 ,*, 吴丹, 韩彪, 王艳山东省林木种质资源中心, 济南 250102
,*, 吴丹, 韩彪, 王艳山东省林木种质资源中心, 济南 250102Effect of Conservation Conditions on Restricting Conservation of Acer rubrum cv. ‘Somerset’
Yang Xian, Xin Dong, Xiaoman Xie,*, Dan Wu, Biao Han, Yan WangShandong Forest Germplasm Resources Center, Jinan 250102, China通讯作者:
收稿日期:2018-01-11接受日期:2018-08-1网络出版日期:2019-01-30
| 基金资助: |
Corresponding authors:
Received:2018-01-11Accepted:2018-08-1Online:2019-01-30

摘要
关键词:
Abstract
Keywords:
PDF (2087KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
咸洋, 董昕, 解孝满, 吴丹, 韩彪, 王艳. 光照和温度对红花槭限制生长保存的影响. 植物学报, 2019, 54(1): 64-71 doi:10.11983/CBB18010
Xian Yang, Dong Xin, Xie Xiaoman, Wu Dan, Han Biao, Wang Yan.
卓越红花槭(Acer rubrum cv. ‘Somerset’)隶属槭树科槭属, 是从北美红花槭(Acer rubrum)中选育的优良品种之一, 其干形优美, 叶色鲜艳, 红叶期长, 适应性强, 观赏价值高。山东省林木种质资源中心于2007年从美国引进该树种, 并开展了其组培快繁研究, 已建立了完善的组培快繁体系, 繁殖效率达8.0 (咸洋等, 2013)。为防止品种退化, 保护优良品种, 现阶段应积极开展种质资源保存研究。针对已有成熟组培快繁体系的种质资源, 限制生长保存较为简便, 同时也能够保持其优良性状, 节约成本。限制生长保存是一种离体保存方法, 通过改变培养物的外界环境条件, 限制其生长, 但不死亡, 从而达到延长继代培养时间的目的。限制生长保存可采用的方法很多, 从操作方式上大致可分为2类。一是调整培养基成分, 如调整碳含量、改变激素种类和含量以及增加渗透调节物质; 二是改变保存条件, 如光照、温度和含氧量(徐刚标, 2000; 周逊和向长萍, 2008; 兰伟和陈发棣, 2010)。上述方法虽然原理不同, 但都能达到限制生长的目的。本研究以卓越红花槭为试材, 探讨了光照和温度变化对离体材料生长量的影响, 无论对组织培养还是限制生长保存都具有重要的借鉴意义。
1 材料与方法
1.1 实验材料
实验材料为山东省林木种质资源中心保存的卓越红花槭(Acer rubrum L. cv. ‘Somerset’)组培苗。其外植体母株来自山东省林木种质资源中心枣园库定植的卓越红花槭植株1-2年生枝条。1.2 方法
1.2.1 试验设计根据卓越红花槭组培苗生长特性, 选取4种光照时间(8、12、16和24小时)、2种光照强度(31.25和62.50 μmol·m-2·s-1)及4种温度(8、15、25和25/18°C), 设置12组离体保存处理(表1)。主要考察光照时间(T1-T8)、温度(T2、T9、T10和T11)、昼夜温差及光照强度(T2、T6、T11和T12)等因素对离体保存材料生长状态的影响。所有处理组均采用统一的培养基G23 (DKW+0.025 mg?L-1 TDZ+0.03 mg?L-1 6-BA+ 0.8 g?L-1 AC+30 g?L-1蔗糖)。每组约32个外植体, 接种16或17瓶, 每个外植体上2或3个芽点, 重复3次。每7天调查1次生长情况, 保存至182天。
Table 1
表1
表1卓越红花槭离体保存试验设计
Table 1
| Treatment | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Photoperiod (h) | 24 | 16 | 12 | 8 | 24 | 16 | 12 | 8 | 16 | 16 | 16 | 16 |
| Light intensity (μmol·m-2·s-1) | 62.50 | 62.50 | 62.50 | 62.50 | 31.25 | 31.25 | 31.25 | 31.25 | 62.50 | 62.50 | 62.50 | 31.25 |
| Temperature (°C) | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 15 | 8 | 25/18* | 25/18* |
新窗口打开|下载CSV
1.2.2 生长量调查
每7天进行1次生长调查。调查指标包括外植体分化率(分化外植体数量/接种外植体数量; 外植体上形成叶、芽、根等器官或愈伤组织时, 视其已发生分化, 本研究中多数为分化形成芽)、平均出芽数(丛生芽总数量/接种外植体数量)、生根率(生根苗数量/接种外植体数量)和平均株高(0.5 cm以上植株平均高度)等。其中平均出芽数是衡量分化能力的主要指标。为了减少调查中的误差, 同时更准确地反映离体材料生长状况, 根据生长的不同阶段, 将丛生芽分为3类: A类芽为萌动(芽形态肉眼可见, 叶片形态未显现); B类芽为萌发(叶片形态显现); C类芽为生长(芽高0.5 cm以上, 需测量高度)。
根据生长量调查结果绘制生长曲线, 并选取部分数据使用SPSS 18.0软件作进一步分析。
1.2.3 活力恢复
将所有瓶苗接种至新鲜的G23培养基上, 恢复培养28天, 调查生长量。培养条件: 光照强度为62.50 μmol·m-2·s-1, 光照16小时, 培养温度为25/18°C (在有光照时温度为25°C, 无光照时温度为18°C)。
2 结果与讨论
2.1 生长曲线分析
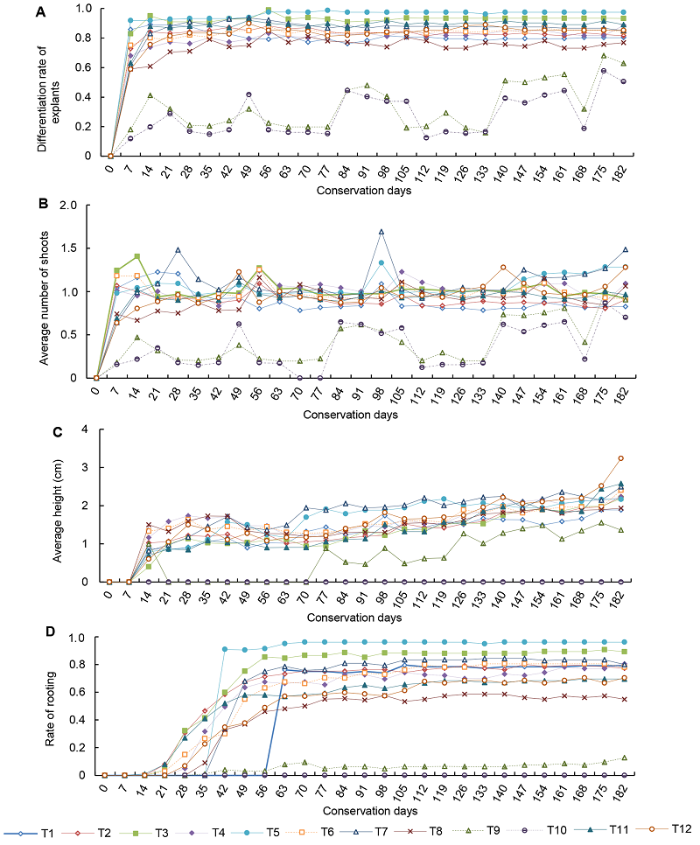
根据每次调查结果的平均值绘制离体材料182天的生长曲线(图1)。182天后的生长状态如图2所示。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1卓越红花槭生长曲线
(A) 外植体分化率; (B) 平均出芽数; (C) 平均高度; (D) 生根率。T1-T12同
Figure 1Growth curve of Acer rubrum cv. ‘Somerset’
(A) Differentiation rate of explants; (B) Average number of shoots; (C) Average height; (D) Rate of rooting. T1-T12 see
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2卓越红花槭离体保存182天
(A) T1; (B) T2; (C) T3; (D) T4; (E) T5; (F) T6; (G) T7; (H) T8; (I) T9; (J) T10; (K) T11; (L) T12。T1-T12同
Figure 2In vitro conservation for 182 days of Acer rubrum cv. ‘Somerset’
(A) T1; (B) T2; (C) T3; (D) T4; (E) T5; (F) T6; (G) T7; (H) T8; (I) T9; (J) T10; (K) T11; (L) T12. T1-T12 see
2.1.1 外植体分化率
由图1A可知, 大部分处理组分化率在第14天达到80%以上, 第56天又出现1个小高峰, 之后一直到调查结束变化都比较平稳。而T9和T10处理组却呈曲线上升, 调查期间间断出现分化率降低, 这与培养温度过低导致刚分化出的A类芽生长缓慢或死亡有关。根据外植体生长情况, 选取大部分处理出现峰值时期(即第7、14、56和175天)的数值作进一步数据分析。
2.1.2 平均出芽数
平均出芽数指总的平均出芽数, 是分析外植体材料生长分化能力的重要指标(图1B)。整个保存期间, 大部分处理的平均出芽数呈缓慢上升趋势。芽分化最快的阶段是0-14天, 14天后出现回落, 结合图1C表明此时平均高度仍保持较快增长, 因此, 此阶段转向营养生长。直至第28天, 大部分处理芽的增长都不明显, 甚至出现降低现象, 随后直至调查结束又出现几个生长高峰, 如第49-56天、第91天及第147天。高峰的间断出现说明外植体一直保持分化能力, 但是分化出的芽却无法生长。从单个处理组来看, T3处理组最早(第14天)达到峰值(1.4), 说明T3处理组生长较快, 可在短期内迅速扩繁。T7处理组分化开始时间较晚, 但是分化能力较强, 第28天首次达到峰值(1.48)后开始回落, 第98天平均出芽数达到最高(1.69), 第182天仍保持同时期最高值(1.48), 说明T7处理组能一直保持比较好的分化能力, 比较适合长期保存。T9和T10处理组出芽数明显低于其它处理组, 也间断有峰值和回落的情况, 但是从整个保存期来看, 其增长趋势最明显, 后期甚至超过常温的T2处理组。结合生长情况, 取峰值较明显的第7、14、28、49、98、147和175天的调查结果作进一步分析。
2.1.3 平均高度
平均高度主要反映外植体的营养生长情况。为避免由于芽过小引起测量误差, 我们只对C类芽, 即高度在0.5 cm以上的芽进行测量。由图1C可知, 几乎所有处理的平均高度都呈上升趋势, 最高的T12处理组达3.24 cm。7-14天平均高度增长最快, T8处理组达到同时期最高值(1.5 cm), 但T8处理组光照时间短, 光强弱, 叶色嫩绿, 植株细弱, 明显表现出光照不足引起的徒长, 在保存后期植株生长较弱, 内生菌污染严重(图2)。T9处理组平均高度明显低于其它处理组, 呈曲线增长, 不断出现回落, 是因为长出的植株不耐低温死亡, 曲线增长说明离体材料在逐渐适应低温环境。由于温度过低, T10处理组一直没有C类芽长成。根据生长情况取生长变化较明显的第14天、生长稳定后的第35天及最终生长量第182天的数据作进一步分析, 并同时分析随机抽取的第91、126及154天的数据。
2.1.4 生根率
虽然生根与否对离体保存效果影响不大, 但是由于我们发现很多离体材料出现生根现象, 因此简单探讨对生根产生影响的因素。调查显示, 生根情况有比较类似的规律(图1D), 相对芽分化, 生根时间较晚, 最早的第14天生根, 大多数处理组集中在第21-35天开始生根, 在35-70天之间生根率增长最快。其中处理组T1、T5及T7增长最迅速, 开始生根后几乎呈直线型增长, 在70天之后生根率基本呈较稳定的态势。取生根率变化最明显的第21-70天及第182天的数据进行分析。
2.2 离体保存条件
2.2.1 光照时间和光照强度对离体保存的影响T1-T8处理组为不同光照时间和光照强度的对照实验。通过生长曲线选取外植体分化率、平均出芽数、平均高度及生根率等方面有代表性的数据, 利用SPSS 18.0软件进行双因素方差分析, 结果见表2。光照时间对外植体分化率和平均出芽数的影响均在前期(外植体分化率在第7、14和56天, 平均出芽数在第7、14和28天)达极显著水平(P<0.01), 对平均高度的影响仅在第91天达显著水平, 对生根率的影响在整个保存期都达极显著水平。光照强度对外植体分化率和平均出芽数的影响均在前期(第7和14天)达显著或极显著水平, 对平均出芽数的影响在后期(第147和182天)仍可达显著或极显著水平, 对平均高度的影响在中期(第91和126天)达极显著水平, 对生根率的影响在第21-56天均达显著或极显著水平。同时, 二者有较强的交互作用。
2.2.2 温度对离体保存的影响
T2、T9、T10及T11处理组为不同温度下的对照实验。表2显示, 温度对外植体分化率、平均出芽数、平均高度和生根率的影响大部分都达到极显著水平。仅有的2次不显著, 一是第14天的平均株高, 此时正是组培苗开始生长的时期, 株高区别不明显; 二是第182天的平均出芽数。结合图1B发现, 随着保存时间的延长, T9和T10两个低温处理组平均出芽数逐步上升, 后期甚至达到常温处理组水平。由此推断, 随着处理时间的延长, 低温对离体材料芽分化能力的抑制作用正在减弱, 或离体材料正在逐步适应低温环境, 而其作用机理和182天之后的作用效果有待进一步研究与验证。
Table 2
表2
表2光照和温度对卓越红花槭离体保存的影响
Table 2
| Conservation days | Significance testing of P and LI | Significance testing of Tm | Significance testing of DTV and LI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | LI | P×LI | Tm | DTV | LI | DTV×LI | ||||
| Differentiation rate of explants | 7 | 0** | 0.018* | 0.002** | 0** | 0.05 | 0.84 | 0.589 | ||
| 14 | 0** | 0.004** | 0.036* | 0.005** | 0.914 | 0.009** | 0.115 | |||
| 56 | 0.002** | 0.075 | 0.001** | 0** | 0.771 | 0.283 | 0.302 | |||
| 182 | 0.297 | 0.907 | 0.01* | 0** | 0.322 | 0.86 | 0.564 | |||
| Average number of shoots | 7 | 0** | 0** | 0** | 0** | 0** | 0.695 | 0.248 | ||
| 14 | 0** | 0.003** | 0.002** | 0.009** | 0.013* | 0.772 | 0.008** | |||
| 28 | 0** | 0.236 | 0** | 0** | 0.8 | 0.488 | 0.479 | |||
| 49 | 0.017* | 0.248 | 0.033* | 0** | 0.003** | 0.244 | 0.563 | |||
| 98 | 0.129 | 0.113 | 0.237 | 0.001** | 0.036* | 0.209 | 0.574 | |||
| 147 | 0.167 | 0.011* | 0.193 | 0.006** | 0.383 | 0.217 | 0.992 | |||
| 182 | 0.057 | 0.001** | 0.008** | 0.131 | 0.016* | 0.03* | 0.063 | |||
| Average height | 14 | 0.589 | 0.516 | 0.751 | 0.081 | 0.071 | 0.641 | 0.680 | ||
| 35 | 0.113 | 0.322 | 0.942 | 0** | 0.845 | 0.077 | 0.744 | |||
| 91 | 0.021* | 0** | 0.004** | 0.002** | 0.464 | 0** | 0.670 | |||
| 126 | 0.733 | 0.002** | 0.039* | 0.004** | 0.636 | 0.016* | 0.422 | |||
| 154 | 0.207 | 0.291 | 0.093 | 0.003** | 0.646 | 0.549 | 0.501 | |||
| 182 | 0.187 | 0.294 | 0.182 | 0** | 0.009** | 0.034* | 0.185 | |||
| Rate of rooting | 21 | 0.004** | 0.041* | 0.213 | 0.037* | 0.541 | 0.025* | 0.504 | ||
| 28 | 0** | 0** | 0.002** | 0.001** | 0.225 | 0.004** | 0.658 | |||
| 35 | 0** | 0** | 0.012* | 0** | 0.409 | 0.008** | 0.885 | |||
| 42 | 0.885 | 0.35 | 0** | 0** | 0.896 | 0.003** | 0.336 | |||
| 49 | 0** | 0.002** | 0** | 0** | 0.018* | 0.006** | 0.415 | |||
| 56 | 0** | 0.001** | 0** | 0** | 0.005** | 0.036* | 0.905 | |||
| 63 | 0** | 0.329 | 0.004** | 0** | 0.01* | 0.49 | 0.432 | |||
| 70 | 0** | 0.156 | 0.001** | 0** | 0.054 | 0.448 | 0.423 | |||
| 182 | 0.015* | 0.284 | 0.007** | 0** | 0.09 | 0.713 | 0.901 | |||
新窗口打开|下载CSV
2.2.3 昼夜温差和光照强度对离体保存的影响
昼夜温差对外植体分化率影响不显著, 对平均高度和生根率的影响仅在中后期达显著或极显著水平(平均高度在第182天, 生根率为第49-63天), 而在芽分化的各个时期对平均出芽数的影响都达显著或极显著水平。光照强度对外植体分化率的影响在第14天达极显著水平, 对平均出芽数和平均高度的影响在中后期(平均出芽数为第182天, 平均高度为第91、126和182天)达显著或极显著水平, 对生根率的影响在第21- 56天均达到显著或极显著水平。从整体来看, 二者没有明显的交互作用(表2)。
2.3 活力恢复
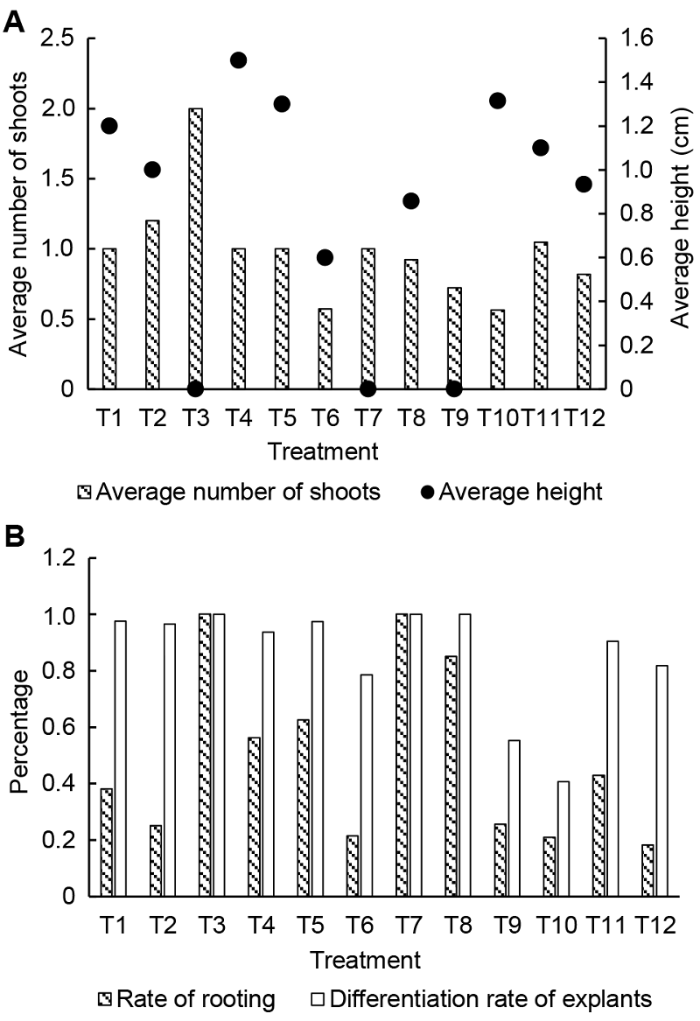
对恢复培养28天的离体保存材料进行生长量调查, 结果如图3, 生长状态如图4。在平均出芽数方面(图3A), T3处理组最高(2), T10处理组最低(0.56)。平均高度(图3A)最高为T4处理组(1.5), 其次为T5 (1.3)和T10 (1.3)处理组, 而T3、T7和T9处理组平均高度均为0, 说明在恢复期内没有C类芽形成。大部分处理的外植体分化率在80%以上(图3B), 其中T3和T7处理组达到100%, T9和T10处理组较低(60%以下)。生根率方面(图3B), T3和T7处理组达到100%, T8处理组为85%, T5处理组为63%, 其余均在60%以下。值得注意的是, T3处理组的平均出芽数、外植体分化率及生根率都达到最大值, 但是平均高度却是0, 说明该处理组植株有较强的分化能力, 但营养生长并不旺盛。T9和T10两个低温处理组相比, T9处理组在平均出芽数、外植体分化率和生根率上略高于T10处理组, 但远远低于25°C下的处理组; 而在平均高度方面, T10处理组远远高于T9处理组, 甚至超过了部分25°C下的处理组。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3卓越红花槭活力恢复培养28天
(A) 平均出芽数和平均高度; (B) 外植体分化率和生根率。T1- T12同
Figure 3Recovering culture for 28 days of Acer rubrum cv. ‘Somerset’
(A) Average number of shoots and average height; (B) Differentiation rate of explants and rate of rooting. T1-T12 see
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4卓越红花槭活力恢复28天
Figure 4Recovering culture for 28 days of Acer rubrum cv. ‘Somerset’
(A) T1; (B) T2; (C) T3; (D) T4; (E) T5; (F) T6; (G) T7; (H) T8; (I) T9; (J) T10; (K) T11; (L) T12。T1-T12同表1。Bars=1 cm
(A) T1; (B) T2; (C) T3; (D) T4; (E) T5; (F) T6; (G) T7; (H) T8; (I) T9; (J) T10; (K) T11; (L) T12. T1-T12 see Table 1. Bars=1 cm
2.4 讨论
卓越红花槭离体材料在25°C下生长规律可概括为: 前期以分化为主; 后期以营养生长为主, 分化能力有所减弱, 而且分化出的芽不能全部长大。原因可能有2种, 一是培养基内的养分和激素已耗尽, 无力供给更多的芽生长; 二是前期长成的植株挤占了有限的生长空间, 致使幼芽无法生长。而在8°C和15°C条件下, 生长规律有所不同, 外植体分化率和平均出芽数增长缓慢, 但是在最后2次调查中上述二指标接近25°C下的处理组, 说明低温限制了离体材料的生长, 但是离体材料也在逐渐适应低温环境。限制生长保存中, 低温保存是较常用的方法, 不同种质资源对低温的耐受性不同。例如, 赤松(Pinus densiflora)的丛生芽适合在17°C保存, 在14°C长期保存会影响其恢复(张红岩等, 2010); 欧洲山杨(Populus tremula)可以在10°C下保存1年, 而不影响活力恢复, 但是在低温下能检测到脱落酸、脯氨酸及超氧化物歧化酶等合成活跃, 低温处理3天即能检测到2种新合成蛋白(Jouve et al., 2000)。大量研究证明低温胁迫能引发植物的一系列生理变化, 甚至影响基因表达, 但还不明确低温胁迫是否会引起遗传变异(徐呈祥, 2012)。光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等。多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(Kozai et al., 1995; Ahmad et al., 2014; 燕丽萍等, 2016)。此外, 不同光质也会影响组培苗的生长和分化(任桂萍等, 2016)。在卓越红花槭离体保存中, 我们发现光照时间和光照强度都对离体材料的生长及分化有影响, 而且影响效果持久, 交互作用显著, 不能以单一因素而论。例如, 光照24小时条件下, 图1中所有指标都是弱光高于强光(图1中T1和T5的对比), 随着光照时间减少, 这种差异越来越小(图1中T2和T6的对比), 并逐渐转为强光高于弱光(图1中T3和T7以及T4和T8的对比)。在对文心兰(Oncidium hybridum) (郑卫杰等, 2011)和非洲菊(Gerbera jamesonii) (杨博等, 2012)的研究中, 较小的昼夜温差有利于试管苗形态建成, 较大的昼夜温差有助于干物质积累。本研究表明, 昼夜温差对卓越红花槭离体材料的平均出芽数和生根率影响较大, 尤其是有利于芽的形成, 影响持久且显著, 这与前人的研究结果不同。
在最佳保存条件筛选上, 25°C下的处理组活力恢复情况最好, 但在较低温度下可以有效控制污染。最佳保存条件的选择可根据保存环境和保存期限综合考虑。但无论采用哪种方式, 其核心应是保证种质材料的分化能力, 从而使种质材料能够迅速恢复生长。如T3 (图2)和T7 (图2) 处理组都有比较旺盛的分化能力, 较适于保存, 而T2 (图2)和T12 (图2)处理组虽然平均出芽数也较高, 但植株生长过于旺盛, 影响后期芽的生长和分化, 在活力恢复时可用的有效芽偏少, 因此保存效果稍差。
北美红花槭是引进的槭树科种质资源, 我国槭树科种质资源非常丰富(刘静波等, 2012), 但很多种已处于濒危或极危状态, 如血皮槭(A. griseum)、剑叶槭(A. lanceolatum)、梧桐槭(A. firmianioides)及羊角槭(A. yanjuechi)。有关槭树科组培繁育技术方面的研究较多, 但是其种质资源保存仍以原地和异地保存为主, 离体保存尚未见报道。了解卓越红花槭离体材料在限制生长保存期间的生长规律和主要影响因素, 对于发展槭树科组培繁育技术和离体保存技术都具有重要意义。
限制生长保存是以组培技术为基础发展起来的保存方式, 是离体保存的重要组成部分, 在种质资源保存中得到广泛应用。英国邱园千年种子库通过微体快繁技术繁育和保存了超过3 000份植物种质资源的离体材料(Sarasan et al., 2006)。在限制生长保存技术研究中, 快繁和保存一体化的研究趋势也越发明显(Silveira et al., 2009; Salcedo-Morales et al., 2009)。因此, 组培技术的完善是生长保存技术发展的基础。目前, 很多植物已开发出成熟的组培快繁体系, 但由于种和品种间的差异, 对组培条件要求各有其特殊性, 仍有多种植物的组培快繁体系尚未建立, 尤其是木本植物, 如血皮槭和蒙古栎(Xylosma racemosum), 这大大阻碍了种质资源保存技术的发展。因此, 作为一种经典的繁育技术, 组培技术在种质资源保存工作中仍然具有广阔的研究和应用前景。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3969/j.issn.1004-4329.2010.02.019URL [本文引用: 1]

植物种质资源是人类赖以生存和发展的重要物质基础,保持其遗传多样性对实现农业可持续发展具有重要的战略意义。缓慢生长保存法是植物种质资源保存切实可行的方法之一,目前已被广泛应用于植物种质保存中。本中综述了植物种质缓慢生长法保存的原理、方法、影响因素及其遗传稳定性的鉴定等,分析了目前存在的问题,并提出了对策。
DOI:10.3969/j.issn.1000-6850.2012.25.001URL [本文引用: 1]
More and more attentions are paid to Acer resources for their characteristics as ornamental trees, lumber trees, medicinal plants. In order to explore such a precious resource of Acer to China, the research advances of Acer were summarized, including the aspects of resource investigation, propagation technology, genetic basis, introduction and domestication. Economic values of Acer were concluded as ornamental trees, lumber trees, medicinal plants. In future, people paid attention to collect and protect the gene resources, introduction and domestication, integrative research and development utilization, and in order to realize rational utilization and detailed research of plant resources.
DOI:10.11983/CBB14196URL [本文引用: 1]
以蝴蝶兰(Phalaenopsis ssp.)栽培种绿熊(Green Bear)和大辣椒(Big Chilli)为培养材料,研究不同光质的发光二极管(LED)组合对组织培养过程中增殖及生根的影响。结果表明,红光更有利于蝴蝶兰单芽增殖、干重、鲜重以及株高的增加,但不利于叶片叶绿素的积累;蓝光有利于叶片叶绿素的积累,并能提高根系活力;远红光则对根长和根系活力的增加作用更显著。增殖扩繁阶段的最适LED为暖白,2种蝴蝶兰芽增殖系数分别比白色荧光灯(对照)高出53.17%和46.37%。生根诱导阶段的最适LED组合为红:蓝:远红=3:6:1,根长及根系活力均较对照显著增加。该研究结果为LED光源在蝴蝶兰组织培养中的应用奠定了基础。
[本文引用: 1]
DOI:10.5846/stxb201106260945URL [本文引用: 1]
低温胁迫是世界范围内影响植物产量和品质的主要非生物胁迫。植物抗寒生理生态研究是比较活跃和发展很快的领域。文章综述了提高植物抗寒性机理的研究进展。大量科学研究和生产实践表明,气象因素与植物自身因素是影响植物抗寒性的关键因素,前者主要是温度、光周期和水分,后者主要是植物的遗传学基础、生长时期、发育水平以及低温胁迫下细胞的抗氧化能力。保证植物抗寒基因充分表达对提高植物抗寒性有重要意义。植物抗寒性的遗传机制与调控主要通过5条路径实现:丰富多样的植物低温诱导蛋白,低温转录因子DREB/CBF可同时调控多个植物低温诱导基因的表达,DREB/CBF与辅助因子相互作用调控下游基因表达,Ca2+、ABA及蛋白质磷酸化上游调控低温诱导基因表达,以及不饱和脂肪酸酶基因的表达。基因工程改良植物抗寒性已获重要进展,但距产业化尚有许多开创性的工作要做,目前主要通过导入抗寒调控基因和抗寒功能基因而实现,后者主要是导入抗渗透胁迫相关基因、抗冻蛋白基因、脂肪酸去饱和代谢关键酶基因、SOD等抗氧化系统的基因以及与植物激素调节有关的基因。农林技术对提高植物抗寒性有重大实用价值,其中的不少技术蕴涵着深刻的科学机理,重点评述了抗寒育种、抗砧嫁接、抗寒锻炼、水肥耦合及化学诱导五大技术提高植物抗寒性的作用机理。展望了提高植物抗寒性的研究。
DOI:10.3969/j.issn.1673-923X.2000.04.008URL [本文引用: 1]
植物种质资源是物种进化、遗传学研究及植物育种的物质基础,也是人类生存与持续发展的基本依存条件,为此,国内外****对植物种质保存的原理与技术进行了全面研究。在简要概述常规种质保存的基础上,着重阐述了离体保存原理,全面综述了该项技术在植物种质保存中的主要方法、程序及保存效果评价等的最新研究进展。
DOI:10.11983/CBB15222URL [本文引用: 1]
该文探讨了基本培养基、外植体、培养条件以及植物生长调节剂配比对绒毛白蜡(Fraxinus velutina)体胚诱导的影响。结果表明,胚根是诱导体胚发生的最佳外植体;体胚诱导的最适培养基为改良MS+2 mg·L^-1 6-BA+0.1 mg·L^-1 NAA、30g·L^-1蔗糖、5.0 g·L^-1琼脂;暗培养20天后进行光照培养(14小时光照/10小时黑暗),光密度为100^-120μmol·m^-2·s^-1,昼温度(25±2)°C,夜温度(18±2)°C;成功诱导出体细胞胚并获得再生植株,体胚诱导率可达59.8%,体胚萌发率达81.2%。壮苗最适培养基为改良WPM+0.5 mg·L^-1 6-BA+0.2 mg·L^-1 ZT+0.01 mg·L^-1 NAA。生根最适培养基为改良1/2MS+1.0 mg·L^-1IBA+0.05 mg·L^-1 NAA+20 g·L^-1蔗糖,生根率高达97.3%,试管苗移栽成活率达97.8%。
DOI:10.3969/j.issn.1001-7461.2012.02.19URL [本文引用: 1]
以非洲菊‘瑞扣’试管苗为材料,研究不同昼夜温差(0℃、3℃、6℃、9℃、12℃)处理对其生长的影响。结果表明,0℃昼夜温差处理下试管苗株高、叶数、最大叶长、根数、最大根长、总鲜干重、地上部分鲜干重、地下部分鲜干重、叶绿素a、叶绿素b、总叶绿素含量及根系活力均高于其他处理。昼夜温差9℃处理时试管苗的可溶性糖含量及干物率均优于其他处理,达到最大值。故在昼夜温差0℃处理时有利于促进非洲菊试管苗的形态生长、叶绿素合成及根部生长,在昼夜温差9℃处理时有利于试管苗的可溶性糖合成和干物质积累。
DOI:10.3969/j.issn.1000-2006.2010.05.002URL [本文引用: 1]
从抗病赤松丛生芽的保存温度和保存时间以及山梨醇浓度、NAA和6-BA组合等方面探讨了离体条件下赤松组培丛生芽的低温保存条件。结果表明:保存温度为17℃时,丛生芽存活率达到96.7%;培养基中单独添加0.1mg/L的山梨醇或0.1mg/L多效唑均可缓解冷害,添加0.2mg/L的6-BA+(0.1~0.2)mg/L的NAA组合保存效果最好,可使恢复生长后丛生芽的增殖系数提高到5.9~6.0;不同基因型的丛生芽低温保存效果差异显著,其中,1^#-A、7^#-7、16^#-2保存效果较好。
URL [本文引用: 1]
以文心兰‘柠檬黄’试管苗为材料,探讨了不同昼夜温差对其生长的影响。结果表明:昼夜温差 3℃处理下文心兰试管苗的株高、叶长、总鲜重、地上部鲜重、总干重、地上部干重均达到最大值,且与其他处理差异性显著。昼夜温差12℃时文心兰试管苗根 数、最大根长、地下部鲜重及干重最大,且在此处理下可溶性糖含量和根系活力也达到最大值。叶绿素a、叶绿素b、总叶绿素含量和干物率均随着昼夜温差的不断 增大呈现出增大的趋势,且在昼夜温差15℃时达到最大值。综上分析,在昼夜温差3℃时最有利于促进文心兰试管苗的形态生长,昼夜温差12℃时最有利于根部 的生长,昼夜温差15℃时最有利于干物质的积累和叶绿素的合成。
URL [本文引用: 1]
综述了缓慢生长离体保存技术在植物种质资源保存方面的研究进展,并列述了植物种质资源离体保存过程中的遗传变异概况。
.
[本文引用: 1]
.
DOI:10.1016/S0176-1617(00)80144-3URL [本文引用: 1]
Microcuttings of Populus tremula L. were successfully in vitro conserved for more than 1 year at 10 C on Murashige and Skoog (MS) medium devoid of any growth regulator. During storage, an important limitation of growth was observed. Survival was unaffected and the recovery of multiplication ability after transfer to normal conditions was satisfactory. From the first days of treatment microcuttings cultured at 10 C when compared with cultures grown at 23 C presented important biochemical changes. Endogenous contents of ABA and proline were affected during cold exposure as well as activities of several free radicals scavenging involved enzymes such as superoxide dismutase (SOD), dehydroascorbate reductase (DHAR) or glutathione reductase (GR). Moreover, de novo synthesis and accumulation of two new proteins with a molecular weight of about 120 kDa were observed after 3 days of chilling treatment.
.
DOI:10.1016/0304-4238(95)00828-4URL [本文引用: 1]
Stem elongation and growth of potato plantlets under three DIF (difference in photoperiod and dark period temperatures) levels, 619, 0 and +9, combined with two PPF (photosynthetic photon flux) levels, 70 (low) and 140 (high) μmol m 612 s 611 provided by white cool fluorescent lamps, under 16 h day 611 (long) or 8 h day 611 (short) photoperiods, were studied. Four nodal cuttings were cultured for 21 days on 0.6 × 10 614 m 3 MS (Murashige and Skoog, 1962, Physiol. Plant. , 15: 473–497) agar (8 kg m 613 ) medium with no added sugar in 3.7 × 10 613 m 3 polycarbonate boxes. Each box had two 10 mm holes covered with microporous filter to facilitate air exchange (3.6 air exchanges per hour). The average daily temperature in the culture room was set the same at 23 °C for all treatments, and CO 2 concentration and relative humidity were maintained at 400–500 μmol mol 611 and 50–70%, respectively. Stem length was significantly suppressed under 0 or 619 DIF, high PPF and long photoperiod. Stem diameter, leaf area and number of leaves were significantly enhanced by long photoperiod and high PPF, but affected little by DIF level. Specific leaf area was little affected by photoperiod, but decreased under high PPF and under low DIF. Long photoperiod and high PPF led to an increase in the fresh and dry weights maintaining similar percentage dry matter and to enhanced root growth. Under the same amount of integrated PPF, fresh and dry weights of leaf, stem, root and whole plantlet were significantly higher under the long photoperiod and low PPF conditions than under the short photoperiod and high PPF conditions. Because of suppressed root growth under short photoperiod, shoot to root dry weight ratio increased under short photoperiod, but was not affected by DIF. It is suggested that under photoautotrophic conditions a combination of high PPF level, long photoperiod, and zero or negative DIF produces potato plantlets in vitro of short and thick stem with similar number and increased area of leaves, which are desirable for transfer to ex vitro conditions.
.
[本文引用: 1]
.
DOI:10.1079/IVP2006769URL [本文引用: 1]
In vitro techniques have found increasing use in the conservation of threatened plants in recent years and this trend is likely to continue as more species face risk of extinction. The Micropropagation Unit at Royal Botanic Gardens, Kew, UK (RBG Kew) has an extensive collection of in vitro plants including many threatened species from throughout the world. The long history of the unit and the range of plants cultured have enabled considerable expertise to be amassed in identifying the problems and developing experimental strategies for propagation and conservation of threatened plants. While a large body of knowledge is available on the in vitro culture of plants, there are limited publications relating to threatened plant conservation. This review highlights the progress in in vitro culture and conservation of threatened plants in the past decade (1995-2005) and suggests future research directions. Works on non-threatened plants are also included wherever methods have applications in rare plant conservation. Recalcitrant plant materials collected from the wild or ex situ collections are difficult to grow in culture. Different methods of sterilization and other treatments to establish clean material for culture initiation are reviewed. Application of different culture methods for multiplication, and use of unconventional materials for rooting and transplantation are reviewed. As the available plant material for culture initiation is scarce and in many cases associated with inherent problems such as low viability and endogenous contamination, reliable protocols on multiplication, rooting, and storage methods are very important. In this context, photoautotrophic micropropagation has the potential for development as a routine method for the in vitro conservation of endangered plants. Long-term storage of material in culture is challenging and the potential applications of cryopreservation are significant in this area. Future conservation biotechnology research and its applications must be aimed at conserving highly threatened, mainly endemic, plants from conservation hotspots.
[本文引用: 1]
植物种质资源缓慢生长法保存研究进展
1
2010
... 卓越红花槭(Acer rubrum cv. ‘Somerset’)隶属槭树科槭属, 是从北美红花槭(Acer rubrum)中选育的优良品种之一, 其干形优美, 叶色鲜艳, 红叶期长, 适应性强, 观赏价值高.山东省林木种质资源中心于2007年从美国引进该树种, 并开展了其组培快繁研究, 已建立了完善的组培快繁体系, 繁殖效率达8.0 (
槭属植物种质资源研究进展
1
2012
... 北美红花槭是引进的槭树科种质资源, 我国槭树科种质资源非常丰富(
不同光质的LED对蝴蝶兰组织培养增殖及生根的影响
1
2016
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
一种红花槭的组培快繁方法
1
2014
... 卓越红花槭(Acer rubrum cv. ‘Somerset’)隶属槭树科槭属, 是从北美红花槭(Acer rubrum)中选育的优良品种之一, 其干形优美, 叶色鲜艳, 红叶期长, 适应性强, 观赏价值高.山东省林木种质资源中心于2007年从美国引进该树种, 并开展了其组培快繁研究, 已建立了完善的组培快繁体系, 繁殖效率达8.0 (
提高植物抗寒性的机理研究进展
1
2012
... 卓越红花槭离体材料在25°C下生长规律可概括为: 前期以分化为主; 后期以营养生长为主, 分化能力有所减弱, 而且分化出的芽不能全部长大.原因可能有2种, 一是培养基内的养分和激素已耗尽, 无力供给更多的芽生长; 二是前期长成的植株挤占了有限的生长空间, 致使幼芽无法生长.而在8°C和15°C条件下, 生长规律有所不同, 外植体分化率和平均出芽数增长缓慢, 但是在最后2次调查中上述二指标接近25°C下的处理组, 说明低温限制了离体材料的生长, 但是离体材料也在逐渐适应低温环境.限制生长保存中, 低温保存是较常用的方法, 不同种质资源对低温的耐受性不同.例如, 赤松(Pinus densiflora)的丛生芽适合在17°C保存, 在14°C长期保存会影响其恢复(
植物种质资源离体保存研究进展
1
2000
... 卓越红花槭(Acer rubrum cv. ‘Somerset’)隶属槭树科槭属, 是从北美红花槭(Acer rubrum)中选育的优良品种之一, 其干形优美, 叶色鲜艳, 红叶期长, 适应性强, 观赏价值高.山东省林木种质资源中心于2007年从美国引进该树种, 并开展了其组培快繁研究, 已建立了完善的组培快繁体系, 繁殖效率达8.0 (
绒毛白蜡体胚诱导和植株再生
1
2016
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
昼夜温差对非洲菊试管苗生长的影响
1
2012
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
抗病赤松组培丛生芽低温保存条件研究
1
2010
... 卓越红花槭离体材料在25°C下生长规律可概括为: 前期以分化为主; 后期以营养生长为主, 分化能力有所减弱, 而且分化出的芽不能全部长大.原因可能有2种, 一是培养基内的养分和激素已耗尽, 无力供给更多的芽生长; 二是前期长成的植株挤占了有限的生长空间, 致使幼芽无法生长.而在8°C和15°C条件下, 生长规律有所不同, 外植体分化率和平均出芽数增长缓慢, 但是在最后2次调查中上述二指标接近25°C下的处理组, 说明低温限制了离体材料的生长, 但是离体材料也在逐渐适应低温环境.限制生长保存中, 低温保存是较常用的方法, 不同种质资源对低温的耐受性不同.例如, 赤松(Pinus densiflora)的丛生芽适合在17°C保存, 在14°C长期保存会影响其恢复(
昼夜温差对文心兰试管苗生长的影响
1
2011
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
植物种质资源缓慢生长离体保存研究进展
1
2008
... 卓越红花槭(Acer rubrum cv. ‘Somerset’)隶属槭树科槭属, 是从北美红花槭(Acer rubrum)中选育的优良品种之一, 其干形优美, 叶色鲜艳, 红叶期长, 适应性强, 观赏价值高.山东省林木种质资源中心于2007年从美国引进该树种, 并开展了其组培快繁研究, 已建立了完善的组培快繁体系, 繁殖效率达8.0 (
Effect of reverse photoperiod on in vitro regene- ration and piperine production in Piper nigrum L
1
2014
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
Poplar acclimation to cold during in vitro conservation at low non-freezing temperature: metabolic and proteic changes
1
2000
... 卓越红花槭离体材料在25°C下生长规律可概括为: 前期以分化为主; 后期以营养生长为主, 分化能力有所减弱, 而且分化出的芽不能全部长大.原因可能有2种, 一是培养基内的养分和激素已耗尽, 无力供给更多的芽生长; 二是前期长成的植株挤占了有限的生长空间, 致使幼芽无法生长.而在8°C和15°C条件下, 生长规律有所不同, 外植体分化率和平均出芽数增长缓慢, 但是在最后2次调查中上述二指标接近25°C下的处理组, 说明低温限制了离体材料的生长, 但是离体材料也在逐渐适应低温环境.限制生长保存中, 低温保存是较常用的方法, 不同种质资源对低温的耐受性不同.例如, 赤松(Pinus densiflora)的丛生芽适合在17°C保存, 在14°C长期保存会影响其恢复(
Stem elongation and growth of Solanum tuberosum L. in vitro in response to photosynthetic photon flux, photoperiod and difference in photoperiod and dark period temperatures
1
1995
... 光照和昼夜温差是植物生长过程中的重要影响因素, 在组织培养研究中经常用光照和昼夜温差来调控生长状态及分化方向等.多数研究证明, 长时间、高强度的光照有助于苗的成长和干物质积累, 而短时间、低强度的光照有利于愈伤及胚性愈伤的形成以及某些次生代谢物的积累(
Propagation and conservation of Castilleja tenu- iflora Benth
1
2009
... 限制生长保存是以组培技术为基础发展起来的保存方式, 是离体保存的重要组成部分, 在种质资源保存中得到广泛应用.英国邱园千年种子库通过微体快繁技术繁育和保存了超过3 000份植物种质资源的离体材料(
Conservation in vitro of threatened plants—progress in the past decade
1
2006
... 限制生长保存是以组培技术为基础发展起来的保存方式, 是离体保存的重要组成部分, 在种质资源保存中得到广泛应用.英国邱园千年种子库通过微体快繁技术繁育和保存了超过3 000份植物种质资源的离体材料(
Micropropagation and in vitro conservation of Neoglaziovia variegata (Arr. Cam.) Mez, a fiber producing bromeliad from Brazil.
1
2009
... 限制生长保存是以组培技术为基础发展起来的保存方式, 是离体保存的重要组成部分, 在种质资源保存中得到广泛应用.英国邱园千年种子库通过微体快繁技术繁育和保存了超过3 000份植物种质资源的离体材料(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}