马宏秀, 王开勇
, * , 张开祥, 孟春梅, 安梦洁石河子大学农学院, 石河子 832000
Effect of Cottonseed Meal on Cotton Physiology and Growth Compensation Under Salinity-alkalinity Stress Hongxiu Ma, Kaiyong Wang
, * , Kaixiang Zhang, Chunmei Meng, Mengjie AnAgricultural College, Shihezi University, Shihezi 832000, China
通讯作者: * E-mail: wky20@163.com 责任编辑: 白羽红
收稿日期: 2018-04-8
接受日期: 2018-08-23
网络出版日期: 2019-03-10
Corresponding authors: * E-mail: wky20@163.com Received: 2018-04-8
Accepted: 2018-08-23
Online: 2019-03-10
摘要 为探究棉粕对盐碱胁迫下棉花(Gossypium hirsutum )的抗盐碱机理, 通过田间试验, 研究添加不同棉粕用量对8 g·kg -1 盐碱胁迫下棉花生理及生长的补偿效应。结果表明, 添加棉粕能够增加盐碱胁迫下棉花不同器官对K + 的吸收, 降低对Na + 的吸收, 维持盐碱胁迫下细胞内K + 和Na + 的平衡, 显著促进棉花生长, 提高叶片叶绿素含量和光合作用效率, 有效缓解盐碱胁迫对棉花的伤害。其中, 以添加6 000 kg·hm -2 棉粕处理的效果最显著, 且盐胁迫下棉粕的改良效果较好。主成分分析结果表明, 盐碱胁迫下棉花生长生理的主要影响因子为叶片K + /Na + 比值、根长、鲜重、干重和胞间CO2 浓度(C i )。 关键词: 棉粕 ;
盐碱胁迫 ;
生理 ;
生长 ;
补偿效应 Abstract We investigated the salt-tolerant mechanism of cottonseed meal in cotton (Gossypium hirsutum ) under sali- nity-alkalinity stress. Salinity-alkalinity stress of 8 g·kg -1 was tested in the field to explore the effect of cottonseed meal dose on the physiology and growth of cotton. Cottonseed meal could increase the absorption of K + and decrease that of Na + . K + and Na + were kept in ion balance in cells under salinity-alkalinity stress. Cottonseed meal could effectively alleviate damage to cotton, significantly promote growth of cotton, and improve chlorophyll content and photosynthesis of leaves. An amount of 6 000 kg·hm -2 cottonseed meal was the most significant treatment, and the improvement effect of cottonseed meal under salt stress was better. According to principal component analysis, the K + /Na + ratio in leaves, root length, fresh weight, dry weight and intercellular CO2 concentration were the main factors involved in salinity-alkalinity stress. Keywords: cottonseed meal ;
salinity-alkalinity stress ;
physiology ;
growth ;
compensation effect PDF (5764KB) 摘要页面 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 马宏秀, 王开勇, 张开祥, 孟春梅, 安梦洁 . 棉粕对盐碱胁迫下棉花生理及生长补偿效应. 植物学报, 2019,
54 (2): 208-216 doi:10.11983/CBB18089
Ma Hongxiu, Wang Kaiyong, Zhang Kaixiang, Meng Chunmei, An Mengjie .
Effect of Cottonseed Meal on Cotton Physiology and Growth Compensation Under Salinity-alkalinity Stress .
Chinese Bulletin of Botany , 2019,
54 (2): 208-216 doi:10.11983/CBB18089
盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称。 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区。新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 )。目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 )。综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势。
新疆作为我国最大的商品棉生产基地, 棉籽原料十分丰富, 棉粕是棉籽榨油形成的副产物, 因此新疆是我国棉粕主产区之一。棉粕价格低廉, 主要用作动物饲料, 但棉粕中的棉酚具有一定的毒性, 极大降低了其饲用价值(
Jalees et al., 2011 ;
陈琪和石剑华, 2013 )。
张娜等(2009) 以棉粕为原料, 对提取的棉蛋 白进行理化性质分析, 发现棉蛋白的提取率可达75.5%。
刘玉国等(2013) 针对盐渍化土壤对几种常见土壤改良剂进行初次筛选, 并对改良后盐渍化土壤电导率和pH值进行测定, 发现用葵粕、棉粕、柠檬酸和沼液改良盐渍化土壤效果较好。
盐碱胁迫是两种不同性质的胁迫。盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 )。研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 )。碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 )。也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 )。将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 )。一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 )。研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 )。目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见。因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义。 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴。
1 材料与方法 1.1 试验条件 于2017年在新疆石河子大学试验站(44°18′42″N, 86°03′21″E)进行田间小区试验。土壤类型为壤土, 其基本理化性质为有机质含量16.19 g·kg
-1 , 土壤电导率(EC
1:5 ) 0.28 dS·m
-1 , 全氮0.5 g·kg
-1 , 速效磷20.40 mg·kg
-1 , 速效钾112 mg·kg
-1 , pH7.73。 棉花品种为早熟陆地棉品种新陆早60号(
Gossypium hirsutum cv. ‘Xinluzao 60’), 在全疆各地广泛种植, 由新疆农垦科学院棉花研究所培育而成。试验田所用肥料为棉粕和复合肥(用高塔硝硫基肥与尿素配制, N:P:K=20: 15:9, v/v/v)。
1.2 试验设计 2016年9月, 采集混合土样测定实验地土壤基本理化性质, 并通过撒施中性盐(NaCl)和碱性盐(Na
2 CO
3 )的方式对试验小区预先进行盐化及碱化处理, 使土壤盐碱含量达到8 g·kg
-1 , 处理后盐化土壤的pH值为8.43, EC
1:5 为1.94 dS·m
-1 ; 碱化土壤pH值为9.89, EC
1:5 为1.03 dS·m
-1 。 随后将预定量的棉粕均匀地撒入不同小区, 并进行0-20 cm浅翻处理, 使其充分混合。于2017年4月播种, 9月收获。 小区面积6 m
2 , 小区间距1.5 m, 并在小区之间设置约50 cm深的双层塑料隔膜。 所有小区均采用地膜滴灌栽培方式, 一膜三管六行, 宽窄行处理, 宽行距60 cm, 窄行距20 cm。一次性施入基肥: 氮300 kg·hm
-2 , 五氧化二磷237 kg·hm
-2 , 氧化钾123 kg·hm
-2 。 全生育期滴灌量4 200 m
3 ·hm
-2 。 其它管理措施同当地大田常规管理。
试验共设7个处理: NaCl+0 kg·hm
-2 棉粕(Y
0 )、NaCl+3 000 kg·hm
-2 棉粕(Y
1 )、NaCl+6 000 kg·hm
-2 棉粕(Y
2 )、Na
2 CO
3 +0 kg·hm
-2 棉粕(J
0 )、Na
2 CO
3 + 3 000 kg·hm
-2 棉粕(J
1 )、Na
2 CO
3 +6 000 kg·hm
-2 棉粕(J
2 )及未进行盐化处理和未撒施棉粕的空白对照(CK)。每个处理设3次重复。
1.3 样品采集与光合参数测定 在棉花花铃期进行样品采集。每小区随机选取5株, 采用常规方法测定株高、根长、鲜重和干重(地上部和地下部)。叶绿素提取采用乙醇提取法, 紫外分光光度法测定其含量(
李合生和陈华癸, 2000 )。分别将烘干的根、茎和叶粉碎过筛, 准确称取200 mg粉末样品, 利用硝酸-双氧水消解, 采用火焰光度计法(
鲍士旦, 2000 )测定Na
+ 和K
+ 含量。光合参数采用便携式光合仪(CARIS-2, 英国PP Systems公司)于中午12:00- 14:30取棉花叶片倒4叶进行测定。测定过程中光强为400 μmol·m
-2 ·s
-1 , 大气温度(25±1)°C, 大气CO
2 浓度(
C a )变化范围为(400±10) μmol·L
-1 。测定指标包括净光合速率(
P n )、胞间CO
2 浓度(
C i )、蒸腾速率(
E )和气孔导度(
G s )。气孔限制值计算公式为
L s =1-
C i /
C a (
C a 为空气中CO
2 浓度)。每个处理重复测定5次, 取平均值。
1.4 主成分分析 为了分析不同处理对棉花生长的影响, 利用DPS V9.01软件进行各因子主成分分析(principal components analysis, PCA), 同时做因子载荷矩阵。
1.5 数据处理 采用Microsoft Excel 2010和DPS V9.01软件处理数据并绘制图表。利用SPSS 19.0软件对数据进行统计分析。差异显著性检验采用Turkey检验法进行多重比较。
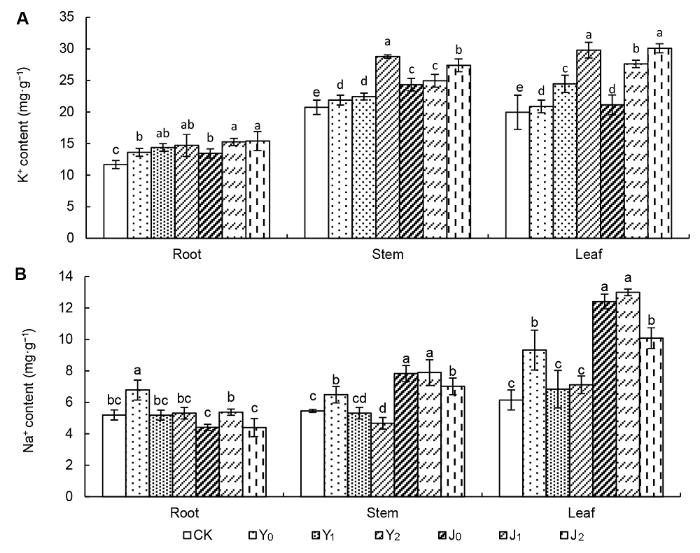
2 结果与讨论 2.1 棉粕对盐碱胁迫下棉花不同器官K+ 、Na+ 累积效应的影响 结果(
图1A )表明, 与对照相比, 不同处理棉花根、茎、叶K
+ 含量均有所增加, 盐碱胁迫下茎和叶中K
+ 累积显著高于根部。 随着棉粕施用量的增加, 盐碱胁迫下棉花的K
+ 含量呈现逐渐增加的趋势。Y
1 处理与Y
0 处理相比差异不显著, Y
2 处理的根、茎、叶中K
+ 含量分别增加了8.09%、31.55%和42.81%。与J
0 处理相比, J
2 处理的根、茎、叶中K
+ 含量分别增加了14.51%、12.54%和42.26%。上述结果表明, 在盐胁迫下, 添加棉粕后棉花不同器官中K
+ 含量变化大小为叶>茎>根; 在碱胁迫下, 添加棉粕后棉花不同器官中K
+ 含量变化大小为叶>根>茎。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1棉粕对盐碱胁迫下棉花不同器官K+ (A)、Na+ (B)含量的影响 CK: 对照; Y
0 : NaCl+0 kg·hm
-2 棉粕; Y
1 : NaCl+3 000 kg·hm
-2 棉粕; Y
2 : NaCl+6 000 kg·hm
-2 棉粕; J
0 : Na
2 CO
3 +0 kg·hm
-2 棉粕; J
1 : Na
2 CO
3 +3 000 kg·hm
-2 棉粕; J
2 : Na
2 CO
3 +6 000 kg·hm
-2 棉粕。不同小写字母表示各处理差异显著(
P ?0.05)。
Figure 1Effect of cottonseed meal on K+ (A) and Na+ (B) content of cotton under salinity-alkalinity stress CK: Control; Y
0 : NaCl+0 kg·hm
-2 cottonseed meal; Y
1 : NaCl+3 000 kg·hm
-2 cottonseed meal; Y
2 : NaCl+6 000 kg·hm
-2 cottonseed meal; J
0 : Na
2 CO
3 +0 kg·hm
-2 cottonseed meal; J
1 : Na
2 CO
3 +3 000 kg·hm
-2 cottonseed meal; J
2 : Na
2 CO
3 +6 000 kg·hm
-2 cottonseed meal. Different lowercase letters indicate significant differences among each treatment (
P ?0.05).
与对照相比, 不同处理棉花根、茎、叶中Na
+ 含量变化规律不同, 盐碱胁迫下茎和叶中Na
+ 累积显著高于根部(
图1B )。与Y
0 处理相比, Y
1 和Y
2 处理棉花根、茎、叶中Na
+ 含量显著降低, 但Y
1 和Y
2 两者差异不显著, 根、茎、叶中Na
+ 含量分别降低了21.85%和23.64%、18.24%和28.01%、23.61%和26.57%。与J
0 处理相比, J
1 处理与其差异不显著, J
2 处理棉花各器官Na
+ 含量显著降低, 根、茎、叶Na
+ 含量分别降低了0.30%、10.34%和18.73%。以上结果表明, 在盐胁迫下, 添加3 000 kg·hm
-2 棉粕后棉花不同器官中Na
+ 含量变化大小依次为叶>茎>根, 添加6 000 kg·hm
-2 棉粕后棉花不同器官中Na
+ 含量变化大小为茎>叶>根; 在碱胁迫下, 添加6 000 kg·hm
-2 棉粕后棉花不同器官中Na
+ 含量变化大小为叶>茎>根。
2.2 棉粕对盐碱胁迫下棉花不同器官K+ /Na+ 比值的影响 与对照相比, 盐胁迫下, 除Y
0 处理外, Y
1 和Y
2 处理棉花不同器官K
+ /Na
+ 比值均有所增加; 碱胁迫下, J
0 、J
1 和J
2 处理棉花根部K
+ /Na
+ 比值均有所增加, 茎部和叶部K
+ /Na
+ 比值均有不同程度降低(
图2 )。盐胁迫下, 茎部和叶部以Y
2 处理K
+ /Na
+ 比值最高, 根部以J
2 处理K
+ /Na
+ 比值最高。与Y
0 处理相比, 随着棉粕施用量的增加, 各处理棉花不同器官K
+ /Na
+ 比值也相应增加。与Y
0 处理相比, Y
2 处理根、茎、叶中K
+ /Na
+ 比值分别增加了38.03%、82.81%和86.88%。碱胁迫下,与J
0 处理相比, J
1 和J
2 处理棉花不同器官K
+ /Na
+ 比值均有所增加, 其中J
2 处理根、茎、叶中K
+ /Na
+ 比值分别增加了14.77%、25.41%和75.47%。以上结果表明, 在盐碱胁迫下, 添加棉粕后棉花不同器官中K
+ /Na
+ 比值变化大小为叶>茎>根。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2棉粕对盐碱胁迫下棉花不同器官K+ /Na+ 比值的影响 CK、Y
0 、Y
1 、Y
2 、J
0 、J
1 和J
2 同图1。不同小写字母表示各处理差异显著(P<0.05)。
Figure 2Effect of cottonseed meal on K+ /Na+ ratio of cotton under salinity-alkalinity stress CK、Y
0 、Y
1 、Y
2 、J
0 、J
1 and J
2 see Figure 1. Different lowercase letters indicate significant differences among each treatment (P<0.05).
2.3 棉粕对盐碱胁迫下棉花生理指标的影响 与对照相比, 不同处理棉花叶片叶绿素含量均有所降低(
表1 )。盐胁迫下, 与Y
0 处理相比, 随着棉粕施用量的增加, 棉花叶绿素含量也相应增加。Y
2 处理叶绿素总含量、叶绿素a、叶绿素b及类胡萝卜素含量分别增加了37.09%、23.08%、36.36%和67.65%。碱胁迫下, 与J
0 处理相比, 随着棉粕施用量的增加, 棉花叶绿素含量呈现不同的变化趋势, 其中, J
2 处理叶绿素a和类胡萝卜素含量分别增加了22.22%和82.14%, 而叶绿素总含量和叶绿素b含量分别降低了13.30%和4.76%。
Table 1 表1 表1 棉粕对盐碱胁迫下棉花叶片叶绿素含量的影响
Table 1
Effect of cottonseed meal on chlorophyll content of cotton leaves under salinity-alkalinity stress Treatment Total chlorophyll content-1 ) Chlorophyll a content-1 ) Chlorophyll b content-1 ) Carotenoid content-1 ) CK 2.51±0.10 a 1.45±0.08 a 0.66±0.06 a 0.64±0.09 a Y0 1.51±0.08 c 1.17±0.10 bc 0.33±0.13 b 0.34±0.09 b Y1 1.80±0.08 bc 1.37±0.08 bc 0.43±0.09 ab 0.46±0.10 ab Y2 2.07±0.13 ab 1.44±0.09 ab 0.45±0.10 ab 0.57±0.13 ab J0 2.18±0.19 ab 1.17±0.13 c 0.42±0.13 ab 0.28±0.08 b J1 1.78±0.28 bc 1.37±0.13 ab 0.33±0.10 b 0.32±0.09 b J2 1.89±0.12 bc 1.43±0.08 ab 0.40±0.11 ab 0.51±0.16 ab
CK、Y0 、Y1 、Y2 、J0 、J1 和J2 同图1。同列不同小写字母表示各处理差异显著(P<0.05)。 CK、Y0 、Y1 、Y2 、J0 、J1 and J2 see Figure 1. Different lowercase letters in the same column indicate that each treatment is significantly different (P<0.05). 新窗口打开 |
下载CSV 与对照相比, Y
0 和J
0 处理的棉花叶片净光合速率(
P n )、气孔导度(
G s )和胞间CO
2 浓度(
C i )均有所降低(
表2 ), 表明盐碱胁迫造成棉花叶片
P n 下降的主要因素是气孔因素。与Y
0 处理相比, Y
1 和Y
2 处理的
P n 分别增加了3.96%和4.91%;
G s 分别增加了8.25%和19.21%; 蒸腾速率(
E )分别增加了4.37%和10.00%;
C i 分别增加了2.74%和7.62%。与J
0 处理相比, J
1 和J
2 处理的
P n 分别增加了11.63%和12.79%;
G s 分别增加了14.13%和14.45%;
E 分别增加了3.16%和5.06%;
C i 分别增加了2.80%和7.05%。以上结果表明, 在盐碱胁迫下, 添加棉粕后棉花叶片
P n 、
G s 、
E 和
C i 均有不同程度的增加, 其中, 碱胁迫下添加棉粕后棉花叶片的
P n 增幅大于盐胁迫。
Table 2 表2 表2 棉粕对盐碱胁迫下棉花光合参数的影响
Table 2
Effect of cottonseed meal on photosynthetic parameters of cotton under salinity-alkalinity stress Treatment P n (μmol·m-2 ·s-1 )G s (mol·m-2 ·s-1 )E (mmol·m-2 ·s-1 )C i (μmol·mol-1 )L s CK 6.73±0.15 a 111.13±1.28 a 1.81±0.25 a 281.20±0.10 b 0.25±0.10 a Y0 6.32±0.38 b 79.17±0.70 d 1.60±1.95 ab 262.79±0.07 e 0.28±0.04 a Y1 6.57±0.15 a 85.7±0.49 c 1.67±1.10 ab 270.00±0.06 d 0.25±0.06 a Y2 6.63±0.27 a 94.38±1.81 b 1.76±1.86 ab 282.81±0.04 b 0.21±0.10 a J0 5.16±0.07 c 75.16±1.63 e 1.58±1.60 b 274.77±0.08 c 0.26±0.05 a J1 5.76±0.18 bc 85.78±1.06 c 1.63±1.54 ab 282.47±0.06 b 0.21±0.10 a J2 5.82±0.25 b 86.02±1.51 c 1.66±1.88 ab 294.13±0.10 a 0.21±0.09 a
Pn : 净光合速率; Gs : 气孔导度; E: 蒸腾速率; Ci : 胞间CO2 浓度; Ls : 气孔限制值。CK、Y0 、Y1 、Y2 、J0 、J1 和 J2 同图1。同列不同小写字母表示各处理差异显著(P<0.05)。 Pn : Net photosynthetic rate; Gs : Stomatal conductance; E: Transpiration rate; Ci : Intercellular CO2 concentration; Ls : Stomatal limit value. CK、Y0 、Y1 、Y2 、J0 、J1 and J2 see Figure 1. Different lowercase letters in the same column indicate significant differences among each treatment(P<0.05). 新窗口打开 |
下载CSV 2.4 棉粕对盐碱胁迫下棉花生长的影响 与对照相比, 各处理棉花的根长、鲜重及干重均有所降低(
表3 )。与Y
0 处理相比, Y
1 和Y
2 处理植株株高分别增加了8.20%和17.32%; 根长分别增加了14.29%和18.66%; 鲜重分别增加了27.47%和33.59%; 干重分别增加了16.68%和45.55%。与J
0 处理相比, J
1 和J
2 处理植株株高分别增加了2.38%和9.51%; 根长分别增加了11.50%和15.89%; 鲜重分别增加了9.34%和11.21%; 干重分别增加了15.49%和35.47%。以上结果表明, 在盐碱胁迫下, 添加棉粕后棉花的株高、根长、鲜重及干重均有不同程度的增加, 其中, 盐胁迫下添加棉粕后棉花的株高、根长、鲜重及干重增加幅度均大于碱胁迫。
Table 3 表3 表3 棉粕对盐碱胁迫下棉花生长的影响
Table 3
Effect of cottonseed meal on cotton growth under salinity-alkalinity stress Treatment Plant height (cm) Root length (cm) Fresh weight (g·plant -1 ) Dry weight (g·plant -1 ) CK 89.4±0.36 b 31.83±0.78 a 184.59±0.23 a 54.49±0.26 a Y0 84.1±0.17 c 26.1±0.61 c 135.81±0.52 g 37.89±0.46 e Y1 91±0.58 b 29.83±0.35 b 173.12±0.88 c 44.21±0.18 c Y2 98.67±0.58 a 30.97±0.40 ab 181.43±0.88 b 55.15±0.34 a J0 74.33±0.58 d 22.78±0.40 d 148.47±0.43 f 34.42±0.34 f J1 76.33±1.15 d 25.4±0.26 c 162.33±0.96 e 39.75±0.27 d J2 82.33±0.58 c 26.4±0.17 c 165.12±0.42 d 46.63±0.71 b
CK、Y0 、Y1 、Y2 、J0 、J1 和J2 同图1。同列不同小写字母表示各处理差异显著(P<0.05)。 CK、Y0 、Y1 、Y2 、J0 、J1 and J2 see Figure 1. Different lowercase letters in the same column indicate that each treatment is significantly different (P<0.05). 新窗口打开 |
下载CSV 2.5 各指标主成分分析 对标准化后各处理的22个指标进行分析, 转换为新的综合指标, 能代替22个单项指标的绝大部分信息且贡献率最大定义为第1主成分(PC1), 其贡献率为46.85%。此外, 能代替单项指标的综合指标定义为第2主成分(PC2), 其贡献率为21.71%。对代表样点理化特性的变量进行排序作图(
图3 ), 同时计算各理化因子载荷(
表4 )。
Table 4 表4 表4 棉花盐碱胁迫下不同处理因子载荷
Table 4
Load of different treatment factors of cotton under salinity-alkalinity stress Index Salt treatment Alkali treatment PC1 PC2 PC1 PC2 K+ Root -0.05 0.94** -0.67 0.69 Stem 0.47 0.78* -0.68 0.70 Leaf 0.73 0.60 -0.09 0.96** Na+ Root -0.83* 0.01 0.41 -0.15 Stem -0.87* -0.36 -0.98** 0.02 Leaf -0.81* 0.23 -0.95** 0.00 K+ /Na+ ratio Root 0.61 0.61 -0.67 0.59 Stem 0.69 0.66 0.72 0.58 Leaf 0.96** 0.15 0.93** 0.28 Physiology index Total chlorophyll content 0.79* -0.52 0.63 -0.55 Chlorophyll a content 0.82* -0.07 0.60 0.55 Chlorophyll b content 0.56 -0.68 0.78* -0.35 Carotenoid content 0.78* -0.29 0.83* 0.19 P n 0.61 -0.16 0.94** 0.10 G s 0.75* -0.61 0.98** -0.03 C i 0.94** -0.11 0.16 0.97** E 0.74 -0.36 0.78* 0.06 L s -0.36 -0.12 0.02 -0.27 Growth index Plant height 0.82* 0.52 0.97** 0.17 Root length 0.96** -0.10 0.97** 0.11 Fresh weight 0.97** -0.11 0.97** 0.16 Dry weight 0.96** -0.13 0.95** 0.28
Pn 、Gs 、 Ci 、E和 Ls 同表2。 * 因子与主成分相关性显著(P<0.05)。** 因子与主成分相关性极显著(P<0.01)。 Pn 、Gs 、 Ci 、E和 Ls see Table 2; * Significant correlation(P<0.05)。** Extremely significant correlation(P<0.01)。 新窗口打开 |
下载CSV 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3棉花盐(A)和碱(B)胁迫下各指标主成分(PC)分析 CK、Y
0 、Y
1 、Y
2 、J
0 、J
1 和 J
2 同图1。
Figure 3Analysis of principal components (PC) of each index of cotton under salinity (A) and alkalinity (B) stress CK、Y
0 、Y
1 、Y
2 、J
0 、J
1 and J
2 see Figure 1.
PC1主轴正方向为Y
0 、Y
1 和Y
2 ; PC2主轴正方向为CK和Y
2 (
图3A )。由
图3B 可知, PC1主轴正方向为J
1 和J
2 ; PC2主轴正方向为CK。主成分相关性分析表明(
表4 ), 盐胁迫下, 第1主成分PC1与叶中K
+ /Na
+ 比值、
C i 、根长、鲜重、干重呈极显著正相关, 与叶绿素总含量、叶绿素a含量、类胡萝卜素含量、
G s 、株高呈显著正相关, 与根、茎、叶中Na
+ 含量呈显著负相关; 第2主成分PC2与根中K
+ 含量呈极显著正相关, 与茎中K
+ 含量呈显著正相关。以上结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 尤其是对叶中K
+ /Na
+ 比值、
C i 、根长、鲜重及干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感。
碱胁迫下, 第1主成分PC1与叶中K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重呈极显著正相关, 与叶绿素b含量、类胡萝卜素含量、
E 呈显著正相关, 与茎、叶中Na
+ 含量呈极显著负相关; 第2主成分PC2与叶中K
+ 含量、
C i 呈极显著正相关。这说明碱胁迫下添加棉粕对棉花生长生理指标影响较大, 尤其是对叶中K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感。
综上所述, 盐碱胁迫下棉花生长生理补偿机制的影响因子有所不同。盐胁迫下, 气孔关闭, 阻碍CO
2 进入细胞内部, 因此
C i 成为主要影响因子; 盐碱胁迫均会导致棉花叶片
P n 和
G s 下降, 但在碱胁迫下, 棉花叶片
P n 和
G s 值极低, 说明叶片的光合作用发生适应性下调。由于
P n 逐渐增大, 消耗大量CO
2 , 同时
G s 逐渐减小, 影响外界CO
2 向细胞内部扩散, 从而导致
C i 下降。通常植株地上部受害情况比地下部表现更为明显, 碱胁迫较盐胁迫对植物造成的伤害更大(
Zhang and Mu, 2009 ;
焦伟红, 2011 ), 因此
P n 、
G s 及株高成为主要影响因子。综上, 盐碱胁迫下棉花生长生理的主要影响因子有叶中K
+ /Na
+ 比值、根长、鲜重、干重及胞间CO
2 浓度(
C i )。
2.6 讨论 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 )。研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 )。保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 )。本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 )。叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 )。植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 )。本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 )。这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 )。本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 )。其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 )。植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 )。本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 )。王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加。本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用。主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 )。
综上, 本研究得到以下结论。(1) 添加棉粕能够增加盐碱胁迫下棉花各器官对K
+ 的吸收, 降低对Na
+ 的吸收, 以维持细胞内K
+ 和Na
+ 的平衡; (2) 添加棉粕能够缓解盐碱胁迫对棉花的伤害, 可促进棉花生长, 提高叶片叶绿素含量和光合作用效率; (3) 盐碱胁迫条件下添加6 000 kg·hm
-2 棉粕处理的改良效果最显著, 同时在盐胁迫下的作用优于碱胁迫。主成分分析表明, 盐碱胁迫下棉花生长生理的主要影响因子有叶中K
+ /Na
+ 比值、根长、鲜重、干重及胞间CO
2 浓度(
C i )。
The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。 [1] 阿曼古丽·买买提阿力 , 拉扎提·努尔布拉提 , 高丽丽 , 张巨松 , 田立文 ( 2017 ). 盐胁迫对海岛棉和陆地棉幼苗生长及生理特性的影响植物学报 52, 465 -473 . [本文引用: 1] [2] 鲍士旦 ( 2000 ). 土壤农化分析(第3版) . 北京: 中国农业出版社. pp.270 -271 . [本文引用: 1] [3] 陈俊 ( 2006 ). 碱地肤幼苗抗氧化酶系统对盐碱混合胁迫的生理响应特点长春: 东北师范大学 . pp.4 -10 . [本文引用: 1] [4] 陈琪 , 石剑华 ( 2013 ). 棉籽饼使用脱毒剂和生物发酵脱毒的营养分析当代畜禽养殖业 ( 10), 6 -7 ,8. [本文引用: 1] [5] 郭凯 , 巨兆强 , 封晓辉 , 李晓光 , 刘小京 ( 2016 ). 咸水结冰灌溉改良盐碱地的研究进展及展望中国生态农业学报 24, 1016 -1024 . [本文引用: 1] [6] 黄清荣 , 祁琳 , 柏新富 ( 2018 ). 根环境供氧状况对盐胁迫下棉花幼苗光合及离子吸收的影响生态学报 38, 528 -536 . [本文引用: 1] [7] 贾玉珍 , 朱禧月 , 唐予迪 , 蔡养廉 , 林同保 , 罗先宝 , 杨兆庚 , 韩海江 ( 1987 ). 棉花出苗及苗期耐盐性指标的研究河南农业大学学报 21, 30 -41 . [本文引用: 1] [8] 焦伟红 ( 2011 ). 燕麦耐盐碱渗透调节机制研究呼和浩特: 内蒙古农业大学 . pp.15 -20 . [本文引用: 1] [9] 李合生 , 陈华癸 ( 2000 ). 21世纪农林本科生物系列课程改革的研究与实践(上)中国农业教育 ( 4 ), 19 -22 . [本文引用: 1] [10] 蔺吉祥 , 李晓宇 , 唐佳红 , 张兆军 , 李卓琳 , 高战武 , 穆春生 ( 2011 ). 盐碱胁迫对小麦种子萌发、早期幼苗生长及Na + 、K + 代谢的影响 麦类作物学报 31, 1148 -1152 . [本文引用: 1] [11] 刘玉国 , 谭兰兰 , 卞龙 , 王开勇 ( 2013 ). 盐渍化土壤改良剂的筛选农业科技与信息 (20), 48 -49 , 50 . [本文引用: 1] [12] 牛花朋 , 李胜荣 , 申俊峰 , 李祯 , 佟景贵 ( 2006 ). 粉煤灰与若干有机固体废弃物配施改良土壤的研究进展地球与环境 34(2 ), 27 -34 . [本文引用: 1] [13] 石德成 , 殷丽娟 ( 1993 ). 盐(NaCl)与碱(Na2 CO3 )对星星草胁迫作用的差异植物学报 35, 144 -149 . [本文引用: 1] [14] 田长彦 , 周宏飞 , 刘国庆 ( 2000 ). 21世纪新疆土壤盐渍化调控与农业持续发展研究建议干旱区地理 23, 177 -181 . [本文引用: 1] [15] 王安平 , 吕云峰 , 张军民 , 赵青余 , 王加启 , 田科雄 ( 2010 ). 我国棉粕和棉籽蛋白营养成分和棉酚含量调研华北农学报 25(S1 ), 301 -304 . DOI:
10.7668/hbnxb.2010.S1.068 Magsci 本试验旨在调查研究我国不同产地的棉粕和棉籽蛋白的营养成分和棉酚含量。采集全国不同主产区的5个棉粕样品和5个棉籽蛋白样品测定其常规营养成分和棉酚含量。结果表明,棉粕的粗蛋白、粗脂肪、酸性洗涤纤维营养成分含量分别为39.28%,0.28%,21.60%,棉籽蛋白的粗蛋白、粗脂肪、酸性洗涤纤维营养成分含量分别为51.96%,0.75%,13.29%,其中,在棉粕和棉籽蛋白所有营养成分中粗脂肪和钙两项变异系数最大,粗蛋白变异系数最小。棉粕和棉籽蛋白的游离棉酚含量分别为1021.14mg/kg和687.75mg/kg。不同主产区棉粕和棉籽蛋白的粗蛋白含量稳定,而粗脂肪和钙含量差异较大;棉粕游离棉酚含量较高。
[16] 王文杰 , 贺海升 , 祖元刚 , 赵修华 , 杨磊 , 关宇 , 许慧男 , 于兴洋 ( 2009 ). 施加改良剂对重度盐碱地盐碱动态及杨树生长的影响生态学报 29, 2272 -2278 . [本文引用: 1] [17] 韦本辉 , 申章佑 , 周佳 , 甘秀芹 , 劳承英 , 周灵芝 , 刘斌 , 胡泊 , 李艳英 ( 2017 ). 粉垄改造利用盐碱地效果初探中国农业科技导报 19(10 ), 107 -112 . [本文引用: 1] [18] 翁永玲 , 宫鹏 ( 2006 ). 土壤盐渍化遥感应用研究进展地理科学 26, 369 -375 . [本文引用: 1] [19] 杨国会 , 石德成 ( 2011 ). 盐碱胁迫对小冰麦相对生长率及茎叶离子积累的影响河南农业科学 40, 45 -47, 57 . [本文引用: 1] [20] 杨淑萍 , 危常州 , 梁永超 ( 2010 ). 盐胁迫对不同基因型海岛棉光合作用及荧光特性的影响中国农业科学 43, 1585 -1593 . [本文引用: 1] [21] 员学锋 , 汪有科 , 吴普特 , 冯浩 ( 2005 ). PAM对土壤物理性状影响的试验研究及机理分析水土保持学报 19(2 ), 37 -40 . [本文引用: 1] [22] 张华宁 , 李孟军 , 郭秀林 , 张艳敏 , 刘子会 ( 2017 ). 盐胁迫下不同K+ 吸收抑制剂对小麦根系K+ /Na+ 比和质膜相关蛋白活性的影响 华北农学报 32(5 ), 154 -162 . [本文引用: 1] [23] 张景云 , 缪南生 , 白雅梅 , 万新建 , 吕文河 ( 2014 ). 盐胁迫下二倍体马铃薯叶绿素含量和抗氧化酶活性的变化作物杂志 ( 5 ), 59 -63 . [本文引用: 1] [24] 张娜 , 潘思轶 , 侯旭杰 ( 2009 ). 棉籽蛋白提取工艺及其主要理化性质研究食品研究与开发 30(7 ), 36 -38 . [本文引用: 1] [25] 张树文 , 杨久春 , 李颖 , 张养贞 , 常丽萍 ( 2010 ). 1950s中期以来东北地区盐碱地时空变化及成因分析自然资源学报 25, 435 -442 . [本文引用: 1] [26] 赵可夫 , 王韶唐 ( 1990 ). 作物抗性生理 . 北京: 农业出版社. pp. 300 -304 . [本文引用: 1] [27] 周志林 , 唐君 , 曹清河 , 赵冬兰 , 张安 ( 2017 ). NaCl胁迫对甘薯植株体内K+ 、Na+ 和Cl- 含量及生长的影响 中国农业科技导报 19(4 ), 17 -23 . [本文引用: 1] [28] 罗宾BA ( 陈恺元等译) ( 1983 ). 棉花生理学 . 上海: 上海科学技术出版社. pp. 116 -118 . [本文引用: 1] [29] Boscaiu M Estrelles E Soriano P Vicente O 2005 ). Effects of salt stress on the reproductive biology of the halophyte Plantago crassifolia Biol Plant 141 -143 . [本文引用: 1] [30] Jalees MM Khan MZ Saleemi MK Khan A 2011 ). Effects of cottonseed meal on hematological, biochemical and behavioral alterations in male Japanese quail (Coturnix japonica )Pak Vet J 211 -214 . [本文引用: 1] [31] Li RL Shi FC Fukuda K Yang YL 2010 ). Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa ( Medicago sativa L.)Soil Sci Plant Nutr 725 -733 . [本文引用: 1] [32] Munns R 2002 ). Comparative physiology of salt and water stressPlant Cell Environ 239 -250 . DOI:
10.1046/j.0016-8025.2001.00808.x URL [本文引用: 1] [33] Nedjimi B Daoud Y 2009 ). Ameliorative effect of CaCl2 on growth, membrane permeability and nutrient uptake in Atriplex halimus subspschweinfurthii grown at high (NaCl) salinity. Desalination 163 -166 . [本文引用: 1] [34] Schroeder JI Ward JM Gassmann W 1994 ). Perspectives on the physiology and structure of inward-rectifying K+ channels in higher plants: biophysical implications for K+ uptake Annu Rev Biophys Biomol Struct 441 -471 . DOI:
10.1146/annurev.bb.23.060194.002301 URL [本文引用: 1] [35] Štefanić PP Koffler T Adler G Bar-Zvi D 2013 ). Chloroplasts of salt-grown Arabidopsis seedlings are impaired in structure, genome copy number and transcript levelsPLoS One e82548 . DOI:
10.1371/journal.pone.0082548 URL [本文引用: 1] [36] Wang LW Showalter AM Ungar IA 2005 ). Effects of intraspecific competition on growth and photosynthesis of Atriplex prostrata Aquat Bot 187 -192 . [本文引用: 1] [37] Zhang JT Mu CS 2009 ). Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius Soil Sci Plant Nutr 685 -697 . [本文引用: 1] [38] Zhang TB Zhan XY Kang YH Wan SQ Feng H 2017 ). Improvements of soil salt characteristics and nutrient status in an impermeable saline-sodic soil reclaimed with an improved drip irrigation while ridge planting Lycium barbarum LJ Soils Sediments 1126 -1139 . [本文引用: 1] 盐胁迫对海岛棉和陆地棉幼苗生长及生理特性的影响
1
2017
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
1
2000
... 在棉花花铃期进行样品采集.每小区随机选取5株, 采用常规方法测定株高、根长、鲜重和干重(地上部和地下部).叶绿素提取采用乙醇提取法, 紫外分光光度法测定其含量(
李合生和陈华癸, 2000 ).分别将烘干的根、茎和叶粉碎过筛, 准确称取200 mg粉末样品, 利用硝酸-双氧水消解, 采用火焰光度计法(
鲍士旦, 2000 )测定Na
+ 和K
+ 含量.光合参数采用便携式光合仪(CARIS-2, 英国PP Systems公司)于中午12:00- 14:30取棉花叶片倒4叶进行测定.测定过程中光强为400 μmol·m
-2 ·s
-1 , 大气温度(25±1)°C, 大气CO
2 浓度(
C a )变化范围为(400±10) μmol·L
-1 .测定指标包括净光合速率(
P n )、胞间CO
2 浓度(
C i )、蒸腾速率(
E )和气孔导度(
G s ).气孔限制值计算公式为
L s =1-
C i /
C a (
C a 为空气中CO
2 浓度).每个处理重复测定5次, 取平均值. ...
碱地肤幼苗抗氧化酶系统对盐碱混合胁迫的生理响应特点
1
2006
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
棉籽饼使用脱毒剂和生物发酵脱毒的营养分析
1
2013
... 新疆作为我国最大的商品棉生产基地, 棉籽原料十分丰富, 棉粕是棉籽榨油形成的副产物, 因此新疆是我国棉粕主产区之一.棉粕价格低廉, 主要用作动物饲料, 但棉粕中的棉酚具有一定的毒性, 极大降低了其饲用价值(
Jalees et al., 2011 ;
陈琪和石剑华, 2013 ).
张娜等(2009) 以棉粕为原料, 对提取的棉蛋 白进行理化性质分析, 发现棉蛋白的提取率可达75.5%.
刘玉国等(2013) 针对盐渍化土壤对几种常见土壤改良剂进行初次筛选, 并对改良后盐渍化土壤电导率和pH值进行测定, 发现用葵粕、棉粕、柠檬酸和沼液改良盐渍化土壤效果较好. ...
咸水结冰灌溉改良盐碱地的研究进展及展望
1
2016
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
根环境供氧状况对盐胁迫下棉花幼苗光合及离子吸收的影响
1
2018
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
棉花出苗及苗期耐盐性指标的研究
1
1987
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
燕麦耐盐碱渗透调节机制研究
1
2011
... 综上所述, 盐碱胁迫下棉花生长生理补偿机制的影响因子有所不同.盐胁迫下, 气孔关闭, 阻碍CO
2 进入细胞内部, 因此
C i 成为主要影响因子; 盐碱胁迫均会导致棉花叶片
P n 和
G s 下降, 但在碱胁迫下, 棉花叶片
P n 和
G s 值极低, 说明叶片的光合作用发生适应性下调.由于
P n 逐渐增大, 消耗大量CO
2 , 同时
G s 逐渐减小, 影响外界CO
2 向细胞内部扩散, 从而导致
C i 下降.通常植株地上部受害情况比地下部表现更为明显, 碱胁迫较盐胁迫对植物造成的伤害更大(
Zhang and Mu, 2009 ;
焦伟红, 2011 ), 因此
P n 、
G s 及株高成为主要影响因子.综上, 盐碱胁迫下棉花生长生理的主要影响因子有叶中K
+ /Na
+ 比值、根长、鲜重、干重及胞间CO
2 浓度(
C i ). ...
21世纪农林本科生物系列课程改革的研究与实践(上)
1
2000
... 在棉花花铃期进行样品采集.每小区随机选取5株, 采用常规方法测定株高、根长、鲜重和干重(地上部和地下部).叶绿素提取采用乙醇提取法, 紫外分光光度法测定其含量(
李合生和陈华癸, 2000 ).分别将烘干的根、茎和叶粉碎过筛, 准确称取200 mg粉末样品, 利用硝酸-双氧水消解, 采用火焰光度计法(
鲍士旦, 2000 )测定Na
+ 和K
+ 含量.光合参数采用便携式光合仪(CARIS-2, 英国PP Systems公司)于中午12:00- 14:30取棉花叶片倒4叶进行测定.测定过程中光强为400 μmol·m
-2 ·s
-1 , 大气温度(25±1)°C, 大气CO
2 浓度(
C a )变化范围为(400±10) μmol·L
-1 .测定指标包括净光合速率(
P n )、胞间CO
2 浓度(
C i )、蒸腾速率(
E )和气孔导度(
G s ).气孔限制值计算公式为
L s =1-
C i /
C a (
C a 为空气中CO
2 浓度).每个处理重复测定5次, 取平均值. ...
盐碱胁迫对小麦种子萌发、早期幼苗生长及Na
+ 、K
+ 代谢的影响
1
2011
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
盐渍化土壤改良剂的筛选
1
2013
... 新疆作为我国最大的商品棉生产基地, 棉籽原料十分丰富, 棉粕是棉籽榨油形成的副产物, 因此新疆是我国棉粕主产区之一.棉粕价格低廉, 主要用作动物饲料, 但棉粕中的棉酚具有一定的毒性, 极大降低了其饲用价值(
Jalees et al., 2011 ;
陈琪和石剑华, 2013 ).
张娜等(2009) 以棉粕为原料, 对提取的棉蛋 白进行理化性质分析, 发现棉蛋白的提取率可达75.5%.
刘玉国等(2013) 针对盐渍化土壤对几种常见土壤改良剂进行初次筛选, 并对改良后盐渍化土壤电导率和pH值进行测定, 发现用葵粕、棉粕、柠檬酸和沼液改良盐渍化土壤效果较好. ...
粉煤灰与若干有机固体废弃物配施改良土壤的研究进展
1
2006
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
盐(NaCl)与碱(Na
2 CO
3 )对星星草胁迫作用的差异
1
1993
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
21世纪新疆土壤盐渍化调控与农业持续发展研究建议
1
2000
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
我国棉粕和棉籽蛋白营养成分和棉酚含量调研
0
2010
施加改良剂对重度盐碱地盐碱动态及杨树生长的影响
1
2009
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
粉垄改造利用盐碱地效果初探
1
2017
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
土壤盐渍化遥感应用研究进展
1
2006
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
盐碱胁迫对小冰麦相对生长率及茎叶离子积累的影响
1
2011
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
盐胁迫对不同基因型海岛棉光合作用及荧光特性的影响
1
2010
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
PAM对土壤物理性状影响的试验研究及机理分析
1
2005
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
盐胁迫下不同K
+ 吸收抑制剂对小麦根系K
+ /Na
+ 比和质膜相关蛋白活性的影响
1
2017
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
盐胁迫下二倍体马铃薯叶绿素含量和抗氧化酶活性的变化
1
2014
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
棉籽蛋白提取工艺及其主要理化性质研究
1
2009
... 新疆作为我国最大的商品棉生产基地, 棉籽原料十分丰富, 棉粕是棉籽榨油形成的副产物, 因此新疆是我国棉粕主产区之一.棉粕价格低廉, 主要用作动物饲料, 但棉粕中的棉酚具有一定的毒性, 极大降低了其饲用价值(
Jalees et al., 2011 ;
陈琪和石剑华, 2013 ).
张娜等(2009) 以棉粕为原料, 对提取的棉蛋 白进行理化性质分析, 发现棉蛋白的提取率可达75.5%.
刘玉国等(2013) 针对盐渍化土壤对几种常见土壤改良剂进行初次筛选, 并对改良后盐渍化土壤电导率和pH值进行测定, 发现用葵粕、棉粕、柠檬酸和沼液改良盐渍化土壤效果较好. ...
1950s中期以来东北地区盐碱地时空变化及成因分析
1
2010
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
1
1990
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
NaCl胁迫对甘薯植株体内K
+ 、Na
+ 和Cl
- 含量及生长的影响
1
2017
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
1
1983
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
Effects of salt stress on the reproductive biology of the halophyte
Plantago crassifolia 1
2005
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
Effects of cottonseed meal on hematological, biochemical and behavioral alterations in male Japanese quail (
Coturnix japonica )
1
2011
... 新疆作为我国最大的商品棉生产基地, 棉籽原料十分丰富, 棉粕是棉籽榨油形成的副产物, 因此新疆是我国棉粕主产区之一.棉粕价格低廉, 主要用作动物饲料, 但棉粕中的棉酚具有一定的毒性, 极大降低了其饲用价值(
Jalees et al., 2011 ;
陈琪和石剑华, 2013 ).
张娜等(2009) 以棉粕为原料, 对提取的棉蛋 白进行理化性质分析, 发现棉蛋白的提取率可达75.5%.
刘玉国等(2013) 针对盐渍化土壤对几种常见土壤改良剂进行初次筛选, 并对改良后盐渍化土壤电导率和pH值进行测定, 发现用葵粕、棉粕、柠檬酸和沼液改良盐渍化土壤效果较好. ...
Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (
Medicago sativa L.)
1
2010
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
Comparative physiology of salt and water stress
1
2002
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
Ameliorative effect of CaCl
2 on growth, membrane permeability and nutrient uptake in
Atriplex halimus subsp
1
2009
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
Perspectives on the physiology and structure of inward-rectifying K
+ channels in higher plants: biophysical implications for K
+ uptake
1
1994
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
Chloroplasts of salt-grown Arabidopsis seedlings are impaired in structure, genome copy number and transcript levels
1
2013
... 盐碱胁迫下, 植物体内积累大量Na
+ 进而抑制了对K
+ 的吸收, 导致K
+ 含量下降, 离子稳态失衡(
Schroeder et al., 1994 ;
Nedjimi and Daoud, 2009 ).研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官的Na
+ 含量显著降低, 促进其对K
+ 的吸收(
图1 ,
图2 ).保持较高的K
+ /Na
+ 比值是植株耐盐的重要策略(
周志林等, 2017 ).本研究表明, 在盐碱胁迫下, 添加棉粕可以使棉花不同器官K
+ /Na
+ 比值增加, 盐胁迫下棉花不同器官的K
+ /Na
+ 比值的变化幅度大于碱胁迫(
图3 ), 说明添加棉粕有助于维持棉花不同器官的K
+ 、Na
+ 平衡, 对棉花生长生理有促进作用可能是由于其增加对K
+ 的选择性吸收, 而降低对Na
+ 的吸收, 进而通过增加K
+ /Na
+ 比来维持细胞内的离子平衡(
张华宁等, 2017 ).叶绿体不仅是植物进行光合作用的重要部位, 也是对盐胁迫最敏感的细胞器(
Štefanić et al., 2013 ).植株在正常生长过程中, 叶绿素含量呈现动态平衡, 而盐胁迫会打破这种平衡, 导致叶绿素含量发生变化(
张景云等, 2014 ).本研究表明, 添加棉粕能够增加盐胁迫下棉花叶片的叶绿素含量, 提高光合作用效率, 从而促进棉花生长, 其中以添加6 000 kg·hm
-2 棉粕处理效果最显著, 而碱胁迫下效果不显著(
表1 ).这可能是由于碱胁迫下叶绿素分解酶活力增强, 促进叶绿素的分解(
陈俊, 2006 ).本研究表明, 盐碱胁迫下棉花叶片
P n 下降的主要原因是气孔因素, 这与前人的研究结果一致(
杨淑萍等, 2010 ).其中
P n 增加与
G s 和
C i 的升高有关, 表明添加棉粕能够有效调节棉花叶片的气孔开度(
表2 ).植物的生长过程对盐碱胁迫较为敏感, 而生长量是植物耐盐性的直接体现(
Boscaiu et al., 2005 ).本研究表明, 添加棉粕在一定程度上缓解了盐碱胁迫对棉花生长的抑制作用, 添加后棉花的株高、根长、鲜重和干重与单独盐碱胁迫处理相比均有显著差异, 其中添加6 000 kg·hm
-2 棉粕处理的效果最显著且盐胁迫下棉花生长指标的变化幅度较大(
表3 ).王安平等(2010)用棉粕开发的氨基酸肥能提高小麦的株高、叶面积、叶绿素和干物质含量, 使小麦产量显著增加.本研究也得到类似的结果, 这可能由于棉粕中除了含有蛋白质外, 还有粗脂肪、酸性及中性洗涤纤维等营养成分, 这些营养成分可能对棉花的生长有一定的促进作用.主成分分析结果表明, 盐胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
C i 、根长、鲜重和干重, 其中根、茎中K
+ 含量在次要影响因子中较为敏感; 碱胁迫下添加棉粕对棉花生长生理指标影响较大, 主要影响因子有叶中的K
+ /Na
+ 比值、
P n 、
G s 、株高、根长、鲜重、干重及茎和叶中Na
+ 含量, 其中叶片K
+ 含量和
C i 在次要影响因子中较为敏感(
表4 ). ...
Effects of intraspecific competition on growth and photosynthesis of
Atriplex prostrata 1
2005
... 盐碱胁迫是两种不同性质的胁迫.盐胁迫的危害主要体现在渗透胁迫和离子毒害两个方面(
Munns, 2002 ;
Wang et al., 2005 ), 而碱胁迫还包括高pH值引起的胁迫(
石德成和殷丽娟, 1993 ).研究表明, 随着盐碱浓度的增大, 碱胁迫下小冰麦(
Triticum aestivum-Agropyronin termedium )的生长率降幅大于盐胁迫, 并且碱胁迫对离子积累的干扰也较大(
杨国会和石德成, 2011 ).碱胁迫使细胞内离子稳态失衡是导致净光合速率(
P n )降低的原因(
Li et al., 2010 ).也有****认为低浓度盐短期处理时, 植物净光合速率降低主要是由气孔因素引起的; 而高浓度盐长期处理时, 则主要受非气孔因素的影响(
黄清荣等, 2018 ).将盐胁迫处理后的小麦(
T. aestivum )种子放回蒸馏水中能继续萌发, 而在碱胁迫下却多半死亡, 表明盐碱胁迫下, 小麦种子活力、生理及生长补偿效应机制不同(
蔺吉祥等, 2011 ).一般认为, 棉花(
Gossypium hirsutum )幼苗期和开花结铃期是对盐分较为敏感的时期(
赵可夫和王韶唐, 1990 ).研究表明, 棉花耐盐能力以出苗时最弱(
贾玉珍等, 1987 ), 且随着生育时期的不断推进而逐渐提高, 在蕾期、初花期耐盐能力又开始下降, 并于开花结铃盛期耐盐能力达到最强(
罗宾, 1983 ).目前, 多数研究集中于盐碱胁迫下植株幼苗期的生长生理变化(
阿曼古丽·买买提阿力等, 2017 ), 而对棉花花铃期耐盐机理的研究较少, 通过添加棉粕改良盐碱地的方式更为鲜见.因此, 深入研究棉粕在盐碱地治理方面的利用途径具有重要的实际意义. 为此, 本研究以新疆典型的2种盐渍化土壤(中性盐NaCl和碱性盐Na
2 CO
3 )为研究对象, 通过田间试验模拟重度盐碱土壤条件, 探讨添加棉粕对盐碱胁迫下棉花的生理生长指标和离子吸收的影响, 以期揭示棉粕改良盐渍化土壤的作用机理, 为开发盐渍化土壤棉花栽培技术提供借鉴. ...
Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage
Lathyrus quinquenervius 1
2009
... 综上所述, 盐碱胁迫下棉花生长生理补偿机制的影响因子有所不同.盐胁迫下, 气孔关闭, 阻碍CO
2 进入细胞内部, 因此
C i 成为主要影响因子; 盐碱胁迫均会导致棉花叶片
P n 和
G s 下降, 但在碱胁迫下, 棉花叶片
P n 和
G s 值极低, 说明叶片的光合作用发生适应性下调.由于
P n 逐渐增大, 消耗大量CO
2 , 同时
G s 逐渐减小, 影响外界CO
2 向细胞内部扩散, 从而导致
C i 下降.通常植株地上部受害情况比地下部表现更为明显, 碱胁迫较盐胁迫对植物造成的伤害更大(
Zhang and Mu, 2009 ;
焦伟红, 2011 ), 因此
P n 、
G s 及株高成为主要影响因子.综上, 盐碱胁迫下棉花生长生理的主要影响因子有叶中K
+ /Na
+ 比值、根长、鲜重、干重及胞间CO
2 浓度(
C i ). ...
Improvements of soil salt characteristics and nutrient status in an impermeable saline-sodic soil reclaimed with an improved drip irrigation while ridge planting
Lycium barbarum L
1
2017
... 盐碱土是盐土、碱土以及不同种盐化土、碱化土的总称. 我国盐碱地总面积约3.67×10
7 hm
2 , 具有分布广、面积大、种类多的特点, 主要分布于西部干旱、半干旱地区.新疆是典型的干旱区, 土壤盐渍化非常严重, 不仅成为制约当地农业发展的主要障碍, 也是影响绿洲生态稳定的重要因素(
田长彦等, 2000 ;
翁永玲和宫鹏, 2006 ;
张树文等, 2010 ).目前, 国内外有关盐碱地治理技术的研究较多(
王文杰等, 2009 ;
郭凯等, 2016 ;
韦本辉等, 2017 ;
Zhang et al., 2017 ), 其中对沸石、绿肥、粉煤灰、污泥和聚丙烯酰胺(
员学锋等, 2005 )等土壤改良剂研究较多, 相关技术逐渐成熟, 但是存在改良效果不佳和二次污染等负面效应(
牛花朋等, 2006 ).综合来看, 目前探索改良剂的治理技术时偏重应用效果, 忽略了与当地资源结合的优势. ...
备案号:
京ICP备16067583号-21 版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由
北京玛格泰克科技发展有限公司 设计开发
 ,*, 张开祥, 孟春梅, 安梦洁石河子大学农学院, 石河子 832000
,*, 张开祥, 孟春梅, 安梦洁石河子大学农学院, 石河子 832000
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT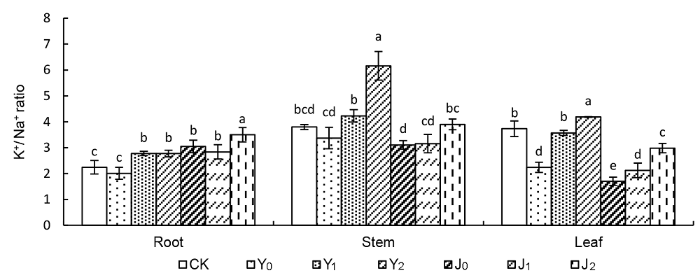 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT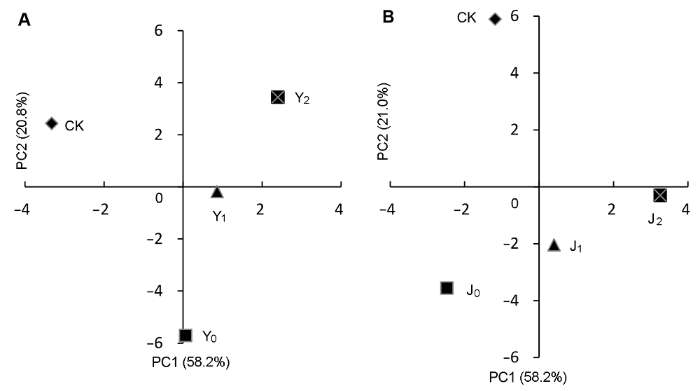 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}