刘进
1 , 2 , 姚晓云
1 , 2 , 余丽琴
1 , 李慧
1 , 周慧颖
1 , 王嘉宇
, 2 , * , 黎毛毛
, 1 , * 1 江西省农业科学院水稻研究所/水稻国家工程实验室(南昌)/国家水稻改良中心南昌分中心, 南昌 330200 2 沈阳农业大学水稻研究所, 沈阳 110866 Detection and Analysis of Dynamic Quantitative Trait Loci at Three Years for Seed Storability in Rice (Oryza sativa ) Jin Liu
1 , 2 , Xiaoyun Yao
1 , 2 , Liqin Yu
1 , Hui Li
1 , Huiying Zhou
1 , Jiayu Wang
, 2 , * , Maomao Li
, 1 , * 1 Nanchang Sub-center, National Rice Improvement Center/National Engineering Laboratory for Rice (Nanchang)/Rice Research Institute, Jiangxi Academy of Agricultural Sciences, Nanchang 330200, China 2 Rice Research Institute, Shenyang Agricultural University, Shenyang 110866, china 通讯作者: * E-mail: ricewjy@126.com * E-mail: Lmm3056@163.com 责任编辑: 朱亚娜
收稿日期: 2018-09-3
接受日期: 2019-02-11
网络出版日期: 2019-07-10
基金资助: 国家重点研发计划 (2016YFD0100101-11 )江西现代农业科研协同创新专项 (JXXTCX201701-01 )江西省青年科学基金 (20171BAB214026 )
Corresponding authors: * E-mail: ricewjy@126.com * E-mail: Lmm3056@163.com Received: 2018-09-3
Accepted: 2019-02-11
Online: 2019-07-10
摘要 种子耐储藏特性是粮食作物的特殊农艺性状之一, 耐储藏性能对种子生产和种质资源保存有重要意义。以粳型超级稻龙稻5 (LD5)和高产籼稻中优早8 (ZYZ8)杂交衍生的重组自交系(RILs)群体(共180个株系)为实验材料, 自然高温高湿条件下放置1年、2年和3年后, 对不同储藏时段种子发芽率进行比较, 并利用223个分子标记的遗传图谱进行动态QTL鉴定。结果表明, 不同储藏时段龙稻5的发芽率均显著低于中优早8, 株系间耐储性存在较大差异; 不同储藏时段发芽率显著相关, 相邻存储时段发芽率关系紧密。共检测到17个耐储性相关的QTLs, 3个老化时段分别检测到5、4和3个, 检测到5个动态条件QTLs, 单一QTL解释5.60%-32.76%的表型变异, 加性效应在-16.78%-16.95%范围内。主效QTL簇qSSC2 、qSSC6 、qSSC7 和qSSC8 能调控不同储藏时段的发芽率, qSSC6 具有明显降低发芽率的效应。共检测到26对上位性互作位点, 主效QTL qSS1 和qSS4 参与上位性互作, 这表明上位性互作是调控耐储藏性状的重要遗传组成。研究结果为水稻(Oryza sativa )耐储性相关QTL的精细定位奠定基础, 同时丰富了耐储性分子标记辅助选择育种的基因资源。 关键词: 水稻 ;
自然老化 ;
种子耐储藏 ;
QTL分析 ;
上位性互作 Abstract Seed storability is one of the most important special agronomic traits of grain crops, which has great significance for seed production and germplasm conservation. In this study, we obtained a set of recombinant inbred lines (RILs), containing 180 lines and 223 molecular markers for a genetic map, derived from the cross between Longdao5 (LD5) and Zhongyouzao8 (ZYZ8). The set was used to identify quantitative trait loci (QTL) for seed storability traits under natural aging (higher temperature and humidity) at 1, 2 and 3 years after rice harvest. Then the germination rates of seeds at different storage stages were compared and identified by dynamic QTLs. The germination rate of the parents (the germination rate of LD5 was significantly lower than ZYZ8 under different storage stages) and RILs population was significant different under different storage stages, with a significant correlation of germination rate under different storage stages. A total of 17 QTLs were detected to control seed storability; 5, 4 and 3 QTLs were detected for 3 natural aging stages; and 5 dynamic conditional QTLs were detected; these QTLs explained 5.60% to 32.76% of the phenotypic variation, with an additive effect of -16.78% to 16.95%. Among these QTLs, 4 major clusters qSSC2 , qSSC6 , qSSC7 and qSSC8 were stably and reliably detected under 3 environments; the remaining QTLs were expressed in a single environment. qSSC6 reduced the germination rate. We detected 26 pairs of epistatic interaction sites. The major QTLs qSS1 and qSS4 were also involved in the epistasis effect, so epistatic interaction was an important genetic component for seed storability. These results will provide information for genetic analysis and related QTL fine mapping and will also enrich the gene resources for molecular marker-assisted selection of seed storability. Keywords: rice ;
natural aging ;
seed storability ;
QTL analysis ;
epistasis interaction PDF (2382KB) 摘要页面 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 刘进, 姚晓云, 余丽琴, 李慧, 周慧颖, 王嘉宇, 黎毛毛 . 水稻耐储藏特性三年动态鉴定与QTL分析. 植物学报, 2019,
54 (4): 464-473 doi:10.11983/CBB18188
Liu Jin, Yao Xiaoyun, Yu Liqin, Li Hui, Zhou Huiying, Wang Jiayu, Li Maomao .
Detection and Analysis of Dynamic Quantitative Trait Loci at Three Years for Seed Storability in Rice (Oryza sativa ) .
Chinese Bulletin of Botany , 2019,
54 (4): 464-473 doi:10.11983/CBB18188
稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 )。种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 )。水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 )。储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 )。近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重。鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 )。
水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 )。其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标。不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 )。此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质。水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 )。不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 )。不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 )。
水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 )。以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 )。近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 )。有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 )。发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义。本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源。
1 材料与方法 1.1 实验材料与设计 以水稻(
Oryza sativa L.)品种龙稻5 (Longdao 5, LD5)和中优早8 (Zhongyouzao 8, ZYZ8)杂交衍生的重组自交系(recombinant inbred lines, RILs)群体为实验材料, 共180个株系(F
7 )。大田试验于2015年在沈阳农业大学试验基地进行, RILs群体每系种植3行, 单苗栽插, 田间管理同当地常规大田栽培管理。
1.2 性状测定 水稻成熟后混合收取1穴稻穗, 混合脱粒, 取亲本和RILs群体各株系材料, 于高温、高湿条件(
表1 )下保存于江西省农业科学院水稻研究所。发芽试验参照
Li等(2017) 的方法进行, 于存储后12个月(2016年)、24个月(2017年)和36个月(2018年)分别测定发芽率, 并以发芽率评价耐储藏特性。每个处理50粒种子, 设3次重复, 取平均值进行后续统计分析。
Table 1 表1 表1 2016-2018南昌市年温度与降雨情况
Table 1
Summary of temperature and rainfall conditions from 2016-2018 in Nanchang city Years Rainy days Max temperature (°C) Days of max temperature (>30°C) Max mean temperature (°C) Mean min temperature (°C) Mean temperature (°C) 2016 192 38 101 22.68 16.10 19.39 2017 184 39 97 23.03 16.10 19.57 2018 (1-9 months) 130 38 119 25.34 17.71 21.53
新窗口打开 |
下载CSV 1.3 遗传图谱构建 在抽穗期采集亲本及RILs群体各家系叶片, 采用CTAB法(
Dogle and Dogle, 1987 )提取水稻基因组DNA。PCR扩增总体积(12 µL)包括2.0 µL DNA模板, 上下游引物(10 μmol∙L
-1 )各1.5 µL, 6 µL 2×Mix混合体系, 1.0 µL ddH
2 O。扩增程序为: 94°C4分钟; 94°C45秒, 55-58°C45秒, 72°C45秒, 32个循环; 72°C10分钟, 12°C保存。PCR扩增产物用4%-5%的琼脂糖凝胶电泳检测。利用亲本间具有明显多态性的标记构建遗传图谱, 共包括223个分子标记(SSR、InDel和ILP标记), 图谱覆盖水稻基因组约1 514.0 cM, 标记间的平均距离为6.79 cM, 平均每对染色体上的标记数为18.58。
1.4 QTL分析 采用QTL ICI Mapping v4.0的完备区间作图(ICIM)方法(
Wang et al., 2014 )进行加性效应、加性×加性上位互作效应QTL分析; LOD的阈值设为2.50, 当实际求得的LOD值大于LOD阈值时, 就判定该区段存在1个QTL; 估算每个QTL的加性效应值和贡献率, 同时应用ICIM-EPI进行上位性分析。QTL的命名遵循
McCouch等(1997) 的原则。
测定第1、2和3年环境下RILs各株系和亲本发芽率, 设定为非条件表型值, 即某一时刻的表型值。根据
朱军(1997) 提出的动态条件遗传效应值的统计分析方法, 可以获得给定的1-2年和2-3年动态变化的发芽率条件表型值, 动态条件QTL的加性效应则为某一时间段内基因表达的净加性效应。以1年和2年差值表示储藏1-2年发芽率变化的动态条件值, 2年和3年差值表示储藏2-3年发芽率变化的动态条件值, 以动态条件值计算动态条件QTL。
2 结果与讨论 2.1 表型分析 不同存储年份时段下, 双亲间发芽率均存在极显著差异。龙稻5发芽率均较低, 中优早8则表现出较高的发芽率, 存储3年后龙稻5的发芽率基本为0, 而中优早8的发芽率下降较慢, 存储3年后具有56.12%的发芽率(
图1A -C); RILs群体存储1年后发芽率变化不明显(
图1A ), 2年后发芽率显著下降(
图1B ), 3年后多数株系发芽率下降至0, 但仍有部分株系表现较强的活力(
图1C ); RILs群体中不同株系发芽率和发芽率动态变化值存在较大幅度变异, 呈现双向超亲分离, 近似于连续的正态分布(
图1A -E;
表2 ), 这表明不同存储时段下发芽率性状均为多基因控制的数量性状, 符合QTL作图的要求。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1自然老化条件下不同存储时间水稻重组自交系(RILs)群体发芽率(GR)分布情况 (A)-(C) 分别表示自然老化(高温高湿)条件下存储1、2和3年后的发芽率; (D), (E) 分别表示1-2年和2-3年环境下发芽率动态变化值。亲本龙稻5 (LD5)和中优早8 (ZYZ8)发芽率分别采用实线和虚线箭头表示。
Figure 1Distribution of germination rates (GR) under different seed dormancy time in the recombinant inbred lines (RILs) population of rice (A)-(C) GR of natural storage condition for 1, 2 and 3 years, respectively; (D), (E) Dynamic GR of natural storage condition for 1-2 and 2-3 years, respectively. GR of the parents (LD5 and ZYZ8) are indicated by black and dotted line arrows, respectively.
Table 2 表2 表2 不同存储年份水稻亲本和重组自交系(RILs)群体发芽率(GR)分布情况
Table 2
Distribution of germination rate (GR) after different aging treatments for parents and recombinant inbred lines (RILs) population of rice Trait Environment (Year) Parents RILs population LD5 ZYZ8 Means±SD Range Skewness Kurtosis GR (%) 1 63.97±6.10 95.95±4.51** 56.34±26.89 0.00-100.00 -0.30 -0.88 2 43.19±4.30 59.79±2.20** 32.92±29.32 0.00-96.55 0.49 -1.04 3 0.00±0.00 56.12±2.05** 6.89±16.80 0.00-87.68 2.92 3.36 Dynamic GR 1-2 20.78±5.20 36.16±2.75** 23.42±29.59 -5.70-97.79 -0.03 0.01 2-3 43.19±5.63 3.67±3.50** 26.02±28.28 -2.9-96.55 0.64 -0.45
**表示在0.01水平差异显著。** represent significant differences at the 0.01 level. 新窗口打开 |
下载CSV 自然老化条件下, 不同储藏时段与发芽率间均呈显著或极显著正相关(
表3 )。相邻储藏时段的发芽率相关系数较大, 储藏1年与2年、2年和3年的发芽率相关系数分别为0.448和0.347, 储藏1年与3年的相关系数仅为0.166。这表明不同储藏时段的发芽率具有相关性, 随着存储时间的推移发芽率逐步降低, 发芽率具有明显的动态变化规律。
Table 3 表3 表3 自然老化条件下不同存储时间水稻发芽率(GR)性状间的相关系数
Table 3
Correlation coefficients among germination rates (GR) of rice under different natural storage conditions Traits Environment (Year) 1 2 3 GR 1 1.000 2 0.448** 1.000 3 0.166* 0.347** 1.000
*和**分别表示在0.05和0.01水平差异显著。* and ** represent significant differences at the 0.05 and 0.01 level, respectively. 新窗口打开 |
下载CSV 2.2 QTL分析 我们共检测到17个控制耐储藏性状相关的QTLs, 3个自然老化时段下分别检测到5、4和3个, 并检测到5个动态条件QTLs, 分布于第1、2、3、4、6、7、8和12号染色体上, LOD值介于2.53-6.52之间, 可解释5.60%-32.76%的表型变异, 加性效应在-16.78% -16.95%范围内(
表4 ;
图2 )。我们还检测到5个调控自然老化1年条件下发芽率的QTLs, 分别位于第3、4、6、7和8号染色体上;
qSS4 增效等位基因来自ZYZ8, 具有提高发芽率的效应;
qSS3 、
qSS6a 、
qSS7a 和
qSS8a 增效等位基因来自LD5, 具有降低存储条件下发芽率的效应;
qSS4 和
qSS6a 具有较高的表型贡献率和加性效应值, 分别为14.49%、13.53%和10.27%、-10.27%。我们共检测到4个控制储存2年后的发芽率QTLs, 分别位于第2、6、7和8号染色体上, LOD值介于2.71-3.86之间, 表型贡献率在5.78%-12.12%, 其中
qSS6b 和
qSS8b 效应明显, 表型贡献率和效应值均超过10.00%, 增效等位基因来自LD5, 具有明显的降低发芽率的功能。我们检测到3个调控存储3年后发芽率位点, 分布在第1、2和7号染色体上,
qSS1 、
qSS2b 和
qSS7c 增效等位基因均来自ZYZ8,
qSS1 贡献率和效应值较大。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2自然存储条件下水稻种子耐储藏性状QTL在重组自交系(RILs)群体中的染色体分布 Figure 2Location of QTLs detected for seed storability under natural storage conditions in rice on the genetic map of recom- binant inbred lines (RILs) Table 4 表4 表4 自然存储条件下水稻种子耐储藏性状QTL定位
Table 4
Putative QTL for seed storability were detected under natural storage conditions in rice Trait Locus Position Marker LOD value PVE (%) Additive effect 1 year qSS3 75.5 RM6676-STS3.8 2.53 10.58 -8.77 qSS4 26.1 R4M17-RM5688 2.85 14.49 10.27 qSS6a 121.4 RM5814-RM412 5.28 13.53 -10.27 qSS7a 96.2 RM8261-RM1209 2.85 7.29 -7.27 qSS8a 85.5 R8M33-RM6976 2.79 6.88 -7.04 2 years qSS2a 40.5 RM5897-RM5699 3.86 10.47 9.69 qSS6b 107.4 RM6395-RM5814 2.88 11.27 -10.19 qSS7b 108.2 RM3555-RM1306 2.71 5.78 -7.03 qSS8b 89.5 R8M33-RM6976 3.79 12.12 -10.18 3 years qSS1 102.5 R1M30-RM3240 3.13 9.20 5.78 qSS2b 50.5 RM5699-RM300 3.01 6.46 4.29 qSS7c 64.2 RM1135-RM11 3.21 10.05 5.32 1-2 years qss2 41.5 RM5897-RM5699 2.98 10.13 -9.59 2-3 years qss3 40.5 STS3.3-STS3.4 4.46 25.22 16.95 qss6 107.4 RM6395-RM5814 6.52 32.76 -16.78 qss8 91.5 R8M33-RM6976 2.54 5.60 -6.68 qss12 86 STS12.2-RM1226 2.89 24.42 -14.64
SS: Seed storability; ss: Dynamic seed storability under different years; LOD: The peak logarithm of odds for QTL; PVE: Percentage of total phenotypic variance explained by the QTL SS: 种子耐储藏; ss: 不同年份年动态耐储藏特性; LOD: QTL的LOD值峰值; PVE: QTL的表型贡献率 新窗口打开 |
下载CSV 我们共检测到5个耐储藏特性动态条件相关的QTLs, 分别位于2、3、6、8和12号染色体上。在存储1-2年时段仅检测到1个相关的QTL
qss2 , 其LOD值、表型贡献率和加性效应值分别为2.98、10.13%和-9.59%, 增效等位基因来自ZYZ8。我们共检测4个与存储2-3年动态条件相关的QTLs,
qss8 的加性效应值较小,
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能, 其中仅
qss3 增效等位基因来自ZYZ8, 具有明显增加发芽率的效应, 其余位点增效等位基因均来自LD5, 具有明显的降低发芽率的功能。
此外, 通过比较分析发现有4个QTLs所在的相关区域能持续调控不同存储时段的发芽率, 分别位于第2、6、7和8号染色体上, 主效QTL簇
qSSC2 (
qSS2a 、
qSS2b 和
qss2 )、
qSSC6 (
qSS6a 、
qSS6b 和
qss6 )、
qSSC7 (
qSS7a 和
qSS7b )和
qSSC8 (
qSS8a 、
qSS8b 和
qss8 );
qSSC6 、
qSSC7 和
qSSC8 能调控存储1和2年后的发芽率, 其中
qSSC6 具有明显降低发芽率的效应, 增效等位基因来自LD5, 贡献率和效应值均值分别介于11.27%-32.76%和-16.78%-10.19%之间;
qSSC2 能持续调控存储2年和3年后的发芽率, 贡献率和效应值分别介于6.46%-10.47%和-9.59%- 9.69%之间, 效应值相对较小。
2.3 上位性分析 采用ICIM-EPI模型检测耐储藏性状上位性效应位点, 共检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75%, 加性效应介于-20.45%-17.72%之间(
图3 )。位于第1、4、5和7号染色体上相应的染色体区域对具有明显的上位性作用, 其中主效QTL
qSS1 所在的第1号染色体R1M30-RM3240区域与第1-7和12号染色体间存在明显的上位性效应;第4号染色体RM5688-RM471区域的主效QTL
qSS4 与第1、5、7、8、10和12号染色体间存在明显的上位性效应。此外, 第5号染色体的R5M13-RM3476区域非加性效应位点与第1-7、9和12号染色体的相应区域存在较强的上位性效应; 第7号染色体RM5711-RM1135区域非加性效应位点与第1、3、4、5和7号染色体的相应区域存在较强的上位性效应。上述研究结果表明, 本研究RILs群体中包含多个调控耐储藏性状的上位性互作位点, 上位性互作效应既可存在于加性效应QTL间和加性效应QTL与非加性效应位点间, 也可存在于非加性效应位点间, 这也表明上位性互作效应是调控水稻耐储藏特性的重要遗传组成之一。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3自然存储条件下水稻种子耐储藏性状QTL的上位性互作效应分析 Figure 3QTL with epistasis effect estimate for seed storability in rice under natural storage conditions 2.4 讨论与结论 本研究表明, 不同储藏时段双亲发芽率均存在极显著差异, 龙稻5耐储藏性低于中优早8, 中优早8存储3年后仍表现较高的发芽率; RILs群体不同株系发芽率存在较大幅度变异, 呈现双向超亲分离; 自然老化条件下, 不同储藏时段发芽率关系紧密, 相邻储藏时段的发芽率相关系数较大, 随着存储时间的推移发芽率逐步降低。存储1年的种子发芽率下降相对较小, 存储2年后种子活力显著下降但仍存在部分种子活力较强的株系, 存储3年后仅少数几个株系具有发芽能力, 这表明稻谷种子最适宜的存储时间为1-2年。
种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 )。外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 )。目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 )。
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb。
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体。
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 )。
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强。
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 。
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应。
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 )。本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率。本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能。此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性。与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 )。
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇。后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制。
不同等位基因间的互作效应对表型值的影响称为上位性互作效应。上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作。诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 )。上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 )。
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素。
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应。
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用。
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异。本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应。这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间。水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制。
[1] 龚继平 , 吴方喜 , 吴跃进 , 郑家团 , 黄庭旭 , 王乌齐 , 张建福 , 谢华安 ( 2008 ). 籼稻脂肪酶基因的遗传分析及定位中国水稻科学 22, 125 -130 . [本文引用: 1] [2] 贺梅 , 张文忠 , 宋冬明 , 王嘉宇 , 谢文孝 ( 2007 ). 不同储藏温度及储藏时间对稻米品质的影响沈阳农业大学学报 38, 472 -477 . [本文引用: 1] [3] 江川 , 王金英 , 丁红萍 , 缪小红 ( 2000 ). 影响水稻种子贮藏的因素福建稻麦科技 18(4 ), 46 -47 . [本文引用: 1] [4] 江良荣 , 黄荣裕 , 黄育民 , 王侯聪 , 郑景生 ( 2017 ). 稻米垩白性状的QTL检测、上位性及环境效应分析分子植物育种 15, 1385 -1394 . [本文引用: 2] [5] 李茂柏 , 王慧 , 朴钟泽 , 沈国辉 , 温广月 ( 2010 ). 杂草稻人工老化和耐储藏特性的初步研究作物杂志 ( 5 ), 30 -33 . [本文引用: 2] [6] 李清华 , 郑苹立 , 吴方喜 , 林玲娜 , 王乌齐 ( 2009 ). 优质耐储藏水稻的初步研究和探讨中国农学通报 25(17 ), 47 -51 . [本文引用: 1] [7] 柳武革 , 王丰 , 刘振荣 , 廖亦龙 , 李金华 ( 2006 ). 水稻耐储藏特性研究进展生物技术通报 (增刊) , 50- 52, 62 . [本文引用: 2] [8] 刘喜 , 林秋云 , 孙爱玲 , 曹鹏辉 , 姜一梅 , 陈亮明 , 江玲 , 万建民 ( 2015 ). 水稻种子耐贮性QTL qSS-9 的精细定位南京农业大学学报 38, 877 -882 . [本文引用: 2] [9] 任德勇 , 何光华 , 凌英华 , 桑贤春 , 杨正林 , 赵芳明 ( 2010 ). 基于单片段代换系的水稻穗长QTL加性及其上位性效应植物学报 45, 662 -669 . [本文引用: 1] [10] 任淦 , 彭敏 , 唐为江 , 徐才国 , 邢永忠 ( 2005 ). 水稻种子衰老相关基因定位作物学报 31, 183 -187 . [本文引用: 1] [11] 商连光 , 高振宇 , 钱前 ( 2017 ). 作物杂种优势遗传基础的研究进展植物学报 52, 10 -18 . [本文引用: 1] [12] 沈圣泉 , 庄杰云 , 王淑珍 , 杨国花 , 夏英武 ( 2005 ). 水稻种子耐贮藏性QTL主效应和上位性效应分析分子植物育种 3, 323 -328 . [本文引用: 1] [13] 吴方喜 , 朱永生 , 谢鸿光 , 张建福 , 谢华安 ( 2010 ). 中国水稻微核心种质的耐储藏特性初步研究中国粮油学报 25(10 ), 124 -128 . [本文引用: 2] [14] 吴贻开 , 陈文杰 , 李清华 , 李清华 , 郑向华 ( 2000 ). 相对湿度对水稻种子贮藏寿命的影响福建农业科技 ( 4 ), 6 -7 . [本文引用: 1] [15] 吴跃进 , 卢义宣 , 吴敬德 , 余增亮 , 张瑛 , 童继平 , 郑乐娅 , 佘德红 ( 2004 ). 耐储藏专用型水稻选育及相关技术研究中国稻米 ( 3 ), 6 -7 . [本文引用: 2] [16] 吴跃进 , 吴先山 , 沈宗海 , 张瑛 , 吴敬德 , 卢义宣 , 余增亮 ( 2005 ). 水稻耐储藏种质创新及相关技术研究粮食储藏 34, 17 -20 . [本文引用: 2] [17] 许惠滨 , 江敏榕 , 连玲 , 朱永生 , 蒋家焕 , 谢鸿光 , 谢华安 , 张建福 ( 2017 ). 稻谷耐储性研究进展福建农业科技 ( 11 ), 41 -44 . [本文引用: 2] [18] 许惠滨 , 魏毅东 , 连玲 , 朱永生 , 谢华安 , 王宗华 , 张建福 ( 2013 ). 水稻种子人工老化与自然老化的分析比较分子植物育种 11, 552 -556 . [19] 余丽琴 , 熊玉珍 , 黎二姝 , 饶淑芳 ( 2008 ). 水稻耐贮藏种质资源的筛选江西农业学报 20(4 ), 17 -19 . [本文引用: 1] [20] 曾大力 , 钱前 , 国广泰史 , 滕胜 , 藤本宽 ( 2002 ). 稻谷储藏特性及其与籼粳特性的关系研究作物学报 28, 551 -554 . [本文引用: 2] [21] 张安鹏 , 钱前 , 高振宇 ( 2018 ). 水稻种子活力的研究进展中国水稻科学 32, 296 -303 . [本文引用: 1] [22] 张文明 , 倪安丽 , 王昌初 ( 1998 ). 杂交水稻种子活力的研究种子 ( 2 ), 7 -10 . [本文引用: 1] [23] 张瑛 , 滕斌 , 吴敬德 , 吴跃进 , 宣红 , 朱学桂 ( 2010 ). 水稻种子高温高湿人工加速老化试验方法研究中国粮油学报 25(10 ), 8 -12 . [本文引用: 1] [24] 张玉兰 , 汪晓峰 , 景新明 , 林坚 ( 2005 ). 水稻种子含水量及其对贮藏寿命的影响中国农业科学 38, 1480 -1486 . [本文引用: 1] [25] 周玉亮 , 刘春保 , 潘招远 , 谭斌 , 曾瑞珍 ( 2016 ). 水稻种子休眠的QTL定位研究进展中国科技论文 11, 2837 -2844 . [本文引用: 1] [26] 朱军 ( 1997 ). 遗传模型分析方法 . 北京: 中国农业出版社. pp. 240 -292 . [本文引用: 1] [27] Cai HW Morishima H 2000 ). Genomic regions affecting seed shattering and seed dormancy in riceTheor Appl Genet 840 -846 . [本文引用: 1] [28] Cheng JP Wang L Du WL Lai YY Huang X Wang ZF Zhang HS 2014 ). Dynamic quantitative trait locus analysis of seed dormancy at three development stages in riceMol Breeding 501 -510 . [本文引用: 2] [29] Dong XY Fan SX Liu J Wang Q Li MR Jiang X Liu ZY Yin YC Wang JY 2017 ). Identification of QTLs for seed storability in rice under natural aging conditions using two RILs with the same parent Shennong 265J Integr Agr 1084 -1092 . [本文引用: 3] [30] Doyle JJ Doyle JL 1987 ). A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 11 -15 . [本文引用: 1] [31] Gao YM Zhu J 2007 ). Mapping QTLs with digenic epistasis under multiple environments and predicting heterosis ba- sed on QTL effectsTheor Appl Genet 325 -333 . [本文引用: 1] [32] Gu XY Kianian SF Foley ME 2004 ). Multiple loci and epistases control genetic variation for seed dormancy in weedy rice (Oryza sativa )Genetics 1503 -1516 . [本文引用: 1] [33] Guo LB Zhu LH Xu YB Zeng DL Wu P Qian Q 2004 ). QTL analysis of seed dormancy in rice (Oryza sativa L.)Euphytica 155 -162 . [本文引用: 2] [34] Hang NT Lin QY Liu LL Liu X Liu SJ Wang WY Li LF He NQ Liu Z Jiang L Wan JM 2015 ). Mapping QTLs related to rice seed storability under natural and artificial aging storage conditionsEuphytica 673 -681 . [本文引用: 4] [35] Jiang WZ Lee J Jin TM Qiao YL Piao RH Jang SM Woo MO Kwon SW Liu XH Pan HY Du XL Koh HJ 2011 ). Identification of QTLs for seed germination capability after various storage periods using two RIL populations in riceMol Cells 385 -392 . [本文引用: 1] [36] Jing W Jiang L Zhang WW Zhai HQ Wan JM 2008 ). Mapping QTL for seed dormancy in weedy riceActa Agron Sin 737 -742 . [本文引用: 1] [37] Li CS Shao GS Wang L Wang ZF Mao YJ Wang XQ Zhang XH Liu ST Zhang HS 2017 ). QTL Identification and fine mapping for seed storability in rice (Oryza sativa L.)Euphytica 127 . [本文引用: 5] [38] Li LF Lin QY Liu SJ Liu X Wang WY Hang NT Liu F Zhao ZG Jiang L Wan JM 2012 ). Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.)Plant Breed 739 -743 . [本文引用: 1] [39] Lu BY Xie K Yang CY Wang SF Liu X Zhang L Jiang L Wan JM 2011 ). Mapping two major effect grain dormancy QTL in riceMol Breeding 453 -462 . [本文引用: 2] [40] McCouch SR Cho YG Yano M Paul E Blinstrub M Morishima H Kinoshita T 1997 ). Report on QTL nomenclatureRice Genet Newsl 11 -13 . [本文引用: 1] [41] Miura K Lin Y Yano M Nagamine T 2002 ). Mapping quantitative trait loci controlling seed longevity in rice (Oryza sativa L.)Theor Appl Genet 981 -986 . [本文引用: 1] [42] Sasaki K Fukuta K Sato T 2005 ). Mapping of quantitative trait loci controlling seed longevity of rice (Oryza sativa L.) after various periods of seed storagePlant Breed 361 -366 . [本文引用: 1] [43] Shigemune A Miura K Sasahara H Goto A Yoshida T 2008 ). Role of maternal tissues in qLG-9 control of seed longevity in rice( Oryza sativa L.)Breed Sci 1 -5 . [本文引用: 2] [44] Wan JM Cao YJ Wang CM Ikehashi H 2005 ). Quantitative trait loci associated with seed dormancy in riceCrop Sci 712 -716 . [本文引用: 1] [45] Wang JK Li HH Zhang LY 2014 ). QTL ICI Mapping V4.0 [2014]. URL [本文引用: 1] [46] Xie K Jiang L Lu BY Yang CY Li LF Liu X Zhang L Zhao ZG Wan JM 2011 ). Identification of QTLs for seed dormancy in rice (Oryza sativa L.)Plant Breed 328 -332 . [本文引用: 1] [47] Xue Y Zhang SQ Yao QH Peng RH Xiong AS Li X Zhu WM Zhu YY Zha DS 2008 ). Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.)Euphytica 739 -744 . [本文引用: 1] [48] Zeng DL Guo LB Xu YB Yasukumi K Zhu LH Qian Q 2006 ). QTL analysis of seed storability in ricePlant Breed 57 -60 . [本文引用: 2] [49] Zhang ZH Yu SB Yu T Huang Z Zhu YG 2005 ). Mapping quantitative trait loci (QTLs) for seedling-vigor using recombinant inbred lines of rice (Oryza sativa L.)Field Crop Res 161 -170 . [本文引用: 1] [50] Zhuang JY Fan YY Rao ZM Wu JL Xia YW Zheng KL 2002 ). Analysis on additive effects and additive- by-additive epistatic effects of QTLs for yield traits in a recombinant inbred line population of riceTheor Appl Genet 1137 -1145 . [本文引用: 1] 籼稻脂肪酶基因的遗传分析及定位
1
2008
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
不同储藏温度及储藏时间对稻米品质的影响
1
2007
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
影响水稻种子贮藏的因素
1
2000
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
稻米垩白性状的QTL检测、上位性及环境效应分析
2
2017
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
... ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
杂草稻人工老化和耐储藏特性的初步研究
2
2010
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
... ;
李茂柏等, 2010 ). ...
优质耐储藏水稻的初步研究和探讨
1
2009
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
水稻耐储藏特性研究进展
2
2006
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
... ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
水稻种子耐贮性QTL
qSS-9 的精细定位
2
2015
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
基于单片段代换系的水稻穗长QTL加性及其上位性效应
1
2010
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
水稻种子衰老相关基因定位
1
2005
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
作物杂种优势遗传基础的研究进展
1
2017
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
水稻种子耐贮藏性QTL主效应和上位性效应分析
1
2005
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
中国水稻微核心种质的耐储藏特性初步研究
2
2010
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
... ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
相对湿度对水稻种子贮藏寿命的影响
1
2000
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
耐储藏专用型水稻选育及相关技术研究
2
2004
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
水稻耐储藏种质创新及相关技术研究
2
2005
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
稻谷耐储性研究进展
2
2017
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
... ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
水稻种子人工老化与自然老化的分析比较
0
2013
水稻耐贮藏种质资源的筛选
1
2008
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
稻谷储藏特性及其与籼粳特性的关系研究
2
2002
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
... ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
水稻种子活力的研究进展
1
2018
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
杂交水稻种子活力的研究
1
1998
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
水稻种子高温高湿人工加速老化试验方法研究
1
2010
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
水稻种子含水量及其对贮藏寿命的影响
1
2005
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
水稻种子休眠的QTL定位研究进展
1
2016
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
1
1997
... 测定第1、2和3年环境下RILs各株系和亲本发芽率, 设定为非条件表型值, 即某一时刻的表型值.根据
朱军(1997) 提出的动态条件遗传效应值的统计分析方法, 可以获得给定的1-2年和2-3年动态变化的发芽率条件表型值, 动态条件QTL的加性效应则为某一时间段内基因表达的净加性效应.以1年和2年差值表示储藏1-2年发芽率变化的动态条件值, 2年和3年差值表示储藏2-3年发芽率变化的动态条件值, 以动态条件值计算动态条件QTL. ...
Genomic regions affecting seed shattering and seed dormancy in rice
1
2000
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
Dynamic quantitative trait locus analysis of seed dormancy at three development stages in rice
2
2014
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
Identification of QTLs for seed storability in rice under natural aging conditions using two RILs with the same parent Shennong 265
3
2017
... 稻谷储藏过程中陈化变质、种子活力和发芽率下降一直是水稻(
Oryza sativa )生产、储存与经营中面临的主要问题, 也是水稻育种工作的难题之一(
许惠滨等, 2017 ;
张安鹏等, 2018 ).种子耐储藏特性是重要的农艺性状, 显著影响种子发芽率和植株活力, 与其子代的生产和管理密切相关(
张文明等, 1998 ).水稻是不耐储藏的粮食作物之一, 稻谷储藏时由于呼吸、氧化和酶的作用而发生一系列化学变化, 储存2年后开始劣变, 适宜的储存年限一般为2-3年(
余丽琴等, 2008 ;
许惠滨等, 2017 ).储藏过程中种子活力和发芽率随着储藏时间的延长而降低, 陈化的种子糙米表面色泽变暗、蒸煮食味品质变差、营养成分被破坏, 导致种子的播种品质和食用品质下降(
江川等, 2000 ;
柳武革等, 2006 ;
贺梅等, 2007 ).近年来, 我国稻谷连年丰收使得稻谷储藏问题越来越凸显, 在气候湿热的南方尤为严重.鉴定和利用优异种质与基因, 培育耐储藏新品种对提高水稻耐储特性和粮食贮藏、生产及种质资源保存具有重要的理论和现实意义(
柳武革等, 2006 ;
刘喜等, 2015 ;
Dong et al., 2017 ). ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
A rapid DNA isolation procedure for small quantities of fresh leaf tissue
1
1987
... 在抽穗期采集亲本及RILs群体各家系叶片, 采用CTAB法(
Dogle and Dogle, 1987 )提取水稻基因组DNA.PCR扩增总体积(12 µL)包括2.0 µL DNA模板, 上下游引物(10 μmol∙L
-1 )各1.5 µL, 6 µL 2×Mix混合体系, 1.0 µL ddH
2 O.扩增程序为: 94°C4分钟; 94°C45秒, 55-58°C45秒, 72°C45秒, 32个循环; 72°C10分钟, 12°C保存.PCR扩增产物用4%-5%的琼脂糖凝胶电泳检测.利用亲本间具有明显多态性的标记构建遗传图谱, 共包括223个分子标记(SSR、InDel和ILP标记), 图谱覆盖水稻基因组约1 514.0 cM, 标记间的平均距离为6.79 cM, 平均每对染色体上的标记数为18.58. ...
Mapping QTLs with digenic epistasis under multiple environments and predicting heterosis ba- sed on QTL effects
1
2007
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
Multiple loci and epistases control genetic variation for seed dormancy in weedy rice (
Oryza sativa )
1
2004
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
QTL analysis of seed dormancy in rice (
Oryza sativa L.)
2
2004
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
Mapping QTLs related to rice seed storability under natural and artificial aging storage conditions
4
2015
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Identification of QTLs for seed germination capability after various storage periods using two RIL populations in rice
1
2011
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
Mapping QTL for seed dormancy in weedy rice
1
2008
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
QTL Identification and fine mapping for seed storability in rice (
Oryza sativa L.)
5
2017
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 水稻成熟后混合收取1穴稻穗, 混合脱粒, 取亲本和RILs群体各株系材料, 于高温、高湿条件(
表1 )下保存于江西省农业科学院水稻研究所.发芽试验参照
Li等(2017) 的方法进行, 于存储后12个月(2016年)、24个月(2017年)和36个月(2018年)分别测定发芽率, 并以发芽率评价耐储藏特性.每个处理50粒种子, 设3次重复, 取平均值进行后续统计分析. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
Identification of quantitative trait loci for seed storability in rice (
Oryza sativa L.)
1
2012
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Mapping two major effect grain dormancy QTL in rice
2
2011
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Report on QTL nomenclature
1
1997
... 采用QTL ICI Mapping v4.0的完备区间作图(ICIM)方法(
Wang et al., 2014 )进行加性效应、加性×加性上位互作效应QTL分析; LOD的阈值设为2.50, 当实际求得的LOD值大于LOD阈值时, 就判定该区段存在1个QTL; 估算每个QTL的加性效应值和贡献率, 同时应用ICIM-EPI进行上位性分析.QTL的命名遵循
McCouch等(1997) 的原则. ...
Mapping quantitative trait loci controlling seed longevity in rice (
Oryza sativa L.)
1
2002
... 水稻耐储藏性常采用发芽率、发芽势和发芽指数等指标进行评价(
张瑛等, 2010 ).其中发芽率是指在规定时间内正常发芽种子数占供试种子数的百分率, 测定方法简单且贴近生产, 常被作为种子耐储性研究的直接指标.不同品种间耐储藏特性存在显著差异, 提高水稻种子耐储藏特性有利于提高种子活力和延长水稻贮存时间(
李茂柏等, 2010 ).此外, 具有良好耐储性的种子在长时间储藏后仍然表现出较高的出苗能力和较好的食用品质.水稻的耐储性受内外因素综合影响, 外在因素主要包括种子成熟度、含水量、储藏条件及加工处理等环境因素; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴贻开等, 2000 ;
曾大力等, 2002 ;
张玉兰等, 2005 ;
吴方喜等, 2010 ).不同品种类型的水稻耐储性存在明显差异, 籼粳形态指数与耐储性存在极显著正相关, 一般籼稻耐储性强于粳稻(
Miura et al., 2002 ;
曾大力等, 2002 ).不同来源的种质耐储藏特性差异很大, 而良好耐储特性资源具有较好的利用价值(
吴方喜等, 2010 ;
李茂柏等, 2010 ). ...
Mapping of quantitative trait loci controlling seed longevity of rice (
Oryza sativa L.) after various periods of seed storage
1
2005
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Role of maternal tissues in
qLG-9 control of seed longevity in rice(
Oryza sativa L.)
2
2008
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Quantitative trait loci associated with seed dormancy in rice
1
2005
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
1
2014
... 采用QTL ICI Mapping v4.0的完备区间作图(ICIM)方法(
Wang et al., 2014 )进行加性效应、加性×加性上位互作效应QTL分析; LOD的阈值设为2.50, 当实际求得的LOD值大于LOD阈值时, 就判定该区段存在1个QTL; 估算每个QTL的加性效应值和贡献率, 同时应用ICIM-EPI进行上位性分析.QTL的命名遵循
McCouch等(1997) 的原则. ...
Identification of QTLs for seed dormancy in rice (
Oryza sativa L.)
1
2011
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
Identification of quantitative trait loci for seed storability in rice (
Oryza sativa L.)
1
2008
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
QTL analysis of seed storability in rice
2
2006
... 水稻谷粒包含3种脂肪氧化酶同工酶(Lox1、Lox2和Lox3), 其中Lox3含量相对较多, 其活性降低后可减轻脂质氧化反应、延缓种子衰老、增加稻谷的耐储藏性, 对稻谷的耐储性具有决定作用(
任淦等, 2005 ;
龚继平等, 2008 ;
李清华等, 2009 ).以往主要通过化学药剂、控温和控湿来延长贮藏时间, 其成本高且易污染稻米, 而对耐储藏品种和基因资源的筛选和利用研究相对较少(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ).近年来, 研究者利用不同的群体开展了水稻耐储性相关研究(
Cai and Morishima, 2000 ;
Jiang et al., 2011 ), 相关QTL分布于水稻12条染色体上, 仅有少数几个QTL被精细定位, 贡献率较大的主效QTL主要有
qLG9 (
Shigemune et al., 2008 )、
qSdn-1 和
qSdn-5 (
Lu et al., 2011 )、
qSSh3 和
qSSh7 (
Hang et al., 2015 )、
qSS9 (
刘喜等, 2015 )和
qGP9 (
Li et al., 2017 ).有关水稻耐储藏特性的遗传与分子调控机理仍不甚清楚, 诸多问题亟待解决, 仍需对稻谷耐储藏特性相关基因的分子调控机理进行深入研究(
Xue et al., 2008 ;
Jing et al., 2008 ;
Xie et al., 2011 ).发掘水稻耐储藏种质资源和优异基因, 进一步利用优异基因从根本上解决稻谷储藏过程中发芽率和活力下降与食味品质变差的难题, 对培育新的耐储藏良种具有重要意义.本研究以龙稻5 (不耐储藏)和中优早8 (耐储藏)杂交衍生的重组自交系(RILs)群体为实验材料, 自然老化(高温高湿)条件下存储1、2和3年后对耐储藏特性进行动态鉴定和QTL分析, 探究不同存储时间下种子活力动态变化规律和遗传特性, 发掘新的主效QTL, 为培育强耐储水稻品种奠定理论基础和提供基因资源. ...
... 种子耐储藏性是复杂的数量性状, 受种子发育期间环境条件、储藏条件和遗传基因等因素的共同影响(
Sasaki et al., 2005 ;
Wan et al., 2005 ;
Li et al., 2012 ).外在因素主要包括种子成熟度、储藏条件和加工处理等; 内在因素主要是遗传因素, 包括籼粳特性、脂肪酸含量、淀粉特性、抗氧化物和籽粒大小等(
吴跃进等, 2004 ,
2005 ;
Zeng et al., 2006 ;
周玉亮等, 2016 ).目前, 已有许多研究者利用不同的群体相继开展了水稻种子耐储性相关QTL分析, 耐储性是受多基因控制的复杂的数量性状, 相关QTL分布于1、2、3、4、5、6、7、9、11和12号染色体(
Hang et al., 2015 ;
Li et al., 2017 ;
Dong et al., 2017 ).
Li等(2017) 利用DH和CSSL群体检测到19个耐储藏相关的QTL位点, 主效QTL成簇分布于第1、3、4、8和9号染色体上, 并将具有明显效应的主效QTL
qGP9 精细定位在第9号染色体的标记P6-P8之间, 物理距离为90.2 kb.
Lu等(2011) 检测到2个耐储藏相关的QTLs (
qSdn-1 和
qSdn-5 ), 分别位于第1和5号染色体.
Shigemune等(2008) 利用CSSL群体检测到3个QTLs (
qLG-2 、
qLG-4 和
qLG-9 ).
Gu等(2004) 利用DH群体检测到4个耐储藏相关QTLs, 分别位于第3、5、6和11号染色体, 其中
qSD4 在不同群体中重演性较强.
Hang等(2015) 利用BILs和CSSL群体检测到16个耐储藏相关的QTLs, 分布于第1、2、3、4、5、7、11和12号染色体, 其中
qSSh-2-1 和
qSSh-2-2 能在不同环境下被检测到, 其增效等位基因来自Habataki, 并利用CSSL验证了
qSSh-1 、
qSSh-3 、
qSSh-4 、
qSSh-7 和
qSSh-11 .
Cheng等(2014) 利用RILs群体对水稻耐储藏特性进行QTL分析, 检测到10个耐储藏相关的QTL位点, 分别位于第1、2、3、4、6、8和9号染色体上, 其中
qSD3.1 、
qSD3.2 、
qSD4 和
qSD9 贡献率和效应值均较大, 具有明显的遗传效应.
沈圣泉等(2005) 采用人工老化方法, 检测到2个与耐储藏相关的主效QTLs (
qSS4 和
qSS9 ).本研究共检测到控制耐储藏性状相关的17个QTLs, 3个自然老化时段下分别检测到5、4和3个, 位于第2、6、7和8号染色体上的主效QTL簇
qSSC2 、
qSSC6 、
qSSC7 和
qSSC8 能调控不同存储时段的发芽率QTL,
qSSC6 、
qSSC7 和
qSSC8 具有明显遗传效应, 能调控存储2-3年后的发芽率.本研究还检测到5个耐储藏性动态条件相关的QTLs (
qss2 、
qss3 、
qss6 、
qss8 和
qss12 ), 其中
qss3 、
qss6 和
qss12 的LOD值、表型贡献率和加性效应值均较大, 具有明显的调控发芽率的功能.此外, 通过比较分析, 发现非条件QTL
qSS2 、
qSS6 和
qSS8 与条件QTL
qss2 、
qss6 和
qss8 所在区间基本一致, 其它条件与非条件QTL定位区间不同, 这表明控制耐储藏特性的QTL可能分段表达, 与耐储藏性和发芽率降低动态变化过程相符, 耐储藏相关基因的表达具有时空性.与前人研究结果相比, 本研究检测到的主效QTL簇
qSSC6 与
Guo等(2004) 和
Dong等(2017) 检测到的
qSD6 和
qSC6 位置相近,
qSSC7 与
qSSh-7-1 、
qSSh-7-2 位于相同区间(
Hang et al., 2015 ).
qSSC2 和
qSSC8 是新鉴定的2个主效QTL簇.后续我们将进一步对2个新的主效QTL簇进行遗传解析, 以期精细定位相关QTL, 并阐述相关的基因功能与生理机制. ...
Mapping quantitative trait loci (QTLs) for seedling-vigor using recombinant inbred lines of rice (
Oryza sativa L.)
1
2005
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
Analysis on additive effects and additive- by-additive epistatic effects of QTLs for yield traits in a recombinant inbred line population of rice
1
2002
... 不同等位基因间的互作效应对表型值的影响称为上位性互作效应.上位性互作效应的类型主要有3种, 即主效QTL间、主效QTL与非QTL位点间及2个非主效QTL间的互作.诸多研究表明, 上位性互作在数量性状遗传体系中具有普遍性, QTL普遍参与互作且互作效应存在于各种位点间(
Zhuang et al., 2002 ;
Gao and Zhu, 2007 ;
江良荣等, 2017 ).上位性互作是复杂数量性状遗传的重要组成部分, 也是作物杂种优势的遗传基础和进化的动力(
Zhang et al., 2005 ;
任德勇等, 2010 ;
商连光等, 2017 ).
江良荣等(2017) 对垩白性状进行QTL分析, 共检测到5对上位性效应QTLs, 上位性互作QTL单一位点贡献率不大, 互作位点较多, 综合贡献率较大, 上位性互作QTL也是稻米垩白形成的一个重要因素.
Guo等(2004) 利用DH群体检测到9对耐储藏相关的上位性QTLs, 位于第1、2、3和6号染色体的相应染色体区域具有明显的上位性互作效应.
Cheng等(2014) 利用RILs群体对耐储藏特性进行上位性QTL分析, 共检测到8对耐储藏相关的上位性位点, 其中第1、2和5号染色体上的区域存在明显的上位性作用.
Li等(2017) 利用DH和CSSL群体检测到9对耐储藏相关的上位性位点, 多个主效QTL参与上位性互作, 上位性位点能引起较大的表型变异.本研究检测到26对上位性互作位点, 分布于1-10和12号染色体上, LOD值和表型贡献率均值分别为13.71和24.75, 加性效应介于-20.45%-17.72%之间; 其中部分上位性位点在主效QTL分析中被检测到, 占上位性互作位点总数的40.00%, 而其它位点在主效QTL分析中未检测到; 位于第1、4、5和7号染色体上的相应染色体区域具有明显的上位性效应, 其中主效QTL
qSS1 和
qSS4 与相应染色体区域的上位性互作效应较强; 而第5和7号染色体的R5M13-RM3476区间和RM5711-RM1135区间的非加性效应位点所在的区域与相关染色体区域存在明显的上位性互作效应.这表明, 上位性互作是调控水稻耐储藏性状的重要遗传组成, 存在多个调控耐储藏特性上位性互作位点, 这些上位性互作存在于加性效应QTL间, 也可存在于加性效应QTL与非加性效应位点间及非加性效应位点间.水稻耐储藏特性不仅由加性效应QTL决定, 还受非等位基因之间的互作或多重互作效应的基因位点影响, 耐储藏特性具有受多基因调控的复杂分子网络机制. ...
备案号:
京ICP备16067583号-21 版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由
北京玛格泰克科技发展有限公司 设计开发
 ,2,*, 黎毛毛
,2,*, 黎毛毛
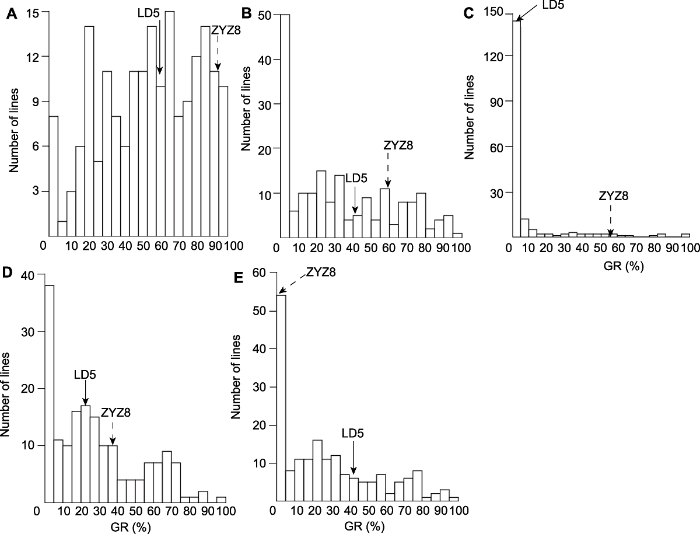 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT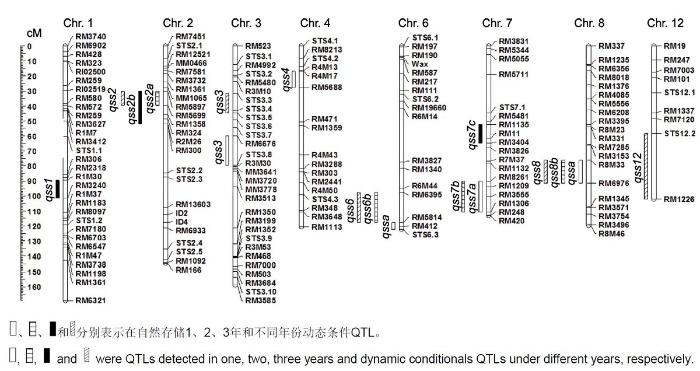 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT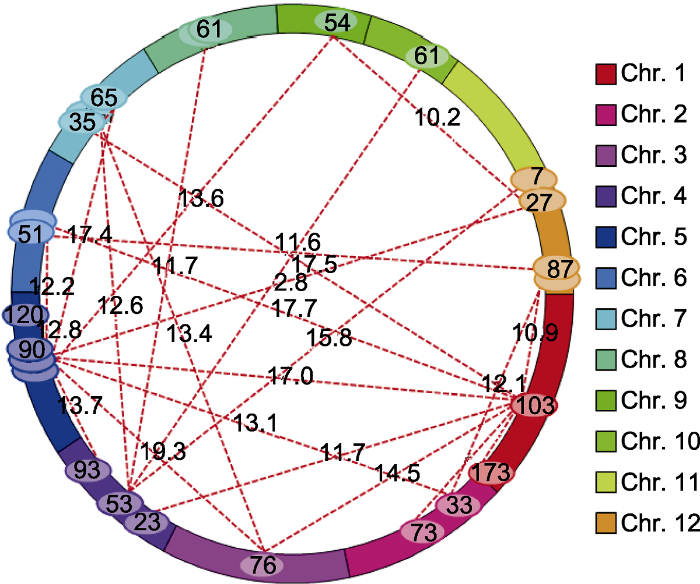 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}