
山东农业大学生命科学学院, 作物生物学国家重点实验室, 泰安 271000
引用本文
贡献者
基金资助
接受日期:2017-10-12网络出版日期:2018-11-1
-->Copyright
2018《植物学报》编辑部
Contributors
History
Received:Online:
摘要:
Abstract:
Key words:
番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高。由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(Gillaspy et al., 1993; Tanksley, 2004; Klee and Giovannoni, 2011; Azzi et al., 2015)。2012年, 番茄全基因组测序的完成极大地推动了番茄中诸多重要功能及调控基因研究, 是肉质果实研究领域的一个重要突破(The Tomato Genome Consortium, 2012)。
在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化。番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (Cong et al., 2008)。因此, 在番茄的整个生长发育过程中, 人们对其果实, 特别是形态、大小和品质等方面尤为感兴趣。番茄果实发育主要分为早期果实发育和果实成熟2个时期, 但对果实形态结构和大小而言, 主要决定于早期果实发育时期(图1)。番茄果实的早期发育大致分为3个阶段: (1) 子房发育及果实坐果; (2) 果实中细胞开始迅速分裂, 细胞数量及层数迅速增加; (3) 果实中细胞开始迅速增大, 从而促进番茄果实的生长发育(Gillaspy et al., 1993; Giovannoni, 2004; McAtee et al., 2013; Shinozaki et al., 2015)。
图 1
Figure 1

下载原图ZIP
生成PPT
图 1
番茄果实发育时期示意图
番茄果实发育过程主要分为2个时期: 果实早期发育期与果实成熟发育期。番茄果实的形态结构与大小主要决定于番茄果实早期发育期, 该时期主要特点为细胞分裂和增大。在细胞分裂过程中, 果皮细胞层数不断增加, 番茄果实逐渐增大; 而在细胞增大过程中, 细胞层数已不再增加, 细胞开始膨大, 导致番茄果实迅速增大。番茄果实成熟发育期的主要特点为果实颜色及果实中代谢产物的变化。
Figure 1
Fruit development in tomato
The development of tomato fruit can be divided into early fruit growth and fruit ripening, but for fruit morphological structure and fruit size, they are mainly determined the early stage of fruit development. In the period of tomato fruit early development, the characteristic is cell division and cell expansion. For the cell division process, the number of cell layers across the pericarp will be increased and fruit will be bigger than before. Fruit growth then proceeds into the cell expansion phase, in this phase, cell layers will not increase but cell enlargement occurred, leading to size increase of tomato fruit rapidly. The main characteristic of fruit ripening is the changes of fruit color and metabolites in tomato.
番茄果实生长发育是一个极其复杂的过程, 包含了诸多调控途径。本文重点综述番茄早期果实发育时期植物激素、细胞周期循环、表观遗传和物质代谢等多方面调控番茄果实发育的分子机制, 以期为深入研究番茄果实发育提供参考。
1 植物激素调控番茄果实的早期发育植物激素是植物体内产生的一类在很低浓度时即可对植物的生理过程产生重要调控作用的代谢产物。植物激素作用广泛, 在植物生长发育的各个过程及环境应答中均发挥重要作用(许智宏和李家洋, 2006)。
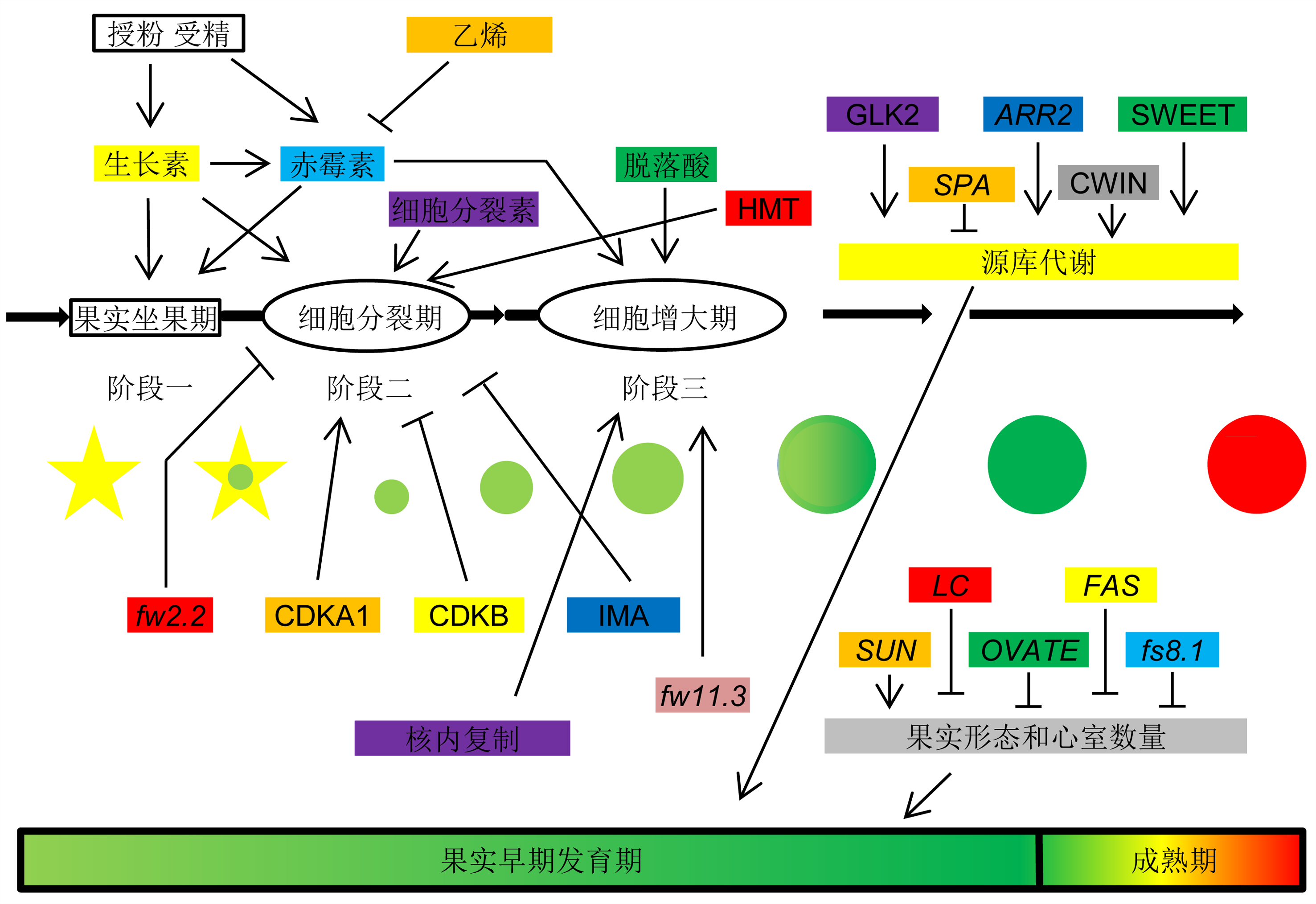
果实发育是一个复杂的过程, 受多种因素的共同调控。其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(Klee and Giovannoni, 2011; Ruan et al., 2012; Seymour et al., 2013; Kumar et al., 2014)。前期研究表明, 生长素、细胞分裂素、赤霉素、脱落酸和乙烯5类植物激素在果实的生长发育过程中起作用。随着研究的不断深入, 人们发现除上述5类激素外, 多胺、水杨酸、茉莉酸、油菜素内酯以及独脚金内酯等激素也在果实发育过程中起极其重要的作用(Li et al., 1992; Mehta et al., 2002; Vardhini and Rao, 2002; Ozga and Rienecke, 2003; Sheng et al., 2003; Srivastava and Handa, 2005; Kumar et al., 2014) (图2)。
图 2
Figure 2
下载原图ZIP
生成PPT
图 2
番茄果实早期发育时期相关途径调控示意图
箭头表示正调控, 障碍线表示负调控。CDKA1: 细胞周期依赖性激酶A1; CDKB: 细胞周期依赖性激酶B; CWIN: 细胞壁转化酶; GLK2: 类Golden 2转录因子; HMT: 组蛋白甲基转移酶; IMA: 分生组织活性抑制子; SWEET: 糖转运蛋白
Figure 2
Related pathways regulate early fruit growth in tomato
Arrows represent positive regulation, barred arrows represent negative regulation. CDKA1: Cyclin-dependent protein kinase A1; CDKB: Cyclin-dependent protein kinase B; CWIN: Cell wall invertase; GLK2: Golden 2-like; HMT: Histone methyltransferase; IMA: Inhibitor of meristem activity; SWEET: Sugar transporter
在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育。生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂。在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(Srivastava and Handa, 2005; Kumar et al., 2014)。当过表达番茄生长素响应因子(auxin response factor, ARF) SlARF9时, 果实细胞的分裂能力下降, 从而导致转基因番茄的果实明显小于野生型(de Jong et al., 2015)。de Jong等(2011)研究发现, SlARF7不仅作为生长素响应因子在番茄的坐果期起负调控作用, 还可作为赤霉素的一个调控因子调控果实的发育。细胞分裂素在番茄果实坐果和早期发育过程中也具重要作用。在番茄果实发育过程中, 其细胞数量逐渐增多, 主要由于内源性细胞分裂素水平升高所造成(Matsuo et al., 2012)。另外, 外施细胞分裂素会产生单性结实现象(Mariotti et al., 2011; Matsuo et al., 2012; Kumar et al., 2014)。除上述2种激素外, 赤霉素、乙烯和脱落酸在番茄果实早期发育过程中的作用也不可或缺(Serrani et al., 2007)。对番茄花朵外施赤霉素可导致其子房细胞明显增大, 甚至产生单性结实(Fos et al., 2000)。乙烯可通过调控赤霉素途径来抑制番茄坐果, 外施乙烯抑制剂1-MCP会导致单性结实产生(Shinozaki et al., 2015)。脱落酸则主要参与番茄果实增大的过程, 其缺失突变体的果实明显小于野生型(Gillaspy et al., 1993; Nitsch et al., 2012) (图2)。
甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质。但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作。在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(Gao et al., 2004; Zhang et al., 2006; Hancock et al., 2011; Choudhary et al., 2012; Noreen et al., 2013; Masood et al., 2016)。另外, Park等(2007)研究发现, 在番茄中表达抗逆基因codA, 在没有胁迫的条件下, 能够增大生殖器官花和果实的大小, 该现象可能是codA基因调控细胞分裂相关基因所致。本研究组的研究表明, 在正常生长条件下, 转codA和BADH基因番茄果实的果皮中生长素、细胞分裂素、赤霉素、脱落酸和油菜素内酯的含量均高于野生型(未发表数据), 故推测甜菜碱可能通过调控植物激素的合成从而调控细胞的分裂和增大, 最终促进番茄果实的生长发育。但其中的分子机理及甜菜碱与植物激素产生互作的机制目前并不清楚, 尚待进一步研究。
植物激素对番茄果实生长发育是必需的, 其参与植物生长发育的各个阶段。植物激素在调控果实发育时, 形成一个调控网络, 多种激素通过协同或拮抗作用行使功能。
2 细胞分裂、膨大与核内复制共同调控番茄果实的早期发育植物器官的生长发育依赖于细胞分裂和膨大过程。细胞分裂的活性决定了细胞的数量从而决定了器官的形成; 细胞膨大的活性则决定其最终大小(Chevalier et al., 2014)。
番茄果实的发育起始于花雌蕊子房的膨大, 其中主要包括细胞分裂和膨大, 即番茄果实增大是细胞分裂和膨大的结果。
在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作。果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(Alpert et al., 1995; Alpert and Tanksley, 1996; Frary et al., 2000)。作为一个主要的数量性状位点(quantitative trait locus, QTL), fw2.2的差异表达可导致驯化番茄的果重比野生型增加30%。fw2.2在细胞分裂过程中起负调控因子的作用, 其通过抑制细胞分裂来影响番茄果实的最终大小。另一个重要基因fw11.3 (Fruit weight 11.3)则在细胞增大的过程中起作用, 过表达fw11.3基因可显著增大番茄果实果皮细胞的大小, 从而起到增大果实的作用(Mu et al., 2017)。细胞周期依赖性激酶(cyclin-dependent protein kinases, CDKs)是细胞周期循环途径的重要调控因子, 其需与细胞周期蛋白(Cyclin)互作方可发挥作用(Joubès et al., 2000; Adachi et al., 2009)。CDKA是细胞周期依赖性激酶家族的重要组分之一, 可参与细胞增殖能力的维持, 促进雌、雄配子体、胚和胚乳的发育, 也是核内复制现象产生的必要因素(Hemerly et al., 1993; Segers et al., 1996; Joubès et al., 1999, 2000; Leiva-Neto et al., 2004; Verkest et al., 2005; Iwakawa et al., 2006; Gonzalez et al., 2007; Zhao et al., 2012; Czerednik et al., 2015)。CDKA1是细胞周期过程中的关键调控因子, 过表达CDKA1基因会导致番茄果实产生更大的胎座、更厚的果皮和更少的种子数量, 观察果皮可以发现其细胞层数和单位面积的细胞数量都明显增加(Czerednik et al., 2015)。过表达CDKB1和CDKB2则可产生较野生型略小的果实, 与降低CDKA1基因表达量的结果相似(Czerednik et al., 2012)。IMA (inhibitor of meristem activity)微型锌指蛋白是分生组织活性抑制剂, 在番茄果实中参与细胞分裂、分化和激素调控相关的多重调控途径(Sicard et al., 2008)。过表达IMA的植株, 心皮细胞数量减少, 心皮变小, 产生了更小的花和果实, 表明IMA的表达抑制了细胞分裂; 沉默IMA的植株心皮更大且数量增多(Sicard et al., 2008) (图2)。
核内复制在番茄果实早期发育过程中也具有重要作用。核内复制广泛存在于植物中, 但在番茄果实发育中尤为突出。在番茄果实早期发育时期, 果皮细胞存在内多倍性现象, 即各层细胞中DNA含量不同, 此为核内复制引起(Chevalier et al., 2014)。根据核质比理论, 核内复制产生了较高水平的核内DNA倍数, 细胞本身为了适应高倍数的DNA, 从而增大本身体积(Cheniclet et al., 2005)。与此同时, 核内复制还会引起细胞核与细胞质的有效交流, 并通过建立一个高度结构化的细胞系统来帮助维持细胞质成分的稳态平衡; 在这个细胞系统中, 多重的生理功能整合到一起共同在果实发育期间促进细胞生长(Azzi et al., 2015) (图2)。
细胞分裂、膨大和核内复制协同调控番茄果实的早期发育, 共同构建生长发育模型并为果实器官形成提供生长动力, 最终决定果实的重量和大小。因此, 通过修饰改造有丝分裂和核内复制相关靶基因的表达模式来调控番茄果实的生长发育过程将是今后研究的热点和重点(Azzi et al., 2015)。
3 果实形态与心室数量调控番茄果实的早期发育在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(Cong et al., 2008; Rodrlguez et al., 2011; van der Knaap et al., 2014)。番茄果实形态与心室数量变化主要发生于番茄果实早期发育阶段。相关研究表明, 调控番茄果实形态的主要基因为SUN、OVATE和fs8.1 (Xiao et al., 2008; Rodrlguez et al., 2011; van der Knaap et al., 2014; Sun et al., 2015); 而调控番茄果实心室数量的主要基因为FAS (FASCIATED)和LC (LOCULE NUMBER)。此外, FAS和LC基因还可调控番茄果实呈扁平形态(Cong et al., 2008; Munos et al., 2011; Rodrlguez et al., 2011; van der Knaap et al., 2014)。
SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(Xiao et al., 2008; Wu et al., 2011)。OVATE基因编码OVATE蛋白家族成员, 参与转录抑制过程从而调控番茄果实的形态变化(Liu et al., 2002; van der Knaap et al., 2014)。fs8.1基因位于番茄的第8号染色体上。形态学和组织学分析显示, fs8.1的作用主要是通过增加从近端至远端方向上的细胞数量进而导致番茄果实伸长(Sun et al., 2015)。LC基因编码的产物可能是WUSCHEL的直系同源物, 其可控制分生组织的大小和心室数量, 从而调控番茄果实的形态与大小(Munos et al., 2011; van der Knaap et al., 2014)。FAS基因属于YABBY家族成员, 其可调控番茄果实心室数量增多, 从而使番茄果实呈扁平状或牛心樱桃状(Cong et al., 2008; Munos et al., 2011)。值得一提的是, fw11.3与FAS基因均位于番茄的第11号染色体的长臂端, 且位置十分接近, 但fw11.3与FAS不是等位基因, 而是两个不同的基因, 具有不同的调控功能(Huang and van der Knaap, 2011) (图2)。
Lin等(2014)研究表明, 在番茄长期驯化过程中, 野生番茄果实在形态、大小、重量和颜色等方面均发生了显著变化。他们通过群体遗传学分析, 发现番茄果实的变大经历了两步驯化过程, 即从野生醋栗番茄到樱桃番茄再到大栽培番茄。该研究揭示了在驯化过程中分别有5个(fw1.1、fw5.2、fw7.2、fw12.1和lcn12.1)和13个(fw2.1、fw2.2、fw2.3、lcn2.1、lcn2.2、fw3.2、lcn3.1、fw9.1、fw9.3、lcn10.1、fw11.1、fw11.2和fw11.3)果实重量基因受到人类的定向选择。与此同时, 他们还构建了高密度的番茄遗传变异组图谱, 为阐明番茄的进化历史、基因挖掘和分子育种奠定了基础。
4 表观遗传调控番茄果实的早期发育表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变。这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递。表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(Vermaak et al., 2003; Chan et al., 2005; Reyes, 2006; Li et al., 2007; Teyssier et al., 2015)。目前, 表观遗传调控功能主要在模式植物拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和玉米(Zea mays)中进行研究, 而在大多数作物中并未开展。然而, 越来越多的证据表明, 表观遗传调控在相关性状表现过程中不可缺少。例如, 植物对外界环境的适应性(Mirouze and Paszkowski, 2011; Sahu et al., 2013)、杂种优势(Shen et al., 2012)、开花时间的调控(Michaels, 2009; He, 2012)、果实品质(Seymour et al., 2008)以及配子形成和种子发育(Feng et al., 2010; Wollmann and Berger, 2012)等。
表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程。DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(Law and Jacobsen, 2010)或响应外界环境时, DNA甲基化水平和模式会发生动态变化(Chinnusamy and Zhu, 2009; Sahu et al., 2013; Teyssier et al., 2015)。对果实而言, DNA甲基化动态变化的研究仍然很少, 目前仅在番茄果实中进行。越来越多的研究表明, 在番茄果实早期发育过程中, 果皮细胞的DNA失去甲基化(Teyssier et al., 2008; Zhong et al., 2013)。但这种现象仅出现在果皮组织中, 在腔室组织中并未发现, 可能具有组织特异性。另外, 在番茄果实发育过程中, 不同组织的DNA甲基转移酶(DNA methyltransferase, DNMT)基因会出现差异性表达(Teys- sier et al., 2008)。
除了DNA甲基化, 组蛋白修饰在番茄果实早期发育过程中也起重要作用。组蛋白翻译后修饰能够影响染色体的结构与基因表达。它们通过多种途径影响染色体的组装, 其中主要包括核小体净电荷修饰(Strahl and Allis, 2000)。在番茄果实早期发育过程中, 通过组蛋白翻译后修饰途径, 使相关基因发生差异性表达。多个组蛋白甲基转移酶(histone methyltransferase, HMT)基因在番茄果实早期发育过程中的细胞分裂阶段发生表达(Cigliano et al., 2013)。例如, 基因SLEZ2 (Enhancer of zeste 2)可编码zeste结构域的增强子HMT (Kit et al., 2010), 说明组蛋白甲基化在番茄果实发育过程中的细胞分裂阶段格外活跃(图2)。
此外, Lauria和Rossi (2011)研究发现, 表观遗传调控在番茄果实生长发育过程中行使重要功能。表观基因组多态性现象(DNA甲基化和组蛋白修饰)在基因组功能发挥过程中具重要作用, 且可能导致表型多样性, 控制复杂的形状特性甚至可被遗传(Zhang et al., 2008; Johannes et al., 2009; Schmitz et al., 2011)。
5 番茄果实早期发育过程中的源库代谢平衡在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理。然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (Tilman et al., 2011; Bermúdez et al., 2014)。目前, 更多采用植物育种来实现增产的目的。植物育种是通过提高收获指数(harvest index, HI), 从而提高作物的产量(Lippman et al., 2007; Yang and Zhang, 2010)。植物产量的形成实际是光合同化物生产与分配的过程, 植物获得高产的基本途径是尽量增加光合同化物的积累量, 并使之尽可能多地分配转运到产品器官中(álvaro et al., 2008; Yu et al., 2015)。
番茄果实具有农业和基础研究价值。由于番茄具有生长周期短、繁殖速度快和自花授粉等特点, 其已成为研究肉质果实的模式植物(Klee and Giovannoni, 2011; Azzi et al., 2015)。果实是番茄的主要库器官, 其果实的生长发育及产量不仅与叶片产生的碳同化物有关, 还与同化物是否有效地分配到果实这个库器官密切相关(Carrari and Fernie, 2006)。
在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量。转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(Powell et al., 2012; Nguyen et al., 2014)。同样, 番茄APRR2-LIKE (ARABIDOPSIS PSEUDO RESPONSE REGULATOR 2-LIKE)基因可影响番茄果实细胞中质体的数量和区域分布, 进而影响未成熟果实的叶绿素和成熟果实的类胡萝卜素含量(Pan et al., 2013)。SPA (sugar partitioning-affecting)基因与糖分配相关, 其可作用于质体代谢过程, 限制蔗糖从叶片转运至果实的速率(Bermúdez et al., 2008)。在SPA沉默株系中, 其源叶中的可溶性糖和淀粉含量降低, 而成熟果实中的淀粉量却增加了2倍多, 沉默株系结果更多且果实更重, 收获指数更高(Bermúdez et al., 2014)。细胞壁转化酶(cell wall invertase, CWIN)的活性对于果实和种子的发育及最终的作物产量也至关重要(Ruan, 2014; Liu et al., 2016)。在番茄株系中沉默CWIN基因会导致种子败育(Jin et al., 2009; Zanor et al., 2009)。除上述功能外, CWIN还可调控番茄果实中蔗糖的卸载途径及己糖和生物量的积累(Damon et al., 1988; Ruan and Patrick, 1995; Ho, 1996; Balibrea et al., 1999; McCurdy et al., 2010; Albacete et al., 2014; Liu et al., 2016)。另外, 有研究表明, SWEET (sugars will eventually be exported transporters)蛋白是糖流的携带者, 其在蔗糖转运体SUT1 (sucrose transporter 1)和H+-ATPases的协助下可将蔗糖流从韧皮部薄壁组织运输至筛管伴胞复合体并最终转运至库器官, 从而调控源库之间的代谢平衡(Chen et al., 2010, 2012)。本研究组最新的研究表明, 甜菜碱可调控番茄果实的源库代谢, 在转codA和BADH基因株系中, 番茄叶片的光合作用强于野生型株系, 且番茄果实中的同化物含量明显升高(未发表数据), 表明甜菜碱可能在调控番茄果实的源库代谢平衡方面起重要作用(图2)。
6 研究展望番茄是重要的蔬菜作物, 也是肉质果实的模式植物, 同时还是最早用于转基因的作物之一。番茄果实的生长发育是一个极其复杂且受到精密调控的过程, 其中包括细胞周期、植物激素、表观遗传、源库代谢以及果实形态和心室数量调控等。而对于番茄果实的产量和质量, 其果实的早期发育过程占据了十分重要的地位。番茄果实早期发育过程主要包括细胞分裂和增大, 期间受到生长素、赤霉素和细胞分裂素等多种激素以及表观遗传(如DNA甲基化和组蛋白修饰等)的调控, 同时光合作用产生同化物并运输至果实细胞中进行填充, 从而完成果实生长与发育。
番茄果实发育的调控研究中, 前人已在细胞周期、激素调节、源库代谢、表观遗传以及果实形态和心室数量调控等方面做了大量工作。然而由于果实的发育及最终果实的重量和质量是多种因素综合调控的结果, 故明确番茄果实不同发育时期的主要调控机制和重要的调控基因, 对促进果实生长发育及提高果实产量和质量具有重要的理论指导意义。
虽然目前已完成番茄全基因组测序, 但仍有许多重要的功能及调控基因需要揭示, 故对番茄果实生长发育过程生理和分子机制的研究还有很长的路要走。对番茄果实的研究不仅有助于人们进一步认识植物生长发育的基本生物学问题, 还有助于促进基础理论研究成果在生产中的应用。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | DOI:10.3969/j.issn.1674-3466.2006.05.001URLThis special issue of “Plant Hormones” by Chinese Bulletin of Botany is dedicated to the “Xiangshan Conference—Plant Hormones and Green Revolution”, which will be held in October 2006 in Beijing. Here, we present an overview on the plant hormone research in China for readers of interest. Until 1980s, research on plant hormones in China mainly focused on physiological studies. With rapid growth of the Chinese economy, a rising generation of young scientists has been able to make significant progress in elucidating the regulatory mechanisms of plant hormone metabolism, transport and signal transduction. In particular, Chinese scientists have made substantial contributions to our current understanding to the field by the identification of hormone receptors, the hormone-regulated plant architecture formation as well as interactions among different hormone signaling pathways. Finally, based on the trends of the field as well as the potentials of Chinese scientists, we offer a prospective view on the future studies on plant hormones in China. [本文引用: 1] |
| [2] | DOI:10.1016/j.ydbio.2009.03.002PMID:19285489URLA-type cyclin-dependent kinase (CDKA) is an ortholog of yeast Cdc2/Cdc28p, and is assumed to have an essential function in plant growth and organogenesis. Previous studies revealed that its kinase activity is controlled by post-translational modifications, such as binding to cyclins and phosphorylations, but its transcriptional regulation is poorly understood. Here, we generated a promoter dissection series of Arabidopsis ( Arabidopsis thaliana) CDKA;1, and used -glucuronidase ( GUS) gene-fused reporter constructs for expression analyses in planta. The results revealed two types of transcriptional control in shoots: general quantitative regulation and cell type-specific regulation. We identified a promoter region that promotes CDKA;1 expression in the leaf epidermis, but not in the L1 layer of the shoot apical meristem. This region also directed abaxial side-biased expression, which may be linked to the adaxial/abaxial side specification. Another reporter construct showed that CDKA;1 expression in the inner layers of leaves is controlled by a distinct regulatory region in the promoter. These results suggest that the transcriptional regulation of CDKA;1 may play a key role in proper development of leaves by coordinating cell division and differentiation of different cell types. [本文引用: 1] |
| [3] | DOI:10.1093/jxb/eru347.PMID:4203140URLCytokinins and cell wall invertase are positive players in regulating fruit sink strength, growth, and yield under salinity as components of the same signalling cascade establishing and developing sink organs. Salinization of water and soil has a negative impact on tomato (Solanum lycopersicum L.) productivity by reducing growth of sink organs and by inducing senescence in source leaves. It has been hypothesized that yield stability implies the maintenance or increase of sink activity in the reproductive structures, thus contributing to the transport of assimilates from the source leaves through changes in sucrolytic enzymes and their regulation by phytohormones. In this study, classical and functional physiological approaches have been integrated to study the influence of metabolic and hormonal factors on tomato fruit sink activity, growth, and yield: (i) exogenous hormones were applied to plants, and (ii) transgenic plants overexpressing the cell wall invertase (cwInv) gene CIN1 in the fruits and de novo cytokinin (CK) biosynthesis gene IPT in the roots were constructed. Although salinity reduces fruit growth, sink activity, and trans-zeatin (tZ) concentrations, it increases the ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC) during the actively growing period (25 days after anthesis). Indeed, exogenous application of the CK analogue kinetin to salinized actively growing fruits recovered sucrolytic activities (mainly cwInv and sucrose synthase), sink strength, and fruit weight, whereas the ethylene-releasing compound ethephon had a negative effect in equivalent non-stressed fruits. Fruit yield was increased by both the constitutive expression of CIN1 in the fruits (up to 4-fold) or IPT in the root (up to 30%), owing to an increase in the fruit number (lower flower abortion) and in fruit weight. This is possibly related to a recovery of sink activity in reproductive tissues due to both (i) increase in sucrolytic activities (cwInv, sucrose synthase, and vacuolar and cytoplasmic invertases) and tZ concentration, and (ii) a decrease in the ACC levels and the activity of the invertase inhibitor. This study provides new functional evidences about the role of metabolic and hormonal inter-regulation of local sink processes in controlling tomato fruit sink activity, growth, and yield under salinity. [本文引用: 1] |
| [4] | DOI:10.1007/BF00223911PMID:24169988URLWe have shown that a major QTL for fruit weight ( fw2.2 ) maps to the same position on chromosome 2 in the green-fruited wild tomato species, Lycopersicon pennellii and in the red-fruited wild tomato species, L. pimpinellifolium . An introgression line F 2 derived from L. esculentum (tomato) x L. pennellii and a backcross 1 (BC 1 ) population derived from L. esculentum x L. pimpinellifolium both place fw2.2 near TG91 and TG167 on chromosome 2 of the tomato highdensity linkage map. fw2.2 accounts for 30% and 47% of the total phenotypic variance in the L. pimpinellifolium and L. pennellii populations, respectively, indicating that this is a major QTL controlling fruit weight in both species. Partial dominance (d/a of 0.44) was observed for the L. pennellii allele of fw 2.2 as compared with the L. esculentum allele. A QTL with very similar phenotypic affects and gene action has also been identified and mapped to the same chromosomal region in other wild tomato accessions: L. cheesmanii and L. pimpinellifolium . Together, these data suggest that fw2.2 represents an orthologous QTL (i.e., derived by speciation as opposed to duplication) common to most, if not all, wild tomato species. High-resolution mapping may ultimately lead to the cloning of this key locus controlling fruit development in tomato. [本文引用: 1] |
| [5] | DOI:10.1073/pnas.93.26.15503URLA high-resolution physical and genetic map of a major fruit weight quantitative trait locus (QTL), fw2.2, has been constructed for a region of tomato chromosome 2. Using an F2nearly isogenic line mapping population (3472 individuals) derived from Lycopersicon esculentum (domesticated tomato) Lycopersicon pennellii (wild tomato), fw2.2 has been placed near TG91 and TG167, which have an interval distance of 0.13 0.03 centimorgan. The physical distance between TG91 and TG167 was estimated to be 150 kb by pulsed-field gel electrophoresis of tomato DNA. A physical contig composed of six yeast artificial chromosomes (YACs) and encompassing fw2.2 was isolated. No rearrangements or chimerisms were detected within the YAC contig based on restriction fragment length polymorphism analysis using YAC-end sequences and anchored molecular markers from the high-resolution map. Based on genetic recombination events, fw2.2 could be narrowed down to a region less than 150 kb between molecular markers TG91 and HSF24 and included within two YACs: YAC264 (210 kb) and YAC355 (300 kb). This marks the first time, to our knowledge, that a QTL has been mapped with such precision and delimited to a segment of cloned DNA. The fact that the phenotypic effect of the fw2.2 QTL can be mapped to a small interval suggests that the action of this QTL is likely due to a single gene. The development of the high-resolution genetic map, in combination with the physical YAC contig, suggests that the gene responsible for this QTL and other QTLs in plants can be isolated using a positional cloning strategy. The cloning of fw2.2 will likely lead to a better understanding of the molecular biology of fruit development and to the genetic engineering of fruit size characteristics. [本文引用: 1] |
| [6] | DOI:10.2135/cropsci2007.10.0545URLABSTRACT To understand yield limitations imposed by the source–sink relationship and the effects that breeding had on it, the source–sink ratio was modifi ed on 24 durum wheat cultivars released in different periods. Field experiments were conducted during 3 yr, with treatments consisting of removing the fl ag-leaf blade, degraining half of the spike, and intact checks. Grains from near-basal spikelets tended to be the most sensitive to source–sink modifications. Degraining increased grain weight up to 15% in modern cultivars, which were more source limited and more responsive to source–sink modifications than the old ones. Our results suggest a source–sink colimitation for grain fi lling in Mediterranean environments. Translocation of preanthesis assimilates was responsible for 25, 27, and 33% of grain weight in old, intermediate, and modern cultivars, respectively. Defoliation did not alter dry matter translocation (DMT) from the main stem, as the reduction in translocation of leaf blades was compensated with translocation increases from the chaff. Sink demand determined DMT that was mostly regulated by the stem. Our results indicate that future yield improvements may be achieved by strengthening the source capacity. [本文引用: 1] |
| [7] | DOI:10.1093/jxb/eru527PMID:25573859URLAbstract Tomato (Solanum lycopersicum Mill.) represents a model species for all fleshy fruits due to its biological cycle and the availability of numerous genetic and molecular resources. Its importance in human nutrition has made it one of the most valuable worldwide commodities. Tomato fruit size results from the combination of cell number and cell size, which are determined by both cell division and expansion. As fruit growth is mainly driven by cell expansion, cells from the (fleshy) pericarp tissue become highly polyploid according to the endoreduplication process, reaching a DNA content rarely encountered in other plant species (between 2C and 512C). Both cell division and cell expansion are under the control of complex interactions between hormone signalling and carbon partitioning, which establish crucial determinants of the quality of ripe fruit, such as the final size, weight, and shape, and organoleptic and nutritional traits. This review describes the genes known to contribute to fruit growth in tomato. The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com. [本文引用: 4] |
| [8] | DOI:10.1071/PP99026URL [本文引用: 1] |
| [9] | DOI:10.1111/tpj.12418PMID:24372694URLLimitations in our understanding about the mechanisms that underlie source-sink assimilate partitioning are increasingly becoming a major hurdle for crop yield enhancement via metabolic engineering. By means of a comprehensive approach, this work reports the functional characterization of a DnaJ chaperone related-protein (named as SPA; sugar partition-affecting) that is involved in assimilate partitioning in tomato plants. SPA protein was found to be targeted to the chloroplast thylakoid membranes. SPA-RNAi tomato plants produced more and heavier fruits compared with controls, thus resulting in a considerable increment in harvest index. The transgenic plants also displayed increased pigment levels and reduced sucrose, glucose and fructose contents in leaves. Detailed metabolic and enzymatic activities analyses showed that sugar phosphate intermediates were increased while the activity of phosphoglucomutase, sugar kinases and invertases was reduced in the photosynthetic organs of the silenced plants. These changes would be anticipated to promote carbon export from foliar tissues. The combined results suggested that the tomato SPA protein plays an important role in plastid metabolism and mediates the source-sink relationships by affecting the rate of carbon translocation to fruits. [本文引用: 2] |
| [10] | DOI:10.1093/jxb/ern146PMID:18552354URLIn tomato, numerous wild-related species have been demonstrated to be untapped sources of valuable genetic variability, including pathogen-resistance genes, nutritional, and industrial quality traits. From a collection ofS. pennelliiintrogressed lines, 889 fruit metabolic loci (QML) and 326 yield-associated loci (YAL), distributed across the tomato genome, had been identified previously. By using a combination of molecular marker sequence analysis, PCR amplification and sequencing, analysis of allelic variation, and evaluation of co-response between gene expression and metabolite composition traits, the present report, provides a comprehensive list of candidate genes co-localizing with a subset of 106 QML and 20 YAL associated either with important agronomic or nutritional characteristics. This combined strategy allowed the identification and analysis of 127 candidate genes located in 16 regions of the tomato genome. Eighty-five genes were cloned and partially sequenced, totalling 45 816 and 45 787 bases fromS. lycopersicumandS. pennellii, respectively. Allelic variation at the amino acid level was confirmed for 37 of these candidates. Furthermore, out of the 127 gene-metabolite co-locations, some 56 were recovered following correlation of parallel transcript and metabolite profiling. Results obtained here represent the initial steps in the integration of genetic, genomic, and expressional patterns of genes co-localizing with chemical compositional traits of the tomato fruit. [本文引用: 1] |
| [11] | DOI:10.1093/jxb/erj020PMID:16449380URLAbstract The development and maturation of tomato fruits has received considerable attention because of both the uniqueness of such processes to the biology of plants and the importance of these fruits as a component of the human diet. Molecular and genetic analysis of fruit development, and especially ripening of fleshy fruits, has resulted in significant gains in knowledge over recent years. A large amount of knowledge has been gathered on ethylene biosynthesis and response, cell wall metabolism, and environmental factors, such as light, that impact ripening. Considerably less attention has been paid directly to the general metabolic shifts that underpin these responses. Given the vast complexity of fruit metabolism, the focus chosen for this review is on primary metabolites and those secondary metabolites that are important with respect to fruit quality. Here, recent advances in dissecting tomato metabolic pathways are reviewed. Also discussed are recent examples in which the combined application of metabolic and transcriptional profiling, aimed at identifying candidate genes for modifying metabolite contents, was used. [本文引用: 1] |
| [12] | [本文引用: 1] |
| [13] | DOI:10.1038/nature09606PMID:3000469URLAbstract Sugar efflux transporters are essential for the maintenance of animal blood glucose levels, plant nectar production, and plant seed and pollen development. Despite broad biological importance, the identity of sugar efflux transporters has remained elusive. Using optical glucose sensors, we identified a new class of sugar transporters, named SWEETs, and show that at least six out of seventeen Arabidopsis, two out of over twenty rice and two out of seven homologues in Caenorhabditis elegans, and the single copy human protein, mediate glucose transport. Arabidopsis SWEET8 is essential for pollen viability, and the rice homologues SWEET11 and SWEET14 are specifically exploited by bacterial pathogens for virulence by means of direct binding of a bacterial effector to the SWEET promoter. Bacterial symbionts and fungal and bacterial pathogens induce the expression of different SWEET genes, indicating that the sugar efflux function of SWEET transporters is probably targeted by pathogens and symbionts for nutritional gain. The metazoan homologues may be involved in sugar efflux from intestinal, liver, epididymis and mammary cells. [本文引用: 1] |
| [14] | DOI:10.1126/science.1213351PMID:22157085URLPlants transport fixed carbon predominantly as sucrose, which is produced in mesophyll cells and imported into phloem cells for translocation throughout the plant. It is not known how sucrose migrates from sites of synthesis in the mesophyll to the phloem, or which cells mediate efflux into the apoplasm as a prerequisite for phloem loading by the SUT sucrose-H(proton) cotransporters. Using optical sucrose sensors, we identified a subfamily of SWEET sucrose efflux transporters. AtSWEET11 and 12 localize to the plasma membrane of the phloem. Mutant plants carrying insertions in AtSWEET11 and 12 are defective in phloem loading, thus revealing a two-step mechanism of SWEET-mediated export from parenchyma cells feeding H-coupled import into the sieve element-companion cell complex. We discuss how restriction of intercellular transport to the interface of adjacent phloem cells may be an effective mechanism to limit the availability of photosynthetic carbon in the leaf apoplasm in order to prevent pathogen infections. [本文引用: 1] |
| [15] | DOI:10.1104/pp.105.068767URL [本文引用: 1] |
| [16] | DOI:10.1093/jxb/ert366PMID:24187421URLThe growth of a plant organ depends upon the developmental processes of cell division and cell expansion. The activity of cell divisions sets the number of cells that will make up the organ; the cell expansion activity then determines its final size. Among the various mechanisms that may influence the determination of cell size, endopolyploidy by means of endoreduplication appears to be of great importance in plants. Endoreduplication is widespread in plants and supports the process of differentiation of cells and organs. Its functional role in plant cells is not fully understood, although it is commonly associated with ploidy-dependent cell expansion. During the development of tomato fruit, cells from the (fleshy) pericarp tissue become highly polyploid, reaching a DNA content barely encountered in other plant species (between 2C and 512C). Recent investigations using tomato fruit development as a model provided new data in favour of the long-standing karyoplasmic ratio theory, stating that cells tend to adjust their cytoplasmic volume to the nuclear DNA content. By establishing a highly structured cellular system where multiple physiological functions are integrated, endoreduplication does act as a morphogenetic factor supporting cell growth during tomato fruit development. [本文引用: 2] |
| [17] | DOI:10.1016/j.pbi.2008.12.006PMID:19179104URLGene expression driven by developmental and stress cues often depends on nucleosome histone post-translational modifications and sometimes on DNA methylation. A number of studies have shown that these DNA and histone modifications play a key role in gene expression and plant development under stress. Most of these stress-induced modifications are reset to the basal level once the stress is relieved, while some of the modifications may be stable, that is, may be carried forward as ‘stress memory’ and may be inherited across mitotic or even meiotic cell divisions. Epigenetic stress memory may help plants more effectively cope with subsequent stresses. Comparative studies on stress-responsive epigenomes and transcriptomes will enhance our understanding of stress adaptation of plants. [本文引用: 1] |
| [18] | [本文引用: 1] |
| [19] | DOI:10.1186/1471-2164-14-57PMID:23356725URLpAbstract/p pBackground/p pHistone post-translational modifications (HPTMs) including acetylation and methylation have been recognized as playing a crucial role in epigenetic regulation of plant growth and development. Although itSolanum lycopersicum/it is a dicot model plant as well as an important crop, systematic analysis and expression profiling of histone modifier genes (itHMs/it) in tomato are sketchy./p pResults/p pBased on recently released tomato whole-genome sequences, we identified itin silico/it 32 histone acetyltransferases (HATs), 15 histone deacetylases (HDACs), 52 histone methytransferases (HMTs) and 26 histone demethylases (HDMs), and compared them with those detected in Arabidopsis (itArabidopsis thaliana/it), maize (itZea mays/it) and rice (itOryza sativa/it) orthologs. Comprehensive analysis of the protein domain architecture and phylogeny revealed the presence of non-canonical motifs and new domain combinations, thereby suggesting for HATs the existence of a new family in plants. Due to species-specific diversification during evolutionary history tomato has fewer HMs than Arabidopsis. The transcription profiles of itHMs/it within tomato organs revealed a broad functional role for some itHMs/it and a more specific activity for others, suggesting key itHM/it regulators in tomato development. Finally, we explored itS. pennellii/it introgression lines (ILs) and integrated the map position of itHMs/it, their expression profiles and the phenotype of ILs. We thereby proved that the strategy was useful to identify itHM/it candidates involved in carotenoid biosynthesis in tomato fruits./p pConclusions/p pIn this study, we reveal the structure, phylogeny and spatial expression of members belonging to the classical families of HMs in tomato. We provide a framework for gene discovery and functional investigation of itHMs/it in other itSolanaceae/it species./p [本文引用: 1] |
| [20] | DOI:10.1038/ng.144PMID:18469814URLPlant domestication represents an accelerated form of evolution, resulting in exaggerated changes in the tissues and organs of greatest interest to humans (for example, seeds, roots and tubers). One of the most extreme cases has been the evolution of tomato fruit. Cultivated tomato plants produce fruit as much as 1,000 times larger than those of their wild progenitors. Quantitative trait mapping studies have shown that a relatively small number of genes were involved in this dramatic transition, and these genes control two processes: cell cycle and organ number determination. The key gene in the first process has been isolated and corresponds to fw2.2, a negative regulator of cell division. However, until now, nothing was known about the molecular basis of the second process. Here, we show that the second major step in the evolution of extreme fruit size was the result of a regulatory change of a YABBY-like transcription factor (fasciated) that controls carpel number during flower and/or fruit development. [本文引用: 4] |
| [21] | DOI:10.1111/pbi.12268PMID:25283700URLSummary Tomato is one of the most cultivated vegetables in the world and an important ingredient of the human diet. Tomato breeders and growers face a continuous challenge of combining high quantity (production volume) with high quality (appearance, taste and perception for the consumers, processing quality for the processing industry). To improve the quality of tomato, it is important to understand the regulation of fruit development and of fruit cellular structure, which is in part determined by the sizes and numbers of cells within a tissue. The role of the cell cycle therein is poorly understood. Plant cyclin-dependent kinases (CDKs) are homologues of yeast cdc2, an important cell cycle regulator conserved throughout all eukaryotes. CDKA1 is constitutively expressed during the cell cycle and has dual functions in S- and M-phase progression. We have produced transgenic tomato plants with increased expression of CDKA1 under the control of the fruit-specific TPRP promoter, which despite a reduced number of seeds and diminished amount of jelly, developed fruits with weight and shape comparable to that of wild-type fruits. However, the phenotypic changes with regard to the pericarp thickness and placenta area were remarkable. Fruits of tomato plants with the highest expression of CDKA1 had larger septa and columella (placenta), compared with wild-type fruits. Our data demonstrate the possibility of manipulating the ratio between cell division and expansion by changing the expression of a key cell cycle regulator and probably its activity with substantial effects on structural traits of the harvested fruit. [本文引用: 2] |
| [22] | DOI:10.1093/jxb/err451PMID:3346228URLGrowth of tomato fruits is determined by cell division and cell expansion, which are tightly controlled by factors that drive the core cell cycle. The cyclin-dependent kinases (CDKs) and their interacting partners, the cyclins, play a key role in the progression of the cell cycle. In this study the role of CDKA1, CDKB1, and CDKB2 in fruit development was characterized by fruit-specific overexpression and down-regulation. CDKA1 is expressed in the pericarp throughout development, but is strongly up-regulated in the outer pericarp cell layers at the end of the growth period, when CDKB gene expression has ceased. Overexpression of the CDKB genes at later stages of development and the down-regulation of CDKA1 result in a very similar fruit phenotype, showing a reduction in the number of cell layers in the pericarp and alterations in the desiccation of the fruits. Expression studies revealed that CDKA1 is down-regulated by the expression of CDKB1/2 in CDKB1 and CDKB2 overexpression mutants, suggesting opposite roles for these types of CDK proteins in tomato pericarp development. [本文引用: 1] |
| [23] | DOI:10.1104/pp.87.3.731PMID:16666216URLAnalysis of [3H]-(fructosyl)-sucrose translocation in tomato (Lycopersicon esculentum Mill.) indicates that phloem unloading in the fruit occurs, at least in part, to the apoplast followed by extracellular hydrolysis. Apoplastic sucrose, glucose, and fructose concentrations were estimated as 1 to 7, 12 to 49, and 8 to 63 millimolar, respectively in the tomato fruit pericarp tissue. Hexose concentrations were at least fourfold greater than sucrose at all developmental stages. Short-term uptake of [14C]sucrose, -glucose, and -fructose in tomato pericarp disks showed first order kinetics over the physiologically relevant concentration range. The uptake rate of [14C]-(glucosyl)-1-fluorosucrose was identical to the rate of [14C]sucrose uptake, suggesting sucrose may be taken up directly without prior extracellular hydrolysis. Short-term uptake of all three sugars was insensitive to 10 micromolar carbonyl cyanide m-chlorophenylhydrazone and to 10 micromolar p-chloromercuribenzene sulfonic acid. However, long-term accumulation of glucose was sensitive to carbonyl cyanide m-chlorophenylhydrazone. Together these results suggest that although sucrose is at least partially hydrolyzed in the apoplast, sucrose may enter the metabolic carbohydrate pool directly. In addition, sugar uptake across the plasma membrane does not appear to be energy dependent, suggesting that sugar accumulation in the tomato fruit is driven by subsequent intracellular metabolism and/or active uptake at the tonoplast. [本文引用: 1] |
| [24] | DOI:10.1093/jxb/erq293PMID:20937732URL[EN] Transgenic tomato plants (Solanum lycopersicum L.) with reduced mRNA levels of AUXIN RESPONSE FACTOR 7(SlARF7) form parthenocarpic fruits with morphological characteristics that seem to be the result of both increasedauxin and gibberellin (GA) responses during fruit growth. This paper presents a more detailed analysis of thesetransgenic lines. Gene expression analysis of auxin-responsive genes show that SlARF7 may regulate only part ofthe auxin signalling pathway involved in tomato fruit set and development. Also, part of the GA signalling pathwaywas affected by the reduced levels of SlARF7 mRNA, as morphological and molecular analyses display similaritiesbetween GA-induced fruits and fruits formed by the RNAi SlARF7 lines. Nevertheless, the levels of GAs were stronglyreduced compared with that in seeded fruits. These findings indicate that SlARF7 acts as a modifier of both auxinand gibberellin responses during tomato fruit set and development. |
| [25] | DOI:10.1093/jxb/erv152PMID:4449553URLAbstract The transformation of the ovary into a fruit after successful completion of pollination and fertilization has been associated with many changes at transcriptomic level. These changes are part of a dynamic and complex regulatory network that is controlled by phytohormones, with a major role for auxin. One of the auxin-related genes differentially expressed upon fruit set and early fruit development in tomato is Solanum lycopersicum AUXIN RESPONSE FACTOR 9 (SlARF9). Here, the functional analysis of this ARF is described. SlARF9 expression was found to be auxin-responsive and SlARF9 mRNA levels were high in the ovules, placenta, and pericarp of pollinated ovaries, but also in other plant tissues with high cell division activity, such as the axillary meristems and root meristems. Transgenic plants with increased SlARF9 mRNA levels formed fruits that were smaller than wild-type fruits because of reduced cell division activity, whereas transgenic lines in which SlARF9 mRNA levels were reduced showed the opposite phenotype. The expression analysis, together with the phenotype of the transgenic lines, suggests that, in tomato, ARF9 negatively controls cell division during early fruit development. The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology. [本文引用: 1] |
| [26] | DOI:10.1073/pnas.1002720107PMID:20395551URLCytosine DNA methylation is a heritable epigenetic mark present in many eukaryotic organisms. Although DNA methylation likely has a conserved role in gene silencing, the levels and patterns of DNA methylation appear to vary drastically among different organisms. Here we used shotgun genomic bisulfite sequencing (Bs-Seq) to compare DNA methylation in eight diverse plant and animal genomes. We found that patterns of methylation are very similar in flowering plants with methylated cytosines detected in all sequence contexts, whereas CG methylation predominates in animals. Vertebrates have methylation throughout the genome except for CpG islands. Gene body methylation is conserved with clear preference for exons in most organisms. Furthermore, genes appear to be the major target of methylation in Ciona and honey bee. Among the eight organisms, the green alga Chlamydomonas has the most unusual pattern of methylation, having non-CG methylation enriched in exons of genes rather than in repeats and transposons. In addition, the Dnmt1 cofactor Uhrf1 has a conserved function in maintaining CG methylation in both transposons and gene bodies in the mouse, Arabidopsis, and zebrafish genomes. [本文引用: 1] |
| [27] | DOI:10.1104/pp.122.2.471URLWe investigated the role of gibberellins (GAs) in the effect of pat-2, a recessive mutation that induces facultative parthenocarpic fruit development in tomato (Lycopersicon esculentum Mill.) using near-isogenic lines with two different genetic backgrounds. Unpollinated wild-type Madrigal (MA/wt) and Cuarenteno (CU/wt) ovaries degenerated, but GA3application induced parthenocarpic fruit growth. On the contrary, parthenocarpic growth of MA/pat-2 and CU/pat-2 fruits, which occurs in the absence of pollination and hormone application, was not affected by GA3. Pollinated MA/wt and parthenocarpic MA/pat-2 ovary development was negated by paclobutrazol, and this inhibitory effect was counteracted by GA3. The main GAs of the early-13-hydroxylation pathway ($\text{GA}_{1}$, GA3,$\text{GA}_{8}$,$\text{GA}_{19}$,$\text{GA}_{20}$,$\text{GA}_{29}$,$\text{GA}_{44}$,$\text{GA}_{53}$, and, tentatively,$\text{GA}_{81}$) and two GAs of the non-13-hydroxylation pathway (GA9and$\text{GA}_{34}$) were identified in MA/wt ovaries by gas chromatography-selected ion monitoring. GAs were quantified in unpollinated ovaries at flower bud, pre-anthesis, and anthesis. In unpollinated MA/pat-2 and CU/pat-2 ovaries, the$\text{GA}_{20}$content was much higher (up to 160 times higher) and the$\text{GA}_{19}$content was lower than in the corresponding non-parthenocarpic ovaries. The application of an inhibitor of 2-oxoglutarate-dependent dioxygenases suggested that$\text{GA}_{20}$is not active per se. The pat-2 mutation may increase GA 20-oxidase activity in unpollinated ovaries, leading to a higher synthesis of$\text{GA}_{20}$, the precursor of an active GA. [本文引用: 1] |
| [28] | DOI:10.1126/science.289.5476.85PMID:10884229URLDomestication of many plants has correlated with dramatic increases in fruit size. In tomato, one quantitative trait locus (QTL), fw2.2, was responsible for a large step in this process. When transformed into large-fruited cultivars, a cosmid derived from the fw2.2 region of a small-fruited wild species reduced fruit size by the predicted amount and had the gene action expected for fw2.2. The cause of the QTL effect is a single gene, ORFX, that is expressed early in floral development, controls carpel cell number, and has a sequence suggesting structural similarity to the human oncogene c-H-ras p21. Alterations in fruit size, imparted by fw2.2 alleles, are most likely due to changes in regulation rather than in the sequence and structure of the encoded protein. [本文引用: 1] |
| [29] | DOI:10.1111/j.1365-3040.2004.01167.xPMID:15215509URLJasmonic acid (JA) is known to be involved in the response of plants to environmental stresses such as drought, and betaine (glycinebetaine) is an osmopretectant accumulated in plants under environmental stresses including drought. However, it remains currently unclear whether JA is involved in the water-stress-induced betaine accumulation in plant leaves. The present experiment, performed with the whole pear plant ( Pyrus bretschneideri Redh. cv. Suli), revealed that the exogenously applied JA induced a significant increase of the betaine level in the pear leaves when the plants were not yet stressed by drought, and when the plants were subjected to water stress, the 'JA plus drought' treatment induced a significant higher betaine level than did the drought treatment alone. Meanwhile, the 'JA plus drought' treatment induced higher levels of betaine aldehyde dehydrogenase (BADH, E C 1.2.1.8) and activities in the leaves than did the drought treatment alone. These results obtained in the whole plant experiments were supported by the results of detached leaf experiments. In detached leaves JA induced significant increases in betaine levels, BADH activities and BADH protein amounts in a time- and concentration-dependent manner. These data demonstrate that JA is involved in the drought-induced betaine accumulation in pear leaves. [本文引用: 1] |
| [30] | DOI:10.1105/tpc.5.10.1439URL [本文引用: 3] |
| [31] | DOI:10.1105/tpc.019158PMID:15010516URLThe LeMADS-RIN gene itself is induced at the onset of ripening without substantial influence by ethylene thus indicating higher order regulatory control. Comparative gene expression analysis in rin and nor fruit suggests an interesting sub-class of ethylene responsive genes, including E8, which respond to developmental signals and ethylene in rin but not nor fruit. E8 expression is induced to approximately 30% of maximal ripening levels in mature green rin fruit at a time consistent with the onset of ripening and attains normal expression in response to exogenous ethylene. The bi-modal regulation of E8 expression in rin and the absence of expression in nor, combined with the inability to induce ripening in either mutant via ethylene, defines a minimal regulatory network during ripening. In this network, ethylene regulates a subset of ripening genes either directly or in concert with developmental signals influenced by LeMADS-RIN and/or the nor gene product. E8 represents a regulatory motif in which nor but not LeMADS-RIN provides developmental control. In this instance, LeMADS-RIN impacts E8 gene expression mainly via activation of autocatalytic ethylene synthesis. Future characterization of ripening gene expression in nor will facilitate further definition of developmental regulation during ripening which is clearly impacted by both mutations in ways that do not overlap. The nor locus has been cloned and encodes a putative transcription factor with no relationship to MADS-box genes. The availability of this sequence should promote refinement of the developmental regulatory network governing fruit ripening. [本文引用: 1] |
| [32] | DOI:10.1111/j.1365-313X.2007.03167.xPMID:17587306URLSummary Top of page Summary Introduction Results Discussion Experimental procedures Acknowledgements References Tomato fruit size results from the combination of cell number and cell size which are respectively determined by cell division and cell expansion processes. As fruit growth is mainly sustained by cell expansion, the development of pericarp and locular tissues is characterized by the concomitant arrest of mitotic activity, inhibition of cyclin-dependent kinase (CDK) activity, and numerous rounds of endoreduplication inducing a spectacular increase in DNA ploidy and mean cell size. To decipher the molecular basis of the endoreduplication-associated cell growth in fruit, we investigated the putative involvement of the WEE1 kinase (Solly;WEE1). We here report a functional analysis of Solly;WEE1 in tomato. Impairing the expression of Solly;WEE1 in transgenic tomato plants resulted in a reduction of plant size and fruit size. In the most altered phenotypes, fruits displayed a reduced number of seeds without embryo development. The reduction of plant-, fruit- and seed size originated from a reduction in cell size which could be correlated with a decrease of the DNA ploidy levels. At the molecular level downregulating Solly;WEE1 in planta resulted in the increase of CDKA activity levels originating from a decrease of the amount of Y15-phosphorylated CDKA, thus indicating a release of the negative regulation on CDK activity exerted by WEE1. Our data indicated that Solly;WEE1 participates in the control of cell size and/or the onset of the endoreduplication process putatively driving cell expansion. [本文引用: 1] |
| [33] | DOI:10.1016/j.plantsci.2011.03.017URL [本文引用: 1] |
| [34] | DOI:10.1016/j.tplants.2012.05.001URL [本文引用: 1] |
| [35] | DOI:10.2307/3869688PMID:8305869URLA key regulator of the cell cycle is a highly conserved protein kinase whose catalytic subunit, p34 cdc2, is encoded by the cdc2 gene. We studied the control of the expression of the Arabidopsis cdc2a gene in cell suspensions and during plant development. In cell cultures, arrest of the cell cycle did not significantly affect cdc2a mRNA levels, but nutrient conditions were important for cdc2a expression. During plant development, the pattern of cdc2a expression was strongly correlated with the cell proliferation potential. The effects of external signals on cdc2a expression were analyzed. Wounding induced expression in leaves. Lack of light altered temporal regulation of cdc2a in the apical but not root meristem of seedlings. Differential cdc2a responses were obtained after different hormone treatments. Signals present only in intact plants were necessary to mediate these responses. Although other control levels have yet to be analyzed, these results suggest that the regulation of cdc2a expression may contribute greatly to spatial and temporal regulation of cell division in plants. Our results also show that cdc2a expression is not always coupled with cell proliferation but always precedes it. We propose that cdc2a expression may reflect a state of competence to divide, and that the release of other controls is necessary for cell division to occur. [本文引用: 1] |
| [36] | DOI:10.1093/jxb/47.Special_Issue.1239PMID:21245255URLThe limitation of tomato yield was investigated in a number of cultivars with contrasting fruiting habits. Unless light is limiting, yield is mainly restricted by the number or the size of the fruit (i.e. the sink strength) rather than the supply of assimilate (i.e. the source strength). Fruit size is determined by both cell number and cell size. The rate of fruit expansion is affected by assimilate supply, temperature and water relations in the plant. The size or the growth rate of a tomato fruit is regulated by the import of assimilate and water. The sink strength for assimilate of a tomato fruit measured by the rate of assimilate import may be related to the routes of sugar transport into the sink cells during fruit development. Enzymic regulation of the hydrolysis of sucrose by sucrose synthase and the accumulation of starch by ADPG pyrophosphorylase may determine the rate of assimilate import in the young fruit. Vacuolar invertase activity may determine the sugar composition of a mature fruit, but may not affect the overall dry matter accumulation of a tomato fruit. While yield is determined by the balance between source and sink strengths of the plant, quality is determined by the transport and metabolism of sugars within the fruit. [本文引用: 1] |
| [37] | [本文引用: 1] |
| [38] | [本文引用: 1] |
| [39] | DOI:10.1105/tpc.108.063719PMID:19574437URLInvertase plays multiple pivotal roles in plant development. Thus, its activity must be tightly regulated in vivo. Emerging evidence suggests that a group of small proteins that inhibit invertase activity in vitro appears to exist in a wide variety of plants. However, little is known regarding their roles in planta. Here, we examined the function of INVINH1, a putative invertase inhibitor, in tomato [Solanum lycopersicum). Expression of a INVINH1: green fluorescent protein fusion revealed its apoplasmic localization. Ectopic overexpression of INVINH1 in Arabidopsis thaliana specifically reduced cell wall invertase activity. By contrast, silencing its expression in tomato significantly increased the activity of cell wall invertase without altering activities of cytoplasmic and vacuolar invertases. Elevation of cell wall invertase activity in RNA interference transgenic tomato led to (1) a prolonged leaf life span involving in a blockage of abscisic acid-induced senescence and (2) an increase in seed weight and fruit hexose level, which is likely achieved through enhanced sucrose hydrolysis in the apoplasm of the fruit vasculature. This assertion is based on (1) coexpression of INVINH1 and a fruit-specific cell wall invertase Lin5 in phloem parenchyma cells of young fruit, including the placenta regions connecting developing seeds; (2) a physical interaction between INVINH1 and Lin5 in vivo; and (3) a symplasmic discontinuity at the interface between placenta and seeds. Together, the results demonstrate that INVINH1 encodes a protein that specifically inhibits the activity of cell wall invertase and regulates leaf senescence and seed and fruit development in tomato by limiting the invertase activity in planta. [本文引用: 1] |
| [40] | DOI:10.1371/journal.pgen.1000530PMID:2696037URLLoss or gain of DNA methylation can affect gene expression and is sometimes transmitted across generations. Such epigenetic alterations are thus a possible source of heritable phenotypic variation in the absence of DNA sequence change. However, attempts to assess the prevalence of stable epigenetic variation in natural and experimental populations and to quantify its impact on complex traits have been hampered by the confounding effects of DNA sequence polymorphisms. To overcome this problem as much as possible, two parents with little DNA sequence differences, but contrasting DNA methylation profiles, were used to derive a panel of epigenetic Recombinant Inbred Lines (epiRILs) in the reference plant Arabidopsis thaliana. The epiRILs showed variation and high heritability for flowering time and plant height ( approximately 30%), as well as stable inheritance of multiple parental DNA methylation variants (epialleles) over at least eight generations. These findings provide a first rationale to identify epiallelic variants that contribute to heritable variation in complex traits using linkage or association studies. More generally, the demonstration that numerous epialleles across the genome can be stable over many generations in the absence of selection or extensive DNA sequence variation highlights the need to integrate epigenetic information into population genetics studies. [本文引用: 1] |
| [41] | [本文引用: 1] |
| [42] | DOI:10.1046/j.1365-2923.2000.00553.xPMID:10557234URLFollowing fruit set, the early development of tomato (Lycopersicon esculentum Mill.) fruit comprises two distinct phases: a cell division phase and a consecutive phase of cell expansion until the onset of ripening. In this study, we analyzed cytological and molecular changes characterizing these early phases of tomato fruit development. First we investigated the spatial and temporal regulation of the mitotic activity during fruit development. The DNA content of isolated nuclei from the different fruit tissues was determined by flow cytometry analysis. The results confirm the data of mitotic activity measurements and show that cell differentiation, leading to expanded cells, is characterized by endoreduplication. Second, we isolated two cDNAs, named Lyces;CDKA1 (accession no. Y17225) and Lyces;CDKA2 (accession no. Y17226), encoding tomato homologs of the cyclin-dependent kinase (CDK) p34cdc2. Tomato CDKA gene expression was followed at both the transcriptional and translational levels during fruit development. The transcripts for Lyces;CDKA1 and Lyces;CDKA2 and the corresponding CDKA proteins are predominantly accumulated during the phase of cell division between anthesis and 5 d post anthesis (DPA). In whole fruits, the maximum CDK activity was obtained between 5 and 10 DPA. The determination of the kinase activity using protein extracts from the different fruit tissues was in agreement with mitotic activity analysis. It showed the particular disappearance of the activity in the gel tissue as early as 15 DPA. The overall data of CDK activity measurements suggest a strong post-translational regulation of CDK at the temporal and spatial levels during early tomato fruit development. [本文引用: 1] |
| [43] | [本文引用: 1] |
| [44] | DOI:10.1146/annurev-genet-110410-132507PMID:22060040URLTomato ripening is a highly coordinated developmental process that coincides with seed maturation. Regulated expression of thousands of genes controls fruit softening as well as accumulation of pigments, sugars, acids, and volatile compounds that increase attraction to animals. A combination of molecular tools and ripening-affected mutants has permitted researchers to establish a framework for the control of ripening. Tomato is a climacteric fruit, with an absolute requirement for the phytohormone ethylene to ripen. This dependence upon ethylene has established tomato fruit ripening as a model system for study of regulation of its synthesis and perception. In addition, several important ripening mutants, including rin, nor, and Cnr, have provided novel insights into the control of ripening processes. Here, we describe how ethylene and the transcription factors associated with the ripening process fit together into a network controlling ripening. [本文引用: 3] |
| [45] | DOI:10.1093/jxb/eru277PMID:25028558URLAbstract Plant hormones have been extensively studied for their roles in the regulation of various aspects of plant development. However, in the last decade important new insights have been made into their action during development and ripening, in both dry and fleshy fruits. Emerging evidence suggests that relative functions of plant hormones are not restricted to a particular stage, and a complex network of more than one plant hormone is involved in controlling various aspects of fruit development. Though some areas are extensively covered, considerable gaps in our knowledge and understanding still exist in the control of hormonal networks and crosstalk between different hormones during fruit expansion, maturation, and various other aspects of ripening. Here, we evaluate the new knowledge on their relative roles during tomato fruit development with a view to understand their mechanism of action in fleshy fruits. For a better understanding, pertinent evidences available on hormonal crosstalk during fruit development in other species are also discussed. We envisage that such detailed knowledge will help design new strategies for effective manipulation of fruit ripening. The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com. [本文引用: 4] |
| [46] | DOI:10.1016/j.bbagrm.2011.03.002PMID:21414429URL78 Mechanisms modulating chromatin structure control gene regulation. 78 Distinct epigenetic modifications show specific correlations with transcription. 78 Transcription factors and chromatin modifiers interact to control gene expression. 78 Repeats regulate expression of adjacent genes through epigenetic marks diffusion. 78 Specific epigenetic regulation may be associated with plant life cycle features. |
| [47] | DOI:10.1038/nrg2719URL [本文引用: 1] |
| [48] | DOI:10.1105/tpc.022178URL [本文引用: 1] |
| [49] | DOI:10.1016/j.cell.2007.01.015PMID:17320508URLChromatin structure imposes significant obstacles on all aspects of transcription that are mediated by RNA polymerase II. The dynamics of chromatin structure are tightly regulated through multiple mechanisms including histone modification, chromatin remodeling, histone variant incorporation, and histone eviction. In this Review, we highlight advances in our understanding of chromatin regulation and discuss how such regulation affects the binding of transcription factors as well as the initiation and elongation steps of transcription. [本文引用: 1] |
| [50] | DOI:10.1007/BF00040664PMID:1371404URLRegulation of wound-inducible 1-aminocyclopropane-1-carboxylic acid (ACC) synthase expression was studied in tomato fruit ( Lycopersicon esculentum cv. Pik-Red). A 70 base oligonucleotide probe homologous to published ACC synthase cDNA sequences was successfully used to identify and analyze regulation of a wound-inducible transcript. The 1.8 kb ACC synthase transcript increased upon wounding the fruit as well as during fruit ripening. Salicylic acid, an inhibitor of wound-responsive genes in tomato, inhibited the wound-induced accumulation of the ACC synthase transcript. Further, polyamines (putrescine, spermidine and spermine) that have anti-senescence properties and have been shown to inhibit the development of ACC synthase activity, inhibited the accumulation of the wound-inducible ACC synthase transcript. The inhibition by spermine was greater than that caused by putrescine or spermidine. The transcript level of a wound-repressible glycine-rich protein gene and that of the constitutively expressed rRNA were not affected as markedly by either salicylic acid or polyamines. These data suggest that salicylic acid and polyamines may specifically regulate ethylene biosynthesis at the level of ACC synthase transcript accumulation. [本文引用: 1] |
| [51] | DOI:10.1038/ng.3117PMID:25305757URLThe histories of crop domestication and breeding are recorded in genomes. Although tomato is a model species for plant biology and breeding, the nature of human selection that altered its genome remains largely unknown. Here we report a comprehensive analysis of tomato evolution based on the genome sequences of 360 accessions. We provide evidence that domestication and improvement focused on two independent sets of quantitative trait loci (QTLs), resulting in modern tomato fruit 090304100 times larger than its ancestor. Furthermore, we discovered a major genomic signature for modern processing tomatoes, identified the causative variants that confer pink fruit color and precisely visualized the linkage drag associated with wild introgressions. This study outlines the accomplishments as well as the costs of historical selection and provides molecular insights toward further improvement. [本文引用: 1] |
| [52] | DOI:10.1016/j.gde.2007.07.007PMID:17723293URLResolving natural phenotypic variation into genetic and molecular components is a major objective in biology. Over the past decade, tomato interspecific introgression lines (ILs), each carrying a single ‘exotic’ chromosome segment from a wild species, have exposed thousands of quantitative trait loci (QTL) affecting plant adaptation, morphology, yield, metabolism, and gene expression. QTL for fruit size and sugar composition were isolated by map-based cloning, while others were successfully implemented in marker-assisted breeding programs. More recently, integrating the multitude of IL-QTL into a single database has unraveled some unifying principles about the architecture of complex traits in plants. [本文引用: 1] |
| [53] | DOI:10.1073/pnas.162485999PMID:12242331URLA common, recurring theme in domesticated plants is the occurrence of pear-shaped fruit. A major quantitative trait locus (termed ovate) controlling the transition from round to pear-shaped fruit has been cloned from tomato. OVATE is expressed early in flower and fruit development and encodes a previously uncharacterized, hydrophilic protein with a putative bipartite nuclear localization signal, Von Willebrand factor type C domains, and an 鈮70-aa C-terminal domain conserved in tomato, Arabidopsis, and rice. A single mutation, leading to a premature stop codon, causes the transition of tomato fruit from round- to pear-shaped. Moreover, ectopic, transgenic expression of OVATE unevenly reduces the size of floral organs and leaflets, suggesting that OVATE represents a previously uncharacterized class of negative regulatory proteins important in plant development. [本文引用: 1] |
| [54] | DOI:10.1104/pp.16.00959PMID:27462084URLReduced cell wall invertase (CWIN) activity has been shown to be associated with poor seed and fruit set under abiotic stress. Here, we examined whether genetically increasing native CWIN activity would sustain fruit set under long-term moderate heat stress (LMHS), an important factor limiting crop production, by using transgenic tomato (Solanum lycopersicum) with its CWIN inhibitor gene silenced and focusing on ovaries and fruits at 2 d before and after pollination, respectively. We found that the increase of CWIN activity suppressed LMHS-induced programmed cell death in fruits. Surprisingly, measurement of the contents of H2O2 and malondialdehyde and the activities of a cohort of antioxidant enzymes revealed that the CWIN-mediated inhibition on programmed cell death is exerted in a reactive oxygen species-independent manner. Elevation of CWIN activity sustained Suc import into fruits and increased activities of hexokinase and fructokinase in the ovaries in response to LMHS. Compared to the wild type, the CWIN-elevated transgenic plants exhibited higher transcript levels of heat shock protein genes Hsp90 and Hsp100 in ovaries and HspII17.6 in fruits under LMHS, which corresponded to a lower transcript level of a negative auxin responsive factor IAA9 but a higher expression of the auxin biosynthesis gene ToFZY6 in fruits at 2 d after pollination. Collectively, the data indicate that CWIN enhances fruit set under LMHS through suppression of programmed cell death in a reactive oxygen species-independent manner that could involve enhanced Suc import and catabolism, HSP expression, and auxin response and biosynthesis. [本文引用: 2] |
| [55] | DOI:10.1007/s00344-011-9204-1URL [本文引用: 1] |
| [56] | [本文引用: 1] |
| [57] | DOI:10.1093/jxb/ers207PMID:3444270URLCytokinins (CKs) are thought to play important roles in fruit development, especially cell division. However, the mechanisms and regulation of CK activity have not been well investigated. This study analysed CK concentrations and expression of genes involved in CK metabolism in developing tomato (Solanum lycopersicum) ovaries. The concentrations of CK ribosides and isopentenyladenine and the transcript levels of the CK biosynthetic genes SlIPT3, SlIPT4, SlLOG6, and SlLOG8were high at anthesis and decreased immediately afterward. In contrast,trans-zeatin concentration and the transcript levels of the CK biosynthetic genes SlIPT1, SlIPT2, SlCYP735A1, SlCYP735A2, and SlLOG2increased after anthesis. The expression of type-A response regulator genes was high in tomato ovaries from pre-anthesis to early post-anthesis stages. These results suggest that the CK signal transduction pathway is active in the cell division phase of fruit development. This study also investigated the effect of CK application on fruit set and development. Application of a synthetic CK,N-(2-chloro-pyridin-4-yl)-N-phenylurea (CPPU), to unpollinated tomato ovaries induced parthenocarpic fruit development. The CPPU-induced parthenocarpic fruits were smaller than pollinated fruits, because of reduction of pericarp cell size rather than reduced cell number. Thus, CPPU-induced parthenocarpy was attributable to the promotion of cell division, not cell expansion. Overall, the results provide evidence that CKs are involved in cell division during development of tomato fruit. [本文引用: 2] |
| [58] | DOI:10.3389/fpls.2013.00079PMID:3628358URLPlant species that bear fruit often utilize expansion of an ovary (carpel) or accessory tissue as a vehicle for seed dispersal. While the seed(s) develop, the tissue(s) of the fruit follow a common progression of cell division and cell expansion, promoting growth of the fruit. Once the seed is fully developed, the fruit matures and the surrounding tissue either dries or ripens promoting the dissemination of the seed. As with many developmental processes in plants, plant hormones play an important role in the synchronization of signals between the developing seed and its surrounding fruit tissue(s), regulating each phase of fruit development. Following pollination, fruit set is achieved through a de-repression of growth and an activation of cell divisionviathe action of auxin and/or cytokinin and/or gibberellin. Following fruit set, growth of the fruit is facilitated through a relatively poorly studied period of cell expansion and endoreduplication that is likely regulated by similar hormones as in fruit set. Once the seeds reach maturity, fruit become ready to undergo ripening and during this period there is a major switch in relative hormone levels of the fruit, involving an overall decrease in auxin, gibberellin, and cytokinin and a simultaneous increase in abscisic acid and ethylene. While the role of hormones in fruit set and ripening is well documented, the knowledge of the roles of other hormones during growth, maturation, and some individual ripening components is sketchy. [本文引用: 1] |
| [59] | DOI:10.1093/mp/ssq050PMID:20833733URLHexoses accumulate to high concentrations (6520065mM) in storage parenchyma cells of tomato fruit. Hexoses are sourced from the fruit apoplasm as hydrolysis products of phloem-imported sucrose. Three hexose transporters (LeHT1, LeHT2, LeHT3), expressed in fruit storage parenchyma cells, may contribute to hexose uptake by these cells. An analysis of their full-length sequences demonstrated that all three transporters belong to the STP sub-family of monosaccharide transporters that localize to plasma membranes. Heterologous expression of LeHT1 (and previously LeHT2, Gear et al., 2000), but not LeHT3, rescued a hexose transport-impaired yeast mutant when raised on glucose or fructose as the sole carbon source. Biochemically, LeHT1, similarly to LeHT2, exhibited transport properties consistent with a high-affinity glucose/H+ symporter. Significantly, LeHT1 and LeHT2 also functioned as low-affinity fructose/H+ symporters with apparent Km values commensurate with those of fruit tissues. A substantial reduction (80–90%) in fruit expression levels of all LeHT genes by RNAi-mediated knockdown caused a 55% decrease in fruit hexose accumulation. In contrast, photoassimilate production by source leaves and phloem transport capacity to fruit were unaffected by transporter knockdown. Collectively, these findings demonstrate that LeHTs play key roles in driving accumulation of hexoses into storage parenchyma cells during tomato fruit development. [本文引用: 1] |
| [60] | DOI:10.1038/nbt0602-613PMID:12042867URLAbstract Polyamines, ubiquitous organic aliphatic cations, have been implicated in a myriad of physiological and developmental processes in many organisms, but their in vivo functions remain to be determined. We expressed a yeast S-adenosylmethionine decarboxylase gene (ySAMdc; Spe2) fused with a ripening-inducible E8 promoter to specifically increase levels of the polyamines spermidine and spermine in tomato fruit during ripening. Independent transgenic plants and their segregating lines were evaluated after cultivation in the greenhouse and in the field for five successive generations. The enhanced expression of the ySAMdc gene resulted in increased conversion of putrescine into higher polyamines and thus to ripening-specific accumulation of spermidine and spermine. This led to an increase in lycopene, prolonged vine life, and enhanced fruit juice quality. Lycopene levels in cultivated tomatoes are generally low, and increasing them in the fruit enhances its nutrient value. Furthermore, the rates of ethylene production in the transgenic tomato fruit were consistently higher than those in the nontransgenic control fruit. These data show that polyamine and ethylene biosynthesis pathways can act simultaneously in ripening tomato fruit. Taken together, these results provide the first direct evidence for a physiological role of polyamines and demonstrate an approach to improving nutritional quality, juice quality, and vine life of tomato fruit. [本文引用: 1] |
| [61] | DOI:10.1016/j.pbi.2008.09.005PMID:2644822URLMany of the molecular details regarding the promotion of flowering in response to prolonged exposure to cold temperatures (vernalization) and daylength have recently been elucidated in Arabidopsis. The daylength and vernalization pathway converge in the regulation of floral promoters referred to as floral integrators. In the meristem, vernalization promotes flowering through the epigenetic repression of the floral repressor FLOWERING LOCUS C. This allows for the induction of floral integrators by CONSTANS under inductive long days. In the vasculature of leaves, CONSTANS protein is produced only in long days where it acts to promote the expression of FLOWERING LOCUS T ( FT). FT protein is then translocated to the meristem where it acts to promote floral induction. Thus a detailed molecular framework for the regulation of flowering time has now been established in Arabidopsis. [本文引用: 1] |
| [62] | DOI:10.1016/j.pbi.2011.03.004PMID:21450514URLHighlights? Epigenetic regulation buffers phenotypic variation. ? Destabilization of epigenetic regulation leads to novel epialleles and transposon mobility. ? New epialleles and relocated transposons broaden plant phenotypic variation. ? Epigenetically induced new traits are likely to improve adaptation to environmental stresses. [本文引用: 1] |
| [63] | DOI:10.1371/journal.pgen.1006930PMID:5560543URLIncreases in fruit weight of cultivated vegetables and fruits accompanied the domestication of these crops. Here we report on the positional cloning of a quantitative trait locus (QTL) controlling fruit weight in tomato. The derived allele ofCell Size Regulator(CSR-D) increases fruit weight predominantly through enlargement of the pericarp areas. The expanded pericarp tissues result from increased mesocarp cell size and not from increased number of cell layers. The effect ofCSRon fruit weight and cell size is found across different genetic backgrounds implying a consistent impact of the locus on the trait. In fruits,CSRexpression is undetectable early in development from floral meristems to the rapid cell proliferation stage after anthesis. Expression is low but detectable in growing fruit tissues and in or around vascular bundles coinciding with the cell enlargement stage of the fruit maturation process.CSRencodes an uncharacterized protein whose clade has expanded in the Solanaceae family. The mutant allele is predicted to encode a shorter protein due to a 1.4 kb deletion resulting in a 194 amino-acid truncation. Co-expression analyses and GO term enrichment analyses suggest association of CSR with cell differentiation in fruit tissues and vascular bundles. The derived allele arose inSolanum lycopersicumvarcerasiformeand appears completely fixed in many cultivated tomato market classes. This finding suggests that the selection of this allele was critical to the full domestication of tomato from its intermediate ancestors. Starting about 10,000 years ago, during the Neolithic period, human societies began the transformation from a hunting and gathering-dependent lifestyle to an agrarian lifestyle. This transformation was accompanied by plant and animal domestication. Tomato shows a huge increase in fruit weight that has arisen as a consequence of its domestication. We identified a gene that encodes a poorly characterized protein that controls fruit weight in tomato. The mutation that led to the increase in fruit weight arose early during the cultivation of tomato and is now incorporated in all large tomato varieties. The gene regulates cell size in the fruit and is calledCell Size Regulator. The increases in cell size are proposed to relate to cellular maturation that accompanies fruit growth. [本文引用: 1] |
| [64] | DOI:10.1104/pp.111.173997PMID:21673133URLIn tomato (Solarium lycopersicum) fruit, the number of locules (cavities containing seeds that are derived from carpels) varies from two to up to 10 or more. Locule number affects fruit shape and size and is controlled by several quantitative trait loci (QTLs). The large majority of the phenotypic variation is explained by two of these QTLs, fasciated (fas) and locule number (lc), that interact epistatically with one another. FAS has been cloned, and mutations in the gene are described as key factors leading to the increase in fruit size in modern varieties. Here, we report the map-based cloning of k. The lc QTL includes a 1,600-bp region that is located 1,080 bp from the 3' end of WUSCHEL, which encodes a homeodomain protein that regulates stem cell fate in plants. The molecular evolution of lc showed a reduction of diversity in cultivated accessions with the exception of two single-nucleotide polymorphisms. These two single-nucleotide polymorphisms were shown to be responsible for the increase in locule number. An evolutionary model of locule number is proposed herein, suggesting that the fas mutation appeared after the mutation in the lc locus to confer the extreme high-locule-number phenotype. [本文引用: 3] |
| [65] | DOI:10.1105/tpc.113.118794PMID:24510723URLFruit ripening is the summation of changes rendering fleshy fruit tissues attractive and palatable to seed dispersing organisms. For example, sugar content is influenced by plastid numbers and photosynthetic activity in unripe fruit and later by starch and sugar catabolism during ripening. Tomato fruit are sinks of photosynthate, yet unripe green fruit contribute significantly to the sugars that ultimately accumulate in the ripe fruit. Plastid numbers and chlorophyll content are influenced by numerous environmental and genetic factors and are positively correlated with photosynthesis and photosynthate accumulation. GOLDEN2-LIKE (GLK) transcription factors regulate plastid and chlorophyll levels. Tomato (Solanum lycopersicum), like most plants, contains two GLKs (i.e., GLK1 and GLK2/UNIFORM). Mutant and transgene analysis demonstrated that these genes encode functionally similar peptides, though differential expression renders GLK1 more important in leaves, while GLK2 is predominant in fruit. A latitudinal gradient of GLK2 expression influences the typical uneven coloration of green and ripe wild-type fruit. Transcriptome profiling revealed a broader fruit gene expression gradient throughout development. The gradient influenced general ripening activities beyond plastid development and was consistent with the easily observed yet poorly studied ripening gradient present in tomato and many fleshy fruits. [本文引用: 1] |
| [66] | DOI:10.1016/j.jplph.2012.02.004PMID:22424572URLAbscisic acid (ABA) deficient mutants, such as notabilis and flacca, have helped elucidating the role of ABA during plant development and stress responses in tomato (Solanum lycopersicum L.). However, these mutants have only moderately decreased ABA levels. Here we report on plant and fruit development in the more strongly ABA-deficient notabilis/flacca (not/flc) double mutant. We observed that plant growth, leaf-surface area, drought-induced wilting and ABA-related gene expression in the different genotypes were strongly correlated with the ABA levels and thus most strongly affected in the not/flc double mutants. These mutants also had reduced fruit size that was caused by an overall smaller cell size. Lower ABA levels in fruits did not correlate with changes in auxin levels, but were accompanied by higher ethylene evolution rates. This suggests that in a wild-type background ABA stimulates cell enlargement during tomato fruit growth via a negative effect on ethylene synthesis. [本文引用: 1] |
| [67] | [本文引用: 1] |
| [68] | DOI:10.1007/s00344-003-0024-9URL [本文引用: 1] |
| [69] | DOI:10.1104/pp.112.212654PMID:23292788URLCarotenoids represent some of the most important secondary metabolites in the human diet, and tomato (Solanum lycopersicum) is a rich source of these health-promoting compounds. In this work, a novel and fruit-related regulator of pigment accumulation in tomato has been identified by artificial neural network inference analysis and its function validated in transgenic plants. A tomato fruit gene regulatory network was generated using artificial neural network inference analysis and transcription factor gene expression profiles derived from fruits sampled at various points during development and ripening. One of the transcription factor gene expression profiles with a sequence related to an Arabidopsis (Arabidopsis thaliana) ARABIDOPSIS PSEUDO RESPONSE REGULATOR2-LIKE gene (APRR2-Like) was up-regulated at the breaker stage in wild-type tomato fruits and, when overexpressed in transgenic lines, increased plastid number, area, and pigment content, enhancing the levels of chlorophyll in immature unripe fruits and carotenoids in red ripe fruits. Analysis of the transcriptome of transgenic lines overexpressing the tomato APPR2-Like gene revealed up-regulation of several ripening-related genes in the overexpression lines, providing a link between the expression of this tomato gene and the ripening process. A putative ortholog of the tomato APPR2-Like gene in sweet pepper (Capsicum annuum) was associated with pigment accumulation in fruit tissues. We conclude that the function of this gene is conserved across taxa and that it encodes a protein that has an important role in ripening. [本文引用: 1] |
| [70] | DOI:10.1111/j.1467-7652.2007.00251.xPMID:17362485URLThe tolerance of various species of plant to abiotic stress has been enhanced by genetic engineering with certain genes. However, the use of such transgenes is often associated with negative effects on growth and productivity under non-stress conditions. The codA gene from Arthrobacter globiformis is of particular interest with respect to the engineering of desirable productive traits in crop plants. The expression of this gene in tomato plants resulted in significantly enlarged flowers and fruits under non-stress conditions. The enlargement of flowers and fruits was associated with high levels of glycinebetaine that accumulated in reproductive organs, such as flower buds and fruits. The enlargement of flowers was related to an increase in the size and number of cells, and reflected the pleiotropic effect of the codA transgene on the expression of genes involved in the regulation of cell division. [本文引用: 1] |
| [71] | DOI:10.1126/science.1222218PMID:22745430URLModern () varieties are bred for uniform ripening (u) light green fruit phenotypes to facilitate harvests of evenly ripened fruit. U encodes a Golden 2-like () transcription factor, SlGLK2, which determines accumulation and distribution in developing fruit. In , two --SlGLK1 and SlGLK2--are expressed in leaves, but only SlGLK2 is expressed in fruit. Expressing increased the content of fruit, whereas SlGLK2 suppression recapitulated the u mutant phenotype. overexpression enhanced fruit and development, leading to elevated carbohydrates and in ripe fruit. SlGLK2 influences in developing fruit, contributing to mature fruit characteristics and suggesting that selection of u inadvertently compromised ripe fruit quality in exchange for desirable production traits. [本文引用: 1] |
| [72] | DOI:10.1016/j.pbi.2005.11.010PMID:16337828URLThe different cell types of a multicellular organism express different sets of genes. Although this is one of the oldest paradigms of developmental genetics, how different patterns of gene expression are established and maintained during subsequent cell division is an active topic of research. Chromatin modifiers play an essential role in controlling gene expression and in establishing epigenetic marks that can be inherited. During the past few years, large number of putative chromatin-associated proteins have been uncovered as controllers of meristem organization and activity, phase transition, and gametophyte and embryo development. [本文引用: 1] |
| [73] | DOI:10.1104/pp.110.167577PMID:21441384Phenotypic diversity within cultivated tomato (Solarium lycopersicum) is particularly evident for fruit shape and size. Four genes that control tomato fruit shape have been cloned. SUN and OVATE control elongated shape whereas FASCIATED (FAS) and LOCULE NUMBER (EC) control fruit locule number and flat shape. We investigated the distribution of the fruit shape alleles in the tomato germplasm and evaluated their contribution to morphology in a diverse collection of 368 predominantly tomato and tomato var. cerasiforme accessions. Fruits were visually classified into eight shape categories that were supported by objective measurements obtained from image analysis using the Tomato Analyzer software. The allele distribution of SUN, OVATE, EC, and FAS in all accessions was strongly associated with fruit shape classification. We also genotyped 116 representative accessions with additional 25 markers distributed evenly across the genome. Through a model-based clustering we demonstrated that shape categories, germplasm classes, and the shape genes were nonrandomly distributed among five genetic clusters (P < 0.001), implying that selection for fruit shape genes was critical to subpopulation differentiation within cultivated tomato. Our data suggested that the EC, FAS, and SUN mutations arose in the same ancestral population while the OVATE mutation arose in a separate lineage. Furthermore, EC, OVATE, and FAS mutations may have arisen prior to domestication or early during the selection of cultivated tomato whereas the SUN mutation appeared to be a postdomestication event arising in Europe. [本文引用: 3] |
| [74] | DOI:10.1146/annurev-arplant-050213-040251PMID:24579990URLAbstract Sucrose metabolism plays pivotal roles in development, stress response, and yield formation, mainly by generating a range of sugars as metabolites to fuel growth and synthesize essential compounds (including protein, cellulose, and starch) and as signals to regulate expression of microRNAs, transcription factors, and other genes and for crosstalk with hormonal, oxidative, and defense signaling. This review aims to capture the most exciting developments in this area by evaluating (a) the roles of key sucrose metabolic enzymes in development, abiotic stress responses, and plant-microbe interactions; (b) the coupling between sucrose metabolism and sugar signaling from extra- to intracellular spaces; (c) the different mechanisms by which sucrose metabolic enzymes could perform their signaling roles; and (d) progress on engineering sugar metabolism and transport for high yield and disease resistance. Finally, the review outlines future directions for research on sugar metabolism and signaling to better understand and improve plant performance. [本文引用: 1] |
| [75] | DOI:10.1007/BF00203641URLThe cellular pathway of postphloem sugar transport was elucidated in the outer pericarp of tomato (Lycopersicon esculentum Mill cv. Floradade) fruit at 13—14 and 23—25 days after anthesis (DAA). These developmental stages are characterized by phloem-imported sugars being accumulated as starch and hexose, respectively. The symplasmic tracer, 5(6)-carboxyfluorescein, loaded into the storage parenchyma cells of pericarp discs, moved readily in the younger fruit but was immobile in fruit at 23—25 DAA. Symplasmic mobility of [14C]glucose was found to be identical to 5(6)-carboxyfluorescein. For the older fruit, the pericarp apoplasm was shown to be freely permeable to the apoplasmic tracer, trisodium 3-hydroxy-5,8,10-pyrenetrisulfonate. Indeed, the transport capacity of the pericarp apoplasm was such that the steady-state rate of in-vitro glucose uptake by pericarp discs accounted fully for the estimated rate of in-vivo glucose accumulation. For fruit at 23—25 DAA, the inhibitory effects of the sulfhydryl group modifier, p-chloromercuribenzenesulfonic acid (PCMBS), on [14C]glucose and [14C]fructose uptake by the pericarp discs depended on the osmolality of the external solution. The inhibition was most pronounced for pericarp discs enriched in storage parenchyma. Consistent with the PCMBS study, strong fluorescent signals were exhibited by the storage parenchyma cells of pericarp discs exposed to the membrane-impermeable thiol-binding fluorochrome, monobromotrimethylammoniobimane. The fluorescent weak acid, sulphorhodamine G, was accumulated preferentially by the storage parenchyma cells. Accumulation of sulphorhodamine G was halted by the ATPase inhibitor erythrosin B, suggesting the presence of a plasma-membrane-bound H+-ATPase. A linkage between the putative H+-ATPase activity and hexose transport was demonstrated by an erythrosin-B inhibition of [14C]glucose and [14C]fructose uptake. In contrast, comparable evidence for an energy-coupled hexose porter could not be found in the pericarp of younger fruit at 13—14 DAA. Overall, the data are interpreted to indicate that: (i) The postphloem cellular pathway in the outer fruit pericarp shifts from the symplasm during starch accumulation (13—14 DAA) to the apoplasm for rapid hexose accumulation (23—25 DAA). (ii) An energy-coupled plasma-membrane hexose carrier is expressed specifically in storage parenchyma cells at the latter stage of fruit development. [本文引用: 1] |
| [76] | DOI:10.1016/j.tplants.2012.06.005PMID:22776090URLSeed and fruit set are established during and soon after fertilization and determine seed and fruit number, their final size and, hence, yield potential. These processes are highly sensitive to biotic and abiotic stresses, which often lead to seed and fruit abortion. Here, we review the regulation of assimilate partitioning, including the potential roles of recently identified sucrose efflux transporters in seed and fruit set and examine the similarities of sucrose import and hydrolysis for both pollen and ovary sinks, and similar causes of abortion. We also discuss the molecular origins of parthenocarpy and the central roles of auxins and gibberellins in fruit set. The recently completed strawberry (Fragaria vesca) and tomato (Solanum lycopersicum) genomes have added to the existing crop databases, and new models are starting to be used in fruit and seed set studies. [本文引用: 1] |
| [77] | DOI:10.1007/s00299-013-1462-xPMID:23719757URLEpigenetics has become one of the hottest topics of research in plant functional genomics since it appears promising in deciphering and imparting stress-adaptive potential in crops and other plant species. Recently, numerous studies have provided new insights into the epigenetic control of stress adaptation. Epigenetic control of stress-induced phenotypic response of plants involves gene regulation. Growing evidence suggest that methylation of DNA in response to stress leads to the variation in phenotype. Transposon mobility, siRNA-mediated methylation and host methyltransferase activation have been implicated in this process. This review presents the current status of epigenetics of plant stress responses with a view to use this knowledge towards engineering plants for stress tolerance. [本文引用: 2] |
| [78] | DOI:10.1126/science.1212959PMID:21921155URLAbstract Epigenetic information, which may affect an organism's phenotype, can be stored and stably inherited in the form of cytosine DNA methylation. Changes in DNA methylation can produce meiotically stable epialleles that affect transcription and morphology, but the rates of spontaneous gain or loss of DNA methylation are unknown. We examined spontaneously occurring variation in DNA methylation in Arabidopsis thaliana plants propagated by single-seed descent for 30 generations. We identified 114,287 CG single methylation polymorphisms and 2485 CG differentially methylated regions (DMRs), both of which show patterns of divergence compared with the ancestral state. Thus, transgenerational epigenetic variation in DNA methylation may generate new allelic states that alter transcription, providing a mechanism for phenotypic diversity in the absence of genetic mutation. [本文引用: 1] |
| [79] | DOI:10.1046/j.1365-313X.1996.10040601.xPMID:8893539URLCell cycle progression is regulated by cyclin-dependent kinases (CDKs). Arabidopsis thaliana contains two cdk genes, cdc2aAt and cdc2bAt . This paper compares the developmental and cell cycle phase-dependent transcription of both cdk genes. In situ hybridizations revealed that cdc2bAt steady-state mRNAs, much like cdc2aAt , are found both in meristematic cells and cells with a high proliferative competence. Cdc2aAt is expressed in every meristematic cell whereas cdc2bAt is found to be expressed in a patchy pattern. An even smaller number of meristematic cells express the mitotic cyc1At . These data indicate that cdc2bAt and cyc1At mRNAs accumulate in a particular cell cycle phase in agreement with evidence provided by hybridization experiments of flow cytometry-sorted nuclei and the use of cell cycle blockers on roots. The data indicate that cdc2bAt is preferentially expressed in S and G 2 phases whereas cdc2aAt expression is constitutive throughout the cell cycle, as shown previously. The existence of two distinct CDK classes in plants is proposed: (i) constitutively expressed CDKs containing a PSTAIRE motif (e.g. cdc2aAt ) and (ii) CDKs with divergent motifs which are expressed during a limited interval of the cell cycle (e.g. cdc2bAt ). [本文引用: 1] |
| [80] | DOI:10.1104/pp.107.098335PMID:17660355URLThe role of gibberellins (GAs) in tomato (Solarium lycopersicum) fruit development was investigated. Two different inhibitors of GA biosynthesis (LAB 198999 and paclobutrazol) decreased fruit growth and fruit set, an effect reversed by GA61 application. LAB 198999 reduced GA69 and GA66 content, but increased that of their precursors GA6261, GA6262, GA6967, and GA6068 in pollinated fruits. This supports the hypothesis that GA69 is the active GA for tomato fruit growth. Unpollinated ovaries developed parthenocarpically in response to GA61 > GA69 = GA62 > GA6068, but not to GA6967, suggesting that GA 20-oxidase activity was limiting in unpollinated ovaries. This was confirmed by analyzing the effect of pollination on transcript levels of SICPS, SIGA20ox1, -2, and -3, and SlGA3ox1 and -2, encoding enzymes of GA biosynthesis. Pollination increased transcript content of SlGA20ox1, -2, and -3, and SICPS, but not of SlGA3ox1 and -2. To investigate whether pollination also altered GA inactivation, full-length cDNA clones of genes encoding enzymes catalyzing GA 2-oxidases (SIGA2ox1, -2, -3, -4, and -5) were isolated and characterized. Transcript levels of these genes did not decrease early after pollination (5-d-old fruits), but transcript content reduction of all of them, mainly of SlGA2ox2, was found later (from 10 d after anthesis). We conclude that pollination mediates fruit set by activating GA biosynthesis mainly through up-regulation of GA20ox. Finally, the phylogenetic reconstruction of the GA2ox family clearly showed the existence of three gene subfamilies, and the phylogenetic position of SlGA2ox1, -2, -3, -4, and -5 was established. [本文引用: 1] |
| [81] | DOI:10.1016/j.pbi.2007.09.003PMID:17997126URLFruits come in a vast variety of forms with both dry and fleshy types being essential components of the human diet. Elegant studies on the dry fruits of Arabidopsis have identified a suite of transcription factors involved in their development and dehiscence. Recent discoveries in tomato have revealed a hitherto unsuspected regulatory network involved in the developmental regulation of ripening in these fleshy fruits. Intriguingly it has become apparent that tomato shares some elements of its regulatory network in common with those involved in fruit development in Arabidopsis. Furthermore epigenetic variation has been shown to influence tomato ripening. These discoveries are likely to have a major impact on strategies for crop improvement in fruit bearing species. [本文引用: 1] |
| [82] | DOI:10.1146/annurev-arplant-050312-120057URL [本文引用: 1] |
| [83] | DOI:10.1105/tpc.111.094870URL [本文引用: 1] |
| [84] | DOI:10.17660/ActaHortic.2003.620.12URLJasmonic acid (JA) is a metabolite of lipoxygenase (LOX) and a growth regulator, which has many physiological functions associated with ethylene. To determine the role of LOX and JA in ethylene biosynthesis during tomato fruit ripening, low-ethylene producing fruits from antisense ACC synthase (ACS) transgenic plants (ethylene production is only 0.5%-3.0% of the wild-type fruits) were compared with fruits from wild-type plants. Expression of a gene encoding LOX, TOMloxB, and relative LOX activity increased continuously in transgenic fruits after treated with 1000 碌L.L-1 ethephon at room temperature; meanwhile, two kinds of fruit treated with exogenous JA were examined to determine the role of JA and ethylene in TOMloxB expression. In wild type fruit, exogenous JA increased relative LOX activity followed continuously increasing TOMloxB transcripts, and promoted color change of fruits from green to yellow/orange within 24h treatment; TOMloxB transcripts decreased almost to zero at 48h. However, exogenous JA had no effect on low ethylene transgenic fruits; no color change was detected within 24h; both relative LOX activity and TOMloxB transcripts remains unchanged in 24h. The effect of LOX on ethylene biosynthesis was less than that on ACC oxidase, suggesting an indirect role of LOX in ethylene biosynthesis. Presence of ethylene appears necessary to affect JA-induced LOX gene expression, suggesting that the JA signal pathway interacts with that of ethylene, and that LOX may play a role in ethylene biosynthesis via JA. [本文引用: 1] |
| [85] | DOI:10.1111/tpj.12882PMID:25996898URLSummary Fruit set in angiosperms marks the transition from flowering to fruit production and a commitment to seed dispersal. Studies with Solanum lycopersicum (tomato) fruit have shown that pollination and subsequent fertilization induce the biosynthesis of several hormones, including auxin and gibberellins (GAs), which stimulate fruit set. Circumstantial evidence suggests that the gaseous hormone ethylene may also influence fruit set, but this has yet to be substantiated with molecular or mechanistic data. Here, we examined fruit set at the biochemical and genetic levels, using hormone and inhibitor treatments, and mutants that affect auxin or ethylene signaling. The expression of system–1 ethylene biosynthetic genes and the production of ethylene decreased during pollination-dependent fruit set in wild-type tomato and during pollination-independent fruit set in the auxin hypersensitive mutant iaa9–3 . Blocking ethylene perception in emasculated flowers, using either the ethylene-insensitive Sletr1–1 mutation or 1–methylcyclopropene (1–MCP), resulted in elongated parthenocarpic fruit and increased cell expansion, whereas simultaneous treatment with the GA biosynthesis inhibitor paclobutrazol (PAC) inhibited parthenocarpy. Additionally, the application of the ethylene precursor 1–aminocyclopropane-1-carboxylic acid (ACC) to pollinated ovaries reduced fruit set. Furthermore, Sletr1–1 parthenocarpic fruits did not exhibit increased auxin accumulation, but rather had elevated levels of bioactive GAs, most likely reflecting an increase in transcripts encoding the GA-biosynthetic enzyme SlGA20ox3, as well as a reduction in the levels of transcripts encoding the GA-inactivating enzymes SlGA2ox4 and SlGA2ox5. Taken together, our results suggest that ethylene plays a role in tomato fruit set by suppressing GA metabolism. [本文引用: 2] |
| [86] | [本文引用: 2] |
| [87] | DOI:10.1007/s00344-005-0015-0URL [本文引用: 2] |
| [88] | DOI:10.1038/47412PMID:10638745URLHistone proteins and the nucleosomes they form with DNA are the fundamental building blocks of eukaryotic chromatin. A diverse array of post-translational modifications that often occur on tail domains of these proteins has been well documented. Although the function of these highly conserved modifications has remained elusive, converging biochemical and genetic evidence suggests functions in several chromatin-based processes. We propose that distinct histone modifications, on one or more tails, act sequentially or in combination to form a 'histone code' that is, read by other proteins to bring about distinct downstream events. [本文引用: 1] |
| [89] | DOI:10.1093/jxb/erv361PMID:4588892URLWe combined fine-mapping, gene expression, genome sequence variation, and morphological and histological analyses to select candidate genes underlying fs8.1, a major tomato fruit shape QTL located in a pericentromeric region. fs8.1 is a major quantitative trait locus (QTL) that controls the elongated shape of tomato (Solanum lycopersicum) fruit. In this study, we fine-mapped the locus from a 47Mb to a 3.03Mb interval on the long arm of chromosome 8. Of the 122 annotated genes found in the fs8.1 region, 51 were expressed during floral development and six were differentially expressed in anthesis-stage ovaries in fs8.1 and wild-type (WT) lines. To identify possible nucleotide polymorphisms that may underlie the fruit shape phenotype, genome sequence analyses between tomato cultivars carrying the mutant and WT allele were conducted. This led to the identification of 158 single-nucleotide polymorphisms (SNPs) and five small indels in the fs8.1 interval, including 31 that could be associated with changes in gene expression or function. Morphological and histological analyses showed that the effects of fs8.1 were mainly on reproductive organ elongation by increasing cell number in the proximal鈥揹istal direction. Fruit weight was also increased in fs8.1 compared with WT, which was predominantly attributed to the increased fruit length. By combining the findings from the different analyses, we consider 12 likely candidate genes to underlie fs8.1, including Solyc08g062580 encoding a pentatricopeptide repeat protein, Solyc08g061560 encoding a putative orthologue of ERECTA, which is known to control fruit morphology and inflorescence architecture in Arabidopsis, Solyc08g061910 encoding a GTL2-like trihelix transcription factor, Solyc08g061930 encoding a protein that regulates cytokinin degradation, and two genes, Solyc08g062340 and Solyc08g062450, encoding 17.6kDa class II small heat-shock proteins. [本文引用: 2] |
| [90] | DOI:10.1016/j.laa.2009.09.019PMID:15131251URLFruit, corresponding to the plant's ovary (or ovaries), protect seed development and serve as the vehicle for seed dispersal to different habitats for species propagation. Fruit also provide humans with a source of nutrition, culinary diversity, and often great pleasure. Humans consume fruit from a wide range of plants, including members of the Rutaceae (citrus), Rosaceae (stone fruits, apples, pears), Solanaceae (eggplant, pepper, tomato, husk tomato), Cucurbitaceae (melons, squash, cucumbers), Ribaceae (berries), Vitaceae (grapes), and a multitude of additional species. [本文引用: 1] |
| [91] | DOI:10.1007/s00425-008-0743-zURL [本文引用: 2] |
| [92] | [本文引用: 2] |
| [93] | DOI:10.1038/nature11119PMID:22660326URLTomato (Solanum lycopersicum) is a major crop plant and a model system for fruit development. Solanum is one of the largest angiosperm genera(1) and includes annual and perennial plants from diverse habitats. Here we present a high-quality genome sequence of domesticated tomato, a draft sequence of its closest wild relative, Solanum pimpinellifolium(2), and compare them to each other and to the potato genome (Solanum tuberosum). The two tomato genomes show only 0.6% nucleotide divergence and signs of recent admixture, but show more than 8% divergence from potato, with nine large and several smaller inversions. In contrast to Arabidopsis, but similar to soybean, tomato and potato small RNAs map predominantly to gene-rich chromosomal regions, including gene promoters. The Solanum lineage has experienced two consecutive genome triplications: one that is ancient and shared with rosids, and a more recent one. These triplications set the stage for the neofunctionalization of genes controlling fruit characteristics, such as colour and fleshiness. [本文引用: 1] |
| [94] | DOI:10.1073/pnas.1116437108URL [本文引用: 1] |
| [95] | DOI:10.3389/fpls.2014.00227PMID:4034497URLDomestication of fruit and vegetables resulted in a huge diversity of shapes and sizes of the produce. Selections that took place over thousands of years of alleles that increased fruit weight and altered shape for specific culinary uses provide a wealth of resources to study the molecular bases of this diversity. Tomato (Solanum lycopersicum) evolved from a wild ancestor (S. pimpinellifolium) bearing small and round edible fruit. Molecular genetic studies led to the identification of two genes selected for fruit weight: FW2.2 encoding a member of the Cell Number Regulator family; and FW3.2 encoding a P450 enzyme and the ortholog of KLUH. Four genes were identified that were selected for fruit shape: SUN encoding a member of the IQD family of calmodulin-binding proteins leading to fruit elongation; OVATE encoding a member of the OVATE family proteins involved in transcriptional repression leading to fruit elongation; LC encoding most likely the ortholog of WUSCHEL controlling meristem size and locule number; FAS encoding a member in the YABBY family controlling locule number leading to flat or oxheart shape. For this article, we will provide an overview of the putative function of the known genes, when during floral and fruit development they are hypothesized to act and their potential importance in regulating morphological diversity in other fruit and vegetable crops. [本文引用: 5] |
| [96] | DOI:10.1016/S0031-9422(02)00223-6PMID:12453577URL28-Homobrassinolide ( 1) and 24-epibrassinolide ( 2) application resulted in acceleration of ripening of tomato pericarp discs. [本文引用: 1] |
| [97] | DOI:10.1105/tpc.105.032383URL [本文引用: 1] |
| [98] | DOI:10.1016/S0955-0674(03)00043-7PMID:12787767URLThe traditional view of chromatin envisions two states: one is ‘active’ and accessible to nucleases, whereas the other is ‘silent’ and relatively inaccessible. Recent evidence that combinations of diverse histone tail modifications represent a spectrum of chromatin states challenges this simple view. Here, we examine inter-relationships between chromatin remodeling, histone modification, DNA methylation, RNA interference, and nucleosome assembly activities. We find that the two-state view can accommodate these new findings, and that nucleosome assembly pathways may ultimately maintain euchromatic and heterochromatic states. [本文引用: 1] |
| [99] | DOI:10.1016/j.pbi.2011.10.001PMID:22035873URLEpigenetic processes such as DNA methylation are crucial for the development of flowering plants, and for protection of genome integrity via silencing of transposable elements (TEs). Recent advances in genome-wide profiling suggest that during reproduction DNA methylation patterns are at least partially transmitted or even enhanced in the next generation to ensure stable silencing of TEs. At the same time, parent-of-origin specific removal of DNA methylation in the accompanying tissue allows imprinted expression of genes. Here we summarize the dynamics of DNA methylation as a major epigenetic regulatory pathway during reproduction and seed development. [本文引用: 1] |
| [100] | [本文引用: 1] |
| [101] | DOI:10.1126/science.1153040PMID:18339939URLEdible fruits, such as that of the tomato plant and other vegetable crops, are markedly diverse in shape and size. SUN, one of the major genes controlling the elongated fruit shape of tomato, was positionally cloned and found to encode a member of the IQ67 domain--containing family. We show that the locus arose as a result of an unusual 24.7-kilobase gene duplication event mediated by the long terminal repeat retrotransposon Rider. This event resulted in a new genomic context that increased SUN expression relative to that of the ancestral copy, culminating in an elongated fruit shape. Our discovery demonstrates that retrotransposons may be a major driving force in genome evolution and gene duplication, resulting in phenotypic change in plants. [本文引用: 2] |
| [102] | DOI:10.1364/AO.36.000144PMID:20421195URLA major challenge in rice (Oryza sativa L.) production is to enhance water use efficiency (WUE) and maintain or even increase grain yield. WUE, if defined as the biomass accumulation over water consumed, may be fairly constant for a given species in given climate. WUE can be enhanced by less irrigation. However, such enhancement is largely a trade-off against lower biomass production. If WUE is defined as the grain production per unit amount of water irrigated, it would be possible to increase WUE without compromising grain yield through the manipulation of harvest index. Harvest index has been shown to be a variable factor in crop production, and in many situations, it is closely associated with WUE and grain yield in cereals. Taking rice as an example, this paper discussed crop management techniques that can enhance harvest index. Several practices such as post-anthesis controlled soil drying, alternate wetting and moderate soil drying regimes during the whole growing season, and non-flooded straw mulching cultivation, could substantially enhance WUE and maintain or even increase grain yield of rice, mainly via improved canopy structure, source activity, sink strength, and enhanced remobilization of pre-stored carbon reserves from vegetative tissues to grains. All the work has proved that a proper crop management holds great promise to enhance harvest index and, consequently, achieve the dual goal of increasing grain production and saving water. [本文引用: 1] |
| [103] | DOI:10.1016/j.tplants.2015.10.009PMID:26603980URLWith the rapidly expanding world population, food production must be substantially increased in the coming several decades to meet increasing global demands for food. Source ink interactions modulate carbon (C) assimilation, translocation, partitioning, and storage throughout the plant and have a pivotal role in determining the productivity of harvestable organs. Crop yield is dependent on the source ink relation, which is influenced by environmental changes. Understanding mechanisms by which plants control source activity and sink strength in the context of global climate changes, such as water limitation and high temperature, is of agronomic significance for the development of efficient breeding programs for improving crops. Many examples have demonstrated that manipulation of source activity and sink strength could significantly enhance crop productivity. However, crop productivity is determined by the combinatorial functions of multiple genes involved in signaling and metabolic pathways, which are still not well understood. Additionally, yield increase has not been evaluated in field trials in most studies. Therefore, more work is needed for the translation of basic findings to open field trials. [本文引用: 1] |
| [104] | DOI:10.1104/pp.109.136598PMID:19439574URLIt has been previously demonstrated, utilizing intraspecific introgression lines, that Lycopersicum Invertase5 (LIN5), which encodes a cell wall invertase, controls total soluble solids content in tomato (Solatium lycopersicum). The physiological role of this protein, however, has not yet been directly studied, since evaluation of data obtained from the introgression lines is complicated by the fact that they additionally harbor many other wild species al eles. To allow a more precise comparison, we generated transgenic tomato in which we silenced the expression of LIN5 using the RNA interference approach. The transformants were characterized by an altered flower and fruit morphology, displaying increased numbers of petals and sepals per flower, an increased rate of fruit abortion, and a reduction in fruit size. Evaluation of the mature fruit revealed that the transformants were characterized by a reduction of seed number per plant. Furthermore, detailed physiological analysis revealed that the transformants displayed aberrant pollen morphology and a reduction in the rate of pollen tube elongation. Metabolite profiling of ovaries and green and red fruit revealed that metabolic changes in the transformants were largely confined to sugar metabolism, whereas transcript and hormone profiling revealed broad changes both in the hormones themselves and in transcripts encoding their biosynthetic enzymes and response elements. These results are discussed in the context of current understanding of the role of sugar during the development of tomato fruit, with particular focus given to its impact on hormone levels and organ morphology. [本文引用: 1] |
| [105] | DOI:10.1016/j.fcr.2005.08.018URLSince their early migration from aquatic environments to the land, plants have had to cope with periodic and unpredictable environmental stresses during growth and development. Surviving such stresses over a long evolutionary scale led plants to acquire mechanisms by which they can sensitively perceive incoming stresses and regulate their physiology accordingly. The plant hormone abscisic acid (ABA) plays a major role in plant responses to stress. Although rapid production of ABA in response to drought and salt stresses is essential to define ABA as a stress hormone, an equally rapid catabolism of ABA when such stresses are relieved is also essential in that role. Since ABA mediates so many stress responses, the initial perception of dehydration and the subsequent changes in gene expression that lead to rapid ABA biosynthesis constitute the most important stress signal transduction pathway among all the plant responses to stresses. Identification of the genes involved and understanding their roles during stress perception and physiological regulation has become an important and exciting research field in recent years. This review covers mainly our understanding of this aspect. ABA-induced changes in gene expression and their roles in physiological regulation are dealt with in less detail. [本文引用: 1] |
| [106] | DOI:10.1371/journal.pgen.1000032PMID:18369451URLAuthor SummaryThe functional expression of DNA sequence depends on the chromatin status. Epigenetic marks at specific loci could affect local chromatin accessibility, thus affect the gene activity of that loci. We applied an enzyme methylome approach to globally detect one type of epigenetic mark, cytosine methylation at CCGG restriction sites. Simultaneous transcriptional profiling allowed gene expression differences to be compared with DNA methylation differences, suggesting functional regulatory regions. Our method reveals natural variation in chromatin patterns which may underlie phenotypic variation. [本文引用: 1] |
| [107] | [本文引用: 1] |
| [108] | DOI:10.1038/nbt.2462PMID:23354102URLRipening of tomato fruits is triggered by the plant hormone ethylene, but its effect is restricted by an unknown developmental cue to mature fruits containing viable seeds. To determine whether this cue involves epigenetic remodeling, we expose tomatoes to the methyltransferase inhibitor 5-azacytidine and find that they ripen prematurely. We performed whole-genome bisulfite sequencing on fruit in four stages of development, from immature to ripe. We identified 52,095 differentially methylated regions (representing 1% of the genome) in the 90% of the genome covered by our analysis. Furthermore, binding sites for RIN, one of the main ripening transcription factors, are frequently localized in the demethylated regions of the promoters of numerous ripening genes, and binding occurs in concert with demethylation. Our data show that the epigenome is not static during development and may have been selected to ensure the fidelity of developmental processes such as ripening. Crop-improvement strategies could benefit by taking into account not only DNA sequence variation among plant lines, but also the information encoded in the epigenome. [本文引用: 1] |
中国植物激素研究: 过去、现在和未来
1
2006
... 植物激素是植物体内产生的一类在很低浓度时即可对植物的生理过程产生重要调控作用的代谢产物.植物激素作用广泛, 在植物生长发育的各个过程及环境应答中均发挥重要作用(
1
2009
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2014
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
1995
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
1996
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2008
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
4
2015
... 番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高.由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(
... 核内复制在番茄果实早期发育过程中也具有重要作用.核内复制广泛存在于植物中, 但在番茄果实发育中尤为突出.在番茄果实早期发育时期, 果皮细胞存在内多倍性现象, 即各层细胞中DNA含量不同, 此为核内复制引起(
... 细胞分裂、膨大和核内复制协同调控番茄果实的早期发育, 共同构建生长发育模型并为果实器官形成提供生长动力, 最终决定果实的重量和大小.因此, 通过修饰改造有丝分裂和核内复制相关靶基因的表达模式来调控番茄果实的生长发育过程将是今后研究的热点和重点(
... 番茄果实具有农业和基础研究价值.由于番茄具有生长周期短、繁殖速度快和自花授粉等特点, 其已成为研究肉质果实的模式植物(
1
1999
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
2
2014
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2008
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2006
... 番茄果实具有农业和基础研究价值.由于番茄具有生长周期短、繁殖速度快和自花授粉等特点, 其已成为研究肉质果实的模式植物(
1
2005
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2010
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2012
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2005
... 核内复制在番茄果实早期发育过程中也具有重要作用.核内复制广泛存在于植物中, 但在番茄果实发育中尤为突出.在番茄果实早期发育时期, 果皮细胞存在内多倍性现象, 即各层细胞中DNA含量不同, 此为核内复制引起(
2
2014
... 植物器官的生长发育依赖于细胞分裂和膨大过程.细胞分裂的活性决定了细胞的数量从而决定了器官的形成; 细胞膨大的活性则决定其最终大小(
... 核内复制在番茄果实早期发育过程中也具有重要作用.核内复制广泛存在于植物中, 但在番茄果实发育中尤为突出.在番茄果实早期发育时期, 果皮细胞存在内多倍性现象, 即各层细胞中DNA含量不同, 此为核内复制引起(
1
2009
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(
1
2012
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
1
2013
... 除了DNA甲基化, 组蛋白修饰在番茄果实早期发育过程中也起重要作用.组蛋白翻译后修饰能够影响染色体的结构与基因表达.它们通过多种途径影响染色体的组装, 其中主要包括核小体净电荷修饰(
4
2008
... 在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化.番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... 基因还可调控番茄果实呈扁平形态(
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
2
2015
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
... 基因会导致番茄果实产生更大的胎座、更厚的果皮和更少的种子数量, 观察果皮可以发现其细胞层数和单位面积的细胞数量都明显增加(
1
2012
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
1988
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
2011
1
2015
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2010
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2000
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2000
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2004
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
3
1993
... 番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高.由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(
... 在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化.番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2004
... 在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化.番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (
1
2007
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2011
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
1
2012
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
1993
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
1996
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2011
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
1
2006
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2009
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2009
... 此外, Lauria和Rossi (2011)研究发现, 表观遗传调控在番茄果实生长发育过程中行使重要功能.表观基因组多态性现象(DNA甲基化和组蛋白修饰)在基因组功能发挥过程中具重要作用, 且可能导致表型多样性, 控制复杂的形状特性甚至可被遗传(
1
2000
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
1999
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2010
... 除了DNA甲基化, 组蛋白修饰在番茄果实早期发育过程中也起重要作用.组蛋白翻译后修饰能够影响染色体的结构与基因表达.它们通过多种途径影响染色体的组装, 其中主要包括核小体净电荷修饰(
3
2011
... 番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高.由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
... 番茄果实具有农业和基础研究价值.由于番茄具有生长周期短、繁殖速度快和自花授粉等特点, 其已成为研究肉质果实的模式植物(
4
2014
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
... ;
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
... ;
2011
1
2010
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(
1
2004
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2007
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
1992
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
1
2014
...
1
2007
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
1
2002
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
2
2016
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
... ;
1
2011
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2016
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
2
2012
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
... ;
1
2013
... 在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化.番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (
1
2010
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2002
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
1
2009
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2011
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2017
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
3
2011
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
... ;
1
2014
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2012
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2013
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
1
2003
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
1
2013
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2007
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
1
2012
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2006
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
3
2011
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... ;
... ;
1
2014
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
1995
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2012
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
2
2013
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(
1
2011
... 此外, Lauria和Rossi (2011)研究发现, 表观遗传调控在番茄果实生长发育过程中行使重要功能.表观基因组多态性现象(DNA甲基化和组蛋白修饰)在基因组功能发挥过程中具重要作用, 且可能导致表型多样性, 控制复杂的形状特性甚至可被遗传(
1
1996
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2007
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2008
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2013
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
1
2012
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2003
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
2
2015
... 在植物进化过程中, 表型变化常是人们关注的焦点, 尤其是营养器官和生殖器官的生长发育变化.番茄在进化过程中果实增大是最为明显的变化(约为其祖先的1 000倍) (
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
2
2008
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
... 的植株心皮更大且数量增多(
2
2005
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
... 在番茄果实早期发育过程中, 多种激素单独或协同调控果实的发育.生长素是调控果实发育最主要的激素之一, 其基本作用是调控细胞的伸长和分裂.在番茄果实早期发育过程中, 生长素在子房发育、坐果和果实增大等时期均起重要作用(
1
2000
... 除了DNA甲基化, 组蛋白修饰在番茄果实早期发育过程中也起重要作用.组蛋白翻译后修饰能够影响染色体的结构与基因表达.它们通过多种途径影响染色体的组装, 其中主要包括核小体净电荷修饰(
2
2015
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
1
2004
... 番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高.由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(
2
2008
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(
... ).但这种现象仅出现在果皮组织中, 在腔室组织中并未发现, 可能具有组织特异性.另外, 在番茄果实发育过程中, 不同组织的DNA甲基转移酶(DNA methyltransferase, DNMT)基因会出现差异性表达(
2
2015
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(
1
2012
... 番茄(Solanum lycopersicum)隶属茄科番茄属, 是一种重要的蔬菜作物, 原产于南美洲, 营养丰富, 栽培面积广且经济效益高.由于其具有生长周期短、繁殖速度快、自花授粉和简单的机理研究通路等优势, 番茄已成为肉质果实植物研究的模式植物之一(
1
2011
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
5
2014
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... ;
... ;
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
... ;
1
2002
... 果实发育是一个复杂的过程, 受多种因素的共同调控.其中, 植物激素的调控是番茄果实早期发育过程中最重要的调控途径之一(
1
2005
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2003
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2012
... 表观遗传(epigenetics)是指DNA序列不发生变化, 但基因表达却发生了可遗传的改变.这种改变是细胞内除了遗传信息以外的其它可遗传物质发生的改变, 且这种改变在发育和细胞增殖过程中能稳定传递.表观遗传信息基于DNA甲基化和组蛋白翻译后修饰(histone post-translational modifications, PTMs), 从而决定染色体的结构状态及调控DNA的转录活性(
1
2011
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
2
2008
... 在番茄的整个驯化过程中, 其果实形态和心室数量发生了巨大变化, 导致现代栽培番茄果实产量较野生型祖先增加1 000倍(
... SUN基因编码一种属于IQD (IQ67-Domain)蛋白家族的钙调蛋白结合蛋白, 可调控番茄果实的伸长, 从而改变番茄果实的形态与大小(
1
2010
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
1
2015
... 在过去几十年中, 改善作物产量主要通过化肥的使用、灌溉面积的扩大以及病虫害的治理.然而事实证明, 这些举措对于可持续发展和环境保护极为不利 (
1
2009
... 在番茄果实中, 光合作用可调控其发育和成熟, 并最终影响其产量与质量.转录因子GLK2 (GOL- DEN 2-LIKE)是番茄果实中叶绿体发育的重要调控因子, 其可影响果实中糖类物质和番茄红素的含量, 因此该转录因子基因是提高产量和质量的重要靶点(
1
2006
... 甜菜碱是一种广泛分布于动物、植物及微生物中的渗透调节物质.但越来越多的研究表明, 其非渗透调节作用更重要, 其中包括甜菜碱与植物激素间的互作.在非生物胁迫条件下, 外施茉莉酸、水杨酸、多胺、油菜素内酯以及脱落酸等植物激素, 均对甜菜碱的合成起显著调控作用(
1
2008
... 此外, Lauria和Rossi (2011)研究发现, 表观遗传调控在番茄果实生长发育过程中行使重要功能.表观基因组多态性现象(DNA甲基化和组蛋白修饰)在基因组功能发挥过程中具重要作用, 且可能导致表型多样性, 控制复杂的形状特性甚至可被遗传(
1
2012
... 在细胞分裂、膨大与核内复制调控番茄果实早期发育的过程中, 存在许多相关基因的差异表达及互作.果实重量基因fw2.2 (Fruit weight 2.2)是第1个被鉴定和克隆的与数量性状相关的基因(
1
2013
... 表观遗传调控在番茄果实早期发育过程中必不可少, 主要表现在DNA甲基化和组蛋白修饰过程.DNA甲基化一直以来被认为是稳定的遗传过程, 然而近期相关研究表明, 在相关发育因素的控制下(

{kind=link}
{kind=link}
{kind=link}
{kind=link}