Xinjie Cheng1, Hengxiu Yu1, Zhukuan Cheng,2,*1 Agriculture College, Yangzhou University, Yangzhou 225009, China 2 Center for Plant Gene Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China
Abstract The development of techniques to analyze pachytene chromosomes has greatly overcome most of the difficulties in cytological studies of rice chromosomes caused by their small size. Visualization of meiotic chromosomes has now become routine in cytogenetic studies of this species. This chapter provides protocols on basic meiotic chromosome preparation, FISH analysis and immunocytology in rice. Keywords:rice;meiosis;chromosome;FISH;immunostaining
PDF (1030KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 程新杰, 于恒秀, 程祝宽. 水稻减数分裂染色体分析方法. 植物学报, 2019, 54(4): 503-508 doi:10.11983/CBB19087 Cheng Xinjie, Yu Hengxiu, Cheng Zhukuan. Protocols for Analyzing Rice Meiotic Chromosomes. Chinese Bulletin of Botany, 2019, 54(4): 503-508 doi:10.11983/CBB19087
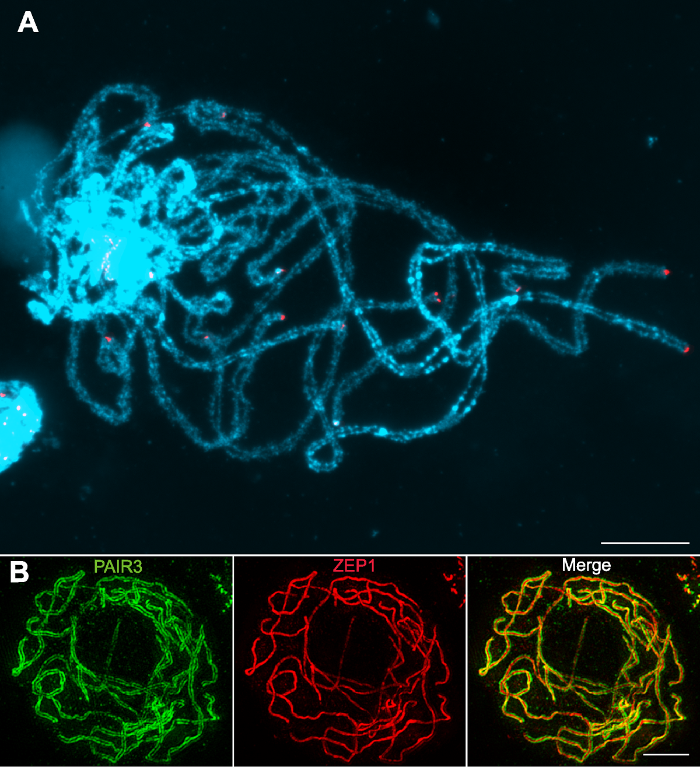
(A) Fluorescent in situ hybridization using rice pachytene chromosomes probed with the telomeric sequence pAtT4 (red); Chromosomes stained with DAPI (blue) (Images taken with a regular fluorescence microscope); (B) Immunodetection of PAIR3 (green) and ZEP1 (red) in rice pachytene chromosomes (Images taken with a super resolution microscope). Bars=5 µm
1 材料和试剂
1.1 染色体制备
(1) 卡诺固定液: 无水乙醇+冰醋酸按3:1 (v/v)配制。
(2) 1%醋酸洋红染液: 将1.0 g洋红粉末和100 mL 45%醋酸加入到500 mL的烧杯中, 在装有回流柱的酒精灯上煮沸8小时。然后转移至冰上快速冷却, 室温保存。
ArumuganathanK, EarleED ( 1991). Estimation of nuclear DNA content of plants by flow cytometry Plant Mol Biol Rep 9, 229-241. [本文引用: 1]
CheL, TangD, WangK, WangM, ZhuK, YuH, GuM, ChengZ ( 2011). OsAM1 is required for leptotene transition in rice Cell Res 21, 654-665. [本文引用: 1]
ChengZ, BuellCR, WingRA, GuM, JiangJ ( 2001 a). Toward a cytological characterization of the rice genome Genome Res 11, 2133-2141.
ChengZ, DongF, LangdonT, OuyangS, BuellCR, GuM, BlattnerFR, JiangJ ( 2002). Functional rice centromeres are marked by a satellite repeat and a centromere-specific retrotransposon Plant Cell 14, 1691-1704. [本文引用: 2]
ChengZ, StuparRM, GuM, JiangJ ( 2001 b). A tandemly repeated DNA sequence is associated with both knob-like heterochromatin and a highly decondensed structure in the meiotic pachytene chromosomes of rice Chromosoma 110, 24-31. [本文引用: 1]
GoffSA ( 1999). Rice as a model for cereal genomics Curr Opin Plant Biol 2, 86-89. [本文引用: 1]
JiJH, TangD, ShenY, XueZH, WangHJ, ShiWQ, ZhangC, DuGJ, LiYF, ChengZK ( 2016). P31 comet, a member of the synaptonemal complex, participates in meiotic DSB formation in rice Proc Natl Acad Sci USA 113, 10577-10582. [本文引用: 1]
KurataN, OmuraT, IwataN ( 1981). Studies on centromere, chromomere and nucleolus in pachytene nuclei of rice, Oryza sativa, microsporocytes Cytologia 46, 791-800. [本文引用: 1]
LiX, ChaoD, WuY, HuangX, ChenK, CuiL, SuL, YeW, ChenH, ChenH, DongN, GuoT, ShiM, FengQ, ZhangP, HanB, ShanJ, GaoJ, LinH ( 2015). Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice Nat Genet 47, 827-833. [本文引用: 1]
LiYF, QinBX, ShenY, ZhangFF, LiuCZ, YouHL, DuGJ, TangD, ChengZK ( 2018). HEIP1 regulates crossover formation during meiosis in rice Proc Natl Acad Sci USA 115, 10810-10815. [本文引用: 1]
LuoQ, LiYF, ShenY, ChengZK ( 2014). Ten years of gene discovery for meiotic event control in rice J Genet Genomics 41, 125-137. [本文引用: 1]
MiaoCB, TangD, ZhangHG, WangM, LiYF, TangSZ, YuHX, GuMH, ChengZK ( 2013). CENTRAL REGION COMPONENT 1, a novel synaptonemal complex component, is essential for meiotic recombination initiation in rice Plant Cell 25, 2998-3009. [本文引用: 1]
NonomuraK, MorohoshiA, NakanoM, EiguchiM, MiyaoA, HirochikaH, KurataN ( 2007). A germ cell specific gene of the ARGONAUTE family is essential for the progression of premeiotic mitosis and meiosis during sporogenesis in rice Plant Cell 19, 2583-2594. [本文引用: 1]
PawlowskiWP, GolubovskayaIN, TimofejevaL, MeeleyRB, SheridanWF, CandeWZ ( 2004). Coordination of meiotic recombination, pairing, and synapsis by PHS1 Science 303, 89-92. [本文引用: 1]
TangX, BaoW, ZhangW, ChengZ ( 2007). Identification of chromosomes from multiple rice genomes using a universal molecular cytogenetic marker system J Integr Plant Biol 49, 953-960. [本文引用: 1]
WangK, TangD, WangM, LuJ, YuH, LiuJ, QianB, GongZ, WangX, ChenJ, GuM, ChengZ ( 2009). MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice J Cell Sci 122, 2055-2063. [本文引用: 1]
WangM, WangK, TangD, WeiC, LiM, ShenY, ChiZ, GuM, ChengZ ( 2010). The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice Plant Cell 22, 417-430. [本文引用: 1]
WangY, XiongG, HuJ, JiangL, YuH, XuJ, FangY, ZengL, XuE, XuJ, YeW, MengX, LiuR, ChenH, JingY, WangY, ZhuX, LiJ, QianQ ( 2015). Copy number variation at the GL7 locus contributes to grain size diversity in rice Nat Genet 47, 944-948. [本文引用: 1]
WuHK ( 1967). Note on preparing of pachytene chromosomes by double mordant Sci Agric 15, 40-44. [本文引用: 1]
YuH, WangM, TangD, WangK, ChenF, GongZ, GuM, ChengZ ( 2010). OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice Chromosoma 119, 625-636. [本文引用: 1]
ZhangF, TangD, ShenY, XueZH, ShiWQ, RenLJ, DuGJ, LiY, ChengZK ( 2017). The F-box protein ZYGO1 mediates bouquet formation to promote homologous pairing, synapsis, and recombination in rice meiosis Plant Cell 29, 2597-2609. [本文引用: 1]
ZhaoTT, RenLJ, ChenXJ, YuHX, LiuCJ, ShenY, ShiWQ, TangD, DuGJ, LiYF, MaBJ, ChengZK ( 2018). The OsRR24/LEPTO1 type-B response regulator is essential for the organization of leptotene chromosomes in rice meiosis Plant Cell 30, 3024-3037. [本文引用: 1]
Estimation of nuclear DNA content of plants by flow cytometry 1 1991
OsAM1 is required for leptotene transition in rice 1 2011
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Toward a cytological characterization of the rice genome 0 2001
Functional rice centromeres are marked by a satellite repeat and a centromere-specific retrotransposon 2 2002
... 水稻粗线期染色体染色的制备方法主要有2种类型: 一种是醋酸洋红染色法(Wu, 1967); 另一种是Giemsa染色法(Kurata et al., 1981).这2种方法都能获得较好的粗线期染色体图片, 并能很好地区分异染色质和常染色质, 对于不同染色体的鉴定有很大帮助.20世纪末, 随着分子生物学研究的发展, 将经典细胞遗传学与现代分子生物学相结合的荧光原位杂交技术, 堪称细胞遗传学研究方法的一场革命.这种显微技术的发展, 使得直接在染色体上观察DNA序列成为可能.随着水稻全基因组序列测定完成, 在水稻基因组中已鉴定出许多类型的串联重复序列, 包括45S rDNA、5S rDNA、CentO (着丝粒重复序列)和Os48 (亚端粒重复序列) (Cheng et al., 2001b, 2002).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
... ).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
A tandemly repeated DNA sequence is associated with both knob-like heterochromatin and a highly decondensed structure in the meiotic pachytene chromosomes of rice 1 2001
... 水稻粗线期染色体染色的制备方法主要有2种类型: 一种是醋酸洋红染色法(Wu, 1967); 另一种是Giemsa染色法(Kurata et al., 1981).这2种方法都能获得较好的粗线期染色体图片, 并能很好地区分异染色质和常染色质, 对于不同染色体的鉴定有很大帮助.20世纪末, 随着分子生物学研究的发展, 将经典细胞遗传学与现代分子生物学相结合的荧光原位杂交技术, 堪称细胞遗传学研究方法的一场革命.这种显微技术的发展, 使得直接在染色体上观察DNA序列成为可能.随着水稻全基因组序列测定完成, 在水稻基因组中已鉴定出许多类型的串联重复序列, 包括45S rDNA、5S rDNA、CentO (着丝粒重复序列)和Os48 (亚端粒重复序列) (Cheng et al., 2001b, 2002).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
P31 comet, a member of the synaptonemal complex, participates in meiotic DSB formation in rice 1 2016
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Studies on centromere, chromomere and nucleolus in pachytene nuclei of rice, Oryza sativa, microsporocytes 1 1981
... 水稻粗线期染色体染色的制备方法主要有2种类型: 一种是醋酸洋红染色法(Wu, 1967); 另一种是Giemsa染色法(Kurata et al., 1981).这2种方法都能获得较好的粗线期染色体图片, 并能很好地区分异染色质和常染色质, 对于不同染色体的鉴定有很大帮助.20世纪末, 随着分子生物学研究的发展, 将经典细胞遗传学与现代分子生物学相结合的荧光原位杂交技术, 堪称细胞遗传学研究方法的一场革命.这种显微技术的发展, 使得直接在染色体上观察DNA序列成为可能.随着水稻全基因组序列测定完成, 在水稻基因组中已鉴定出许多类型的串联重复序列, 包括45S rDNA、5S rDNA、CentO (着丝粒重复序列)和Os48 (亚端粒重复序列) (Cheng et al., 2001b, 2002).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice 1 2015
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
HEIP1 regulates crossover formation during meiosis in rice 1 2018
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Ten years of gene discovery for meiotic event control in rice 1 2014
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
COLD1 confers chilling tolerance in rice 1 2015
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
CENTRAL REGION COMPONENT 1, a novel synaptonemal complex component, is essential for meiotic recombination initiation in rice 1 2013
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
A germ cell specific gene of the ARGONAUTE family is essential for the progression of premeiotic mitosis and meiosis during sporogenesis in rice 1 2007
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Coordination of meiotic recombination, pairing, and synapsis by PHS1 1 2004
OsSPL regulates meiotic fate acquisition in rice 1 2018
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Identification of chromosomes from multiple rice genomes using a universal molecular cytogenetic marker system 1 2007
... 水稻粗线期染色体染色的制备方法主要有2种类型: 一种是醋酸洋红染色法(Wu, 1967); 另一种是Giemsa染色法(Kurata et al., 1981).这2种方法都能获得较好的粗线期染色体图片, 并能很好地区分异染色质和常染色质, 对于不同染色体的鉴定有很大帮助.20世纪末, 随着分子生物学研究的发展, 将经典细胞遗传学与现代分子生物学相结合的荧光原位杂交技术, 堪称细胞遗传学研究方法的一场革命.这种显微技术的发展, 使得直接在染色体上观察DNA序列成为可能.随着水稻全基因组序列测定完成, 在水稻基因组中已鉴定出许多类型的串联重复序列, 包括45S rDNA、5S rDNA、CentO (着丝粒重复序列)和Os48 (亚端粒重复序列) (Cheng et al., 2001b, 2002).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice 1 2009
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice 1 2010
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Copy number variation at the GL7 locus contributes to grain size diversity in rice 1 2015
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
Note on preparing of pachytene chromosomes by double mordant 1 1967
... 水稻粗线期染色体染色的制备方法主要有2种类型: 一种是醋酸洋红染色法(Wu, 1967); 另一种是Giemsa染色法(Kurata et al., 1981).这2种方法都能获得较好的粗线期染色体图片, 并能很好地区分异染色质和常染色质, 对于不同染色体的鉴定有很大帮助.20世纪末, 随着分子生物学研究的发展, 将经典细胞遗传学与现代分子生物学相结合的荧光原位杂交技术, 堪称细胞遗传学研究方法的一场革命.这种显微技术的发展, 使得直接在染色体上观察DNA序列成为可能.随着水稻全基因组序列测定完成, 在水稻基因组中已鉴定出许多类型的串联重复序列, 包括45S rDNA、5S rDNA、CentO (着丝粒重复序列)和Os48 (亚端粒重复序列) (Cheng et al., 2001b, 2002).由于这些串联重复序列可以在特定染色体上产生不同的荧光杂交信号, 为水稻染色体的识别带来了很大便利.此外, 利用水稻基因组DNA构建的细菌人工染色体(BACs), 通过筛选染色体或染色体臂特异BAC克隆, 建立了2套染色体特异的细胞遗传学标记(Cheng et al., 2001a; Tang et al., 2007), 从根本上解决了水稻染色体识别的难题. ...
OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice 1 2010
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
The F-box protein ZYGO1 mediates bouquet formation to promote homologous pairing, synapsis, and recombination in rice meiosis 1 2017
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
The OsRR24/LEPTO1 type-B response regulator is essential for the organization of leptotene chromosomes in rice meiosis 1 2018
... 自水稻基因组序列测序完成以来, 以解析不同基因功能的功能基因组学研究已全面展开.以水稻为研究对象, 众多与水稻形态、发育和代谢相关的基因已逐渐被克隆, 并完成对其生物学功能的解析(Li et al., 2015; Ma et al., 2015; Wang et al., 2015).同时, 利用水稻进行减数分裂调控分子机理研究也有很多优势.例如, 水稻是粮食作物中基因组最小的, 加之已建立其成熟的遗传转化体系、拥有相对完善的基因组信息以及适中的染色体大小和数目, 这些优势使得水稻成为继拟南芥(Arabidopsis thaliana)之后, 研究减数分裂调控分子机制的又一种模式植物.到目前为止, 已在水稻中克隆了40多个调控减数分裂过程的基因(Nonomura et al., 2007; Wang et al., 2009, 2010; Yu et al., 2010; Che et al., 2011; Miao et al., 2013; Luo et al., 2014; Ji et al., 2016; Zhang et al., 2017; Li et al., 2018; Ren et al., 2018; Zhao et al., 2018). ...
 ,2,*
,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}