杨艺
1 , 王娜
1 , 王奎玲
1 , 刘庆华
1 , 李伟
1 , 郭霄
1 , 马颖杰
2 , 刘庆超
, 1 , * 1.
青岛农业大学园林与林学院, 青岛 266109 2.
威海水木原景观设计有限公司, 威海 264205 Megasporogenesis, Microsporogenesis and Development of Male and Female Gametophytes of Clematis heracleifolia Yi Yang
1 , Na Wang
1 , Kuiling Wang
1 , Qinghua Liu
1 , Wei Li
1 , Xiao Guo
1 , Yingjie Ma
2 , Qingchao Liu
, 1 , * 1.
College of Landscape Architecture and Forestry, Qingdao Agricultural University, Qingdao 266109, China 2.
Weihai Shuimuyuan Design Co. Lid, Weihai 264205, China 通讯作者: *, E-mail: liuqingchao7205@126.com 责任编辑: 白羽红
收稿日期: 2018-12-5
接受日期: 2019-06-18
网络出版日期: 2019-09-01
基金资助: 山东省林业科技创新项目 (LYCX03-2018-15 )
Corresponding authors: *, E-mail: liuqingchao7205@126.com Editor: Bai Yu-hong Received: 2018-12-5
Accepted: 2019-06-18
Online: 2019-09-01
摘要 铁线莲属植物在花部形态和结构方面存在较大差异, 遗传背景相对复杂。因此, 在杂交育种前对其进行胚胎学研究具有重要意义。利用石蜡切片技术对大叶铁线莲(Clematis heracleifolia) 大小孢子发生及雌雄配子体发育过程进行研究, 结果显示, 大叶铁线莲具雄株和两性花植株。雄花中, 雄配子体发育偶见败育现象; 而两性花中多数花粉发育异常, 形成功能性雌花。正常发育的两性花中, 雄蕊较雌蕊先发育完全。花药4室, 具腺质绒毡层, 偶见变形绒毡层。胞质分裂为同时型, 以四面体型四分体为主, 偶见左右对称型。成熟花药中, 花药壁由纤维状加厚的表皮及药室内壁构成, 花粉粒为2-细胞型, 近球状, 散沟型。子房1室, 内含少量退化胚珠及1个发育正常的胚珠, 倒生, 单珠被, 薄珠心, 蓼型胚囊, 具线形大孢子四分体及双核反足细胞。大叶铁线莲可能处于相对进化的过渡地位。在杂交育种中, 建议以雄花植株作为父本, 两性花植株仅用作母本; 在两性花花芽大小为0.5-0.8 cm时进行去雄处理。 关键词: 大叶铁线莲 ;
铁线莲属 ;
配子体 ;
毛茛科 ;
孢子体 Abstract In Clematis (Ranunculaceae) species show significant differences in floral morphology and structures, consistent with their complex genetic background. Thus, it is important to study the embryological characteristics prior to any attempts of breeding by hybridization. Here, we report the characterization of microsporogenesis, microgametogenesis, megasporogenesis and macrogametogenesis of Clematis heracleifolia by floral dissection. We show that C. heracleifolia is androdioecy. Aborted microgametophyte cells are observed in the majority of bisexual flowers but occasionally found in male flowers, which results in the formation of functional female flowers. However, in a small number of normal bisexual flowers, male gametophytes mature faster than that of female gametophytes. During microsporogenesis, the tetrasporangiate anther is formed, followed by simultaneous microsporocyte cytokinesis, and the formation of glandular tapetum, occasionally with amoeboid tapetum. After meiosis, the microspores tetrads are mostly tetrahedral, and occasionally symmetrical. The wall of mature anther contains fibrous thickening epidermis and endothecium. Pollen grains are spheroidal, pantocolpate, and 2-celled. The ovary has one chamber with a normal and a few degraded ovules, which contains a anatropous, unitegmic, tenuinucellate, and Polygonum -type embryo-sac. A linear tetrad of megaspores and dikaryocyte antipodal cells can be observed. This species may belong to a transitional clade within Clematis . In breeding practice for C. heracleifolia , it is advised to use plants bearing male flowers and hermaphrodite flowers as male and female parents, respectivley, with the bisexual flowers of 0.5-0.8-cm-long to be emasculated. Keywords: Clematis heracleifolia ;
Clematis ;
gametophyte ;
Ranunculaceae ;
sporophyte PDF (4073KB) 摘要页面 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 杨艺, 王娜, 王奎玲, 刘庆华, 李伟, 郭霄, 马颖杰, 刘庆超 . 大叶铁线莲大小孢子发生及雌雄配子体发育. 植物学报, 2019,
54 (5): 596-605 doi:10.11983/CBB18261
Yang Yi, Wang Na, Wang Kuiling, Liu Qinghua, Li Wei, Guo Xiao, Ma Yingjie, Liu Qingchao .
Megasporogenesis, Microsporogenesis and Development of Male and Female Gametophytes of Clematis heracleifolia .
Chinese Bulletin of Botany , 2019,
54 (5): 596-605 doi:10.11983/CBB18261
毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种。其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值。目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 )。在杂交育种方面, 国外已培育出大量优良品种。我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏。植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础。由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义。
全雪丽等(2012) 利用石蜡切片技术对棉团铁线莲(
C. hexapetala )小孢子及雄配子体发育进行了研究。结果表明, 棉团铁线莲花药4室, 具分泌绒毡层, 成熟花粉粒近球状, 具3条萌发沟, 2-细胞型。
张敏涛等(2016) 观察了东方晨曲铁线莲(
Clematis ‘Ernest Markham’)小孢子及雄配子体发育, 发现其花药4室, 花药壁发育为双子叶型, 胞质分裂为同时型, 成熟花粉粒呈圆球状, 具3条萌发沟, 2-细胞型, 偶见花粉败育现象。
王自芬(2008) 利用扫描电镜观察了绣球藤(
C. montana )、钝齿铁线莲(
C. apiifolia var.
obtusidentata )、灌木铁线莲(
C. fruticosa )和大瓣铁线莲(
C. macropetala )胚珠外部形态。4种铁线莲属植物均为倒生胚珠, 单珠被, 珠柄腹侧具乳突状附属物。国内外有关铁线莲属植物的胚胎学研究相对较少, 且研究对象多为雄蕊, 缺乏对雌蕊发育、雌雄蕊间的发育关系、不同发育时期的雌雄蕊及花芽大小变化的详细研究。同时, 该属植物花芽相对较小, 分化进程较快(
杨艺等, 2019 ), 不利于杂交育种。因此, 有必要对铁线莲属植物的胚胎学特征作进一步研究。
大叶铁线莲(
C. heracleifolia )为毛茛科铁线莲属多年生草本, 花期4-8月, 开花繁茂, 观赏价值较高。其抗逆性强, 养护管理简便, 是一种不可多得的野生花卉资源。目前, 关于大叶铁线莲核型分析(
王娜等, 2017 )、种子萌发(
杨艺等, 2018 )及花芽分化(
杨艺等, 2019 )等已有报道。本研究以大叶铁线莲作为植物材料, 利用石蜡切片技术对其大、小孢子发生和雌、雄配子体发育过程进行观察, 同时对各发育时期的花芽、雄蕊及雌蕊长度进行测定, 以期为后续的杂交育种奠定基础, 同时也为其系统分类和栽培利用提供理论依据。
1 材料与方法 大叶铁线莲(
Clematis heracleifolia DC.)实验材料采于辽宁省丹东市草莓沟。2014年引种栽植于青岛农业大学园林与林学院试验地(120°12′E, 36°20′N)。该地区属暖温带大陆性季风气候, 年平均气温12.6°C, 年平均降水量700 mm。2017年和2018年4-8月采集不同发育阶段的花芽, 用FAA (甲醛:冰醋酸:50%乙醇=1:1:18, v/v/v)固定。采用常规石蜡切片法, 利用轮转式切片机(Leica RM-2145)切片。花药横切, 厚度为8 μm; 子房纵切, 厚度为10 μm。经苏木精-伊红染色后, 用Leica DM500显微镜拍照, 并用Image J和游标卡尺测定各发育时期的花芽大小及雌雄蕊长度。
大叶铁线莲花粉经叔丁醇冷冻干燥及喷金处理(JFC-1600)后, 利用扫描电镜(JSM-7500F, 2 kV)观察并拍照。随机选取30粒花粉, 利用Image J测定花粉粒微刺长度及密度, 观察萌发孔及其外壁纹饰。
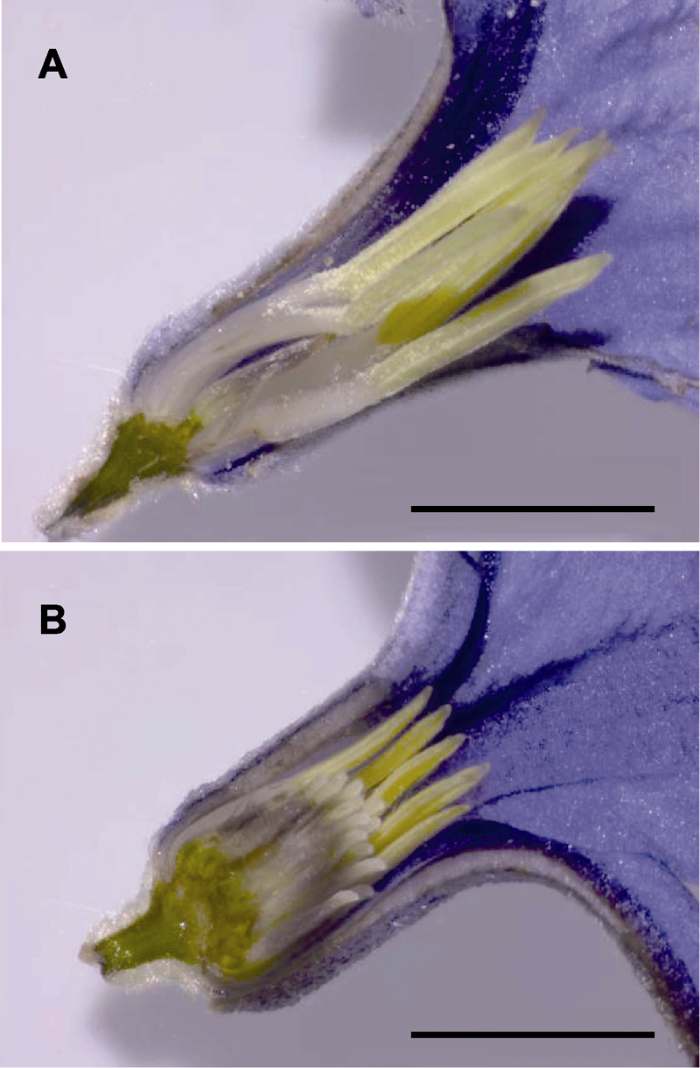
2 结果与讨论 大叶铁线莲聚伞花序顶生或腋生, 具雄株与两性花植株。我们分别对雄花(
图1A )植株的小孢子和雄配子体发育及两性花(
图1B )植株的大小孢子及其配子体发育进行了研究。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1大叶铁线莲花部结构 雄花; (B) 两性花。Bars=5 mm
Figure 1The floral structure of Clematis heracleifolia Staminate flower; (B) Bisexual flower. Bars=5 mm
2.1 雄花小孢子发生及雄配子体发育 图2 和
图3 显示, 大叶铁线莲花药4室, 花药壁发育类型为基本型, 由外向内依次为表皮(1层)、药室内壁(1层)、中层(1-2层)及绒毡层。以腺质绒毡层为主, 偶见分泌绒毡层。小孢子形成过程中, 胞质分裂类型为同时型, 以四面体型四分体为主, 偶见左右对称型。成熟花药中, 花药壁由表皮及药室内壁构成, 二者均存在纤维状加厚, 花粉粒为2-细胞型。
2.1.1 小孢子发生
4月中下旬, 大叶铁线莲花芽开始分化。最初, 雄蕊原基为表皮细胞下一团分裂旺盛的细胞。随后, 花药表皮细胞下分化出孢原细胞, 经平周分裂形成初生壁细胞及初生造孢细胞(
图2A )。初生壁细胞经多次有丝分裂依次形成药室内壁、中层及绒毡层, 初生造孢细胞经有丝分裂形成多个次生造孢细胞(
图2B , C1)。次生造孢细胞经有丝分裂发育形成小孢子母细胞(
图2C 1, D), 因其排列紧密、体积较大、形状不规则且具浓密细胞质及明显的细胞核而易于辨识。
随着周围胼胝质的形成, 小孢子母细胞逐渐由不规则形状变为圆形, 随后进入减数第1次分裂(
图2 E1-E3)。在此期间无细胞壁形成, 只存在双核细胞而非二分体。在减数第1次分裂早期, 绒毡层细胞的细胞核经多次分裂形成多核细胞(
图2 E1-E3)。小孢子母细胞完成减数第2次分裂后(
图2 F1, F2), 小孢子四分体形成, 彼此间以胼胝质相互隔离(
图2 F2)。大叶铁线莲小孢子四分体以四面体型为主, 偶见左右对称型(
图2 F2)。在此期间, 中层及绒毡层细胞逐渐退化或仅留残迹(
图2 E1-G)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2大叶铁线莲雄花小孢子发育 (A) 孢原细胞(箭头所示, 上)及其平周分裂形成初生壁细胞(箭头所示, 上)和初生造孢细胞(箭头所示, 下); (B) 花药壁; (C1) 次生造孢细胞; (C2) 有丝分裂前期(箭头所示); (C3) 有丝分裂中期(箭头所示, 右)和后期(箭头所示, 左); (C4) 有丝分裂末期(箭头所示); (D) 小孢子母细胞; (E1) 减数第1次分裂前期; (E2) 减数第1次分裂中期(箭头所示); (E3) 减数第1次分裂后期(箭头所示, 右)和末期(箭头所示, 左); (F1) 减数第2次分裂后期(箭头所示, 左), 偶见变形绒毡层(箭头所示, 右); (F2) 减数第2次分裂末期, 偶见左右对称型四分体(箭头所示); (G) 四分体解体; (H) 花药发育异步现象(箭头所示); (I1)-(I4) 小孢子败育。Bars=50 μm
Figure 2Microspore development in staminat e flower of Clematis heracleifolia (A) Archesporial cell (arrow, up), primary parietal cell (arrow, up) and primary sporogenous cell (arrow, down); (B) Anther wall; (C1) A row of secondary sporogenous cells; (C2) Prophase (arrow); (C3) Metaphase (arrow, right) and anaphase (arrow, left); (C4) Telophase (arrow); (D) Microspore mother cells; (E1) Microspore mother cells at prophase of prophase I of meiosis; (E2) Microspore mother cells at metaphase of meiosis I (arrow); (E3) Microspore mother cells at anaphase (arrow, right) and telophase (arrow, left) of meiosis I; (F1) Microspore mother cells at anaphase of meiosis II (arrow, left) and amoeboid tapetum (arrow, right); (F2) Microspore mother cells at telophase of meiosis II (arrow) and symmetrical microspore tetrads; (G) Degrading tetrahedral tetrad; (H) The asynchronous phases; (I1)-(I4) Aborted microspores. Bars=50 μm
大叶铁线莲雄花的花药横切结果显示, 小孢子四分体形成期间, 花药各药室间存在发育异步现象,如同一花药的不同药室中, 小孢子可能处于不同发育时期, 同一药室内小孢子发育也存在差异(
图2H )。在大叶铁线莲雄花的花药发育过程中, 偶见小孢子败育现象, 具体表现为细胞核降解、细胞质浓缩及细胞皱缩(
图2I 1-I4)。
2.1.2 雄配子体发育
随着胼胝质的解体, 小孢子四分体彼此分离(
图2G ), 随后小孢子发育进入单核居中期, 此时具加厚细胞壁、浓密细胞质、液泡以及明显中央核(
图3A )。随着液泡增大, 小孢子体积逐渐增大, 细胞核被挤向细胞质一侧(
图3B ), 进入不均等有丝分裂阶段(
图3C 1-C3),最后形成1个大的营养细胞和1个小的生殖细胞, 彼此间由新细胞壁分隔(
图3C 4)。在此期间, 大叶铁线莲花药绒毡层细胞完全解体或仅存残迹(
图3C 1-C4)。随着花粉粒的发育, 2个细胞间的细胞壁消失, 生殖细胞游离于营养细胞的细胞质中, 花药的表皮及药室内壁细胞出现不同程度的纤维状加厚, 随后花药开裂、散粉(
图3D )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3大叶铁线莲雄花雄配子体发育 (A) 单核居中期; (B) 单核靠边期; (C1) 有丝分裂前期(箭头所示); (C2) 有丝分裂中期(箭头所示, 下)和后期(箭头所示, 上); (C3) 有丝分裂后期(箭头所示); (C4) 有丝分裂末期, 形成营养细胞和生殖细胞(箭头所示); (D) 花药开裂, 散粉, 2-细胞花粉粒。(A), (B), (C1), (D) Bars=50 μm; (C2)-(C4) Bars=20 μm
Figure 3Microgametophyte development in staminate flower of Clematis heracleifolia (A) Microspore at monokaryotic stage; (B) Microspore at monokaryotic side stage; (C1) Microspore at prophase (arrow); (C2) Microspore at metaphase (arrow, down) and anaphase (arrow, up); (C3) Microspore at anaphase (arrow); (C4) Microspore at telophase, vegetative cell and germ cell are formed (arrow); (D) The dehiscent anther, and the 2-celled mature pollen grains. (A), (B), (C1), (D) Bars=50 μm; (C2)-(C4) Bars=20 μm
2.2 两性花小孢子发生及雄配子体发育 花芽横切结果显示, 随着小孢子母细胞形成, 多数两性花的花药发育异常, 构成花药壁的表皮、药室内壁和中层细胞的细胞核及细胞质多数降解, 绒毡层细胞及小孢子母细胞发育异常, 降解或仅余残迹(
图 4A -D)。对于少量能够正常形成花粉的两性花(约5%), 小孢子及雄配子体的发育过程与雄花相似, 最终形成2-细胞型花粉粒(
图4G , H), 其花粉形成过程中也偶见败育现象(
图4E , F)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4大叶铁线莲两性花小孢子发生及花粉发育 (A)-(D) 小孢子母细胞败育; (E), (F) 部分小孢子母细胞败育; (G), (H) 花粉正常发育。an: 花药; ov: 子房。(A), (C), (G) Bars=500 μm; (B), (F) Bars=50 μm; (D), (E), (H) Bars=200 μm
Figure 4Microspore and pollen grain development in bisexual flower of Clematis heracleifolia (A)-(D) Aborted microspore mother cells; (E), (F) Some aborted microspore mother cells; (G), (H) Normal pollen grains. an: Anther; ov: Ovary. (A), (C), (G) Bars=500 μm; (B), (F) Bars=50 μm; (D), (E), (H) Bars=200 μm
2.3 花粉形态观察 大叶铁线莲花粉为单粒花粉粒(
图5A ), 近球状, 散沟型(
表1 )。萌发沟向内凹陷, 均匀分布于花粉粒表面(
图5A , B), 花粉粒表面具明显穿孔及微刺(
图5C , D), 微刺平均高度为0.46 μm (
表1 )。
Table 1 表1 表1 大叶铁线莲花粉特性
Table 1
Pollen characteristics of Clematis heracleifolia Characters State Shape Spheroidal Type of aperture Pantocolpate Diameter (μm) 16.09±0.88 Spinule height (μm) 0.46±0.09 Number of spinules (3×3 μm2 ) 3.80±1.08
新窗口打开 |
下载CSV 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5大叶铁线莲花粉扫描电镜观察 (A) Bar=10 μm; (B)-(D) Bars=1 μm
Figure 5Scanning electron microscopy views of pollen grains of Clematis heracleifolia (A) Bar=10 μm; (B)-(D) Bars=1 μm
2.4 两性花大孢子发生及雌配子体发育 大叶铁线莲子房1室, 具少量退化胚珠及1个发育正常胚珠, 倒生, 单珠被, 薄珠心, 蓼型胚囊(
图6A , B)。
4月下旬, 胚珠原基出现, 伴随细胞分裂生长出现一个手指状结构, 并发育成珠心组织。随后, 在胚珠原基的珠心表皮细胞下出现1个孢原细胞(
图6C ), 体积明显大于周边珠心细胞, 并在之后直接发育成大孢子母细胞, 具浓密细胞质及明显细胞核(
图6D )。在此之后, 胚珠弯曲并逐渐与珠柄平行呈倒生状态。与此同时, 大孢子母细胞经减数分裂形成线形四分体(
图6E )。然后, 珠孔端的3个大孢子退化, 合点端的1个大孢子直接发育成功能大孢子(
图6F )。随着其体积增大, 功能大孢子通过有丝分裂相继形成二核胚囊(
图6G )、四核胚囊(
图6H )及八核胚囊(
图6I )。与此同时, 单珠被逐渐形成, 珠孔端具一狭缝(
图6E -I)。
八核胚囊仅存在一小段时间, 然后形成细胞壁, 胚囊发育成熟。大叶铁线莲成熟胚囊中具7细胞8核, 包括珠孔端的2个助细胞和1个卵细胞、1个二倍体的中央细胞以及合点端的3个反足细胞(
图6J -L)。2个助细胞呈对称排列, 具大液泡(
图6J )。卵细胞较助细胞小, 根据其所处位置易于辨认(
图6K , L)。二倍体的中央细胞液泡化明显(
图6K , L)。3个反足细胞体积较大, 具2核(
图6J -L)。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6大叶铁线莲两性花大孢子及雌配子体发育 (A), (B) 子房和胚珠; (C) 孢原细胞(箭头所示); (D) 大孢子母细胞(箭头所示); (E) 线形大孢子四分体(箭头所示); (F) 珠孔端3个大孢子退化, 合点端发育形成功能大孢子(箭头所示); (G) 二核胚囊; (H) 四核胚囊; (I) 八核胚囊; (J)-(L) 成熟胚囊7细胞8核。ac: 反足细胞; cc: 中央极核; ec: 卵细胞; sy: 助细胞。(A), (B) Bars=200 μm; (C)-(L) Bars=50 μm
Figure 6Megaspore and megagametophyte development in bisexual flower of Clematis heracleifolia (A), (B) The ovary and ovule; (C) Archesporial cells (arrow); (D) Megaspore mother cells (arrow); (E) A linear tetrad of megaspores (arrows); (F) The three endostome megaspores were degenerated and the chalazal one was a functional megaspore (arrows); (G) Binucleate embryo sac; (H) Tetranucleate embryo sac; (I) Embryo sac with 8 nuclei; (J)-(L) Mature embryo sac of seven-cell with eight nuclei. ac: Antipodal cell; cc: Central nucleus; ec: Egg cell; sy: Synergids. (A), (B) Bars=200 μm; (C)-(L) Bars=50 μm
2.5 配子体发育与花芽外部形态变化 大叶铁线莲雌雄蕊均为向心式发育。雄花及发育正常的两性花中, 雌雄配子体发育与花芽外部形态变化的关系如
表2 和
表3 所示。
Table 2 表2 表2 大叶铁线莲雄花中配子体发育与花芽形态的关系
Table 2
Relationship between gametophytes development and flower bud morphology in staminate flower of Clematis heracleifolia Bud length (cm) Stamen length (cm) Male gametophyte 0.22-0.40 0.15-0.31 Microsporocyte, meiosis 0.40-0.85 0.31-0.51 Meiosis, microspore 0.85-1.09 0.51-0.86 Microspore, monokaryotic side stage, mitosis 1.09-1.39 0.86-1.12 Mitosis, 2-celled pollen 1.39-2.14 1.12-1.32 2-celled pollen
新窗口打开 |
下载CSV Table 3 表3 表3 大叶铁线莲两性花中配子体发育与花芽形态的关系
Table 3
Relationship between gametophytes development and flower bud morphology in bisexual flower of Clematis heracleifolia Bud length (cm) Stamen length (cm) Pistil length (cm) Male gametophyte Female gametophyte 0.16-0.36 0.10-0.14 0.05-0.11 Microsporocyte, meiosis, microspore - 0.36-0.50 0.14-0.34 0.10-0.18 Microspore monokaryotic side stage, mitosis Archesporium, megasporocyte 0.50-0.88 0.34-0.48 0.18-0.21 Mitosis, 2-celled pollen Megasporocyte, uninucleate embryo sac 0.88-0.95 0.48-0.52 0.21-0.30 2-celled pollen Uninucleate embryo sac, 2-nucleate or 4-nucleate embryo sac 0.95-1.42 0.52-0.60 0.30-0.42 2-celled pollen 4-nucleate or 8-nucleate embryo sac, mature embryo sac
新窗口打开 |
下载CSV 对于正常发育的两性花, 胚珠原基出现时, 雄配子体处于减数第2次分裂至小孢子时期, 雌配子体发育明显晚于雄配子体。
2.6 讨论 2.6.1 大叶铁线莲雄花及两性花雄蕊发育特点
大叶铁线莲有2种类型性别个体, 即雄花植株与两性花植株。2种植株的花芽内, 雄蕊早期发育过程十分相似。但在小孢子母细胞进入第1次减数分裂时期, 雄花植株的花药囊正常发育并最终形成花粉; 两性花植株的花药囊多数发育异常, 花药壁细胞的细胞质稀少、细胞核降解、绒毡层细胞已降解或仅余残迹, 最终形成花药空壳导致雄性不育, 两性花成为功能性雌花(
图4A -D), 这与臭椿(
Ailanthus altissima ) (
王永周等, 2008 )、茄属(
Solanum ) (
Anderson and Symon, 1989 )等植物的两性花植株相似。但与之不同的是, 上述植物的两性花植株中雄蕊完全败育, 而大叶铁线莲仍存在少量能够正常发育并形成成熟花粉的两性花。因此, 大叶铁线莲可能存在3类花芽, 即单性花(雄花), 发育异常的两性花(功能性雌花)及少量正常发育的两性花。从花芽形态轮廓看, 大叶铁线莲两性花短粗, 而雄花相对细长; 两性花雄蕊明显高于雌蕊, 且较雄花短小瘦弱(
图1 ;
表2 ,
表3 )。正常发育的两性花内, 雄配子体发育完全的时间显著早于雌配子体(
表2 ), 这可能有利于减少大叶铁线莲自花授粉概率。
由于未对两性花植株的花芽进行分株采集及保存, 因此本研究发现的少量正常发育的两性花属于偶然现象还是正常发育的两性花和功能上的雌花存在异株现象有待进一步研究。此外, 在杂交育种方面, 可参考雌雄配子体不同发育时期的花芽形态变化(
表2 ,
表3 )确定雌雄配子体发育时期, 进行去雄处理。
2.6.2 大叶铁线莲小孢子发生及雄配子体发育
研究表明, 大叶铁线莲小孢子及雄配子体发育过程与棉团铁线莲(
全雪丽等, 2012 )和东方晨曲铁线莲(
张敏涛等, 2016 )相似。但棉团铁线莲无分泌绒毡层, 其成熟花粉粒近球状, 萌发沟为三沟型(
全雪丽等, 2012 ); 东方晨曲铁线莲花药壁发育为双子叶型, 成熟花粉粒为圆球状, 三沟型, 且在其成熟花药壁中, 仅药室内壁存在纤维状加厚, 表皮细胞未见明显加厚(
张敏涛等, 2016 )。
大叶铁线莲雄花及正常发育的两性花存在少量花粉败育现象, 具体表现为小孢子细胞核降解, 细胞质浓缩, 细胞皱缩(
图3K 1, K2)。
王玖瑞等(2006) 认为, 四分体时期绒毡层异常加厚是导致枣(
Ziziphus jujuba )的小孢子聚集、解体及最终败育的主要原因。
王述彬等(2004) 研究表明, 辣椒(
Capsicum annuum )绒毡层细胞径向异常膨大从而挤压小孢子二分体是导致其裂解及最终不育的主要原因。
张克中等(2003) 发现, 绒毡层细胞前期液泡化、细胞质稀少及非正常膨大是导致王百合(
Lilium regale )小孢子败育的原因之一。通过对大叶铁线莲小孢子发育过程进行观察(
图2I 1-I4,
图4F ), 结果表明绒毡层细胞液泡化、细胞质稀少、非正常膨大、解体时间提前或推迟可能是导致小孢子败育的原因。
大叶铁线莲雄配子体发育过程中存在花药发育异步现象, 多发生于小孢子母细胞减数分裂时期(
图3I , J)。不同药室内多相差1-2个时期, 同一药室内基本同步, 有时相差1-2个时期(
图3 )。散粉阶段, 2-细胞花粉和单核花粉均存在(
图3 )。这与臭椿(
王永周等, 2008 )、东方晨曲铁线莲(
张敏涛等, 2016 )和耐冬山茶(
Camellia japonica (Naidong)) (
Zhang et al., 2017 )相似, 可能与阻碍小孢子间信息交流的胼胝质有关, 是增加花粉活性、延长授粉期及保持种群稳定的一种适应性表现(
赵桦等, 2011 )。
2.6.3 大叶铁线莲大孢子发生及雌配子体发育
研究表明, 大叶铁线莲为单珠被, 胚珠倒生(
图6 ), 这与绣球藤、钝齿铁线莲、灌木铁线莲和大瓣铁线莲(
王自芬, 2008 )相似。由于纵切花芽不能体现胚珠外部形态, 因此大叶铁线莲珠柄腹侧是否也具有乳突状附属物有待进一步观察。
据报道, 毛茛科植物的胚珠中孢原细胞数目不稳定(
宋萍, 2007 ), 如驴蹄草属(
Caltha )、升麻属(
Cimicifuga )、翠雀属(
Delphinium )、毛茛属(
Ranunculus )及唐松草属(
Thalictrum )植物的孢原细胞数量为1-3个不等, 而
Ranunculus septentrionalis 为2-13个不等。本研究表明, 大叶铁线莲仅有1个孢原细胞, 这与类叶升麻属(
Actaea )、铁筷子属(
Helleborus )、白头翁属(
Pulsatilla )及银莲花属(
Anemone )植物相似(
宋萍, 2007 )。研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 )。而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞。此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 )。本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L)。
2.6.4 系统学意义
据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 )。本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群。在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比)。由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群。
3 结论 从外部形态和结构上, 大叶铁线莲存在雄花植株和两性花植株, 具2种花芽。本研究表明, 大叶铁线莲雄花植株雄蕊基本发育正常, 仅少量败育; 两性花植株中, 雄蕊多数败育, 极少数(约5%)花芽的雌雄蕊发育正常。因此, 从配子体育性角度上看, 大叶铁线莲存在3种花芽, 即雄花、发育异常的两性花(功能性雌花)及少量正常发育的两性花。造成两性花中多数雄蕊败育的原因可能是绒毡层细胞液泡化、细胞质稀少、非正常膨大、解体时间提前或推迟。
基于本研究结果, 在杂交育种中, 建议两性花植株仅用作母本, 而雄性花植株用作父本。若为研究铁线莲不同种间或属间的杂交亲和性, 建议在花芽大小为0.5-0.8 cm时进行去雄处理, 此时花芽内雌雄蕊大小差异较明显, 易于操作。若仅以改良种质为目的, 可不对大叶铁线莲两性花花芽进行去雄操作。
[1] 管开云 , 李志坚 , 李景秀 , 匡建 (2002 ). 铁线莲属植物的引种栽培研究初报植物分类与资源学报 24, 392 -396 . [本文引用: 1] [2] 李杨 , 张伟 , 赫雪锋 , 汤海峰 (2018 ). 铁线莲属植物的化学成分及药理作用研究进展中南药学 16, 355 -362 . [本文引用: 1] [3] 刘萍 (2006 ). 铁筷子(毛茛科) 的形态、结构、发育及系统位置的研究硕士论文. 西安: 陕西师范大学 . pp. 15 -35 . [本文引用: 1] [4] 刘庆超 , 王奎玲 , 卢婉佩 , 刘庆华 (2014 ). 我国铁线莲属植物资源研究进展植物遗传资源学报 15, 483 -490, 497 . [本文引用: 1] [5] 全雪丽 , 刘继生 , 吴松权 (2012 ). 棉团铁线莲小孢子发生和雄配子体的形成辽宁林业科技 (1 ), 9 -10 , 32 . [本文引用: 3] [6] 宋春慧 (2006 ). 黄三七属(毛茛科) 植物的形态、结构、发育及系统位置研究硕士论文. 西安: 陕西师范大学. pp . 25 . [本文引用: 1] [7] 宋萍 (2007 ). 金莲花属和驴蹄草属(毛茛科)的比较形态学研究——基于形态、结构、花发育及胚胎学的证据硕士论文. 西安: 陕西师范大学 . pp. 11 -18 . [本文引用: 4] [8] 孙诚 , 谢磊 , 李良千 (2007 ). 铁线莲属尾叶铁线莲组(毛茛科)基于形态学证据的分支系统学植物学通报 24, 87 -98 . [本文引用: 1] [9] 王玖瑞 , 刘玲 , 刘孟军 , 周俊义 (2006 ). 枣树雄性不育新种质的获得园艺学报 33, 374 -377 . [本文引用: 1] [10] 王娜 , 王奎玲 , 刘庆华 , 刘庆超 (2017 ). 七种野生铁线莲属植物核型分析草业学报 26(11 ), 123 -130 . [本文引用: 1] [11] 王述彬 , 罗向东 , 戴亮芳 , 钱春桃 , 陈劲枫 (2004 ). 辣(甜)椒细胞质雄性不育系减数分裂和雄配子发生过程园艺学报 31, 807 -810 . [本文引用: 1] [12] 王文采 , 李良千 (2005 ). 铁线莲属一新分类系统植物分类学报 43, 431 -488 . [本文引用: 2] [13] 王永周 , 古松 , 江莎 , 任艳萍 , 许珂 , 李清河 (2008 ). 臭椿雄蕊的发育和雄配子体的形成园艺学报 35, 577 -586 . [本文引用: 2] [14] 王自芬 (2008 ). 毛茛科植物胚珠形态、结构、发育及其系统学意义博士论文. 西安: 陕西师范大学. pp . 25 . [本文引用: 4] [15] 杨艺 , 王娜 , 马颖杰 , 王奎玲 , 刘庆华 , 刘庆超 (2018 ). 4种铁线莲属植物种子萌发特性草业科学 35, 2395 -2402 . [本文引用: 1] [16] 杨艺 , 王娜 , 王奎玲 , 刘庆华 , 刘庆超 (2019 ). 铁线莲属4种植物的花芽分化研究园艺学报 46, 87 -95 . [本文引用: 2] [17] 张阿娟 (2006 ). 类叶升麻(毛茛科) 形态结构、发育及系统位置的研究硕士论文. 西安: 陕西师范大学 . pp. 14 -27 . [本文引用: 2] [18] 张克中 , 赵祥云 , 梁励 , 黄善武 , 张启翔 (2003 ). 王百合雄性不育突变体小孢子败育的细胞学研究北京林业大学学报 25(3 ), 124 -126 . [本文引用: 1] [19] 张敏涛 , 张荻 , 申晓辉 (2016 ). ‘东方晨曲’铁线莲小孢子发生和雄配子体发育进程的解剖学研究西北植物学报 36, 2213 -2219 . [本文引用: 4] [20] 张燕 , 黎斌 , 李思锋 (2010 ). 铁线莲属植物分类学及园艺学研究进展中国野生植物资源 29(5 ), 6 -10 . [本文引用: 1] [21] 赵桦 , 孟华菊 , 徐娟 (2011 ). 七叶树小孢子发生及雄配子体发育研究西北植物学报 31, 217 -222 . [本文引用: 1] [22] Bhandari NN 1968 ). Studies in the family Ranunculaceae. X. Embryology of Anemone LPhytomorphology 487 -497 . [本文引用: 2] [23] Xie L Li LQ 2012 ). Variation of pollen morphology, and its implications in the phylogeny of Clematis(Ranunculaceae)Plant Syst Evol 1437 -1453 . [本文引用: 1] [24] Zhang Q Hao Q Guo X Liu QC Sun YK Liu QH Wang KL 2017 ). Anther and ovule development in Camellia japonica(Naidong) in relation to winter dormancy: climatic evolution considerationsFlora 127 -139 . [本文引用: 1] 铁线莲属植物的引种栽培研究初报
1
2002
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
铁线莲属植物的化学成分及药理作用研究进展
1
2018
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
铁筷子(毛茛科) 的形态、结构、发育及系统位置的研究
1
2006
... 据报道, 毛茛科植物的胚珠中孢原细胞数目不稳定(
宋萍, 2007 ), 如驴蹄草属(
Caltha )、升麻属(
Cimicifuga )、翠雀属(
Delphinium )、毛茛属(
Ranunculus )及唐松草属(
Thalictrum )植物的孢原细胞数量为1-3个不等, 而
Ranunculus septentrionalis 为2-13个不等.本研究表明, 大叶铁线莲仅有1个孢原细胞, 这与类叶升麻属(
Actaea )、铁筷子属(
Helleborus )、白头翁属(
Pulsatilla )及银莲花属(
Anemone )植物相似(
宋萍, 2007 ).研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
我国铁线莲属植物资源研究进展
1
2014
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
棉团铁线莲小孢子发生和雄配子体的形成
3
2012
...
全雪丽等(2012) 利用石蜡切片技术对棉团铁线莲(
C. hexapetala )小孢子及雄配子体发育进行了研究.结果表明, 棉团铁线莲花药4室, 具分泌绒毡层, 成熟花粉粒近球状, 具3条萌发沟, 2-细胞型.
张敏涛等(2016) 观察了东方晨曲铁线莲(
Clematis ‘Ernest Markham’)小孢子及雄配子体发育, 发现其花药4室, 花药壁发育为双子叶型, 胞质分裂为同时型, 成熟花粉粒呈圆球状, 具3条萌发沟, 2-细胞型, 偶见花粉败育现象.
王自芬(2008) 利用扫描电镜观察了绣球藤(
C. montana )、钝齿铁线莲(
C. apiifolia var.
obtusidentata )、灌木铁线莲(
C. fruticosa )和大瓣铁线莲(
C. macropetala )胚珠外部形态.4种铁线莲属植物均为倒生胚珠, 单珠被, 珠柄腹侧具乳突状附属物.国内外有关铁线莲属植物的胚胎学研究相对较少, 且研究对象多为雄蕊, 缺乏对雌蕊发育、雌雄蕊间的发育关系、不同发育时期的雌雄蕊及花芽大小变化的详细研究.同时, 该属植物花芽相对较小, 分化进程较快(
杨艺等, 2019 ), 不利于杂交育种.因此, 有必要对铁线莲属植物的胚胎学特征作进一步研究. ...
... 研究表明, 大叶铁线莲小孢子及雄配子体发育过程与棉团铁线莲(
全雪丽等, 2012 )和东方晨曲铁线莲(
张敏涛等, 2016 )相似.但棉团铁线莲无分泌绒毡层, 其成熟花粉粒近球状, 萌发沟为三沟型(
全雪丽等, 2012 ); 东方晨曲铁线莲花药壁发育为双子叶型, 成熟花粉粒为圆球状, 三沟型, 且在其成熟花药壁中, 仅药室内壁存在纤维状加厚, 表皮细胞未见明显加厚(
张敏涛等, 2016 ). ...
... )相似.但棉团铁线莲无分泌绒毡层, 其成熟花粉粒近球状, 萌发沟为三沟型(
全雪丽等, 2012 ); 东方晨曲铁线莲花药壁发育为双子叶型, 成熟花粉粒为圆球状, 三沟型, 且在其成熟花药壁中, 仅药室内壁存在纤维状加厚, 表皮细胞未见明显加厚(
张敏涛等, 2016 ). ...
黄三七属(毛茛科) 植物的形态、结构、发育及系统位置研究
1
2006
... 据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
金莲花属和驴蹄草属(毛茛科)的比较形态学研究——基于形态、结构、花发育及胚胎学的证据
4
2007
... 据报道, 毛茛科植物的胚珠中孢原细胞数目不稳定(
宋萍, 2007 ), 如驴蹄草属(
Caltha )、升麻属(
Cimicifuga )、翠雀属(
Delphinium )、毛茛属(
Ranunculus )及唐松草属(
Thalictrum )植物的孢原细胞数量为1-3个不等, 而
Ranunculus septentrionalis 为2-13个不等.本研究表明, 大叶铁线莲仅有1个孢原细胞, 这与类叶升麻属(
Actaea )、铁筷子属(
Helleborus )、白头翁属(
Pulsatilla )及银莲花属(
Anemone )植物相似(
宋萍, 2007 ).研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
... )植物相似(
宋萍, 2007 ).研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
... ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
... s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
铁线莲属尾叶铁线莲组(毛茛科)基于形态学证据的分支系统学
1
2007
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
枣树雄性不育新种质的获得
1
2006
... 大叶铁线莲雄花及正常发育的两性花存在少量花粉败育现象, 具体表现为小孢子细胞核降解, 细胞质浓缩, 细胞皱缩(
图3K 1, K2).
王玖瑞等(2006) 认为, 四分体时期绒毡层异常加厚是导致枣(
Ziziphus jujuba )的小孢子聚集、解体及最终败育的主要原因.
王述彬等(2004) 研究表明, 辣椒(
Capsicum annuum )绒毡层细胞径向异常膨大从而挤压小孢子二分体是导致其裂解及最终不育的主要原因.
张克中等(2003) 发现, 绒毡层细胞前期液泡化、细胞质稀少及非正常膨大是导致王百合(
Lilium regale )小孢子败育的原因之一.通过对大叶铁线莲小孢子发育过程进行观察(
图2I 1-I4,
图4F ), 结果表明绒毡层细胞液泡化、细胞质稀少、非正常膨大、解体时间提前或推迟可能是导致小孢子败育的原因. ...
七种野生铁线莲属植物核型分析
1
2017
... 大叶铁线莲(
C. heracleifolia )为毛茛科铁线莲属多年生草本, 花期4-8月, 开花繁茂, 观赏价值较高.其抗逆性强, 养护管理简便, 是一种不可多得的野生花卉资源.目前, 关于大叶铁线莲核型分析(
王娜等, 2017 )、种子萌发(
杨艺等, 2018 )及花芽分化(
杨艺等, 2019 )等已有报道.本研究以大叶铁线莲作为植物材料, 利用石蜡切片技术对其大、小孢子发生和雌、雄配子体发育过程进行观察, 同时对各发育时期的花芽、雄蕊及雌蕊长度进行测定, 以期为后续的杂交育种奠定基础, 同时也为其系统分类和栽培利用提供理论依据. ...
辣(甜)椒细胞质雄性不育系减数分裂和雄配子发生过程
1
2004
... 大叶铁线莲雄花及正常发育的两性花存在少量花粉败育现象, 具体表现为小孢子细胞核降解, 细胞质浓缩, 细胞皱缩(
图3K 1, K2).
王玖瑞等(2006) 认为, 四分体时期绒毡层异常加厚是导致枣(
Ziziphus jujuba )的小孢子聚集、解体及最终败育的主要原因.
王述彬等(2004) 研究表明, 辣椒(
Capsicum annuum )绒毡层细胞径向异常膨大从而挤压小孢子二分体是导致其裂解及最终不育的主要原因.
张克中等(2003) 发现, 绒毡层细胞前期液泡化、细胞质稀少及非正常膨大是导致王百合(
Lilium regale )小孢子败育的原因之一.通过对大叶铁线莲小孢子发育过程进行观察(
图2I 1-I4,
图4F ), 结果表明绒毡层细胞液泡化、细胞质稀少、非正常膨大、解体时间提前或推迟可能是导致小孢子败育的原因. ...
铁线莲属一新分类系统
2
2005
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
... 据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
臭椿雄蕊的发育和雄配子体的形成
2
2008
... 大叶铁线莲有2种类型性别个体, 即雄花植株与两性花植株.2种植株的花芽内, 雄蕊早期发育过程十分相似.但在小孢子母细胞进入第1次减数分裂时期, 雄花植株的花药囊正常发育并最终形成花粉; 两性花植株的花药囊多数发育异常, 花药壁细胞的细胞质稀少、细胞核降解、绒毡层细胞已降解或仅余残迹, 最终形成花药空壳导致雄性不育, 两性花成为功能性雌花(
图4A -D), 这与臭椿(
Ailanthus altissima ) (
王永周等, 2008 )、茄属(
Solanum ) (
Anderson and Symon, 1989 )等植物的两性花植株相似.但与之不同的是, 上述植物的两性花植株中雄蕊完全败育, 而大叶铁线莲仍存在少量能够正常发育并形成成熟花粉的两性花.因此, 大叶铁线莲可能存在3类花芽, 即单性花(雄花), 发育异常的两性花(功能性雌花)及少量正常发育的两性花.从花芽形态轮廓看, 大叶铁线莲两性花短粗, 而雄花相对细长; 两性花雄蕊明显高于雌蕊, 且较雄花短小瘦弱(
图1 ;
表2 ,
表3 ).正常发育的两性花内, 雄配子体发育完全的时间显著早于雌配子体(
表2 ), 这可能有利于减少大叶铁线莲自花授粉概率. ...
... 大叶铁线莲雄配子体发育过程中存在花药发育异步现象, 多发生于小孢子母细胞减数分裂时期(
图3I , J).不同药室内多相差1-2个时期, 同一药室内基本同步, 有时相差1-2个时期(
图3 ).散粉阶段, 2-细胞花粉和单核花粉均存在(
图3 ).这与臭椿(
王永周等, 2008 )、东方晨曲铁线莲(
张敏涛等, 2016 )和耐冬山茶(
Camellia japonica (Naidong)) (
Zhang et al., 2017 )相似, 可能与阻碍小孢子间信息交流的胼胝质有关, 是增加花粉活性、延长授粉期及保持种群稳定的一种适应性表现(
赵桦等, 2011 ). ...
毛茛科植物胚珠形态、结构、发育及其系统学意义
4
2008
...
全雪丽等(2012) 利用石蜡切片技术对棉团铁线莲(
C. hexapetala )小孢子及雄配子体发育进行了研究.结果表明, 棉团铁线莲花药4室, 具分泌绒毡层, 成熟花粉粒近球状, 具3条萌发沟, 2-细胞型.
张敏涛等(2016) 观察了东方晨曲铁线莲(
Clematis ‘Ernest Markham’)小孢子及雄配子体发育, 发现其花药4室, 花药壁发育为双子叶型, 胞质分裂为同时型, 成熟花粉粒呈圆球状, 具3条萌发沟, 2-细胞型, 偶见花粉败育现象.
王自芬(2008) 利用扫描电镜观察了绣球藤(
C. montana )、钝齿铁线莲(
C. apiifolia var.
obtusidentata )、灌木铁线莲(
C. fruticosa )和大瓣铁线莲(
C. macropetala )胚珠外部形态.4种铁线莲属植物均为倒生胚珠, 单珠被, 珠柄腹侧具乳突状附属物.国内外有关铁线莲属植物的胚胎学研究相对较少, 且研究对象多为雄蕊, 缺乏对雌蕊发育、雌雄蕊间的发育关系、不同发育时期的雌雄蕊及花芽大小变化的详细研究.同时, 该属植物花芽相对较小, 分化进程较快(
杨艺等, 2019 ), 不利于杂交育种.因此, 有必要对铁线莲属植物的胚胎学特征作进一步研究. ...
... 研究表明, 大叶铁线莲为单珠被, 胚珠倒生(
图6 ), 这与绣球藤、钝齿铁线莲、灌木铁线莲和大瓣铁线莲(
王自芬, 2008 )相似.由于纵切花芽不能体现胚珠外部形态, 因此大叶铁线莲珠柄腹侧是否也具有乳突状附属物有待进一步观察. ...
... 据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
... ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
4种铁线莲属植物种子萌发特性
1
2018
... 大叶铁线莲(
C. heracleifolia )为毛茛科铁线莲属多年生草本, 花期4-8月, 开花繁茂, 观赏价值较高.其抗逆性强, 养护管理简便, 是一种不可多得的野生花卉资源.目前, 关于大叶铁线莲核型分析(
王娜等, 2017 )、种子萌发(
杨艺等, 2018 )及花芽分化(
杨艺等, 2019 )等已有报道.本研究以大叶铁线莲作为植物材料, 利用石蜡切片技术对其大、小孢子发生和雌、雄配子体发育过程进行观察, 同时对各发育时期的花芽、雄蕊及雌蕊长度进行测定, 以期为后续的杂交育种奠定基础, 同时也为其系统分类和栽培利用提供理论依据. ...
铁线莲属4种植物的花芽分化研究
2
2019
...
全雪丽等(2012) 利用石蜡切片技术对棉团铁线莲(
C. hexapetala )小孢子及雄配子体发育进行了研究.结果表明, 棉团铁线莲花药4室, 具分泌绒毡层, 成熟花粉粒近球状, 具3条萌发沟, 2-细胞型.
张敏涛等(2016) 观察了东方晨曲铁线莲(
Clematis ‘Ernest Markham’)小孢子及雄配子体发育, 发现其花药4室, 花药壁发育为双子叶型, 胞质分裂为同时型, 成熟花粉粒呈圆球状, 具3条萌发沟, 2-细胞型, 偶见花粉败育现象.
王自芬(2008) 利用扫描电镜观察了绣球藤(
C. montana )、钝齿铁线莲(
C. apiifolia var.
obtusidentata )、灌木铁线莲(
C. fruticosa )和大瓣铁线莲(
C. macropetala )胚珠外部形态.4种铁线莲属植物均为倒生胚珠, 单珠被, 珠柄腹侧具乳突状附属物.国内外有关铁线莲属植物的胚胎学研究相对较少, 且研究对象多为雄蕊, 缺乏对雌蕊发育、雌雄蕊间的发育关系、不同发育时期的雌雄蕊及花芽大小变化的详细研究.同时, 该属植物花芽相对较小, 分化进程较快(
杨艺等, 2019 ), 不利于杂交育种.因此, 有必要对铁线莲属植物的胚胎学特征作进一步研究. ...
... 大叶铁线莲(
C. heracleifolia )为毛茛科铁线莲属多年生草本, 花期4-8月, 开花繁茂, 观赏价值较高.其抗逆性强, 养护管理简便, 是一种不可多得的野生花卉资源.目前, 关于大叶铁线莲核型分析(
王娜等, 2017 )、种子萌发(
杨艺等, 2018 )及花芽分化(
杨艺等, 2019 )等已有报道.本研究以大叶铁线莲作为植物材料, 利用石蜡切片技术对其大、小孢子发生和雌、雄配子体发育过程进行观察, 同时对各发育时期的花芽、雄蕊及雌蕊长度进行测定, 以期为后续的杂交育种奠定基础, 同时也为其系统分类和栽培利用提供理论依据. ...
类叶升麻(毛茛科) 形态结构、发育及系统位置的研究
2
2006
... 据报道, 毛茛科植物的胚珠中孢原细胞数目不稳定(
宋萍, 2007 ), 如驴蹄草属(
Caltha )、升麻属(
Cimicifuga )、翠雀属(
Delphinium )、毛茛属(
Ranunculus )及唐松草属(
Thalictrum )植物的孢原细胞数量为1-3个不等, 而
Ranunculus septentrionalis 为2-13个不等.本研究表明, 大叶铁线莲仅有1个孢原细胞, 这与类叶升麻属(
Actaea )、铁筷子属(
Helleborus )、白头翁属(
Pulsatilla )及银莲花属(
Anemone )植物相似(
宋萍, 2007 ).研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
... 据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
王百合雄性不育突变体小孢子败育的细胞学研究
1
2003
... 大叶铁线莲雄花及正常发育的两性花存在少量花粉败育现象, 具体表现为小孢子细胞核降解, 细胞质浓缩, 细胞皱缩(
图3K 1, K2).
王玖瑞等(2006) 认为, 四分体时期绒毡层异常加厚是导致枣(
Ziziphus jujuba )的小孢子聚集、解体及最终败育的主要原因.
王述彬等(2004) 研究表明, 辣椒(
Capsicum annuum )绒毡层细胞径向异常膨大从而挤压小孢子二分体是导致其裂解及最终不育的主要原因.
张克中等(2003) 发现, 绒毡层细胞前期液泡化、细胞质稀少及非正常膨大是导致王百合(
Lilium regale )小孢子败育的原因之一.通过对大叶铁线莲小孢子发育过程进行观察(
图2I 1-I4,
图4F ), 结果表明绒毡层细胞液泡化、细胞质稀少、非正常膨大、解体时间提前或推迟可能是导致小孢子败育的原因. ...
‘东方晨曲’铁线莲小孢子发生和雄配子体发育进程的解剖学研究
4
2016
...
全雪丽等(2012) 利用石蜡切片技术对棉团铁线莲(
C. hexapetala )小孢子及雄配子体发育进行了研究.结果表明, 棉团铁线莲花药4室, 具分泌绒毡层, 成熟花粉粒近球状, 具3条萌发沟, 2-细胞型.
张敏涛等(2016) 观察了东方晨曲铁线莲(
Clematis ‘Ernest Markham’)小孢子及雄配子体发育, 发现其花药4室, 花药壁发育为双子叶型, 胞质分裂为同时型, 成熟花粉粒呈圆球状, 具3条萌发沟, 2-细胞型, 偶见花粉败育现象.
王自芬(2008) 利用扫描电镜观察了绣球藤(
C. montana )、钝齿铁线莲(
C. apiifolia var.
obtusidentata )、灌木铁线莲(
C. fruticosa )和大瓣铁线莲(
C. macropetala )胚珠外部形态.4种铁线莲属植物均为倒生胚珠, 单珠被, 珠柄腹侧具乳突状附属物.国内外有关铁线莲属植物的胚胎学研究相对较少, 且研究对象多为雄蕊, 缺乏对雌蕊发育、雌雄蕊间的发育关系、不同发育时期的雌雄蕊及花芽大小变化的详细研究.同时, 该属植物花芽相对较小, 分化进程较快(
杨艺等, 2019 ), 不利于杂交育种.因此, 有必要对铁线莲属植物的胚胎学特征作进一步研究. ...
... 研究表明, 大叶铁线莲小孢子及雄配子体发育过程与棉团铁线莲(
全雪丽等, 2012 )和东方晨曲铁线莲(
张敏涛等, 2016 )相似.但棉团铁线莲无分泌绒毡层, 其成熟花粉粒近球状, 萌发沟为三沟型(
全雪丽等, 2012 ); 东方晨曲铁线莲花药壁发育为双子叶型, 成熟花粉粒为圆球状, 三沟型, 且在其成熟花药壁中, 仅药室内壁存在纤维状加厚, 表皮细胞未见明显加厚(
张敏涛等, 2016 ). ...
... ); 东方晨曲铁线莲花药壁发育为双子叶型, 成熟花粉粒为圆球状, 三沟型, 且在其成熟花药壁中, 仅药室内壁存在纤维状加厚, 表皮细胞未见明显加厚(
张敏涛等, 2016 ). ...
... 大叶铁线莲雄配子体发育过程中存在花药发育异步现象, 多发生于小孢子母细胞减数分裂时期(
图3I , J).不同药室内多相差1-2个时期, 同一药室内基本同步, 有时相差1-2个时期(
图3 ).散粉阶段, 2-细胞花粉和单核花粉均存在(
图3 ).这与臭椿(
王永周等, 2008 )、东方晨曲铁线莲(
张敏涛等, 2016 )和耐冬山茶(
Camellia japonica (Naidong)) (
Zhang et al., 2017 )相似, 可能与阻碍小孢子间信息交流的胼胝质有关, 是增加花粉活性、延长授粉期及保持种群稳定的一种适应性表现(
赵桦等, 2011 ). ...
铁线莲属植物分类学及园艺学研究进展
1
2010
... 毛茛科(Ranunculaceae)铁线莲属(
Clematis )植物共355种, 我国约有140多种.其形态多样, 分布广泛, 花量大, 花期长, 具较高观赏价值及一定药用价值.目前, 国内外****已在铁线莲属植物种质资源、系统分类、栽培繁殖及药用价值等方面进行了大量研究(
管开云等, 2002 ;
王文采和李良千, 2005 ;
孙诚等,2007 ;
张燕等, 2010 ;
刘庆超等, 2014 ;
李杨等, 2018 ).在杂交育种方面, 国外已培育出大量优良品种.我国铁线莲属植物资源虽占全球的40%左右, 但多数处于野生状态, 资源利用率较低, 育种研究极度匮乏.植物胚胎学研究常用于揭示植物间的演变及进化水平, 为阐明植物个体发育及物种间的亲缘关系提供依据, 是开展杂交育种研究的基础.由于铁线莲属植物在花部形态、花的结构及生活型等方面存在较大差异, 遗传背景相对复杂, 因此, 在杂交育种前探明其胚胎学特征具有重要意义. ...
七叶树小孢子发生及雄配子体发育研究
1
2011
... 大叶铁线莲雄配子体发育过程中存在花药发育异步现象, 多发生于小孢子母细胞减数分裂时期(
图3I , J).不同药室内多相差1-2个时期, 同一药室内基本同步, 有时相差1-2个时期(
图3 ).散粉阶段, 2-细胞花粉和单核花粉均存在(
图3 ).这与臭椿(
王永周等, 2008 )、东方晨曲铁线莲(
张敏涛等, 2016 )和耐冬山茶(
Camellia japonica (Naidong)) (
Zhang et al., 2017 )相似, 可能与阻碍小孢子间信息交流的胼胝质有关, 是增加花粉活性、延长授粉期及保持种群稳定的一种适应性表现(
赵桦等, 2011 ). ...
Studies in the family Ranunculaceae. X. Embryology of
Anemone L
2
1968
... 大叶铁线莲有2种类型性别个体, 即雄花植株与两性花植株.2种植株的花芽内, 雄蕊早期发育过程十分相似.但在小孢子母细胞进入第1次减数分裂时期, 雄花植株的花药囊正常发育并最终形成花粉; 两性花植株的花药囊多数发育异常, 花药壁细胞的细胞质稀少、细胞核降解、绒毡层细胞已降解或仅余残迹, 最终形成花药空壳导致雄性不育, 两性花成为功能性雌花(
图4A -D), 这与臭椿(
Ailanthus altissima ) (
王永周等, 2008 )、茄属(
Solanum ) (
Anderson and Symon, 1989 )等植物的两性花植株相似.但与之不同的是, 上述植物的两性花植株中雄蕊完全败育, 而大叶铁线莲仍存在少量能够正常发育并形成成熟花粉的两性花.因此, 大叶铁线莲可能存在3类花芽, 即单性花(雄花), 发育异常的两性花(功能性雌花)及少量正常发育的两性花.从花芽形态轮廓看, 大叶铁线莲两性花短粗, 而雄花相对细长; 两性花雄蕊明显高于雌蕊, 且较雄花短小瘦弱(
图1 ;
表2 ,
表3 ).正常发育的两性花内, 雄配子体发育完全的时间显著早于雌配子体(
表2 ), 这可能有利于减少大叶铁线莲自花授粉概率. ...
... 据报道, 毛茛科植物的胚珠中孢原细胞数目不稳定(
宋萍, 2007 ), 如驴蹄草属(
Caltha )、升麻属(
Cimicifuga )、翠雀属(
Delphinium )、毛茛属(
Ranunculus )及唐松草属(
Thalictrum )植物的孢原细胞数量为1-3个不等, 而
Ranunculus septentrionalis 为2-13个不等.本研究表明, 大叶铁线莲仅有1个孢原细胞, 这与类叶升麻属(
Actaea )、铁筷子属(
Helleborus )、白头翁属(
Pulsatilla )及银莲花属(
Anemone )植物相似(
宋萍, 2007 ).研究显示, 毛茛科植物胚珠中, 孢原细胞先进行1次有丝分裂再形成大孢子母细胞, 如川陕金莲花(
Trollius buddae ) (
宋萍, 2007 ).而本研究显示, 大叶铁线莲与驴蹄草(
C. palustri s) (
宋萍, 2007 )及铁筷子(
H. thibetanus ) (
刘萍, 2006 )相似, 胚珠内孢原细胞不进行有丝分裂, 而是直接分化形成大孢子母细胞.此外, 毛茛科多数植物的反足细胞较大, 具二核或多核(
Bhandari, 1968 ), 少数为单核, 如类叶升麻(
A. asatiiaca ) (
张阿娟, 2006 ).本研究中, 大叶铁线莲的反足细胞较大, 具二核(
图6J -L). ...
Variation of pollen morphology, and its implications in the phylogeny of Clematis(Ranunculaceae)
1
2012
... 据报道, 毛茛科植物中雄蕊向心式发育较雄蕊离心式发育原始(
宋春慧, 2006 ), 三沟型萌发沟较散沟型及散孔型萌发沟原始(
Xie and Li, 2012 ), 倒生胚珠较横生胚珠原始(
王自芬, 2008 ), 双珠被较单珠被原始(
王自芬, 2008 ), 单核反足细胞较双核或多核反足细胞原始(
张阿娟, 2006 ).本研究表明, 大叶铁线莲既存在相对进化的特征, 如散沟型萌发沟, 单珠被, 大且双核的反足细胞, 又存在相对原始的特征, 如倒生胚珠, 雄蕊向心式发育, 因此其可能属于铁线莲属植物中相对进化的过渡类群.在已建立的铁线莲属植物分类体系(
王文采和李良千, 2005 )中, 大叶铁线莲位于相对进化的欧洲铁线莲演化干(与原始的绣球藤演化干相比).由于铁线莲属植物的胚胎学信息较少, 本研究虽不能确定大叶铁线莲的系统位置, 但却能与已有分类体系相互印证大叶铁线莲为相对进化的类群. ...
Anther and ovule development in Camellia japonica(Naidong) in relation to winter dormancy: climatic evolution considerations
1
2017
... 大叶铁线莲雄配子体发育过程中存在花药发育异步现象, 多发生于小孢子母细胞减数分裂时期(
图3I , J).不同药室内多相差1-2个时期, 同一药室内基本同步, 有时相差1-2个时期(
图3 ).散粉阶段, 2-细胞花粉和单核花粉均存在(
图3 ).这与臭椿(
王永周等, 2008 )、东方晨曲铁线莲(
张敏涛等, 2016 )和耐冬山茶(
Camellia japonica (Naidong)) (
Zhang et al., 2017 )相似, 可能与阻碍小孢子间信息交流的胼胝质有关, 是增加花粉活性、延长授粉期及保持种群稳定的一种适应性表现(
赵桦等, 2011 ). ...
备案号:
京ICP备16067583号-21 版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由
北京玛格泰克科技发展有限公司 设计开发
 ,1,*1.
,1,*1. 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT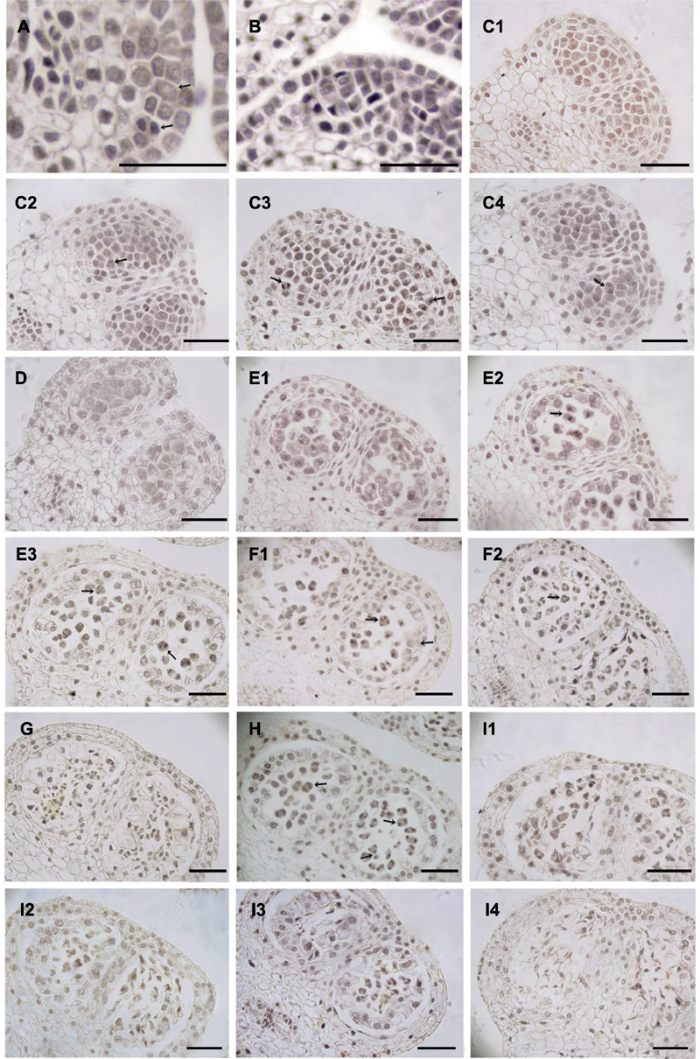 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT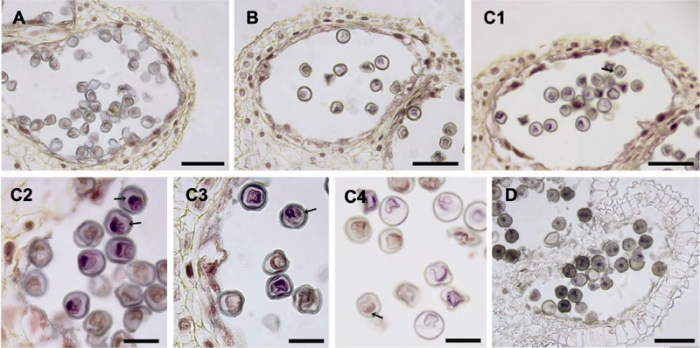 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT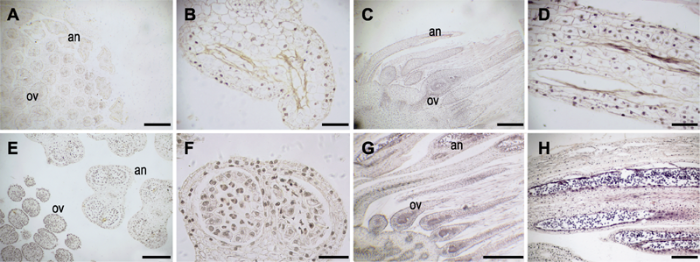 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT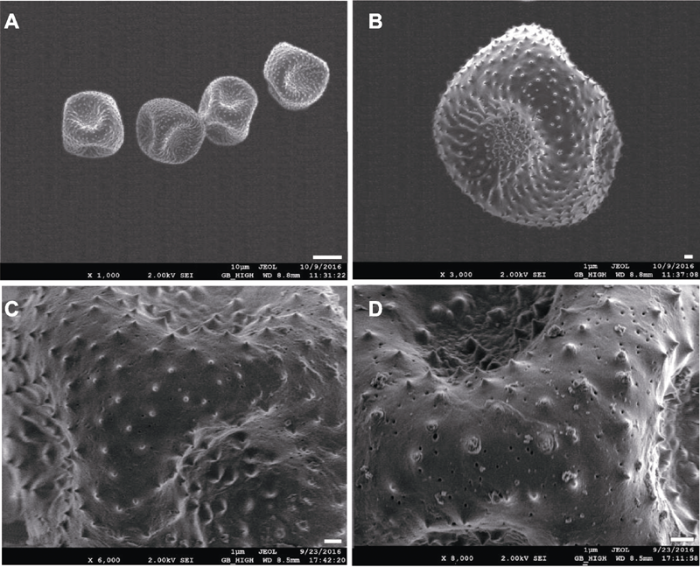 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT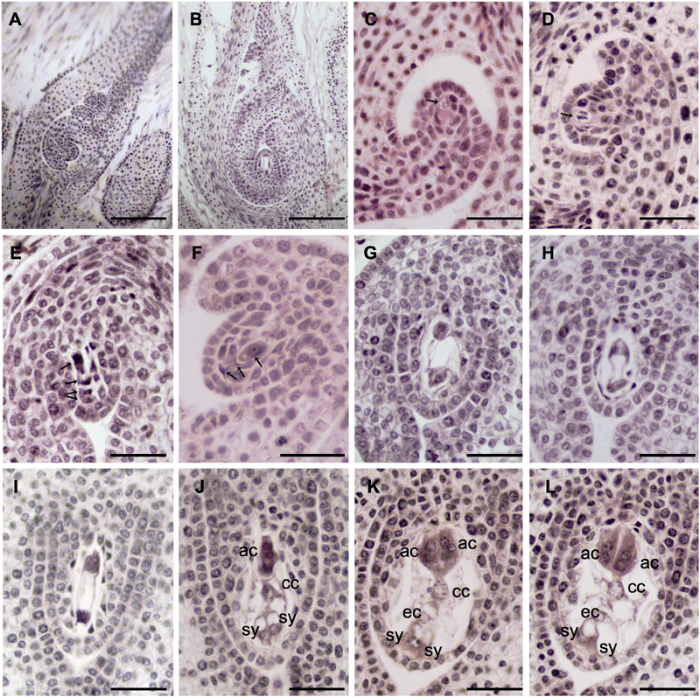 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}