Studies in the Responses of Wheat Root Traits to Drought Stress
Qingxia Miao1,2,3, Yan Fang,1,2,*, Yinglong Chen,1,2,4,*1. State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China; 2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China 3. College of Forestry, Northwest A&F University, Yangling 712100, China; 4. School of Agriculture and Environment, The UWA Institute of Agriculture, the University of Western Australia, Perth 6009, Australia
Abstract Drought stress induces the response of wheat roots, which simultaneously send signals to the aboveground parts stimulating physiological reactions in the aboveground parts, and thus improving drought tolerance of plants. Root architecture traits include morphological traits and three-dimensional geometric structures (i.e, topological structures). The root system architecture not only has genetic stability, but also shows plasticity. The root physiological and biochemical responses to drought stress primarily involve in induced production and changes of root-sourced chemical signals, root cell enzymes, and root osmosis. Under drought stress, plants also alter root anatomical traits and water-uptake kinetics. In this paper, current advances in the studies on root responses to drought stress of wheat (Triticum aestivum) were reviewed with a focus on root system architecture traits, root physiological properties and root anatomical characteristics. The relationship between wheat root properties and drought stress, and the current research constrains were discussed. This review would provide a guidance for future studies on wheat root traits in response to drought stress. Keywords:wheat;root architecture traits;root physiological-biochemical traits;root anatomical traits
PDF (1241KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 苗青霞, 方燕, 陈应龙. 小麦根系特征对干旱胁迫的响应. 植物学报, 2019, 54(5): 652-661 doi:10.11983/CBB19089 Miao Qingxia, Fang Yan, Chen Yinglong. Studies in the Responses of Wheat Root Traits to Drought Stress. Chinese Bulletin of Botany, 2019, 54(5): 652-661 doi:10.11983/CBB19089
AnjumSA, XieXY, WangLC, SaleemMF, ManC, LeiW (2011). Morphological, physiological and biochemical responses of plants to drought stress Afr J Agric Res 6, 2026-2032. [本文引用: 1]
ArmengaudP, BreitlingR, AmtmannA (2004). The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling Plant Physiol 136, 2556-2576. [本文引用: 1]
BandLR, WellsDM, FozardJA, GhetiuT, FrenchAP, PoundMP, WilsonMH, YuL, LiWD, HijaziHI, OhJ, PearceSP, Perez-AmadorMA, YunJ, KramerE, AlonsoJM, GodinC, VernouxT, HodgmanTC, PridmoreTP, SwarupR, KingJR, BennettMJ (2014). Systems analysis of auxin transport in the Arabidopsis root apex Plant Cell 26, 862-875. [本文引用: 1]
BaoF, ShenJJ, BradySR, MudayGK, AsamiT, YangZB (2004). Brassinosteroids interact with auxin to promote lateral root development in Arabidopsis Plant Physiol 134, 1624-1631. [本文引用: 1]
BarracloughPB, KuhlmannH, WeirAH (1989). The effects of prolonged drought and nitrogen fertilizer on root and shoot growth and water uptake by winter wheat J Agron Crop Sci 163, 352-360. [本文引用: 1]
BeckerSR, ByrnePF, ReidSD, BauerleWL, McKayJK, HaleySD (2016). Root traits contributing to drought tolerance of synthetic hexaploid wheat in a greenhouse study Euphytica 207, 213-224. [本文引用: 1]
BerntsonGM (1997). Topological scaling and plant root system architecture: developmental and functional hierarchies New Phytol 135, 621-634. [本文引用: 1]
BothwellJHF, NgCKY (2005). The evolution of Ca 2+ signaling in photosynthetic eukaryotes New Phytol 166, 21-38. [本文引用: 1]
CaseRM, EisnerD, GurneyA, JonesO, MuallemS, VerkhratskyA (2007). Evolution of calcium homeostasis: from birth of the first cell to an omnipresent signaling system Cell Calcium 42, 345-350. [本文引用: 1]
ChenQ, SunJQ, ZhaiQZ, ZhouWK, QiLL, XuL, WangB, ChenR, JiangHL, QiJ, LiXG, PalmeK, LiCY (2011). The basic helix-loop-helix transcription factor MYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis Plant Cell 23, 3335-3352. [本文引用: 1]
de CarvalhoMHC (2008). Drought stress and reactive oxygen species: production, scavenging and signaling Plant Signal Behav 3, 156-165. [本文引用: 1]
DeakKI, MalamyJ (2005). Osmotic regulation of root system architecture Plant J 43, 17-28. [本文引用: 1]
Echevarría-MachadoI, Escobedo-GMRM, Larqué- SaavedraA (2007). Responses of transformed Catharanthus roseus roots to femtomolar concentrations of salicylic acid Plant Physiol Biochem 45, 501-507. [本文引用: 1]
EckertM, KaldenhoffR (2000). Light-induced stomatal movement of selected Arabidopsis thaliana mutants J Exp Bot 51, 1435-1442. [本文引用: 1]
FangB, ShenJJ, BradySR, MudayGK, TadaoA, YangZB (2004). Brassinosteroids interact with auxin to promote lateral root development in Arabidopsis Plant Physiol 134, 1624-1631. [本文引用: 1]
FangY, DuYL, WangJ, WuAJ, QiaoS, XuBC, ZhangSQ, SiddiqueKHM, ChenYL (2017). Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat Front Plant Sci 8, 672. [本文引用: 1]
FitterAH (1987). An architectural approach to the comparative ecology of plant root systems New Phytol 106, 61-77. [本文引用: 1]
GaoJQ, StraussSH, TsaiCJ, FangK, ChenYR, JiangXN, BusovVB (2010). Gibberellins regulate lateral root formation in Populus through interactions with auxin and other hormones Plant Cell 22, 623-639. [本文引用: 1]
GollanT, SchurrU, SchulzeED (1992). Stomatal response to drying soil in relation to changes in the xylem sap composition of Helianthus annuus. I. The concentration of cations, anions, amino acids in, and pH of, the xylem sap Plant Cell Environ 15, 551-559. [本文引用: 1]
GowdaVRP, HenryA, YamauchiA, ShashidharHE, SerrajR (2011). Root biology and genetic improvement for drought avoidance in rice Field Crops Res 122, 1-13. [本文引用: 1]
HanHM, TianZW, FanYH, CuiYK, CaiJ, JiangD, CaoWX, DaiTB (2015). Water-deficit treatment followed by re-watering stimulates seminal root growth associated with hormone balance and photosynthesis in wheat (Triticum aestivum L.) seedlings Plant Growth Regul 77, 201-210. [本文引用: 1]
HartungW, SauterA, HoseE (2002). Abscisic acid in the xylem: where does it come from, where does it go to? J Exp Bot 53, 27-32.
HeXJ, MuRL, CaoWH, ZhangZG, ZhangJS, ChenSY (2005). AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development Plant J 44, 903-916. [本文引用: 1]
IvanovII (2009). Endogenous auxins and branching of wheat roots gaining nutrients from isolated compartments Russ J Plant Physiol 56, 219-223. [本文引用: 1]
KanoM, InukaiY, KitanoH, YamauchiA (2011). Root plasticity as the key root trait for adaptation to various intensities of drought stress in rice Plant Soil 342, 117-128. [本文引用: 1]
KromeK, RosenbergK, DicklerC, KreuzerK, Ludwig- MüllerJ, Ullrich-EberiusC, ScheuS, BonkowskiM (2010). Soil bacteria and protozoa affect root branching via effects on the auxin and cytokinin balance in plants Plant Soil 328, 191-201. [本文引用: 1]
KudoyarovaGR, VysotskayaLB, CherkozyanovaA, DoddIC (2007). Effect of partial rootzone drying on the concentration of zeatin-type cytokinins in tomato (Solanum lycopersicum L.) xylem sap and leaves J Exp Bot 58, 161-168. [本文引用: 1]
LinkohrBI, WilliamsonLC, FitterAH, LeyserHMO (2002). Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis Plant J 29, 751-760. [本文引用: 1]
LiuHY, SunWN, SuWA, TangZC (2006). Co-regulation of water channels and potassium channels in rice Physiol Plant 128, 58-69. [本文引用: 1]
LjungK, BhaleraoRP, SandbergG (2001). Sites and homeostatic control of auxin biosynthesis in Arabidopsis during vegetative growth Plant J 28, 465-474. [本文引用: 1]
LudlowMM, MuchowRC (1990). A critical evaluation of traits for improving crop yields in water-limited environments Adv Agron 43, 107-153. [本文引用: 1]
MacfallJS, JohnsonGA, KramerPJ (1991). Comparative water uptake by roots of different ages in seedlings of loblolly pine (Pinus taeda L.) New Phytol 119, 551-560. [本文引用: 1]
MahouachiJ, ArbonaV, Gómez-CadenasA (2007). Hormonal changes in papaya seedlings subjected to progressive water stress and re-watering Plant Growth Regul 53, 43-51.
NibauC, GibbsDJ, CoatesJC (2008). Branching out in new directions: the control of root architecture by lateral root formation New Phytol 179, 595-614. [本文引用: 1]
OsmontKS, SiboutR, HardtkeCS (2007). Hidden branches: developments in root system architecture Annu Rev Plant Biol 58, 93-113. [本文引用: 1]
ParcyF, GiraudatJ (1997). Interactions between the ABI1 and the ectopically expressed ABI3 genes in controlling abscisic acid responses in Arabidopsis vegetative tissues Plant J 11, 693-702. [本文引用: 1]
PassiouraJB (1991). Soil structure and plant growth Aust J Soil Res 29, 717-728. [本文引用: 1]
ReddyAR, ChaitanyaKV, JuturPP, SumithraK (2004). Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars Environ Exp Bot 52, 33-42. [本文引用: 1]
RichardsRA, PassiouraJB (1989). A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments Aust J Agric Res 40, 943-950.
SaradadeviR, BramleyH, PaltaJA, EdwardsE, SiddiqueKHM (2015). Root biomass in the upper layer of the soil profile is related to the stomatal response of wheat as the soil dries Funct Plant Biol 43, 62-74. [本文引用: 1]
SchachtmanDP, GoodgerJQD (2008). Chemical root to shoot signaling under drought Trends Plant Sci 13, 281-287. [本文引用: 1]
SegalE, KushnirT, MualemY, ShaniU (2008). Water uptake and hydraulics of the root hair rhizosphere Vadose Zone J 7, 1027-1034.
SharmaP, DubeyRS (2005). Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings Plant Growth Regul 46, 209-221. [本文引用: 1]
SiopongcoJDLC, YamauchiA, SalekdehH, BennettJ, WadeLJ (2005). Root growth and water extraction response of doubled-haploid rice lines to drought and rewatering during the vegetative stage Plant Prod Sci 8, 497-508. [本文引用: 1]
SneddenWA, FrommH (2001). Calmodulin as a versatile calcium signal transducer in plants New Phytol 151, 35-66. [本文引用: 1]
SteudleE, PetersonCA (1998). How does water get through roots? J Exp Bot 49, 775-788. [本文引用: 1]
TardieuF, ZhangJ, DaviesWJ (1992). What information is conveyed by an ABA signal from maize roots in drying field soil? Plant Cell Environ 15, 185-191. [本文引用: 1]
TürkanT, BorM, ?zdemirF, KocaH (2005). Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought- sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress Plant Sci 168, 223-231. [本文引用: 1]
TurnerNC (1986). Adaptation to water deficits: a changing perspective Funct Plant Biol 13, 175-190. [本文引用: 1]
UgaY, OkunoK, YanoM (2011). Dro1, a major QTL involved in deep rooting of rice under upland field conditions J Exp Bot 62, 2485-2494.
UgaY, SugimotoK, OgawaS, RaneJ, IshitaniM, HaraN, KitomiY, InukaiY, OnoK, KannoN, InoueH, TakehisaH, MotoyamaR, NagamuraY, WuJZ, MatsumotoT, TakaiT, OkunoK, YanoM (2013). Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions Nat Genet 45, 1097-1102. [本文引用: 1]
VandoorneB, MathieuAS, van den EndeW, VergauwenR, PérilleuxC, JavauxM, LuttsS (2012). Water stress drastically reduces root growth and inulin yield in Cichorium intybus(var. sativum) independently of photosynthesis J Exp Bot 63, 4359-4373. [本文引用: 1]
WangL, HuaDP, HeJN, DuanY, ChenZZ, HongXH, GongZZ (2011). Auxin Response Factor2 (ARF2) and its regulated homeodomain gene HB33 mediate abscisic acid response in Arabidopsis PLoS Genet 7, e1002172. [本文引用: 1]
WassonAP, RichardsRA, ChatrathR, MisraSC, PrasadSVS, RebetzkeGJ, KirkegaardJA, ChristopherJ, WattM (2012). Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops J Exp Bot 63, 3485-3498. [本文引用: 1]
WhiteRG, KirkegaardJA (2010). The distribution and abundance of wheat roots in a dense, structured subsoil-implications for water uptake Plant Cell Environ 33, 133-148.
WuQS, HeXH, ZouYN, LiuCY, XiaoJ, LiY (2012). Arbuscular mycorrhizas alter root system architecture of Citrus tangerine through regulating metabolism of endogenous polyamines Plant Growth Regul 68, 27-35. [本文引用: 1]
XuWF, JiaLG, ShiWM, LiangJS, ZhouF, LiQF, ZhangJH (2013). Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress New Phytol 197, 139-150. [本文引用: 1]
ZhangHM, HanW, De SmetI, TalboysP, LoyaR, HassanA, RongH, JürgensG, PaulKJ, WangMH (2010). ABA promotes quiescence of the quiescent centre and suppresses stem cell differentiation in the Arabidopsis primary root meristem Plant J 64, 764-774. [本文引用: 1]
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
... 根际环境因子(水分除外)包括氮、磷和钾等矿质元素及各种根际微生物(李淑钰和李传友, 2016).矿质元素的吸收对根系构型可塑性有显著影响: 低氮或低磷诱导侧根伸长(Linkohr et al., 2002); 而低钾胁迫抑制侧根生长(Armengaud et al., 2004).根际微生物(丛枝菌根真菌、根瘤菌和根际促生菌等)主要从以下3方面调控根系构型: (1) 影响植物激素的水平和平衡; (2) 调节植株对矿质元素的吸收; (3) 影响碳水化合物的合成和运输(陈伟立等, 2016).水分胁迫下侧根原基受到抑制从而改变根系构型(Deak and Malamy, 2005), 根系构型改变后又反过来影响作物对水分的吸收(Osmont et al., 2007).根系构型的可塑性对作物生长发育及抗旱性至关重要, 因此研究根系理化特性对根系构型可塑性的影响是探究小麦抗旱性的基础. ...
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响 1 2012
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling 1 2004
... 根际环境因子(水分除外)包括氮、磷和钾等矿质元素及各种根际微生物(李淑钰和李传友, 2016).矿质元素的吸收对根系构型可塑性有显著影响: 低氮或低磷诱导侧根伸长(Linkohr et al., 2002); 而低钾胁迫抑制侧根生长(Armengaud et al., 2004).根际微生物(丛枝菌根真菌、根瘤菌和根际促生菌等)主要从以下3方面调控根系构型: (1) 影响植物激素的水平和平衡; (2) 调节植株对矿质元素的吸收; (3) 影响碳水化合物的合成和运输(陈伟立等, 2016).水分胁迫下侧根原基受到抑制从而改变根系构型(Deak and Malamy, 2005), 根系构型改变后又反过来影响作物对水分的吸收(Osmont et al., 2007).根系构型的可塑性对作物生长发育及抗旱性至关重要, 因此研究根系理化特性对根系构型可塑性的影响是探究小麦抗旱性的基础. ...
Systems analysis of auxin transport in the Arabidopsis root apex 1 2014
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Brassinosteroids interact with auxin to promote lateral root development in Arabidopsis 1 2004
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
The effects of prolonged drought and nitrogen fertilizer on root and shoot growth and water uptake by winter wheat 1 1989
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Root traits contributing to drought tolerance of synthetic hexaploid wheat in a greenhouse study 1 2016
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Topological scaling and plant root system architecture: developmental and functional hierarchies 1 1997
The evolution of Ca 2+ signaling in photosynthetic eukaryotes 1 2005
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Evolution of calcium homeostasis: from birth of the first cell to an omnipresent signaling system 1 2007
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
The basic helix-loop-helix transcription factor MYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis 1 2011
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Drought stress and reactive oxygen species: production, scavenging and signaling 1 2008
Osmotic regulation of root system architecture 1 2005
... 根际环境因子(水分除外)包括氮、磷和钾等矿质元素及各种根际微生物(李淑钰和李传友, 2016).矿质元素的吸收对根系构型可塑性有显著影响: 低氮或低磷诱导侧根伸长(Linkohr et al., 2002); 而低钾胁迫抑制侧根生长(Armengaud et al., 2004).根际微生物(丛枝菌根真菌、根瘤菌和根际促生菌等)主要从以下3方面调控根系构型: (1) 影响植物激素的水平和平衡; (2) 调节植株对矿质元素的吸收; (3) 影响碳水化合物的合成和运输(陈伟立等, 2016).水分胁迫下侧根原基受到抑制从而改变根系构型(Deak and Malamy, 2005), 根系构型改变后又反过来影响作物对水分的吸收(Osmont et al., 2007).根系构型的可塑性对作物生长发育及抗旱性至关重要, 因此研究根系理化特性对根系构型可塑性的影响是探究小麦抗旱性的基础. ...
Hormonal interactions and stomatal responses 1 2003
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Responses of transformed Catharanthus roseus roots to femtomolar concentrations of salicylic acid 1 2007
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Light-induced stomatal movement of selected Arabidopsis thaliana mutants 1 2000
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Brassinosteroids interact with auxin to promote lateral root development in Arabidopsis 1 2004
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat 1 2017
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
An architectural approach to the comparative ecology of plant root systems 1 1987
Gibberellins regulate lateral root formation in Populus through interactions with auxin and other hormones 1 2010
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Stomatal response to drying soil in relation to changes in the xylem sap composition of Helianthus annuus. I. The concentration of cations, anions, amino acids in, and pH of, the xylem sap 1 1992
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Root biology and genetic improvement for drought avoidance in rice 1 2011
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Water-deficit treatment followed by re-watering stimulates seminal root growth associated with hormone balance and photosynthesis in wheat (Triticum aestivum L.) seedlings 1 2015
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Abscisic acid in the xylem: where does it come from, where does it go to? 0 2002
AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development 1 2005
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Endogenous auxins and branching of wheat roots gaining nutrients from isolated compartments 1 2009
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Root plasticity as the key root trait for adaptation to various intensities of drought stress in rice 1 2011
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Soil bacteria and protozoa affect root branching via effects on the auxin and cytokinin balance in plants 1 2010
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Effect of partial rootzone drying on the concentration of zeatin-type cytokinins in tomato (Solanum lycopersicum L.) xylem sap and leaves 1 2007
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Cytokinins act directly on lateral root founder cells to inhibit root initiation 1 2007
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis 1 2002
... 根际环境因子(水分除外)包括氮、磷和钾等矿质元素及各种根际微生物(李淑钰和李传友, 2016).矿质元素的吸收对根系构型可塑性有显著影响: 低氮或低磷诱导侧根伸长(Linkohr et al., 2002); 而低钾胁迫抑制侧根生长(Armengaud et al., 2004).根际微生物(丛枝菌根真菌、根瘤菌和根际促生菌等)主要从以下3方面调控根系构型: (1) 影响植物激素的水平和平衡; (2) 调节植株对矿质元素的吸收; (3) 影响碳水化合物的合成和运输(陈伟立等, 2016).水分胁迫下侧根原基受到抑制从而改变根系构型(Deak and Malamy, 2005), 根系构型改变后又反过来影响作物对水分的吸收(Osmont et al., 2007).根系构型的可塑性对作物生长发育及抗旱性至关重要, 因此研究根系理化特性对根系构型可塑性的影响是探究小麦抗旱性的基础. ...
Co-regulation of water channels and potassium channels in rice 1 2006
Hormonal changes in papaya seedlings subjected to progressive water stress and re-watering 0 2007
Branching out in new directions: the control of root architecture by lateral root formation 1 2008
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Hidden branches: developments in root system architecture 1 2007
... 根际环境因子(水分除外)包括氮、磷和钾等矿质元素及各种根际微生物(李淑钰和李传友, 2016).矿质元素的吸收对根系构型可塑性有显著影响: 低氮或低磷诱导侧根伸长(Linkohr et al., 2002); 而低钾胁迫抑制侧根生长(Armengaud et al., 2004).根际微生物(丛枝菌根真菌、根瘤菌和根际促生菌等)主要从以下3方面调控根系构型: (1) 影响植物激素的水平和平衡; (2) 调节植株对矿质元素的吸收; (3) 影响碳水化合物的合成和运输(陈伟立等, 2016).水分胁迫下侧根原基受到抑制从而改变根系构型(Deak and Malamy, 2005), 根系构型改变后又反过来影响作物对水分的吸收(Osmont et al., 2007).根系构型的可塑性对作物生长发育及抗旱性至关重要, 因此研究根系理化特性对根系构型可塑性的影响是探究小麦抗旱性的基础. ...
Interactions between the ABI1 and the ectopically expressed ABI3 genes in controlling abscisic acid responses in Arabidopsis vegetative tissues 1 1997
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Soil structure and plant growth 1 1991
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments 0 1989
Root biomass in the upper layer of the soil profile is related to the stomatal response of wheat as the soil dries 1 2015
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Chemical root to shoot signaling under drought 1 2008
What information is conveyed by an ABA signal from maize roots in drying field soil? 1 1992
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought- sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress 1 2005
Dro1, a major QTL involved in deep rooting of rice under upland field conditions 0 2011
Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions 1 2013
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Water stress drastically reduces root growth and inulin yield in Cichorium intybus(var. sativum) independently of photosynthesis 1 2012
... 根系平面几何构型主要包括根长、根数、根系表面积、生物量、根长密度及其数量和长度等(表1).干旱胁迫下, 小麦根系通过维持较高的根系生物量和增加根长及根密度来增强对水分的吸收(Siopongco et al., 2005; 马富举等, 2012), 但也有根系生长受阻的情况出现, 此时根数和根系表面积减少(Vandoorne et al., 2012), 这可能与小麦品种和干旱程度有关.从土壤分层看, 干旱胁迫下表层土壤的小麦根数减少, 而深层土壤根数、根系生物量和根长密度增加.土壤深层储水可在播种前测量, 故比季节性降雨量更具预测性, 增加深层根系密度有利于充分利用深土层储水(Gowda et al., 2011; Becker et al., 2016; Fang et al., 2017).旱地小麦根虽然可伸长到160 cm以下, 但深层土壤中根系相对较少, 140-160 cm土层的根长密度小于0.1 cm·cm-3, 而表层(0-20 cm)土壤根长密度可达5-9 cm·cm-3, 通过增加80 cm深度以下的根系可以降低干旱带来的减产风险(Barraclough et al., 1989).小麦深层根系主要由初生根发育而来, 因此控制小麦早期初生根的生长发育对旱地小麦抗旱增产具有重要意义(吴永成等, 2004).深层储水利用率较低主要是由于根-土界面的阻力较大, 增加深层根系中根毛的数量和长度可扩大植物根与土壤的接触面积, 从而减小阻力(Passioura, 1991).White和Kirkegaard (2010)观察并量化了澳大利亚田间深层小麦根的位置和根-土界面, 发现根系在土壤中不是均匀生长, 而是利用土壤空隙和通道生长, 并通过根毛接触土壤.在根和土壤间存在间隙的情况下, 主要由根毛负责根与土壤间的水分运输.根系平面几何构型对干旱胁迫的响应因小麦种类、干旱时间及强度而异(Kano et al., 2011).因此, 探索不同小麦品种间根系平面几何构型与水分利用相关机制, 对筛选耐旱品种、维持和提高干旱胁迫条件下作物产量具有重要意义(Uga et al., 2013). ...
Auxin Response Factor2 (ARF2) and its regulated homeodomain gene HB33 mediate abscisic acid response in Arabidopsis 1 2011
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops 1 2012
小麦根系解剖结构及其水分运输途径示意图(<xref ref-type="bibr" 改自rid="b97">Wasson et al., 2012</xref>)
Schematic diagram of wheat root anatomy and its water transport pathway (<xref ref-type="bibr" rid="b97">modified from Wasson et al., 2012</xref>)Figure 2 胁迫. ...
The distribution and abundance of wheat roots in a dense, structured subsoil-implications for water uptake 0 2010
Arbuscular mycorrhizas alter root system architecture of Citrus tangerine through regulating metabolism of endogenous polyamines 1 2012
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress 1 2013
... 根源化学信号主要包括脱落酸、细胞分裂素(CTK)、生长素、木质部pH值和钙离子(Ca2+)等.干旱胁迫下, 小麦根系产生根源信号ABA并运输到地上部, 使叶片中ABA含量显著升高, ABA通过控制气孔开合减弱蒸腾作用, 提高土壤水分的可用性, 减轻干旱胁迫带来的损伤(Tardieu et al., 1992; Saradadevi et al., 2015; 马超等, 2017).此外, 外源喷施ABA也可降低干旱胁迫对小麦产量的抑制影响(谢静静等, 2018).然而, 目前对ABA的研究仍有一定的分歧, Hartung等 (2002)认为表层干燥土壤的根组织中ABA含量增加是导致气孔关闭的主要原因, 而Saradadevi等(2015)在干旱胁迫下并未检测到表层干燥土壤中根组织的ABA浓度增加.目前, 大部分****认为ABA是干旱胁迫下最主要的根源化学信号物质.由于ABA与其它激素具联合作用, 故也受其它激素的影响.例如, IAA与ABA表现出相互拮抗.IAA主要在茎尖分生组织中合成, 通过韧皮部从地上部转移到根部(Ljung et al., 2001), 以促进根分枝和形成不定根(Quint et al., 2009), 并可控制离子的进出和细胞质的pH值, 从而调节保卫细胞的膨压(Eckert and Kal-denhoff, 2000).CTK和IAA共同调节侧根形成以响应干旱胁迫.干旱胁迫下, CTK含量下降(Dodd, 2003). 目前关于CTK的研究主要以玉米素和玉米素核苷为对象, 这些物质由根冠分生组织产生, 影响分生组织细胞的分化速率, 对根系顶端分裂组织大小和根长起负调控作用, 但对地上部生长起促进作用(Ioio et al., 2007; Kudoyarova et al., 2007).水分亏缺条件下, IAA和CTK浓度及其比值(IAA/CTK)均显著降低, 复水后又增加(Han et al., 2015).然而, Mahouachi等(2007)研究表明, 干旱并未改变根组织中的IAA水平, 中度干旱胁迫反而增加了根尖的IAA运输(Xu et al., 2013).因此, 不同程度干旱胁迫下, 调节根系构型的IAA和CTK浓度不同, 具体机制尚不清楚.干旱条件下木质部pH值升高, 其与ABA共同引起气孔关闭(Gollan et al., 1992).Ca2+是干旱胁迫下ABA诱导气孔关闭的第二信使(Bothwell and Ng, 2005; Case et al., 2007), 其主要与蛋白受体互作传递信号到下游. 钙调素(CaM)是目前了解最清楚的Ca2+受体, 其结合Ca2+后激活并调节靶蛋白的活性, 从而引起干旱胁迫下的一系列生理反应(Parcy and Giraudat, 1997; Snedden and Fromm, 2001).根源化学信号是重要的抗旱指标之一, 探明根源化学信号对旱地小麦研究具有重要意义.但目前此方面的研究尚不成熟,其中交互和耦合作用的关键物质及形成的网络还有待揭示. ...
ABA promotes quiescence of the quiescent centre and suppresses stem cell differentiation in the Arabidopsis primary root meristem 1 2010
... 根系构型的可塑性是对逆境适应性的一种体现(陈伟立等, 2016).影响根系构型可塑性的生理因子包括内部激素和根际环境因子(图3).内部激素影响以生长素(IAA)含量及其运输为主(Band et al., 2014), 主要表现在以下3方面: (1) 参与主根的生长; (2) 参与侧根的形成与伸长; (3) 调控根系发育使其适应胁迫环境(Nibau et al., 2008; Krome et al., 2010).干旱胁迫下, 小麦根系IAA的含量增加(王玮等, 2000), 侧根增多(Ivanov, 2009).其它激素(如油菜素内酯(Fang et al., 2004)、茉莉酸(Chen et al., 2011)和脱落酸(ABA) (Zhang et al., 2010; Wang et al., 2011))大部分通过与生长素互作调控根系构型.例如, 油菜素内酯可通过调控IAA的极性运输细微地调节植物的生长发育过程(Bao et al., 2004).外源施加茉莉酸可增强小麦抗旱性并显著增加根系的内源IAA含量(马超等, 2017); ABA通过影响IAA在根系中的含量和极性运输, 进而抑制根尖分生区细胞分裂(袁冰剑等, 2014); 细胞分裂素与IAA的拮抗作用促进小麦分蘖(王振林, 1989), 抑制小麦侧根发育(Laplaze et al., 2007). 此外, 乙烯(He et al., 2005)、赤霉素(Gao et al., 2010)、水杨酸(Echevarría-Machado et al., 2007)和多胺(Wu et al., 2012)也均通过与生长素互作来调节根系的生长发育. ...
 ,1,2,*, 陈应龙
,1,2,*, 陈应龙
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT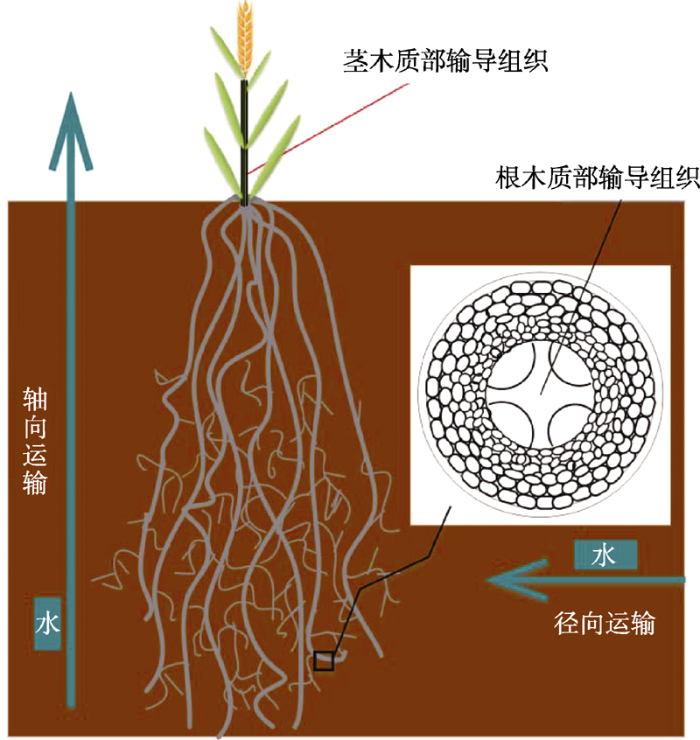 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}