).转录因子SPL7的表达也具有昼夜节律性, 然而在phyB突变体中这种表达特性消失(Mockler et al., 2007).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
... ).受phyB调控, 许多SPL7依赖的Cu缺乏响应基因的表达均具有昼夜节律性, 其振荡相位稍滞后于SPL7 (表达高峰出现在0小时), 大约每22小时达到1次表达高峰(Perea-García et al., 2016b).尽管COPT3的启动子中包含3个GTAC基序, 由于其表达高峰(12小时)与SPL7 (0小时)差距较大, 因此推测有其它转录因子参与COPT3的昼夜节律性表达调控.最近, Andrés-Colás等(2018)对条件过表达转录因子库(Conditional overexpression TF library)进行筛选获得TCP (teosinte branched 1, cycloidea and proliferating cell factor 1)蛋白家族成员TCP16.在Cu缺乏时, 编码该蛋白的基因主要在0小时表达.TCP16能够与COPT3启动子上的顺式作用调控元件(cis-acting regulatory element, CARE) (TTGAGCCCAT)结合, 进而下调COPT3的表达(Martín-Trillo and Cubas, 2010; Coego et al., 2014; Andrés-Colás et al., 2018).如上所述, 生物钟可以调控COPT基因的表达; 同样地, COPT也可以通过其转运底物Cu水平的改变而影响生物钟核心组分的表达, 胞内Cu水平影响生物钟核心基因CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)和GI (gigantea)的表达.例如, 外施Cu能够抑制CCA1和LHY的振幅和GI的转录活性.在一天中的不同时刻于培养基中添加Cu并对相位反应曲线进行观察, 发现Cu水平能够改变LHY的相位和振幅, 但不影响生物钟周期(Andrés-Colás et al., 2010; Mishra and Panigrahi, 2015; Seo and Mas, 2015; Perea-García et al., 2016a).此外还发现, LHY和GI的启动子中分别包含3个和2个GTAC基序, 暗示SPL7可能直接参与Cu缺乏时LHY和GI的上调表达调控(Perea-García et al., 2016a).研究表明, 转录因子SPL7与光形态建成调控因子HY5之间存在相互作用(Zhang et al., 2014).进一步研究表明, Cu稳态与生物钟之间存在紧密联系, 且HY5-SPL7基因网络在介导生物钟和Cu稳态的同步化及协调应答中发挥重要作用. ...
Spx1 is a phosphate-dependent inhibitor of PHOSPHATE STARVATION RESPONSE 1 in Arabidopsis 1 2014
... 大多数离子转运蛋白的表达不仅受到转录水平的调控, 而且也受到蛋白水平的调控.例如磷转运蛋白, 某些磷转运蛋白从内质网的输出取决于其C末端丝氨酸残基的磷酸化状态, 该蛋白的磷酸化可以阻止其从内质网到细胞膜的运输(Lv et al., 2014; Puga et al., 2014).然而, 关于Cu水平变化对COPT蛋白丰度和稳定性影响的报道很少.目前比较清楚的是COPT5的蛋白水平不受外界Cu水平变化的影响(Sancenón et al., 2003; Garcia-Molina et al., 2011).但我们对COPT1的蛋白序列进行分析后发现, 其C末端的赖氨酸残基可能被泛素化.然而外界Cu水平变化是否能够真正影响COPT1蛋白水平, 及其是否能够被泛素化, 且其泛素化程度是否与外界Cu水平相关等需要进一步实验证明. ...
The role of trace metals in photosynthetic electron transport in O2-evolving organisms 1 1999
... Cu是有机体正常发育不可缺少的微量元素之一, 在生理条件下, Cu以还原态Cu+和氧化态Cu2+两种形式存在(房茜等, 2007; 赵雪芹等, 2012).通过在两种状态之间相互转变, 使得Cu在多种植物蛋白中起氧化还原活性辅因子的作用, 影响质体蓝素蛋白、细胞色素c、Cu/Zn超氧化物歧化酶(Cu/Zn-SOD)、漆酶以及乙烯受体ETR1 (ethylene responsive 1)等的生物学活性, 参与植物体内光合作用、呼吸作用、活性氧的消除、细胞壁合成及激素信号转导等多种重要生物学过程(Raven et al., 1999; Gavnholt and Larsen, 2002; Gratão et al., 2005; Yruela, 2009; Komori and Higuchi, 2010; Yuan et al., 2010; 张红晓和张芬琴, 2011; Peñarrubia et al., 2015; 张美琪等, 2018). ...
A copper cofactor for the ethylene receptor ETR1 from Arabidopsis 3 1999
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道.
拟南芥COPT的亚细胞定位与Cu的转运(<xref ref-type="bibr" rid="b53">改自Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质.在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中.推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中.HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质.HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中.HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应.HMA5定位于细胞膜, 介导Cu+的外排. ...
Subcellular localization of COPT and copper traffic in <i>Arabidopsis</i> (modified from <xref ref-type="bibr" rid="b53">Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+. ...
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
The Arabidopsis copper transporter COPT1 functions in root elongation and pollen development 3 2004
... 拟南芥COPT1是植物中第1个被鉴定的COPT家族成员.将COPT1启动子与编码葡糖醛酸酶(GUS)的基因uidA融合表达, 发现COPT1基因在拟南芥主根和侧根根尖的表达最强, 在萌发种子的胚、子叶以及叶片的表皮毛、气孔保卫细胞也能检测到GUS信号; 在花粉发育早期不表达, 仅在花粉发育后期表达(Kampfenkel et al., 1995; Sancenón et al., 2004).COPT2组织表达模式与COPT1非常相似, 在拟南芥的子叶、表皮毛、花药以及成熟的花粉粒、根毛等部位均能检测到其表达; 与COPT1不同, COPT2在根尖的伸长区和分生区中不表达(Sancenón et al., 2003; Perea- García et al., 2013).COPT6主要在拟南芥茎和叶的维管组织中表达, 在子叶、分生组织、表皮毛、侧根和气孔中也能检测到其表达, 在初生根或次生根的根尖中不表达; 在生殖器官中, COPT6在柱头、子房、花丝、花粉以及胚胎中均有表达(Jung et al., 2012; Garcia-Molina et al., 2013).通过对COPT3启动子驱动GUS报告基因表达的转基因植株进行观察后发现, 在营养器官中, 仅在叶片的维管束中可检测到GUS活性(Andrés-Colás et al., 2018); 生殖器官中, 在花药尤其是花粉发育的早期高水平表达, 在花粉发育后期不表达(Bock et al., 2006), 在花柱伸长时的雄蕊花丝中也能检测到COPT3的表达(Andrés-Colás et al., 2018).将COPT5的起始密码子ATG上游357 bp的序列与GUS基因融合后于拟南芥中稳定表达, 对转基因植株中的GUS活性检测后发现, COPT5在主根的内皮层和维管束及根毛中强烈表达, 在地上部的表达相对较弱, 且GUS活性主要集中在胚轴、子叶和叶片的维管束中; 在生殖生长阶段, COPT5在胚珠和雌蕊中有较强表达, 在雄蕊中, 仅在花丝中有表达, 在花粉粒中几乎不表达(GarciaMolina et al., 2011).有关COPT4启动子的活性研究尚未见报道, 而RT-PCR结果显示, COPT4在根中表达很强, 且其表达水平远高于其它几个成员(Sancenón et al., 2003)(表1). ...
... Expression patterns of COPT family genes detected by use of promoter-GUS in Arabidopsis thaliana Table 1
基因名称
表达器官
参考文献
花
叶
茎
根
花粉
花丝
雌蕊
胚珠
子叶
表皮毛
保卫细胞
维管组织
维管组织
胚轴
主根
侧根
根毛
COPT1
++
-
/
/
+
+
+
-
-
+
+
+
/
Kampfenkel et al.,1995; Sancenon et al., 2004
COPT2
++
-
-
/
+
+
/
/
-
/
+
+
+
Sancenón et al., 2003; Perea-García et al., 2013
COPT3
++
+
/
/
/
/
/
+
/
/
/
/
/
Andrés-Colás et al., 2018
COPT5
-
+
++
++
+
+
/
++
+
+
++
+
++
Garcia-Molina et al., 2011
COPT6
+
+
+
+
+
+
+
++
++
/
-
+
/
Jung et al., 2012; Garcia-Molina et al., 2013
+: Expression; ++: Strong expression; -: No expression; /: Not reported ...
Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis 2 2003
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道.
拟南芥COPT的亚细胞定位与Cu的转运(<xref ref-type="bibr" rid="b53">改自Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质.在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中.推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中.HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质.HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中.HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应.HMA5定位于细胞膜, 介导Cu+的外排. ...
Subcellular localization of COPT and copper traffic in <i>Arabidopsis</i> (modified from <xref ref-type="bibr" rid="b53">Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+. ...
Copper transport activity of yeast Ctr1 is down-regulated via its C terminus in response to excess copper 1 2009
... COPT蛋白属于Cu转运蛋白Ctr (copper transporter)家族, 在哺乳动物和酵母(Saccharomyces cerevisiae)中也有其同源蛋白.生物化学、遗传学和拓扑结构学研究表明, 该家族蛋白包含3个保守的跨膜结构域(TMs).其中TM2中存在1个MXXXM基序(M代表甲硫氨酸(Met); X代表任意氨基酸), 在TM3中存在1个GXXXG基序(G代表甘氨酸(Gly); X代表任意氨基酸), 这些结构和Cu离子的结合与转运有关; 其N端具有富含Met的基序, 且游离在细胞外间隙, 能够与胞外Cu离子结合, 介导Cu离子跨膜运输; C端具有富含半胱氨酸(Cys)的基序(CXC基序), 存在于细胞质内, 能够与胞内Cu离子结合, 并将Cu离子传递给胞内的Cu伴侣蛋白, 在Cu过量的情况下能阻断Cu的转运(Wu et al., 2009; Peñarrubia et al., 2010; Wang et al., 2012; 袁金红等, 2016). ...
The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid 1 2012
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
SQUAMOSA promoter binding protein-like7 is a central regulator for copper homeostasis in Arabidopsis 4 2009
... 大多数COPTs基因在转录水平上的表达受到外界Cu水平的影响.研究发现, Cu过量时, COPT1、COPT2和COPT6均下调表达.用1 mmol∙L-1 Cu处理拟南芥幼苗, COPT1的表达显著下调, 而COPT2的表达被完全抑制(Sancenón et al., 2003).用45 μmol∙L-1 Cu处理时叶片中COPT6的表达显著下调(Jung et al., 2012).Cu缺乏时, COPT1、COPT2和COPT6均上调表达, 但其表达模式存在较大差异.分别用1和10 μmol∙L-1 Cu处理拟南芥时, COPT2的表达无明显变化, 而COPT6的表达在1 μmol∙L-1 Cu条件下比在10 μmol∙L-1 Cu条件下增加约2倍; 在100 μmol∙L-1 BCS (bathocuproine disulfonate, 一种Cu离子螯合剂)条件下, COPT2的表达量显著增加, 比1和10 μmol∙L-1 Cu条件下增加约11倍, 而在此条件下COPT6的表达变化较小, 仅分别增加约2和4倍(Yamasaki et al., 2009; Garcia-Molina et al., 2013; Perea-García et al., 2013).尽管较早的实验结果表明, Cu缺乏并不诱导COPT3和COPT5的表达(Sancenón et al., 2003), 然而最近有研究显示COPT3的表达受到Cu缺乏的强烈诱导(Andrés-Colás et al., 2018). ...
... Cu缺乏诱导表达的基因启动子区大多包含GTAC基序, 该基序被称为Cu响应元件(Cu responsive element, CuRE).COPT1、COPT2和COPT6启动子区的600 bp范围内分别包含4、4和2个紧密相连的GTAC基序.COPT3的启动子区500 bp范围内共有3个GTAC基序.然而COPT5的启动子区缺少GTAC基序(Yamasaki et al., 2009; Jung et al., 2012; Andrés-Colás et al., 2018).GTAC基序是转录因子SPL7 (SQUAMOSA promoter-binding protein-like 7)的结合位点.SPL7是拟南芥SBP家族中的一员, 含有SBP结构域, 可以通过该结构域与靶基因启动子中的GTAC基序特异性结合, 进而调控靶基因的表达(Cardon et al., 1999; Yamasaki et al., 2009; Bernal et al., 2012).在Cu缺乏条件下, SPL7通过上述方式上调COPT1、COPT2和COPT6的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
... ; Yamasaki et al., 2009; Bernal et al., 2012).在Cu缺乏条件下, SPL7通过上述方式上调COPT1、COPT2和COPT6的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
... 的表达(Yamasaki et al., 2009; Jung et al., 2012).在Cu充足条件下, SPL7通过结合Cu离子将Zn离子从锌指结构中置换出来, 使其无法结合GTAC基序(Sommer et al., 2010). ...
Perception of the plant immune signal salicylic acid 1 2014
... Cu是植物激素乙烯受体ETR1的辅助因子(Rodríguez et al., 1999).钼辅因子(molybdenum cofactor, MoCo)能够调控ABA和IAA的合成, 而Cu为MoCo合成所必需(Kuper et al., 2004).Cu也与多胺(polyamine, PA)的降解有关(Górecka et al., 2007).此外, Cu稳态在水杨酸(SA)和脱落酸(ABA)的信号转导中具有重要作用(Wu et al., 2012; Yan and Dong, 2014).然而截至目前, 关于植物激素如何调节Cu的吸收和转运还知之甚少.Peñarrubia (2015)对COPT家族成员的启动子序列进行分析后发现, ABA和赤霉素响应元件最多, 其次是乙烯和生长素响应元件, 茉莉酸响应相关元件最少.在Cu充足(1 μmol∙L-1)或过量(25 μmol∙L-1)时, copt2和copt1/copt2/copt6三突变体对ABA处理非常敏感.在这些突变体中, 诸如NCED3、WRKY40、HY5和ABI5的表达受到不同Cu水平调控, 表明在这些突变体中, Cu稳态失衡对ABA合成、转运和信号转导过程均产生影响.此外, ABA也调控SPL7的表达, 这可能是ABA抑制COPT1、COPT2和COPT6表达的原因(Carrió-Seguí et al., 2016).然而, ABA和Cu稳态之间的相互作用机制还需要进一步研究和证实. ...
Copper in plants: acquisition, transport and interactions 3 2009
... Cu是有机体正常发育不可缺少的微量元素之一, 在生理条件下, Cu以还原态Cu+和氧化态Cu2+两种形式存在(房茜等, 2007; 赵雪芹等, 2012).通过在两种状态之间相互转变, 使得Cu在多种植物蛋白中起氧化还原活性辅因子的作用, 影响质体蓝素蛋白、细胞色素c、Cu/Zn超氧化物歧化酶(Cu/Zn-SOD)、漆酶以及乙烯受体ETR1 (ethylene responsive 1)等的生物学活性, 参与植物体内光合作用、呼吸作用、活性氧的消除、细胞壁合成及激素信号转导等多种重要生物学过程(Raven et al., 1999; Gavnholt and Larsen, 2002; Gratão et al., 2005; Yruela, 2009; Komori and Higuchi, 2010; Yuan et al., 2010; 张红晓和张芬琴, 2011; Peñarrubia et al., 2015; 张美琪等, 2018). ...
... 将COPT的开放读码框与绿色荧光蛋白(GFP)基因融合, 在拟南芥叶肉细胞原生质体中瞬时表达后, 发现COPT1、COPT2和COPT6均主要定位于细胞膜(Andrés-Colás et al., 2010; Garcia-Molina et al., 2013; Perea-García et al., 2013).COPT5定位于液泡膜和前液泡体膜(Klaumann et al., 2011; Garcia- Molina et al., 2011).COPT3则定位于内膜系统, 进一步运用蔗糖密度梯度技术对表达COPT3-HA的拟南芥叶片的膜组分进行分离后发现, COPT3在组分中的分布模式与内质网(ER)的marker蛋白SEC12很相似, 推测COPT3可能定位于ER上(Andrés-Colás et al., 2018)(图1).关于COPT4的亚细胞定位尚未见报道.
拟南芥COPT的亚细胞定位与Cu的转运(<xref ref-type="bibr" rid="b53">改自Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+可以被ZIP蛋白吸收或被FRO还原为Cu+, Cu+通过COPT1、COPT2和COPT6蛋白转运进入细胞质.在细胞内, COPT5定位于液泡膜和液泡前体膜, 将液泡和液泡前体中的Cu+转运到细胞质基质中.推测COPT3定位于内质网膜, 并将内质网中的Cu+转运到细胞质基质中.HMA6和HMA1分别定位于叶绿体外膜和内膜, 分别负责将Cu+和Cu2+转运到叶绿体基质.HMA8定位于类囊体膜上, 可将叶绿体基质的Cu+转运到类囊体腔中.HMA7能够转运Cu+通过高尔基体膜, 参与乙烯反应.HMA5定位于细胞膜, 介导Cu+的外排. ...
Subcellular localization of COPT and copper traffic in <i>Arabidopsis</i> (modified from <xref ref-type="bibr" rid="b53">Rodríguez et al., 1999</xref>; <xref ref-type="bibr" rid="b16">Balandin and Castresana, 2002</xref>; <xref ref-type="bibr" rid="b62">Wintz et al., 2003</xref>; <xref ref-type="bibr" rid="b67">Yruela, 2009</xref>; <xref ref-type="bibr" rid="b26">Garcia et al., 2014</xref>) Cu2+can be uptaken by ZIP proteins or reduced to Cu+ by FRO and enters into the cytosol through the COPT1, COPT2 and COPT6 transporters. In the cell, COPT5 localizes in the tonoplast and vacuolar precursor membranes and transports Cu+ to the cytosol. It is speculated that COPT3 localizes to the ER membrane and transports Cu+ to the cytosol. HMA6 and HMA1 are localized in the membranes of the chloroplast and responsible for transporting Cu+ and Cu2+ to the cytosol, respectively. HMA8 is located in the thylakoid membrane and transports the Cu+ of the stroma into the thylakoid cavity. HMA7 transports Cu+ through the golgi membrane and participating in the ethylene reaction. HMA5 localizes in the cell membrane and mediates the efflux of Cu+. ...
Subcellular targeting of bacterial CusF enhances Cu accumulation and alters root to shoot Cu translocation in Arabidopsis 1 2014
Excessive copper induces the production of reactive oxygen species, which is mediated by phospholipase D, nicotinamide adenine dinucleotide phosphate oxidase and antioxidant systems 1 2008

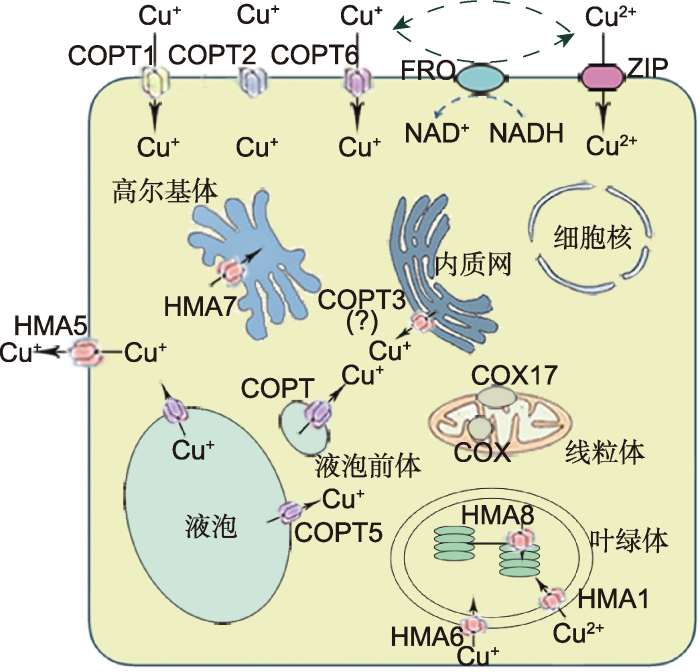 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}