王韵茜 ,
苏延红 ,
杨睿 ,
李鑫 ,
李晶 ,
曾千春 * , ,
罗琼 * , 云南农业大学, 农业生物多样性与病虫害控制教育部重点实验室, 云南生物资源保护与利用国家重点实验室, 昆明 650201
Wang Yunqian ,
Su Yanhong ,
Yang Rui ,
Li Xin ,
Li Jing ,
Zeng Qianchun * , ,
Luo Qiong * , State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, Ministry of Education Key Laboratory of Agriculture Biodiversity for Plant Disease Management, Yunnan Agricultural University, Kunming 650201, China 引用本文 王韵茜 ,
苏延红 ,
杨睿 ,
李鑫 ,
李晶 ,
曾千春 ,
罗琼 . 云南疣粒野生稻稻瘟病抗性. 植物学报, 2018, 53(4): 477-486
贡献者
* 通讯作者。E-mail: zengqianchun@qq.com * 通讯作者。E-mail: qiongbf@aliyun.com † 共同第一作者。 基金资助
国家重点研发计划(No.2016YFD0100600)和国家自然科学基金-云南联合基金(No.U1302261) ;
接受日期:2017-06-5接受日期:2017-08-7网络出版日期:2018-07-1
-->Copyright
2018《植物学报》编辑部
Contributors
* Authors for correspondence. E-mail: zengqianchun@qq.com * Authors for correspondence. E-mail: qiongbf@aliyun.com † These authors contributed equally to this paper History
Received :
Accepted :
Online :
摘要: 野生稻(Oryza rufipogo )保存有许多栽培稻(O. sativa )不具备或已经消失的优异基因资源, 是扩大栽培稻遗传背景、改良产量与品质、提高抗病虫害及抗逆境能力的重要基因库。疣粒野生稻(O. meyeriana )是中国3种野生稻资源之一, 主要分布在云南。为进一步了解其稻瘟病抗性, 首先利用来自不同稻作区的稻瘟病菌株, 通过注射接种法对疣粒野生稻进行系统的稻瘟病抗性鉴定, 发现疣粒野生稻对接种的所有稻瘟病菌株都感病。进一步采用3'/5' RACE方法, 从疣粒野生稻中克隆了水稻同源基因Pid2 和Pid3 , 并构建过表达转基因株系对基因功能进行了研究。结果表明, Pid2 和Pid3 与疣粒野生稻中同源基因间在DNA和氨基酸水平上有较大的序列差异, 过表达转基因的日本晴植株对稻瘟病菌的敏感性与对照相似。推测疣粒野生稻在自然接种条件下, 表现出的抗稻瘟病表型很可能是其旱生叶片结构特征形成了对稻瘟病菌侵染的天然屏障。对控制疣粒野生稻这一类性状基因资源的挖掘和利用, 有利于优良抗性水稻品种的培育。研究结果为疣粒野生稻的研究利用提供了新信息和新思路。 关键词 :
疣粒野生稻 ;
稻瘟病抗性 ;
抗病基因 ;
基因克隆 ;
稻瘟病菌 Abstract :
Wild species of Oryza contain numerous genes of economic importance and are being used as an important cultivated rice gene pool to expand the genetic background, improve yield and quality, and increase the ability for resistance or tolerance to pests as well as bioic and abiotic stresses. O. meyeriana is one of three native wild rice species in China, mainly distributed in Yunnan province. To further understand the disease resistance to rice blast, we performed a systematic investigation of rice blast resistance with O. meyeriana . O. meyeriana plants were inoculated by injection with multiple blast strains from different rice planting areas. Then, we cloned and functionally identified Pid2 and Pid3 orthologous genes in O. meyeriana . O. meyeriana was sensitive to all blast isolates used in this experiment. The DNA and amino acid sequences significantly differed between Pid2 and its orthologs in O. meyeriana , as did Pid3 and its orthologs. Nipponbare plants overexpressing the orthologous genes of Pid2 and Pid3 showed mildly enhanced susceptibility to Magnaporthe oryzae . The orthologs of blast-resistant genes in O. meyeriana may be susceptible alleles. O. meyeriana exhibited resistance to rice blast under natural inoculation likely due to a xeromorphic leaf structure that reduces susceptibility to M. oryzae . Characterization of these genes will help breed elite rice. Our study provides new information and ideas for research and utilization of O. meyeriana . Key words :
wild rice (Oryza meyeriana ) ;
rice blast resistance ;
R gene ;
gene cloning ;
Magnaporthe oryzae 稻瘟病是由稻瘟病菌(
Magnaporthe oryzae )引发并能够侵染水稻(
Oryza sativa )整个生育期的真菌病害, 是水稻生产上最为严重的病害之一。其分布广泛, 危害严重, 造成年均10%-30%的产量损失, 严重影响稻米产量和品质, 威胁世界粮食安全(
Ska- mnioti and Gurr, 2009 ;
Liu et al., 2013 )。目前, 抗病品种的种植和化学农药的使用是防治该病害的主要措施。化学农药的大量使用带来了一系列包括环境污染和食品安全等问题, 而抗病品种的利用是防治稻瘟病最安全有效的措施。但由于稻瘟病菌群体组成复杂、变异快, 导致1个抗病品种大面积推广种植3-5年后抗性便逐渐丧失。为此, 鉴定新的抗病资源、克隆新的抗病基因, 是培育水稻抗病新品种的关键。
自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 )。已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用。例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 )。为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用。例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 )。
疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 )。
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性。
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功。
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆。迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制。
为了进一步了解疣粒野生稻的抗病特性, 有效挖掘和利用疣粒野生稻中的抗病基因资源, 本研究利用分离自不同水稻生态区的稻瘟病菌株(ZB13、ZB15、Zhong-10-8-14、ZE1、LP174、LP29-3和H53), 通过注射接种法对疣粒野生稻进行系统的稻瘟病抗性调查。采用3'/5'RACE方法, 从疣粒野生稻中克隆了
Pid2 (地谷中抗稻瘟病菌株ZB15的主效抗瘟基因) (
Chen et al., 2006 )和
Pid3 (地谷中抗稻瘟病菌株Zhong-10-8-14的主效抗瘟基因) (
Shang et al., 2009 )的同源基因, 并构建过表达转基因株系对基因功能进行研究。研究结果将为疣粒野生稻中优异基因资源的研究和利用提供新信息和新思路。
1 材料与方法1.1 实验材料植物材料: 云南疣粒野生稻(
Oryza meyeriana (Zoll. et Mor. ex. Steud.) Baill), 2013年采自云南省西双版纳嘎洒镇; 感病对照水稻日本晴(Nipponbare)由本实验室保存。
稻瘟病菌株: 7个稻瘟病菌株分别为来自四川籼稻区的优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1由四川省农科院植保所提供; LP174和LP29-3分离自云南罗平籼粳混合区, 由云南农业大学何月秋教授提供; H53为本实验室从2013年哈尔滨五常稻瘟病重灾区病样分离。所有菌株均经过实验室前期的鉴定和筛选。
1.2 方法
1.2.1 稻瘟病菌活化和产孢培养 在超净工作台上用灭菌镊子在酒精灯前挑取-80°C保存的菌株滤纸片于马铃薯葡萄糖平板培养基上; 28°C恒温箱内黑暗活化培养3天后, 用消毒接菌环挑取边沿适量新菌丝, 采用三点接菌法转接于新鲜的马铃薯葡萄糖平板培养基上; 28°C恒温箱内黑暗培养3天后, 用消毒接菌环挑取边沿适量新菌丝块于马铃薯葡萄糖液体培养基中; 28°C恒温振荡(每小时180转)培养2-3天后, 吸取500 μL菌液至番茄燕麦培养基上, 用涂布器将菌液均匀涂于培养基表面; 28°C自然光照培养3-4天后, 用无菌棉签刮去表面菌丝; 28°C自然 光照进行产孢培养3-5天后, 用无菌水洗下孢子, 3层擦镜纸过滤, 将孢子液浓度调制为5×10
4 个∙mL
-1 备用。
1.2.2 稻瘟病菌接种和发病调查 (1) 注射接种
疣粒野生稻注射接种: 取5株疣粒野生稻, 每株选取3个发育良好的孕穗期分蘖, 用1 mL医用注射器在展开的剑叶叶鞘处成40°角向上缓慢注入浓度为5×10
4 个∙mL
-1 的孢子悬浮液, 至喇叭口处有菌液流出时为止, 同时用无菌蒸馏水作对照接种。接种在自然光照下, 25-28°C温室中进行。接种7天后进行发病调查。转基因水稻株系注射接种: 分蘖期进行叶鞘注射接种, 方法同上。
按照
Tharreau (2000) 的6级分类法进行病斑类型调查。0级(HR): 无任何病斑或者不侵染; 1级(R): 有零星直径<0.5 mm的褐色病斑; 2级(MR): 有少数直径为0.5-1 mm的褐色病斑; 3级(MS): 出现直径<3 mm的圆形至椭圆形病斑, 中央灰白色, 边缘褐色; 4级(S): 典型的纺锤形稻瘟病斑, 直径3 mm或更长, 病斑稍有融合或无融合; 5级(HS): 典型的纺锤形稻瘟病斑, 直径3 mm或更长病斑融合成片。
(2) 喷雾接种
用无菌水配制浓度为5×10
4 个∙mL
-1 的稻瘟病菌孢子悬浮液, 加入0.5%的吐温混匀, 喷雾接种3-4叶期水稻幼苗或2-3叶期疣粒野生稻新蘖, 在25-28 °C, 相对湿度95%以上的培养箱中黑暗培育24小时。然后保持温湿度不变, 在12小时光照/12小时黑暗条件下培育7天后调查发病情况。
叶瘟发病分级参照国际水稻研究所的标准评价系统(
Standard Evaluation System for Rice, SES, 2013 ), 即病斑类型分6级。0级: 无任何病斑; 1级: 仅有针尖大小褐点或褐点稍大; 3级: 有小的圆形坏死斑, 直径约为1-2 mm, 有明显褐色边缘或黄色晕; 5级: 出现宽1-2 mm、直径>3 mm的圆形至椭圆形病斑, 边缘褐色; 7级: 出现典型的纺锤形稻瘟病斑, 具有黄色、褐色或紫色边缘; 9级: 白色、灰白色或浅蓝色病斑, 病斑融合无边界。5级、7级和9级为典型感病类型。
1.2.3 cDNA序列同源搜索及全长cDNA克隆 从http://rice.plantbiology.msu.edu/网站下载抗稻瘟病基因
Pid2 和
Pid3 CDS全长序列, 与疣粒野生稻转录组测序结果(另文报道)建立本地文库进行同源性比对, e值<1e-5。根据疣粒野生稻中比对出的同源cDNA片段, 设计基因特异引物, 利用3'-Full RACE Core Set with Prime Script TMR Tase试剂盒(TaKaRa, Cat No.6106)和5'-Full RACE Kit with TAP试剂盒(TaKaRa, Cat No.6107), 按照操作说明进行3'和5'-RACE PCR扩增目的cDNA片段全长, 扩增产物克隆到T-Vector pMD19 (Simple) (Cat No.32710)载体后进行测序分析。
1.2.4 过表达载体构建和遗传转化 将通过测序验证的
OmPid2 和
OmPid3 的全长cDNA通过Seamless Assembly Cloning Kit (Clone Smar- ter Technologies, Cat No.C5891-50)连接到过表达载体pCAMBIA1300-OXHLL, 构建过表达载体pCA- MBIA1300-OXHLL-
OmPid2 (
OmPid2-ox )和pCAM- BIA1300-OXHLL-
OmPid3 (
OmPid3-ox )。参照
Nishimura等(2005) 农杆菌介导的遗传转化方法转入感病材料日本晴。
1.2.5 转基因T0 代植株分子鉴定 提取转基因水稻叶片基因组DNA, 首先利用潮霉素标记基因引物(Forward primer: 5'-GCTTCTGCGG GCGATTTGTGT-3'; Reverse primer: 5'-GGTCGC- GGAGGCTATGGATGC-3')进行PCR鉴定后, 进一步用特异的基因引物OmPid2-G (Forward primer: 5'-CCACCGATATCGTCGCAGATTG-3'; Reverse p- rimer: 5'-CGCAAGACCGGCAACAGGAT-3')和Om- Pid3-G (Forward primer: 5'-CCAGAATTGAAGTTG- CTACCTC GC-3'; Reverse primer: 5'-CGCAAGAC- CGGCAACAGGAT-3')进行PCR检测。PCR反应程序: 94°C预变性5分钟; 94°C变性30秒, 58°C退火30秒, 72°C延伸30秒, 30个循环; 72°C延伸3分钟。PCR产物用1%琼脂糖凝胶电泳检测。
2 结果与讨论2.1 疣粒野生稻稻瘟病抗性鉴定利用7个致病稻瘟病菌株: ZB13、ZB15和Zhong-10- 8-14 (四川籼稻区菌株), ZE1 (四川粳稻区菌株), LP- 174和LP29-3 (云南罗平籼粳混合区菌株), H53 (实验室从2013年哈尔滨稻瘟病重灾区五常病样分离菌株), 注射接种孕穗期疣粒野生稻, 接种后5-10天进行穗颈瘟发病调查。结果表明, 不同菌株接种后的发病速度和程度存在差异。LP29-3、LP174、ZB13、ZB15和Zhong-10-8-14发病快, 接种后5天就能观察到明显的穗颈发病, 白穗。H53接种后7天穗颈病斑明显, 籽粒半瘪。ZE1菌株接种后发病较慢, 接种后10天观察到明显的穗颈病斑, 籽粒半瘪。根据穗颈瘟分级标准, 除ZE1菌株接种出现直径<3 mm的圆形至椭圆形病斑属于中感表型外, 其余菌株(LP29-3、LP- 174、ZB13、H53、ZB15和Zhong-10-8-14)接种后均出现典型的纺锤形、直径远大于3 mm的稻瘟病病斑, 呈现9级以上的高感表型(
图1A )。
图1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_1.png <b>图1</b> 疣粒野生稻稻瘟病抗性鉴定<br/>(A) 注射接种; (B) 喷雾接种; (C) 分别注射接种ZB15和Zhong-10-8-14<br/><b>Figure 1</b> Identification of blast resistance in <i>Oryza meyeriana<br/></i>(A) Injection inoculation; (B) Spray inoculation; (C) Inoculated by blast strains ZB15 and Zhong-10-8-14, respectively Figure 1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_1.png <b>图1</b> 疣粒野生稻稻瘟病抗性鉴定<br/>(A) 注射接种; (B) 喷雾接种; (C) 分别注射接种ZB15和Zhong-10-8-14<br/><b>Figure 1</b> Identification of blast resistance in <i>Oryza meyeriana<br/></i>(A) Injection inoculation; (B) Spray inoculation; (C) Inoculated by blast strains ZB15 and Zhong-10-8-14, respectively 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
疣粒野生稻稻瘟病抗性鉴定
(A) 注射接种; (B) 喷雾接种; (C) 分别注射接种ZB15和Zhong-10-8-14
Figure 1
Identification of blast resistance in
Oryza meyeriana (A) Injection inoculation; (B) Spray inoculation; (C) Inoculated by blast strains ZB15 and Zhong-10-8-14, respectively
上述7个菌株中, ZB15和Zhong-10-8-4菌株对应的抗病基因
Pid2 和
Pid3 在水稻中已完成克隆, 我们利用这2个菌株进一步对疣粒野生稻2-3叶的新蘖进行喷雾接种, 并确保叶片正反面上黏附足够的孢子液。每种菌株各接种3盆(相对于3个水稻株系)疣粒野生稻, 每盆3个新蘖(相对于3株水稻幼苗), 同时也进行了疣粒野生稻孕穗期分蘖的注射接种(
图1C )。结果表明, 2种菌株注射接种后的发病表型与之前一致(
图1A ), 喷雾接种的疣粒野生稻均未发病(
图1B )。推测疣粒野生稻的叶片结构不利于稻瘟病菌的侵染。
2.2 疣粒野生稻中抗病同源基因克隆和序列分析
Pid2 (
Chen et al., 2006 )和
Pid3 (
Shang et al., 2009 )是从籼稻地谷中克隆的2个抗瘟主效基因, 在水稻中的抗病功能均得到了很好的鉴定。
Pid2 对稻瘟病菌株ZB15表现抗性, 而
Pid3 对稻瘟病菌株Zhong-10-8-14表现抗性。为了探明疣粒野生稻对稻瘟病菌抗/感性的分子基础, 我们利用抗病基因
Pid2 和
Pid3 的CDS序列(http://rice.plantbiology.msu.edu/), 通过同源搜索疣粒野生稻转录组测序预测蛋白编码框(CDS)建立的本地文库(转录组测序结果将另文报道), e阈值<1e- 5, 发现1条25 322 bp的Unigene序列与
Pid2 的同源性为92.2%, 1条624 bp的Unigene序列与
Pid3 的同源性为73.0%。将其分别命名为
OmPid2 和
OmPid3 。进一步根据Unigenes的序列设计引物, 采用3'/5' RA- CE-PCR方法, 我们从疣粒野生稻中克隆了这2个基因的全长cDNA。
从疣粒野生稻中克隆到的
OmPid2 cDNA片段全长为3 090 bp, 包括295 bp的5'UTR和260 bp的3'UTR, 编码由844个氨基酸残基组成的蛋白质, 与抗病蛋白Pid2之间存在多个位点的氨基酸差异, 氨基酸序列一致性为91.1% (
图2 )。Chen等(2006)研究表明, 感病蛋白pid2第441位氨基酸为M (甲硫氨酸), 而抗病蛋白Pid2第441位氨基酸为I (异亮氨酸), 第441位氨基酸I是Pid2授予水稻地谷稻瘟病抗性所必需的。而OmPid2第441位氨基酸既不是感病蛋白pid2的M, 也不是抗病蛋白中的I, 而是缬氨酸(V) (
图2 箭头所示)。所以无法根据氨基酸的序列分析推测
OmPid2 是否具有抗稻瘟病的功能, 需要进一步的功能研究。
图2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_2.png <b>图2</b> OmPid2与水稻同源蛋白的氨基酸序列比对<br/>OmPid2: 疣粒野生稻; R-Pid2: 地谷; S-pid2: 日本晴。信号肽结构域(21-52氨基酸、B凝集素结构域(68-184氨基酸)、PAN结构域(355-436氨基酸)和丝氨酸-苏氨酸激酶(STK)结构域(519-789氨基酸)在图中已用下划线标注。箭头示区分稻瘟病抗病基因<i>Pid2</i>抗感等位的关键氨基酸。<br/><b>Figure 2</b> Amino acid sequence alignment of OmPid2 and orthologs in rice<br/>OmPid2: <i>Oryza meyeriana</i>; R-Pid2: Digu; S-pid2: Nipponbare. Signal peptide domain (21-52 amino acids), B-lectin domain (68-184 amino acids), PAN domain (355-436 amino acids) and Serine/threonine kinase (STK) domain (519-789 amino acids) are underlined. Arrow indicates the amino acid that distinguishes resistant and susceptible alleles of rice blast resistance gene <i>Pid2</i>. Figure 2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_2.png <b>图2</b> OmPid2与水稻同源蛋白的氨基酸序列比对<br/>OmPid2: 疣粒野生稻; R-Pid2: 地谷; S-pid2: 日本晴。信号肽结构域(21-52氨基酸、B凝集素结构域(68-184氨基酸)、PAN结构域(355-436氨基酸)和丝氨酸-苏氨酸激酶(STK)结构域(519-789氨基酸)在图中已用下划线标注。箭头示区分稻瘟病抗病基因<i>Pid2</i>抗感等位的关键氨基酸。<br/><b>Figure 2</b> Amino acid sequence alignment of OmPid2 and orthologs in rice<br/>OmPid2: <i>Oryza meyeriana</i>; R-Pid2: Digu; S-pid2: Nipponbare. Signal peptide domain (21-52 amino acids), B-lectin domain (68-184 amino acids), PAN domain (355-436 amino acids) and Serine/threonine kinase (STK) domain (519-789 amino acids) are underlined. Arrow indicates the amino acid that distinguishes resistant and susceptible alleles of rice blast resistance gene <i>Pid2</i>. 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
OmPid2与水稻同源蛋白的氨基酸序列比对
OmPid2: 疣粒野生稻; R-Pid2: 地谷; S-pid2: 日本晴。信号肽结构域(21-52氨基酸、B凝集素结构域(68-184氨基酸)、PAN结构域(355-436氨基酸)和丝氨酸-苏氨酸激酶(STK)结构域(519-789氨基酸)在图中已用下划线标注。箭头示区分稻瘟病抗病基因
Pid2 抗感等位的关键氨基酸。
Figure 2
Amino acid sequence alignment of OmPid2 and orthologs in rice
OmPid2:
Oryza meyeriana ; R-Pid2: Digu; S-pid2: Nipponbare. Signal peptide domain (21-52 amino acids), B-lectin domain (68-184 amino acids), PAN domain (355-436 amino acids) and Serine/threonine kinase (STK) domain (519-789 amino acids) are underlined. Arrow indicates the amino acid that distinguishes resistant and susceptible alleles of rice blast resistance gene
Pid2 .
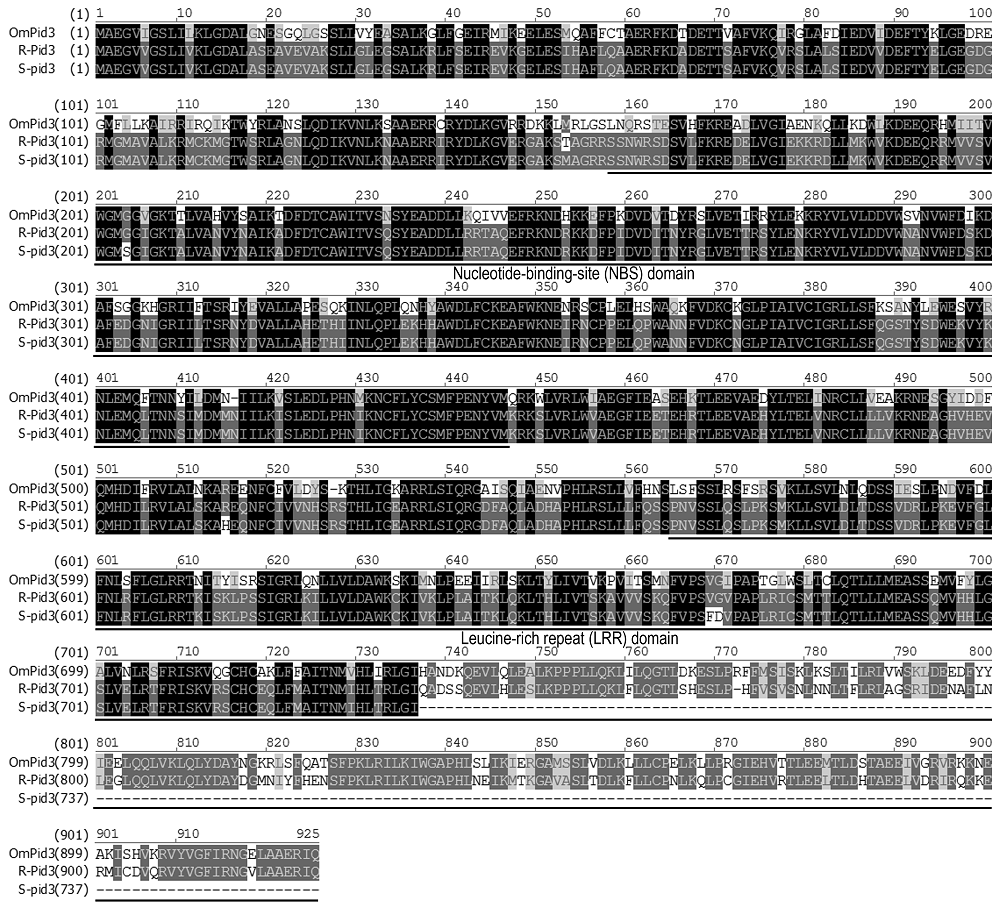
Shang等(2009) 研究表明, 在感病材料日本晴中, 由于
Pid3 基因第2 271位碱基C-T的替换变异形成TAG终止密码子, 造成编码蛋白在第737位氨基酸处提前终止, 从而丧失抗病功能。我们从疣粒野生稻中克隆到
OmPid3 cDNA全长为3 889 bp, 包括993 bp的5'UTR和124 bp的3'UTR, 在编码区没有出现提前终止密码子, 编码由923个氨基酸组成的蛋白质。将OmPid3 (疣粒野生稻)与水稻中同源蛋白Pid3 (地谷)和pid3 (日本晴)氨基酸序列比对分析(
图3 ), 发现OmPid3与水稻同源蛋白氨基酸序列差异较大, 同源性为67.6%。故
OmPid3 的功能也很难通过氨基酸序列比对分析进行预测。
图3 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_3.png <b>图3</b> OmPid3与水稻同源蛋白的氨基酸序列比对<br/>OmPid3: 疣粒野生稻; R-Pid3: 地谷; S-pid3: 日本晴。核酸结合位点(NBS)结构域(158-445氨基酸)和亮氨酸重复(LRR)结构域(563-923氨基酸)在图中已用下划线标注。<br/><b>Figure 3</b> Amino acid sequence alignment of OmPid3 and orthologs in rice<br/>OmPid3: <i>Oryza meyeriana</i>; R-Pid3: Digu; S-pid3: Nipponbare. Nucleotide-binding site (NBS) domain (158-445 amino acids) and Leucine-rich repeat (LRR) domain (563-923 amino acids) are underlined. Figure 3 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_3.png <b>图3</b> OmPid3与水稻同源蛋白的氨基酸序列比对<br/>OmPid3: 疣粒野生稻; R-Pid3: 地谷; S-pid3: 日本晴。核酸结合位点(NBS)结构域(158-445氨基酸)和亮氨酸重复(LRR)结构域(563-923氨基酸)在图中已用下划线标注。<br/><b>Figure 3</b> Amino acid sequence alignment of OmPid3 and orthologs in rice<br/>OmPid3: <i>Oryza meyeriana</i>; R-Pid3: Digu; S-pid3: Nipponbare. Nucleotide-binding site (NBS) domain (158-445 amino acids) and Leucine-rich repeat (LRR) domain (563-923 amino acids) are underlined. 在新窗口打开 下载 下载原图ZIP 生成PPT 图3
OmPid3与水稻同源蛋白的氨基酸序列比对
OmPid3: 疣粒野生稻; R-Pid3: 地谷; S-pid3: 日本晴。核酸结合位点(NBS)结构域(158-445氨基酸)和亮氨酸重复(LRR)结构域(563-923氨基酸)在图中已用下划线标注。
Figure 3
Amino acid sequence alignment of OmPid3 and orthologs in rice
OmPid3:
Oryza meyeriana ; R-Pid3: Digu; S-pid3: Nipponbare. Nucleotide-binding site (NBS) domain (158-445 amino acids) and Leucine-rich repeat (LRR) domain (563-923 amino acids) are underlined.
2.3
OmPid2 和
OmPid3 基因功能为进一步明确疣粒野生稻感病表型是否与
OmPid2 和
OmPid3 基因的功能一致, 我们克隆了这2个基因并构建过表达载体
OmPid2-ox 和
OmPid3-ox 。利用农杆菌介导的转化方法分别将这2个基因转入感病材料日本晴。采用潮霉素标记基因引物和
OmPid2 、
OmPid3 的特异基因引物对转基因T
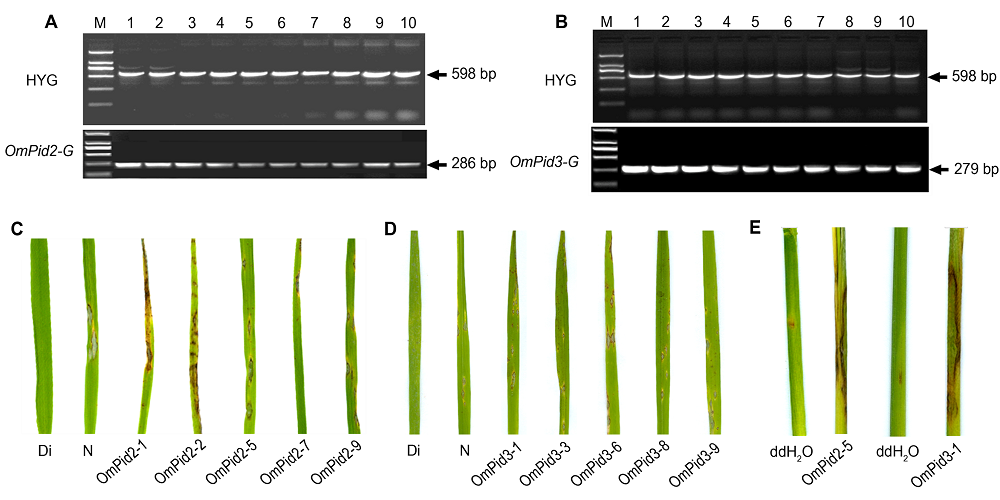
0 植株进行PCR分子检测, 各获得10株阳性转基因植株(
图4A , B)。分别利用稻瘟病菌株ZB15和Zhong-10-8-14喷雾接种
OmPid2-ox 、
OmPid3-ox 转基因幼苗和野生型日本晴幼苗, 转基因植株都表现出与对照日本晴类似或略加重的感病表型(
图4C , D), 与疣粒野生稻注射接种ZB15和Zhong-10-8-14菌株感病表型一致。由于稻瘟病菌株ZB15和Zhong-10-8-14喷雾接种疣粒野生稻不感病, 与注射接种结果不一致。为此, 我们利用ZB15和Zhong-10-8-14对移栽后生长良好的转基因株系
OmPid2-5 和
OmPid3-1 , 于分蘖期分别进行叶鞘注射接种, 转基因水稻感病明显, 与喷雾接种结果一致(
图4E )。这一结果支持了我们关于疣粒野生稻喷雾接种不发病很可能与其叶片结构有关的推测。
图4 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_4.png <b>图4</b> 转基因水稻植株的鉴定<br/><i>OmPid2</i> (A)和<i>OmPid3</i> (B)过表达转基因株系潮霉素和基因引物分子检测; <i>OmPid2</i> (C)和<i>OmPid3</i> (D)过表达转基因株系喷雾接种抗性鉴定; <i>OmPid2</i>和<i>OmPid3</i> (E)过表达转基因株系注射接种ZB15和Zhong-10-8-14抗性鉴定。HYG: 潮霉素; Di: 水稻地谷; N: 野生型对照日本晴<br/><b>Figure 4</b> Characterisation of transgenic rice lines<br/>Molecular detection of transgenic rice lines with Hygromycin primer, <i>OmPid2 </i>gene-specific primer (A) and <i>OmPid3 </i>gene-specific primer (B); Blast resistance idetification of <i>OmPid2 </i>(C) and <i>OmPid3</i> (D) overexpressing transgenic rice lines inoculated by blast strains; Overexpressing transgenic rice lines <i>OmPid2 </i>and<i> OmPid3 </i>(E) were inoculated by blast strains ZB15 and Zhong-10-8-14, respectively. HYG: Hygromycin; Di: The resistant rice variety Digu; N: Wild-type susceptible rice variety Nipponbare Figure 4 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-477/img_4.png <b>图4</b> 转基因水稻植株的鉴定<br/><i>OmPid2</i> (A)和<i>OmPid3</i> (B)过表达转基因株系潮霉素和基因引物分子检测; <i>OmPid2</i> (C)和<i>OmPid3</i> (D)过表达转基因株系喷雾接种抗性鉴定; <i>OmPid2</i>和<i>OmPid3</i> (E)过表达转基因株系注射接种ZB15和Zhong-10-8-14抗性鉴定。HYG: 潮霉素; Di: 水稻地谷; N: 野生型对照日本晴<br/><b>Figure 4</b> Characterisation of transgenic rice lines<br/>Molecular detection of transgenic rice lines with Hygromycin primer, <i>OmPid2 </i>gene-specific primer (A) and <i>OmPid3 </i>gene-specific primer (B); Blast resistance idetification of <i>OmPid2 </i>(C) and <i>OmPid3</i> (D) overexpressing transgenic rice lines inoculated by blast strains; Overexpressing transgenic rice lines <i>OmPid2 </i>and<i> OmPid3 </i>(E) were inoculated by blast strains ZB15 and Zhong-10-8-14, respectively. HYG: Hygromycin; Di: The resistant rice variety Digu; N: Wild-type susceptible rice variety Nipponbare 在新窗口打开 下载 下载原图ZIP 生成PPT 图4
转基因水稻植株的鉴定
OmPid2 (A)和
OmPid3 (B)过表达转基因株系潮霉素和基因引物分子检测;
OmPid2 (C)和
OmPid3 (D)过表达转基因株系喷雾接种抗性鉴定;
OmPid2 和
OmPid3 (E)过表达转基因株系注射接种ZB15和Zhong-10-8-14抗性鉴定。HYG: 潮霉素; Di: 水稻地谷; N: 野生型对照日本晴
Figure 4
Characterisation of transgenic rice lines
Molecular detection of transgenic rice lines with Hygromycin primer,
OmPid2 gene-specific primer (A) and
OmPid3 gene-specific primer (B); Blast resistance idetification of
OmPid2 (C) and
OmPid3 (D) overexpressing transgenic rice lines inoculated by blast strains; Overexpressing transgenic rice lines
OmPid2 and
OmPid3 (E) were inoculated by blast strains ZB15 and Zhong-10-8-14, respectively. HYG: Hygromycin; Di: The resistant rice variety Digu; N: Wild-type susceptible rice variety Nipponbare
结合疣粒野生稻注射接种高感稻瘟病菌株ZB15和Zhong-10-8-14的表型(
图1A ), 我们认为疣粒野生稻中
Pid2 和
Pid3 的同源基因
OmPid2 与
OmPid3 是感病基因; 疣粒野生稻中
OmPid2 和
OmPid3 基因可能维持在较原始的状态, 在疣粒野生稻中可能更多地在其它生长发育过程或抗逆中发挥作用, 对稻瘟病的抗/感性作用不大。进一步对转基因植株性状的详细调查和分析将揭示相关信息。
2.4 讨论疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 )。相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏。
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病。本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异。ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致。可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 )。不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性)。例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 )。疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关。本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病。我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性。疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关。疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义。
疣粒野生稻中
OmPid2 和
OmPid3 基因编码的氨基酸序列与栽培稻同源蛋白间存在较大差异, 它们的过表达转基因水稻幼苗分别接种稻瘟病菌ZB15和Zhong-10-8-14, 其感病表型与野生型感病水稻日本晴相似。我们推测: (1) 由于疣粒野生稻生存环境的特殊性(生长在遮阴的灌木和竹林下干旱土壤的山丘), 疣粒野生稻中栽培稻的抗病同源基因, 在进化过程中没有受到稻瘟病菌压力的选择, 维持在较原始的状态, 对现代的稻瘟病菌株无抗性。这与疣粒野生稻在稻属中属于GG基因组, 与稻属其它种亲缘关系较远, 位于稻属系统进化的最底层, 保存有许多较原始的特性一致(
Zou et al., 2008 ;
Jena, 2010 )。(2) 疣粒野生稻中
OmPid2 和
OmPid3 基因可能维持在较原始的状态, 在疣粒野生稻中可能更多地在其它生长发育过程或抗逆中发挥作用, 对稻瘟病的抗/感性作用不大。进一步对转基因植株其它性状的详细调查和分析将揭示相关信息。
综上所述, 我们初步认为, 疣粒野生稻具有一定的抵抗稻瘟病菌侵染的物理抗性, 很少有或甚至没有“基因对基因假说”的主效抗稻瘟病基因。研究结果对疣粒野生稻资源的基因挖掘和育种利用研究具有重要参考价值。
The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 胡朝芹 , 刘剑宇 , 王韵茜 , 杨睿 , 汪秉琨 , 何月秋 , 曾千春 , 罗琼 (2017 ). 粳稻子预44抗LP11稻瘟病菌基因Pizy6 (t )的定位. 植物学报 52, 61 -69 .[2] 蒋春苗 , 黄兴奇 , 李定琴 , 余腾琼 , 程在全 (2012 ). 云南野生稻叶茎根组织结构特性的比较研究. 西北植物学报 32, 99 -105 .[3] 李冬波 , 张端品 , 林兴华 (2009 ). SSR标记在疣粒野生稻和普通栽培稻中的多态性研究. 华中农业大学学报 28, 1 -4 .[4] 梁斌 , 肖放华 , 黄费元 , 彭绍裘 , 陈勇 , 戴陆国 , 刘二明 (1999 ). 云南野生稻对稻瘟病的抗性评价. 中国水稻科学 13, 183 -185 .[5] 刘继梅 , 程在全 , 杨明挚 , 吴成军 , 王玲仙 , 孙一丁 , 黄兴奇 (2003 ). 云南3种野生稻中抗病基因同源序列的克隆及序列分析. 中国农业科学 36, 273 -280 .[6] 刘进 , 刘建锋 , 曾义 (2001 ). 寄主植物的形态结构抗病性. 四川林业科技 22(3 ), 54 -56 .[7] 彭绍裘 , 魏子生 , 毛昌祥 , 黄河清 , 肖放华 , 罗宽 (1982 ). 云南省疣粒野生稻、药用野生稻和普通野生稻多抗性鉴定. 植物病理学报 12, 60 -62, 74 .[8] 盛腊红 , 何光存 , 舒理慧 , 廖兰杰 (1999 ). 提高疣粒野生稻愈伤组织分化能力的研究. 植物学通报 16, 614 -617 .[9] 文婷 , 梁毅 , 江南 , 李智强 , 金灵 , 杨婷婷 , 戴良英 , 王国梁 , 刘雄伦 (2012 ). 利用pi9 基因序列标记辅助选择改良籼稻稻瘟病抗性. 湖南农业大学学报(自然科学版) 38, 262 -266 .[10] 颜惠霞 , 徐秉良 , 梁巧兰 , 薛应钰 , 陈荣贤 , 梁志福 (2009 ). 南瓜品种对白粉病的抗病性与叶绿素含量和气孔密度的相关性. 植物保护 35, 79 -81 .[11] 杨健源 , 康金平 , 黄显良 , 姜先芽 , 赖添奎 , 陈深 , 李传瑛 , 曾先列 , 朱小源 (2007 ). 稻瘟病田间自然诱发病圃的建立及其在抗性鉴定中的应用. 广东农业科学 (9 ), 59 -61 .[12] 云勇 , 韩义胜 (2014 ). 我国野生稻资源的抗病性鉴定与利用研究进展. 植物遗传资源学报 15, 472 -476, 482 .[13] 张乃群 , 杨莉萍 , 杜敏华 (1999 ). 我国三种野生稻及两个栽培稻品种叶片表面亚显微结构的观察研究. 南都学坛(自然科学版) 19(3 ), 70 -73 .[14] 周镕 , 王波 , 杨睿 , 李书 , 樊琳琳 , 曾千春 , 罗琼 (2015 ). 粳稻子预44中稻瘟病数量抗性位点分析. 植物学报 50, 691 -698 .[15] Chen XW Shang JJ Chen DX Lei CL Zou Y Zhai WX Liu GZ Xu JC Ling ZZ Cao G Ma BT Wang YP Zhao XF Li SG Zhu LH 2006 ). A B-lectin receptor kinase gene conferring rice blast resistance. Plant J 46, 794 -804 .[16] Das A Soubam D Singh PK Thakur S Singh NK Sharma TR 2012 ). A novel blast resistance gene, Pi54rh cloned from wild species of rice, Oryza rhizomatis confers broad spectrum resistance to Magnaporthe oryzae . Funct Integr Genomics 215 -228 .[17] Fu CY Wu T Liu WG Wang F Li JH Zhu XY Huang HJ Liu ZR Liao YL Zhu MS Chen JW Huang YJ 2012 ). Genetic improvement of resistance to blast and bacterial blight of the elite maintainer line Rongfeng B in hybrid rice (Oryza sativa L.) by using marker-assisted selection. Afr J Biotechnol 13104 -13124 .[18] Jena KK 2010 ). The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa . Breed Sci 518 -523 .[19] Jeung JU Kim BR Cho YC Han SS Moon HP Lee YT Jena KK 2007 ). A novel gene, Pi40(t) , linked to the DNA markers derived from NBS-LRR motifs confers broad spectrum of blast resistance in rice. Theor Appl Genet 1163 -1177 .[20] Koide Y Kawasaki A Telebanco-Yanoria MJ Hairmansis A Nguyet NTM Bigirimana J Fujita D Kobayashi N Fukuta Y 2010 ). Development of pyramided lines with two resistance genes, Pish and Pib , for blast disease(Magnaporthe oryzae B. Couch) in rice (Oryza sativa L.). Plant Breed 670 -675 .[21] Liu WD Liu JL Triplett L Leach JE Wang GL 2013 ). Novel insights into rice innate immunity against bacterial and fungal pathogens.Annu Rev Phytopathol 52, 213 -241 .[22] Lv QM Xu X Shang JJ Jiang GH Pang ZQ Zhou ZZ Wang J Liu Y Li T Li XB Xu JC Cheng ZK Zhao XF Li SG Zhu LH 2013 ). Functional analysis of Pid3 -A4 , an ortholog of rice blast resistance gene Pid3 revealed by allele mining in common wild rice. Phytopathology 594 -599 .[23] Nishimura A Ashikari M Lin SY Takashi T Angeles ER Yamamoto T Matsuoka M 2005 ). Isolation of a rice regeneration quantitative trait loci gene and its application to transformation systems.Proc Natl Acad Sci USA 102, 11940 -11944 .[24] Shang JJ Yong T Chen XW Zou Y Lei CL Wang J Li XB Zhao XF Zhang MJ Lu ZK Xu JC Cheng ZK Wan JM Zhu LH 2009 ). Identification of a new rice blast resistance gene, Pid3 , by genomewide comparison of paired nucleotide-binding site-leucine-rich repeat genes and their pseudogene alleles between the two sequenced rice genomes. Genetics 1303 -1311 .[25] Skamnioti P Gurr SJ 2009 ). Against the grain: safeguarding rice from rice blast disease.Trends Biotechnol 27, 141 -150 .[26] Standard Evaluation System (SES) for Rice (2013 ). Authority Files for International Rice Research Institute, 5th edition . .URL [27] Tanweer FA Rafii MY Sijam K Rahim HA Ahmed F Latif MA 2015 ). Current advance methods for the identification of blast resistance genes in rice.CR Biol 338, 321 -334 .[28] Tharreau D Lebrun MH Talbot NJ Notteghem JL 2000 ). Advances in Rice Blast Research . Netherlands: Springer Press. pp. 54 -62 .[29] Vaughan DA 1994 ). The Wild Relatives of Rice. A Genetic Resources Handbook . Manila: International Rice Research Institute. pp. 457 -462 .[30] Xiao N Wu YY Pan CH Yu L Chen Y Liu GQ Li YH Zhang XX Wang ZP Dai ZY Liang CZ Li AH 2016 ). Improving of rice blast resistances in Japonica by pyramiding major R genes. Front Plant Sci 1918 .[31] Zou XH Zhang FM Zhang JG Zhang LL Tang L Wang J Sang T Ge S 2008 ). Analysis of 142 genes resolves the rapid diversification of the rice genus.Genome Biol 9, R49 .
粳稻子预44抗LP11稻瘟病菌基因
Pizy6 (
t )的定位
1
2017
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
云南野生稻叶茎根组织结构特性的比较研究
1
2012
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
SSR标记在疣粒野生稻和普通栽培稻中的多态性研究
1
2009
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
云南野生稻对稻瘟病的抗性评价
2
1999
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
云南3种野生稻中抗病基因同源序列的克隆及序列分析
1
2003
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
寄主植物的形态结构抗病性
1
2001
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
云南省疣粒野生稻、药用野生稻和普通野生稻多抗性鉴定
2
1982
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
提高疣粒野生稻愈伤组织分化能力的研究
1
1999
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
利用
pi9 基因序列标记辅助选择改良籼稻稻瘟病抗性
1
2012
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
南瓜品种对白粉病的抗病性与叶绿素含量和气孔密度的相关性
1
2009
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
稻瘟病田间自然诱发病圃的建立及其在抗性鉴定中的应用
1
2007
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
我国野生稻资源的抗病性鉴定与利用研究进展
1
2014
... 疣粒野生稻(
O. meyeriana )是我国现有3种野生稻资源之一, 主要分布在云南热带和海南西南部地区, 在长期适应各种恶劣环境的进化过程中, 其保存了许多栽培稻不具备的优异基因资源, 如对白叶枯病菌免疫、高抗褐稻虱、抗细菌性条斑病以及耐旱耐荫等(
盛腊红等, 1999 ;
云勇和韩义胜, 2014 ).
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 表明云南疣粒野生稻具有中抗稻瘟病特性.
刘继梅等(2003) 尝试利用同源克隆的方法从疣粒野生稻中克隆抗病同源基因, 但未能成功.
李冬波等(2009) 利用SSR分子标记进行疣粒野生稻和栽培稻IR24基因组多态性分析, 发现两材料间多态性高达96.2%, 认为疣粒野生稻和栽培稻间存在较大的基因组序列差异, 故根据栽培稻基因组序列设计PCR引物, 通过常规PCR方法很难实现疣粒野生稻中抗病同源基因的克隆.迄今为止, 疣粒野生稻可利用的基因组信息非常有限, 其抗病基因克隆研究几乎还是空白, 对疣粒野生稻抗病遗传基础更是缺乏最基本的了解, 这对疣粒野生稻中优异基因资源的研究和利用是极大的限制. ...
我国三种野生稻及两个栽培稻品种叶片表面亚显微结构的观察研究
1
1999
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
粳稻子预44中稻瘟病数量抗性位点分析
1
2015
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
2
2006
... 为了进一步了解疣粒野生稻的抗病特性, 有效挖掘和利用疣粒野生稻中的抗病基因资源, 本研究利用分离自不同水稻生态区的稻瘟病菌株(ZB13、ZB15、Zhong-10-8-14、ZE1、LP174、LP29-3和H53), 通过注射接种法对疣粒野生稻进行系统的稻瘟病抗性调查.采用3'/5'RACE方法, 从疣粒野生稻中克隆了
Pid2 (地谷中抗稻瘟病菌株ZB15的主效抗瘟基因) (
Chen et al., 2006 )和
Pid3 (地谷中抗稻瘟病菌株Zhong-10-8-14的主效抗瘟基因) (
Shang et al., 2009 )的同源基因, 并构建过表达转基因株系对基因功能进行研究.研究结果将为疣粒野生稻中优异基因资源的研究和利用提供新信息和新思路. ...
...
Pid2 (
Chen et al., 2006 )和
Pid3 (
Shang et al., 2009 )是从籼稻地谷中克隆的2个抗瘟主效基因, 在水稻中的抗病功能均得到了很好的鉴定.
Pid2 对稻瘟病菌株ZB15表现抗性, 而
Pid3 对稻瘟病菌株Zhong-10-8-14表现抗性.为了探明疣粒野生稻对稻瘟病菌抗/感性的分子基础, 我们利用抗病基因
Pid2 和
Pid3 的CDS序列(http://rice.plantbiology.msu.edu/), 通过同源搜索疣粒野生稻转录组测序预测蛋白编码框(CDS)建立的本地文库(转录组测序结果将另文报道), e阈值<1e- 5, 发现1条25 322 bp的Unigene序列与
Pid2 的同源性为92.2%, 1条624 bp的Unigene序列与
Pid3 的同源性为73.0%.将其分别命名为
OmPid2 和
OmPid3 .进一步根据Unigenes的序列设计引物, 采用3'/5' RA- CE-PCR方法, 我们从疣粒野生稻中克隆了这2个基因的全长cDNA. ...
1
2012
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2012
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2010
... 疣粒野生稻中
OmPid2 和
OmPid3 基因编码的氨基酸序列与栽培稻同源蛋白间存在较大差异, 它们的过表达转基因水稻幼苗分别接种稻瘟病菌ZB15和Zhong-10-8-14, 其感病表型与野生型感病水稻日本晴相似.我们推测: (1) 由于疣粒野生稻生存环境的特殊性(生长在遮阴的灌木和竹林下干旱土壤的山丘), 疣粒野生稻中栽培稻的抗病同源基因, 在进化过程中没有受到稻瘟病菌压力的选择, 维持在较原始的状态, 对现代的稻瘟病菌株无抗性.这与疣粒野生稻在稻属中属于GG基因组, 与稻属其它种亲缘关系较远, 位于稻属系统进化的最底层, 保存有许多较原始的特性一致(
Zou et al., 2008 ;
Jena, 2010 ).(2) 疣粒野生稻中
OmPid2 和
OmPid3 基因可能维持在较原始的状态, 在疣粒野生稻中可能更多地在其它生长发育过程或抗逆中发挥作用, 对稻瘟病的抗/感性作用不大.进一步对转基因植株其它性状的详细调查和分析将揭示相关信息. ...
1
2007
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2010
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2013
... 稻瘟病是由稻瘟病菌(
Magnaporthe oryzae )引发并能够侵染水稻(
Oryza sativa )整个生育期的真菌病害, 是水稻生产上最为严重的病害之一.其分布广泛, 危害严重, 造成年均10%-30%的产量损失, 严重影响稻米产量和品质, 威胁世界粮食安全(
Ska- mnioti and Gurr, 2009 ;
Liu et al., 2013 ).目前, 抗病品种的种植和化学农药的使用是防治该病害的主要措施.化学农药的大量使用带来了一系列包括环境污染和食品安全等问题, 而抗病品种的利用是防治稻瘟病最安全有效的措施.但由于稻瘟病菌群体组成复杂、变异快, 导致1个抗病品种大面积推广种植3-5年后抗性便逐渐丧失.为此, 鉴定新的抗病资源、克隆新的抗病基因, 是培育水稻抗病新品种的关键. ...
1
2013
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2005
... 将通过测序验证的
OmPid2 和
OmPid3 的全长cDNA通过Seamless Assembly Cloning Kit (Clone Smar- ter Technologies, Cat No.C5891-50)连接到过表达载体pCAMBIA1300-OXHLL, 构建过表达载体pCA- MBIA1300-OXHLL-
OmPid2 (
OmPid2-ox )和pCAM- BIA1300-OXHLL-
OmPid3 (
OmPid3-ox ).参照
Nishimura等(2005) 农杆菌介导的遗传转化方法转入感病材料日本晴. ...
3
2009
... 为了进一步了解疣粒野生稻的抗病特性, 有效挖掘和利用疣粒野生稻中的抗病基因资源, 本研究利用分离自不同水稻生态区的稻瘟病菌株(ZB13、ZB15、Zhong-10-8-14、ZE1、LP174、LP29-3和H53), 通过注射接种法对疣粒野生稻进行系统的稻瘟病抗性调查.采用3'/5'RACE方法, 从疣粒野生稻中克隆了
Pid2 (地谷中抗稻瘟病菌株ZB15的主效抗瘟基因) (
Chen et al., 2006 )和
Pid3 (地谷中抗稻瘟病菌株Zhong-10-8-14的主效抗瘟基因) (
Shang et al., 2009 )的同源基因, 并构建过表达转基因株系对基因功能进行研究.研究结果将为疣粒野生稻中优异基因资源的研究和利用提供新信息和新思路. ...
...
Pid2 (
Chen et al., 2006 )和
Pid3 (
Shang et al., 2009 )是从籼稻地谷中克隆的2个抗瘟主效基因, 在水稻中的抗病功能均得到了很好的鉴定.
Pid2 对稻瘟病菌株ZB15表现抗性, 而
Pid3 对稻瘟病菌株Zhong-10-8-14表现抗性.为了探明疣粒野生稻对稻瘟病菌抗/感性的分子基础, 我们利用抗病基因
Pid2 和
Pid3 的CDS序列(http://rice.plantbiology.msu.edu/), 通过同源搜索疣粒野生稻转录组测序预测蛋白编码框(CDS)建立的本地文库(转录组测序结果将另文报道), e阈值<1e- 5, 发现1条25 322 bp的Unigene序列与
Pid2 的同源性为92.2%, 1条624 bp的Unigene序列与
Pid3 的同源性为73.0%.将其分别命名为
OmPid2 和
OmPid3 .进一步根据Unigenes的序列设计引物, 采用3'/5' RA- CE-PCR方法, 我们从疣粒野生稻中克隆了这2个基因的全长cDNA. ...
...
Shang等(2009) 研究表明, 在感病材料日本晴中, 由于
Pid3 基因第2 271位碱基C-T的替换变异形成TAG终止密码子, 造成编码蛋白在第737位氨基酸处提前终止, 从而丧失抗病功能.我们从疣粒野生稻中克隆到
OmPid3 cDNA全长为3 889 bp, 包括993 bp的5'UTR和124 bp的3'UTR, 在编码区没有出现提前终止密码子, 编码由923个氨基酸组成的蛋白质.将OmPid3 (疣粒野生稻)与水稻中同源蛋白Pid3 (地谷)和pid3 (日本晴)氨基酸序列比对分析(
图3 ), 发现OmPid3与水稻同源蛋白氨基酸序列差异较大, 同源性为67.6%.故
OmPid3 的功能也很难通过氨基酸序列比对分析进行预测. ...
1
2009
... 稻瘟病是由稻瘟病菌(
Magnaporthe oryzae )引发并能够侵染水稻(
Oryza sativa )整个生育期的真菌病害, 是水稻生产上最为严重的病害之一.其分布广泛, 危害严重, 造成年均10%-30%的产量损失, 严重影响稻米产量和品质, 威胁世界粮食安全(
Ska- mnioti and Gurr, 2009 ;
Liu et al., 2013 ).目前, 抗病品种的种植和化学农药的使用是防治该病害的主要措施.化学农药的大量使用带来了一系列包括环境污染和食品安全等问题, 而抗病品种的利用是防治稻瘟病最安全有效的措施.但由于稻瘟病菌群体组成复杂、变异快, 导致1个抗病品种大面积推广种植3-5年后抗性便逐渐丧失.为此, 鉴定新的抗病资源、克隆新的抗病基因, 是培育水稻抗病新品种的关键. ...
3
2013
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
... ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
... 叶瘟发病分级参照国际水稻研究所的标准评价系统(
Standard Evaluation System for Rice, SES, 2013 ), 即病斑类型分6级.0级: 无任何病斑; 1级: 仅有针尖大小褐点或褐点稍大; 3级: 有小的圆形坏死斑, 直径约为1-2 mm, 有明显褐色边缘或黄色晕; 5级: 出现宽1-2 mm、直径>3 mm的圆形至椭圆形病斑, 边缘褐色; 7级: 出现典型的纺锤形稻瘟病斑, 具有黄色、褐色或紫色边缘; 9级: 白色、灰白色或浅蓝色病斑, 病斑融合无边界.5级、7级和9级为典型感病类型. ...
0
2015
2
2000
... 按照
Tharreau (2000) 的6级分类法进行病斑类型调查.0级(HR): 无任何病斑或者不侵染; 1级(R): 有零星直径<0.5 mm的褐色病斑; 2级(MR): 有少数直径为0.5-1 mm的褐色病斑; 3级(MS): 出现直径<3 mm的圆形至椭圆形病斑, 中央灰白色, 边缘褐色; 4级(S): 典型的纺锤形稻瘟病斑, 直径3 mm或更长, 病斑稍有融合或无融合; 5级(HS): 典型的纺锤形稻瘟病斑, 直径3 mm或更长病斑融合成片. ...
... 疣粒野生稻在漫长的进化过程中, 由于所处地理环境和复杂生态环境的影响, 形成了极其丰富的遗传多样性, 具有耐荫、抗旱和对白叶枯病免疫等特性, 使其成为水稻品种改良的重要种质资源(
Vaughan, 1994 ).相比白叶枯病抗性的研究, 对疣粒野生稻稻瘟病抗性的研究比较缺乏.
彭绍裘等(1982) 和
梁斌等(1999) 利用自然病圃对云南野生稻进行稻瘟病抗性评价, 认为云南疣粒野生稻中抗稻瘟病.本研究在温室条件下, 利用四川籼稻区优势菌株ZB13、ZB15和Zhong-10-8-14, 粳稻区优势菌株ZE1, 云南罗平籼粳混合区菌株LP174和LP29-3, 哈尔滨五常稻瘟病重灾区菌株H53, 采用注射接种法对疣粒野生稻进行系统抗性鉴定, 结果表明, 疣粒野生稻对本实验接种的所有稻瘟病菌株均表现为感病或高感(
图1A ), 与之前自然病圃的研究结果存在差异.ZB15和Zhong-10- 8-14对疣粒野生稻的喷雾接种结果表明, 疣粒野生稻表现高抗, 与自然接种结果一致.可能原因为: 一方面, 由于自然诱发病圃对疣粒野生稻稻瘟病抗性的鉴定受到年度间气候条件、诱发圃菌群数量和种类的影响(
杨健源等, 2007 ); 另一方面, 由于植物同病原菌的互作是一个较复杂的系统, 病原菌浸染寄主引起发病的程度受寄主表皮细胞的结构(包括蜡质层厚度, 表皮毛或刺的有无和多少, 气孔、水孔的密度和大小, 表皮细胞的木质化程度, 后壁组织的结构)等影响(
刘进等, 2001 ).不同的物理结构会使植物在自然条件下表现出不同的抗病性(物理抗性).例如, 关于南瓜(
Cucurbita moschata )叶片气孔的密度与白粉病抗性关系的研究表明
, 气孔的密度与植株的抗病性呈负相关, 叶片上气孔密度低, 病菌侵入的机会就少, 植株表现抗病(
颜惠霞等, 2009 ).疣粒野生稻对自然诱发接种所表现出抗稻瘟病特性, 很可能与疣粒野生稻生长在落叶灌木、竹林下的干旱土壤, 长期适应旱生环境而具有较厚的叶片、无主脉结构和气孔较少等(
张乃群等, 1999 ;
蒋春苗等, 2012 )叶片结构所形成的限制稻瘟病菌浸染的天然屏障有关.本研究利用注射接种法对疣粒野生稻稻瘟病抗性进行研究, 发现疣粒野生稻高感稻瘟病.我们推测, 由于注射接种有助于病原菌克服侵入的物理屏障而克制寄主的物理抗性.疣粒野生稻在自然诱发条件下表现抗稻瘟病很可能与其叶片结构形成的物理抗性有关.疣粒野生稻中控制这一类性状的基因资源的挖掘和育种利用, 对改良栽培稻的植株特性并控制病虫害具有重要的理论和现实意义. ...
1
1994
... 自1999年克隆了第1个广谱抗瘟基因
Pib 以来, 截至目前, 在水稻基因组中已鉴定出100个以上稻瘟病抗性位点(
Pi loci)和300多个QTLs (
Tanweer et al., 2015 ;
周镕等, 2015 ;
胡朝芹等, 2017 ).已完成克隆和功能鉴定的抗稻瘟病基因有25个(
Tanweer et al., 2015 ;
Xiao et al., 2016 ), 其中一些基因已应用于育种中, 对稻瘟病的控制起到了积极作用.例如, 利用功能性分子标记
Pi9-a 将广谱抗瘟基因
Pi9 导入丰源B、II32B、天龙香103、明恢63、轮回422、R747和25H003等水稻材料中, 均筛选到抗性明显改良的BC
3 F
1 材料(
文婷等, 2012 ); 通过分子标记辅助选择将
Pish 和
Pib 聚合于水稻品种C039, 将
Pi1 、
Pi2 和
Xa21 聚合于保持系Rong-Feng B, 均显著提高了受体的抗瘟性(
Koide et al., 2010 ;
Fu et al., 2012 ).为进一步拓宽栽培稻的抗性遗传背景, 近年来育种家越来越多地重视从栽培稻的近缘野生种中挖掘抗病基因并进行利用.例如, 从澳洲野生稻中克隆的
Pi- 40 (
t ), 对来自韩国和菲律宾的强致病菌株均有很好的抗性(
Jeung et al., 2007 ); 从二倍体根茎野生稻中克隆的
Pi54 , 对来自印度不同区域的7个稻瘟病菌小种表现高抗(
Das et al., 2012 ); 从普通野生稻A4中克隆的
A4-Pid3 , 具有比
Pid3 更宽的抗谱(
Lv et al., 2013 ). ...
1
2016
... 疣粒野生稻中
OmPid2 和
OmPid3 基因编码的氨基酸序列与栽培稻同源蛋白间存在较大差异, 它们的过表达转基因水稻幼苗分别接种稻瘟病菌ZB15和Zhong-10-8-14, 其感病表型与野生型感病水稻日本晴相似.我们推测: (1) 由于疣粒野生稻生存环境的特殊性(生长在遮阴的灌木和竹林下干旱土壤的山丘), 疣粒野生稻中栽培稻的抗病同源基因, 在进化过程中没有受到稻瘟病菌压力的选择, 维持在较原始的状态, 对现代的稻瘟病菌株无抗性.这与疣粒野生稻在稻属中属于GG基因组, 与稻属其它种亲缘关系较远, 位于稻属系统进化的最底层, 保存有许多较原始的特性一致(
Zou et al., 2008 ;
Jena, 2010 ).(2) 疣粒野生稻中
OmPid2 和
OmPid3 基因可能维持在较原始的状态, 在疣粒野生稻中可能更多地在其它生长发育过程或抗逆中发挥作用, 对稻瘟病的抗/感性作用不大.进一步对转基因植株其它性状的详细调查和分析将揭示相关信息. ...
0
2008
 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}