Abstract: Calculating the number of species on earth has always been an issue of extreme concern, and suspicious species are among the most important obstacles to solve. Presently, the type specimen is the only criteria for identifying the suspicious species but is difficult to observe and study effectively because of its special taxonomic position. Therefore, the mostly effective way is to search the original information for suspicious species in the type locality. As molecular biological method, DNA barcoding, has been widely used in species classification and identification and can be used to identify a suspicious species accurately and quickly. In this study, a suspicious species, Dryopteris yenpingensis, was used as an example. We used DNA barcode analysis with the molecular data for the material from the type locality and combined morphological traits to clarify the doubtful taxonomic status of D. yenpingensis. The results supported D. yenpingensis and D. liboensis as the same species, and D. liboensis should be treated as a synonym. This study provides a reference for clarifying the taxonomic status of other suspicious species.
Key words:Dryopteris yenpingensis ; type locality ; DNA barcode ; taxonomically suspicious species ; biodiversity
全世界有多少物种是整个科学界及全社会共同关注的问题(May, 1988, 1990; Mora et al., 2011)。然而, 要回答这个问题并不容易。一方面, 我们目前尚有大量的新物种有待发现(Scheffers et al., 2012); 另一方面, 已经发现和命名的物种中有多少物种为同物异名尚待进一步研究。据The Plant List网站在线数据统计, 目前已知的1 064 035个物种名称中, 仅有350 699个物种名称(占33%)被接受, 尚有67%的物种名称属于异名或分类学存疑的物种。近年来, 随着分子生物学技术的发展, 对于物种的分类学地位已有许多新认识。特别是随着DNA条形码技术的发展, 科学家们希望用特定的基因片段给每一个物种标上一个基因标签, 用于物种的识别和辨认(Hebert et al., 2003)。然而, 首先需要面对的一个问题就是谁来判定标记物种身份的正确性。
南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)是一个长期处于存疑状态的物种。1938年, 该种发表于《静生生物汇报》, 当时认为这是一个形态奇特的物种, 并与羽裂鳞毛蕨(D. integriloba C. Chr.)形态相似(Ching, 1938), 模式标本为H. H. Chung采集的3508号标本, 现保存在中国科学院植物研究所标本馆(PE)。该种的模式产地位于福建省(Fokien)延平县(Yeping Hsien)半坎(Buong Kan) (据考证现今地址为福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩或半坎)。该种发表后曾被《福建植物志》(第1卷)收录并给予中文名称南平鳞毛蕨, 将其学名改为D. nanpingensis C. Chr. et Ching, 并为该物种绘制了线描图(《福建植物志》编写组, 1991)。但是《中国植物志》出版时, 该种并没有被收录或记载。Flora of China编者因缺少足够的信息而将该种置于Excluded taxa中作为存疑物种(Zhang et al., 2013)。最近出版的《中国生物物种名录——蕨类植物》同样将该种处理为存疑物种(严岳鸿等, 2016)。因此, 该物种的分类学地位成为分类学史上的一桩长期悬而未决的疑案。
2012年, Zhang等(2012)发表贵州蕨类植物新种荔波鳞毛蕨(D. liboensis P. S. Wang, X. Y. Wang et L. B. Zhang), 并认为该种与南平鳞毛蕨形态相似。2017年, 南平鳞毛蕨在《中国茂兰石松类和蕨类植物》一书中再次出现, 张宪春和姚正明(2017)将贵州荔波产的荔波鳞毛蕨鉴定为该种, 并对荔波产的南平鳞毛蕨叶绿体DNA进行测序且构建了该物种的叶绿体DNA系统关系, 将荔波鳞毛蕨处理为南平鳞毛蕨的异名。然而, 鉴定为南平鳞毛蕨的物种DNA序列并不是来自模式标本或模式产地, 因此又一个新的问题出现: 南平鳞毛蕨和荔波鳞毛蕨确定是同一物种吗?为了澄清这一问题, 笔者专程去南平鳞毛蕨的模式产地采集材料并对其叶绿体DNA进行测序, 以期通过DNA条形码技术鉴定和澄清南平鳞毛蕨的分类学地位。
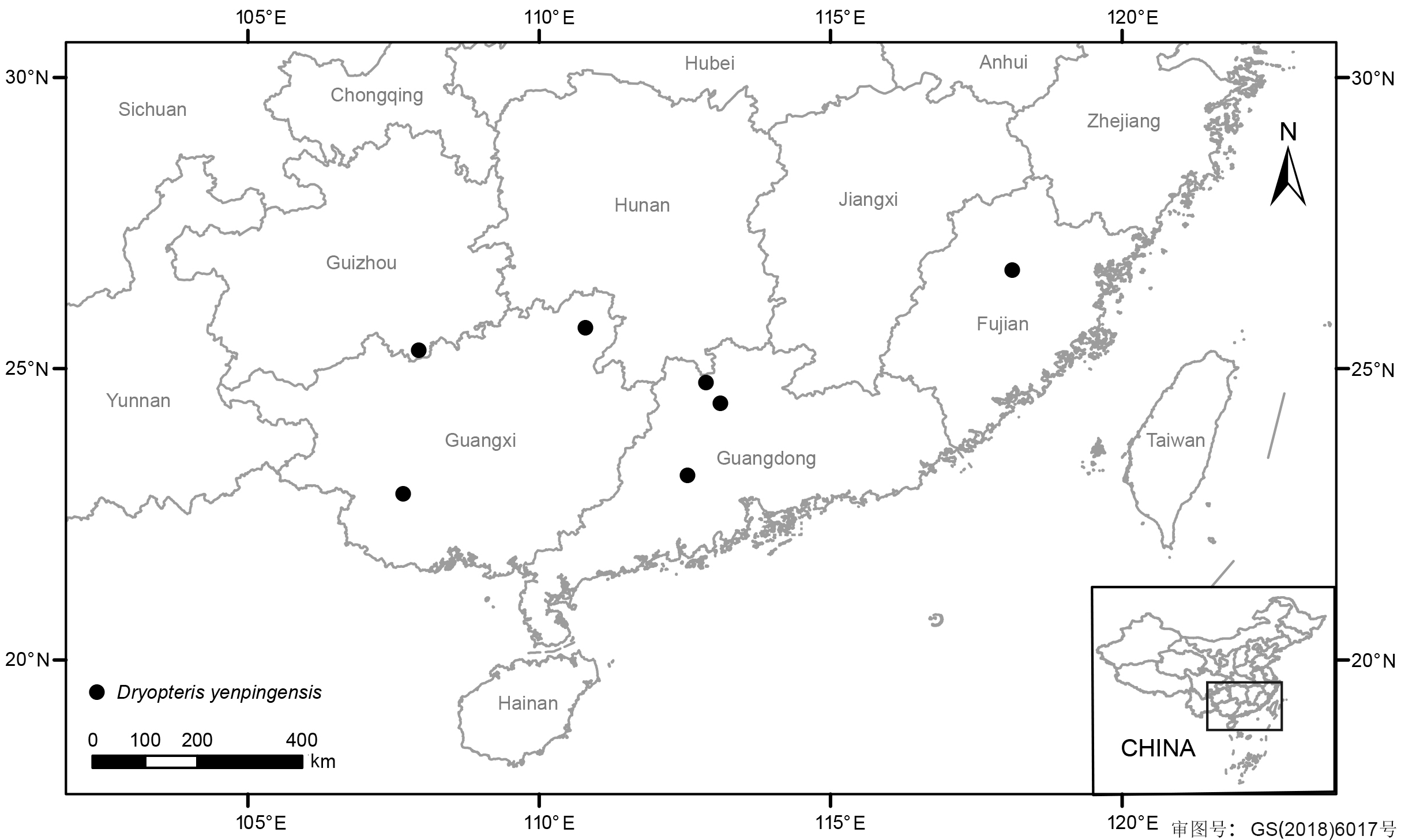
1 材料与方法1.1 研究材料本研究所用南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)来自福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩(或半坎)中途一条废弃的古道。经考证, 此地点应为该物种发表时所记录的地点, 新采集的南平鳞毛蕨位于古道旁边干旱的岩石缝中。同时, 笔者发现该种在广东肇庆鼎湖山山顶也有分布, 同时采集了此处的样本, 并查询了该物种在中国其它地区的采集记录, 绘制地理分布图。本研究使用的材料信息见表1。 表 1 Table 1 表 1 表 1 本研究所用材料的标本信息 Table 1 The information of the specimens in present study
Voucher number
Species name
Collection site
ZXC7305
Dryopteris liboensis
Guizhou, China
ZXC7273
D. liboensis
Guizhou, China
WYD043
D. liboensis
Guangdong, China
WYD066
D. integriloba
Guangdong, China
JSL5149
D. yenpingensis
Fujian, China
JSL3693
D. integriloba
Guangxi, China
JSL5083
D. integriloba
Guangxi, China
表 1 本研究所用材料的标本信息 Table 1 The information of the specimens in present study
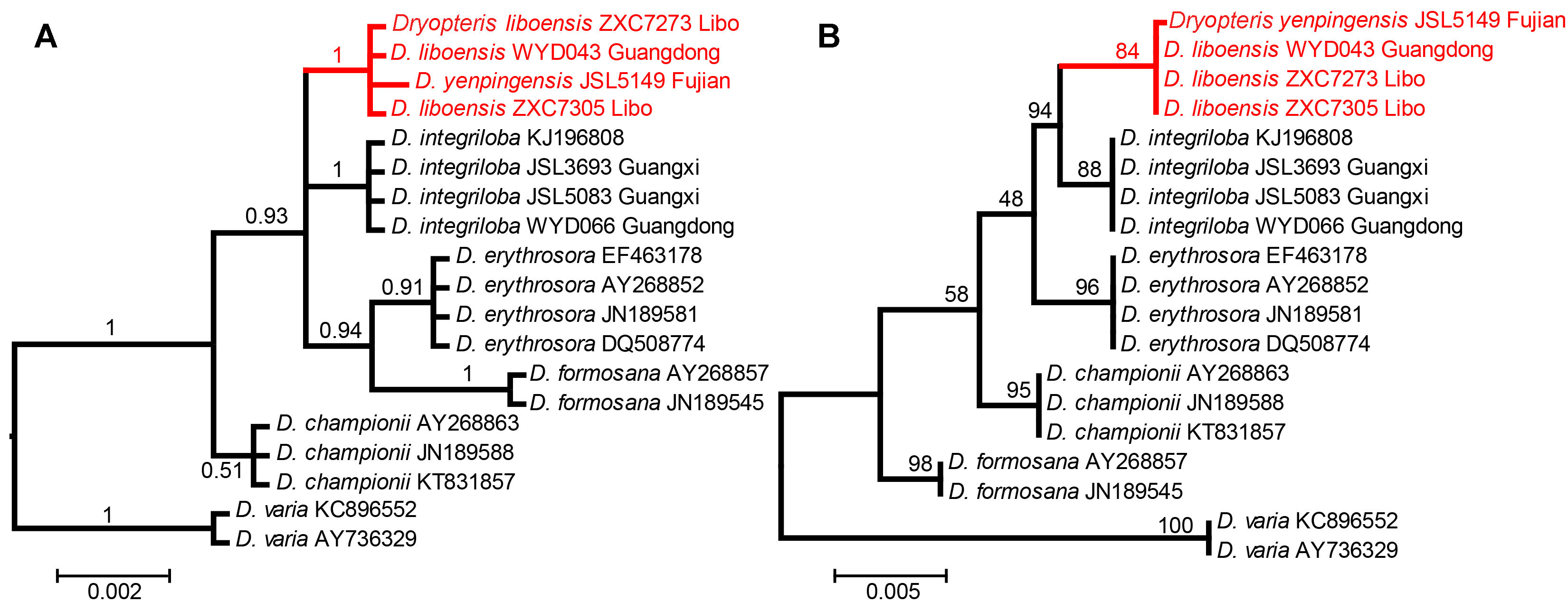
图 1 基于贝叶斯法(A)和邻接法(B)构建的系统发育树 Figure 1 The phylogenetic tree constructed by Bayesian inference (A) and neighbor-joining (B) methods
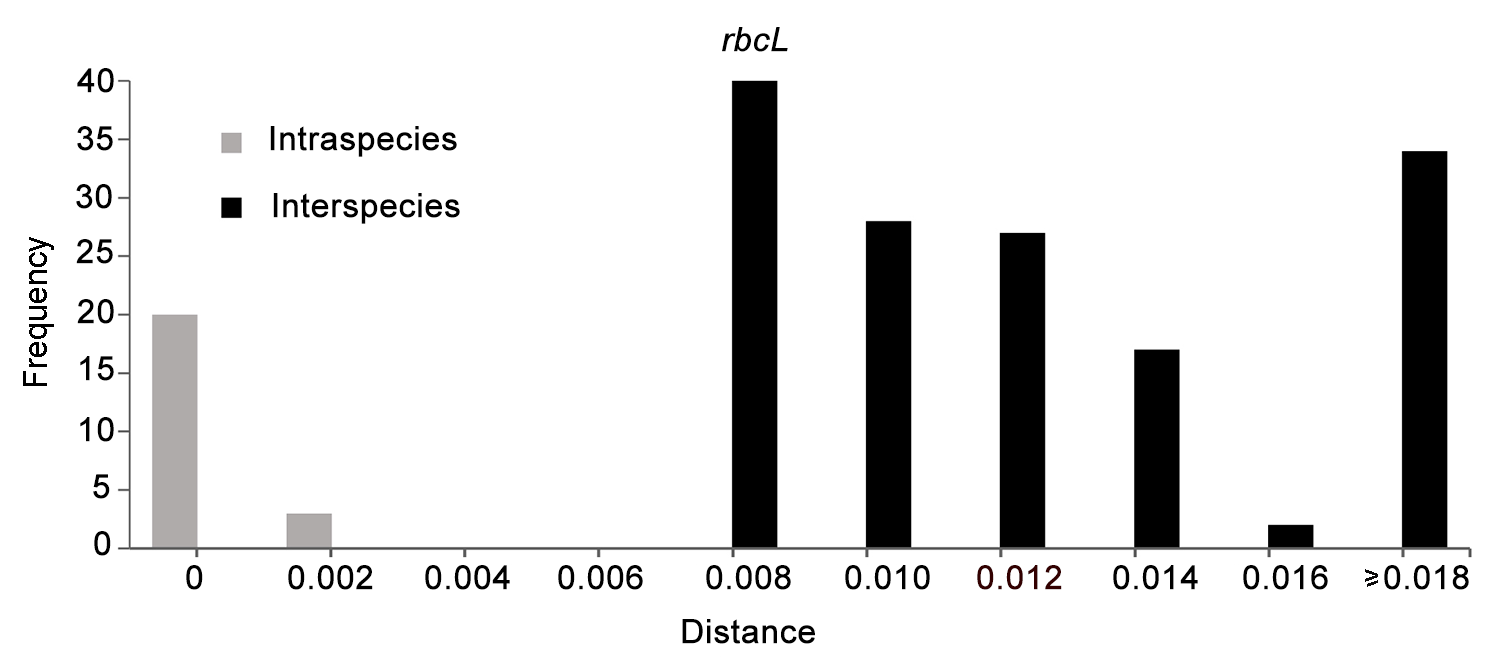
图 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-793/img_2.png<b>图 1</b> 基于<i>rbcL</i>序列的DNA条形码间隔<br/><b>Figure 2</b> The DNA barcoding gap based on <i>rbcL</i> Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-793/img_2.png<b>图 1</b> 基于<i>rbcL</i>序列的DNA条形码间隔<br/><b>Figure 2</b> The DNA barcoding gap based on <i>rbcL</i>
张宪春, 卫然, 刘红梅, 何丽娟, 王丽, 张钢民 (2013). 中国现代石松类和蕨类的系统发育与分类系统. 植物学报 48, 119-137. DOI:10.3724/SP.J.1259.2013.00119URLLycophytes and ferns are spore-bearing vascular plants, and are important groups in the evolution history of land plants. With the progress of molecular systematic, the relationships of the major groups have been resolved, and the traditional concepts have been revised. Meanwhile, new systematic arrangements have been proposed and are continuously being revised and updated. In the present review, the most recent phylogenetic studies were discussed and a new classification system of the extant lycophytes and ferns from China was also proposed. The system comprises five subclasses, 14 orders, 39 families with 12 subfamilies, and about 140 genera of lycophytes and ferns distributed in China. [本文引用: 1]
[13]
张宪春, 邢公侠 (1999). 纪念秦仁昌论文集. 北京: 中国林业出版社.
[本文引用: 1]
[14]
张宪春, 姚正明 (2017). 中国茂兰石松类和蕨类植物. 北京: 科学出版社.
[本文引用: 2]
[15]
CBOL Plant WorkingGroup (2009). A DNA barcode for land plants.Proc Natl Acad Sci USA 106, 12794-12797. DOI:10.1073/pnas.0905845106URL [本文引用: 1]
[16]
ChingRC (1931). The studies of Chinese ferns. V.Bull Fan Mem Inst Biol 2, 185-223. [本文引用: 1]
[17]
ChingRC (1938). A revision of the Chinese and Sikkim HimalayanDryopteris with reference to some species from neighbouring regions. Bull Fan Mem Inst Biol Bot Ser 8, 363-507. [本文引用: 1]
[18]
ChingRC (1941). New family and combinations of ferns.Bull Fan Mem Inst Biol Bot Ser 10, 235-256.
[19]
EbachMC, HoldregeC (2005). DNA barcoding is no substitute for taxonomy.Nature 434, 697. DOI:10.1038/434697bPMID:15815602URLNature is the international weekly journal of science: a magazine style journal that publishes full-length research papers in all disciplines of science, as well as News and Views, reviews, news, features, commentaries, web focuses and more, covering all branches of science and how science impacts upon all aspects of society and life. [本文引用: 1]
[20]
HallTA (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nucl Acids Symp Ser 41, 95-98. [本文引用: 1]
[21]
HebertPDN, CywinskaA, BallSL, deWaardJR (2003). Biological identifications through DNA barcodes.Proc Roy Soc B: Biol Sci 270, 313-321. DOI:10.1098/rspb.2002.2218URL [本文引用: 1]
[22]
HollingsworthPM, GrahamSW, LittleDP (2011). Choosing and using a plant DNA barcode.PLoS One 6, e19254. DOI:10.1371/journal.pone.0019254URL [本文引用: 1]
[23]
HolttumRE (1988). Studies in the fern genera allied to Tectaria Cav. VII. Species of Tectaria sect. Sagenia(Presl) Holttum in Asia excluding Malesia. Kew Bull 43, 475-489. DOI:10.2307/4118979URLThe 23 species here enumerated have in common either entirely free veins or the venation shown by Presl (1836) t.II, fig. 23-25; T.fuscipes bridges the gap between the two conditions. Two species are described as new (T. perdimorpha and T. lacei), most are confined, or almost so, to continental Asia and Ceylon, but four (T. barberi, T. devexa, T. dissecta and T. griffithii) have their main distribution in Malesia. A key to all species is provided, also brief descriptions and synonymy (or reference to the latest adequate one) for those which are not new. T. viridifrons Ching is placed as a synonym of T. coadunata (J. Sm.) C. Chr. with comment on the variability of the latter. New combinations are: T. setulosa (Bak.) Holttum and T. impressa (Fe) Holttum. [本文引用: 1]
[24]
LiouH (1934). Lauracées de Chine et d’Indochine: Contribution à l’étude Systématique et Phytogéographique. Paris: Hermann et Cie. [本文引用: 1]
MayRM (1990). How many species?Philos Trans Roy Soc B: Biol Sci 330, 293-304. DOI:10.1098/rstb.1990.0200URL [本文引用: 1]
[27]
MeusnierI, SingerGAC, LandryJF, HickeyDA, HebertPDN, HajibabaeiM (2008). A universal DNA mini- barcode for biodiversity analysis.BMC Genom 9, 214. DOI:10.1186/1471-2164-9-214URL [本文引用: 1]
[28]
MoraC, TittensorDP, AdlS, SimpsonAGB, WormB (2011). How many species are there on earth and in the ocean?PLoS Biol 9, e1001127. DOI:10.1371/journal.pbio.1001127PMID:21886479URLThe diversity of life is one of the most striking aspects of our planet; hence knowing how many species inhabit Earth is among the most fundamental questions in science. Yet the answer to this question remains enigmatic, as efforts to sample the world's biodiversity to date have been limited and thus have precluded direct quantification of global species richness, and because indirect estimates rely on assumptions that have proven highly controversial. Here we show that the higher taxonomic classification of species (i.e., the assignment of species to phylum, class, order, family, and genus) follows a consistent and predictable pattern from which the total number of species in a taxonomic group can be estimated. This approach was validated against well-known taxa, and when applied to all domains of life, it predicts ~8.7 million (??1.3 million SE) eukaryotic species globally, of which ~2.2 million (??0.18 million SE) are marine. In spite of 250 years of taxonomic classification and over 1.2 million species already catalogued in a central database, our results suggest that some 86% of existing species on Earth and 91% of species in the ocean still await description. Renewed interest in further exploration and taxonomy is required if this significant gap in our knowledge of life on Earth is to be closed. [本文引用: 1]
ScheffersBR, JoppaLN, PimmSL, LauranceWF (2012). What we know and don’t know about Earth’s missing biodiversity.Trends Ecol Evol 27, 501-510. DOI:10.1016/j.tree.2012.05.008URL [本文引用: 1]
[31]
SchindelDE, MillerSE (2005). DNA barcoding a useful tool for taxonomists.Nature 435, 17. [本文引用: 1]
[32]
ShuJP, ShangH, JinDM, WeiHJ, ZhouXL, LiuHM, GuYF, WangY, WangFG, ShenH, ZhangR, AdjieB, YanYH (2017). Re-establishment of species from synonymies based on DNA barcoding and phylogenetic analysis using Diplopterygium simulans(Gleicheniaceae) as an example. PLoS One 12, e0164604. DOI:10.1371/journal.pone.0164604PMID:5351838URLAbstract Because synonymy treatment traditionally relies on morphological judgments, it usually causes many problems in species delimitation and in the biodiversity catalogue. For example, Diplopterygium simulans, which belongs to the Gleicheniaceae family, has been considered to be synonymous with D. glaucum or D. giganteum based mainly on the morphology of its pinna rachis and blade. In the absence of molecular evidence, these revisions remain doubtful. DNA barcoding, which is considered to be a powerful method for species-level identification, was employed to assess the genetic distance among 9 members of the Diplopterygium genus. The results indicate that D. simulans is an independent species rather than a synonymy of D. glaucum or D. giganteum. Moreover, phylogenetic analysis uncovered the sisterhood of D. simulans and D. cantonense, which is supported by their geographical distributions and morphological traits. Incorrect synonymy treatment is prevalent in the characterization of biological diversity, and our study proposes a convenient and effective method for validating synonym treatments and discovering cryptic species. [本文引用: 1]
[33]
TamuraK, StecherG, PetersonD, FilipskiA, KumarS (2013). MEGA6: molecular evolutionary genetics analysis version 6.0.Mol Biol Evol 30, 2725-2729. DOI:10.1093/molbev/mst197PMID:24132122URLWe announce the release of an advanced version of the Molecular Evolutionary Genetics Analysis (MEGA) software, which currently contains facilities for building sequence alignments, inferring phylogenetic histories, and conducting molecular evolutionary analysis. In version 6.0, MEGA now enables the inference of timetrees, as it implements the RelTime method for estimating divergence times for all branching points in a phylogeny. A new Timetree Wizard in MEGA6 facilitates this timetree inference by providing a graphical user interface (GUI) to specify the phylogeny and calibration constraints step-by-step. This version also contains enhanced algorithms to search for the optimal trees under evolutionary criteria and implements a more advanced memory management that can double the size of sequence data sets to which MEGA can be applied. Both GUI and command-line versions of MEGA6 can be downloaded from www.megasoftware.net free of charge. [本文引用: 1]
[34]
TryonAF, LugardonB (1991). Spores of the Pteridophyta. New York: Springer.
[35]
WeiFN, HenkW (2008). Machilus. In: Wu ZY, Raven PH, Hong DY, eds. Flora of China, Vol. 7. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press. pp. 201-224. [本文引用: 1]
[36]
ZhangLB, WangPS, WangXY (2012). Dryopteris liboensis(Dryopteridaceae), a new species in Dryopteris sect. Erythrovariae from Guizhou, China. Novon 22, 256-259. [本文引用: 1]
... 南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)是一个长期处于存疑状态的物种.1938年, 该种发表于《静生生物汇报》, 当时认为这是一个形态奇特的物种, 并与羽裂鳞毛蕨(D. integriloba C. Chr.)形态相似(Ching, 1938), 模式标本为H. H. Chung采集的3508号标本, 现保存在中国科学院植物研究所标本馆(PE).该种的模式产地位于福建省(Fokien)延平县(Yeping Hsien)半坎(Buong Kan) (据考证现今地址为福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩或半坎).该种发表后曾被《福建植物志》(第1卷)收录并给予中文名称南平鳞毛蕨, 将其学名改为D. nanpingensis C. Chr. et Ching, 并为该物种绘制了线描图(《福建植物志》编写组, 1991).但是《中国植物志》出版时, 该种并没有被收录或记载.Flora of China编者因缺少足够的信息而将该种置于Excluded taxa中作为存疑物种(Zhang et al., 2013).最近出版的《中国生物物种名录——蕨类植物》同样将该种处理为存疑物种(严岳鸿等, 2016).因此, 该物种的分类学地位成为分类学史上的一桩长期悬而未决的疑案. ...
... DNA条形码概念的提出, 为实现物种快速准确识别与鉴定提供了新方法, 极大地促进了分类学、演化生物学、生态学和保护生物学等相关学科的发展.在动物中, 线粒体内的细胞色素C氧化酶亚基1 (cytochrome C oxidase subunit 1, CO1)已成功应用于绝大多数动物类群(Meusnier et al., 2008).而在植物中, 暂时没有唯一的单个片段的DNA条形码.生命条形码联盟植物工作组(CBOL Plant Working Group, 2009)建议将rbcL和matK组合作为陆地植物核心DNA条形码.此后, trnH-psbA和ITS被建议作为补充候选片段(Hollingsworth et al., 2011).rbcL基因作为目前植物分子系统学研究中应用最广泛且最重要的分子条形码, 已被应用于蕨类植物各个分类阶元的系统学研究(李春香和陆树刚, 2006; 刘红梅等, 2009; 王凡红等, 2014). ...
鳞毛蕨科的孢子形态研究 2007
铁线蕨属3个种的分布新记录 1 2014
... DNA条形码概念的提出, 为实现物种快速准确识别与鉴定提供了新方法, 极大地促进了分类学、演化生物学、生态学和保护生物学等相关学科的发展.在动物中, 线粒体内的细胞色素C氧化酶亚基1 (cytochrome C oxidase subunit 1, CO1)已成功应用于绝大多数动物类群(Meusnier et al., 2008).而在植物中, 暂时没有唯一的单个片段的DNA条形码.生命条形码联盟植物工作组(CBOL Plant Working Group, 2009)建议将rbcL和matK组合作为陆地植物核心DNA条形码.此后, trnH-psbA和ITS被建议作为补充候选片段(Hollingsworth et al., 2011).rbcL基因作为目前植物分子系统学研究中应用最广泛且最重要的分子条形码, 已被应用于蕨类植物各个分类阶元的系统学研究(李春香和陆树刚, 2006; 刘红梅等, 2009; 王凡红等, 2014). ...
2010
扫描电镜下真蕨目孢子表面纹饰的分类 2003
1 1982
... 在蕨类植物中, 诸如南平鳞毛蕨被《中国植物志》遗漏的例子并非孤例.它们如同分类学名录中的幽灵, 存在众多的物种名录中, 但需清理并开展进一步的深入考察和研究.例如, 蕨类植物中广东叉蕨(Tectaria kwangtungensis Ching)于1931年发表在《静生生物研究所集刊》(Ching, 1931), 模式标本采自广东连州(Kwangtung: Lienchow, C. G. Mattew (Type), Dec. 6, 1907); 之后, Ching (1941)又将该种处理为Pleocnemia kwangtungensis (Ching) Ching; 1982年在《蕨类植物名词与名称》(邢公侠, 1982)中处理为Ctenitopsis kwangtungensis (Ching) Ching et C. H. Wang; Holttum (1988)再次将该种处理为T. kwangtungensis Ching.1999年出版的《纪念秦仁昌论文集》中, 该种以3个不同的名称C. kwangtungensis (Ching) Ching et C. H. Wang in Shing (pp 455)、P. kwangtungensis (Ching) Ching (pp 486)及T. kw- angtungensis Ching (pp 498)各自独立出现(张宪春和邢公侠, 1999).然而此后出版的《中国植物志》、Flora of China及《中国生物物种名录》均没有对该名称进行科学处理, 至今仍未见新的采集报道.据《中国生物物种名录——蕨类植物》记载, 目前中国已知有96种存疑物种需要进一步研究(严岳鸿等, 2016). ...
2
... 南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)是一个长期处于存疑状态的物种.1938年, 该种发表于《静生生物汇报》, 当时认为这是一个形态奇特的物种, 并与羽裂鳞毛蕨(D. integriloba C. Chr.)形态相似(Ching, 1938), 模式标本为H. H. Chung采集的3508号标本, 现保存在中国科学院植物研究所标本馆(PE).该种的模式产地位于福建省(Fokien)延平县(Yeping Hsien)半坎(Buong Kan) (据考证现今地址为福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩或半坎).该种发表后曾被《福建植物志》(第1卷)收录并给予中文名称南平鳞毛蕨, 将其学名改为D. nanpingensis C. Chr. et Ching, 并为该物种绘制了线描图(《福建植物志》编写组, 1991).但是《中国植物志》出版时, 该种并没有被收录或记载.Flora of China编者因缺少足够的信息而将该种置于Excluded taxa中作为存疑物种(Zhang et al., 2013).最近出版的《中国生物物种名录——蕨类植物》同样将该种处理为存疑物种(严岳鸿等, 2016).因此, 该物种的分类学地位成为分类学史上的一桩长期悬而未决的疑案. ... ... 在蕨类植物中, 诸如南平鳞毛蕨被《中国植物志》遗漏的例子并非孤例.它们如同分类学名录中的幽灵, 存在众多的物种名录中, 但需清理并开展进一步的深入考察和研究.例如, 蕨类植物中广东叉蕨(Tectaria kwangtungensis Ching)于1931年发表在《静生生物研究所集刊》(Ching, 1931), 模式标本采自广东连州(Kwangtung: Lienchow, C. G. Mattew (Type), Dec. 6, 1907); 之后, Ching (1941)又将该种处理为Pleocnemia kwangtungensis (Ching) Ching; 1982年在《蕨类植物名词与名称》(邢公侠, 1982)中处理为Ctenitopsis kwangtungensis (Ching) Ching et C. H. Wang; Holttum (1988)再次将该种处理为T. kwangtungensis Ching.1999年出版的《纪念秦仁昌论文集》中, 该种以3个不同的名称C. kwangtungensis (Ching) Ching et C. H. Wang in Shing (pp 455)、P. kwangtungensis (Ching) Ching (pp 486)及T. kw- angtungensis Ching (pp 498)各自独立出现(张宪春和邢公侠, 1999).然而此后出版的《中国植物志》、Flora of China及《中国生物物种名录》均没有对该名称进行科学处理, 至今仍未见新的采集报道.据《中国生物物种名录——蕨类植物》记载, 目前中国已知有96种存疑物种需要进一步研究(严岳鸿等, 2016). ...
... 在蕨类植物中, 诸如南平鳞毛蕨被《中国植物志》遗漏的例子并非孤例.它们如同分类学名录中的幽灵, 存在众多的物种名录中, 但需清理并开展进一步的深入考察和研究.例如, 蕨类植物中广东叉蕨(Tectaria kwangtungensis Ching)于1931年发表在《静生生物研究所集刊》(Ching, 1931), 模式标本采自广东连州(Kwangtung: Lienchow, C. G. Mattew (Type), Dec. 6, 1907); 之后, Ching (1941)又将该种处理为Pleocnemia kwangtungensis (Ching) Ching; 1982年在《蕨类植物名词与名称》(邢公侠, 1982)中处理为Ctenitopsis kwangtungensis (Ching) Ching et C. H. Wang; Holttum (1988)再次将该种处理为T. kwangtungensis Ching.1999年出版的《纪念秦仁昌论文集》中, 该种以3个不同的名称C. kwangtungensis (Ching) Ching et C. H. Wang in Shing (pp 455)、P. kwangtungensis (Ching) Ching (pp 486)及T. kw- angtungensis Ching (pp 498)各自独立出现(张宪春和邢公侠, 1999).然而此后出版的《中国植物志》、Flora of China及《中国生物物种名录》均没有对该名称进行科学处理, 至今仍未见新的采集报道.据《中国生物物种名录——蕨类植物》记载, 目前中国已知有96种存疑物种需要进一步研究(严岳鸿等, 2016). ...
2 2017
... 2012年, Zhang等(2012)发表贵州蕨类植物新种荔波鳞毛蕨(D. liboensis P. S. Wang, X. Y. Wang et L. B. Zhang), 并认为该种与南平鳞毛蕨形态相似.2017年, 南平鳞毛蕨在《中国茂兰石松类和蕨类植物》一书中再次出现, 张宪春和姚正明(2017)将贵州荔波产的荔波鳞毛蕨鉴定为该种, 并对荔波产的南平鳞毛蕨叶绿体DNA进行测序且构建了该物种的叶绿体DNA系统关系, 将荔波鳞毛蕨处理为南平鳞毛蕨的异名.然而, 鉴定为南平鳞毛蕨的物种DNA序列并不是来自模式标本或模式产地, 因此又一个新的问题出现: 南平鳞毛蕨和荔波鳞毛蕨确定是同一物种吗?为了澄清这一问题, 笔者专程去南平鳞毛蕨的模式产地采集材料并对其叶绿体DNA进行测序, 以期通过DNA条形码技术鉴定和澄清南平鳞毛蕨的分类学地位. ... ... 长期被当作存疑物种处理.原因是其生境特殊, 在野外难以发现, 可供研究的材料非常少.笔者专程前往模式标本产地, 经过仔细查找, 终于在非常隐蔽的岩石缝中发现该种.经过仔细研究, 发现荔波鳞毛蕨与南平鳞毛蕨在形态特征上并没有明显区别, 二者都是线状披针形的狭鳞片, 生于叶柄基部(图3), 是泡鳞亚属中比较特殊的类群.而且, DNA条形码分析也支持荔波鳞毛蕨与南平鳞毛蕨为同一个单系(图1), 作为一个物种, 与其它物种间存在明显的遗传间隔(图2).综合形态和分子证据表明, 荔波鳞毛蕨和南平鳞毛蕨确实是同一物种, 再次佐证了张宪春和姚正明(2017)将荔波鳞毛蕨处理为南平鳞毛蕨的异名的观点. ...
1 2009
... DNA条形码概念的提出, 为实现物种快速准确识别与鉴定提供了新方法, 极大地促进了分类学、演化生物学、生态学和保护生物学等相关学科的发展.在动物中, 线粒体内的细胞色素C氧化酶亚基1 (cytochrome C oxidase subunit 1, CO1)已成功应用于绝大多数动物类群(Meusnier et al., 2008).而在植物中, 暂时没有唯一的单个片段的DNA条形码.生命条形码联盟植物工作组(CBOL Plant Working Group, 2009)建议将rbcL和matK组合作为陆地植物核心DNA条形码.此后, trnH-psbA和ITS被建议作为补充候选片段(Hollingsworth et al., 2011).rbcL基因作为目前植物分子系统学研究中应用最广泛且最重要的分子条形码, 已被应用于蕨类植物各个分类阶元的系统学研究(李春香和陆树刚, 2006; 刘红梅等, 2009; 王凡红等, 2014). ...
1 1931
... 在蕨类植物中, 诸如南平鳞毛蕨被《中国植物志》遗漏的例子并非孤例.它们如同分类学名录中的幽灵, 存在众多的物种名录中, 但需清理并开展进一步的深入考察和研究.例如, 蕨类植物中广东叉蕨(Tectaria kwangtungensis Ching)于1931年发表在《静生生物研究所集刊》(Ching, 1931), 模式标本采自广东连州(Kwangtung: Lienchow, C. G. Mattew (Type), Dec. 6, 1907); 之后, Ching (1941)又将该种处理为Pleocnemia kwangtungensis (Ching) Ching; 1982年在《蕨类植物名词与名称》(邢公侠, 1982)中处理为Ctenitopsis kwangtungensis (Ching) Ching et C. H. Wang; Holttum (1988)再次将该种处理为T. kwangtungensis Ching.1999年出版的《纪念秦仁昌论文集》中, 该种以3个不同的名称C. kwangtungensis (Ching) Ching et C. H. Wang in Shing (pp 455)、P. kwangtungensis (Ching) Ching (pp 486)及T. kw- angtungensis Ching (pp 498)各自独立出现(张宪春和邢公侠, 1999).然而此后出版的《中国植物志》、Flora of China及《中国生物物种名录》均没有对该名称进行科学处理, 至今仍未见新的采集报道.据《中国生物物种名录——蕨类植物》记载, 目前中国已知有96种存疑物种需要进一步研究(严岳鸿等, 2016). ...
1 1938
... 南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)是一个长期处于存疑状态的物种.1938年, 该种发表于《静生生物汇报》, 当时认为这是一个形态奇特的物种, 并与羽裂鳞毛蕨(D. integriloba C. Chr.)形态相似(Ching, 1938), 模式标本为H. H. Chung采集的3508号标本, 现保存在中国科学院植物研究所标本馆(PE).该种的模式产地位于福建省(Fokien)延平县(Yeping Hsien)半坎(Buong Kan) (据考证现今地址为福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩或半坎).该种发表后曾被《福建植物志》(第1卷)收录并给予中文名称南平鳞毛蕨, 将其学名改为D. nanpingensis C. Chr. et Ching, 并为该物种绘制了线描图(《福建植物志》编写组, 1991).但是《中国植物志》出版时, 该种并没有被收录或记载.Flora of China编者因缺少足够的信息而将该种置于Excluded taxa中作为存疑物种(Zhang et al., 2013).最近出版的《中国生物物种名录——蕨类植物》同样将该种处理为存疑物种(严岳鸿等, 2016).因此, 该物种的分类学地位成为分类学史上的一桩长期悬而未决的疑案. ...
... 2012年, Zhang等(2012)发表贵州蕨类植物新种荔波鳞毛蕨(D. liboensis P. S. Wang, X. Y. Wang et L. B. Zhang), 并认为该种与南平鳞毛蕨形态相似.2017年, 南平鳞毛蕨在《中国茂兰石松类和蕨类植物》一书中再次出现, 张宪春和姚正明(2017)将贵州荔波产的荔波鳞毛蕨鉴定为该种, 并对荔波产的南平鳞毛蕨叶绿体DNA进行测序且构建了该物种的叶绿体DNA系统关系, 将荔波鳞毛蕨处理为南平鳞毛蕨的异名.然而, 鉴定为南平鳞毛蕨的物种DNA序列并不是来自模式标本或模式产地, 因此又一个新的问题出现: 南平鳞毛蕨和荔波鳞毛蕨确定是同一物种吗?为了澄清这一问题, 笔者专程去南平鳞毛蕨的模式产地采集材料并对其叶绿体DNA进行测序, 以期通过DNA条形码技术鉴定和澄清南平鳞毛蕨的分类学地位. ...
1 2013
... 南平鳞毛蕨(Dryopteris yenpingensis C. Chr. et Ching)是一个长期处于存疑状态的物种.1938年, 该种发表于《静生生物汇报》, 当时认为这是一个形态奇特的物种, 并与羽裂鳞毛蕨(D. integriloba C. Chr.)形态相似(Ching, 1938), 模式标本为H. H. Chung采集的3508号标本, 现保存在中国科学院植物研究所标本馆(PE).该种的模式产地位于福建省(Fokien)延平县(Yeping Hsien)半坎(Buong Kan) (据考证现今地址为福建省南平市延平区茫荡山闽赣古道三千八百坎的半岩或半坎).该种发表后曾被《福建植物志》(第1卷)收录并给予中文名称南平鳞毛蕨, 将其学名改为D. nanpingensis C. Chr. et Ching, 并为该物种绘制了线描图(《福建植物志》编写组, 1991).但是《中国植物志》出版时, 该种并没有被收录或记载.Flora of China编者因缺少足够的信息而将该种置于Excluded taxa中作为存疑物种(Zhang et al., 2013).最近出版的《中国生物物种名录——蕨类植物》同样将该种处理为存疑物种(严岳鸿等, 2016).因此, 该物种的分类学地位成为分类学史上的一桩长期悬而未决的疑案. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}