
山东农业大学园艺科学与工程学院, 作物生物学国家重点实验室, 泰安 271018
引用本文
贡献者
基金资助
接受日期:2016-11-14接受日期:2016-12-11网络出版日期:2017-11-1
-->Copyright
2017《植物学报》编辑部
Contributors
History
Received:Accepted:Online:
摘要:
Abstract:
Key words:
我国葡萄的主产区分布在西北、华北、东北以及沿海等地, 然而受气候变化及不合理耕作等因素的影响, 上述地区的土壤盐渍化程度不断扩大(黄毅和张玉龙, 2004)。由此造成的盐胁迫已成为制约我国葡萄生产的主要逆境因素, 冬季寒冷则是主要的气候胁迫因素。因此, 在西北干旱和半干旱葡萄产区, 常用抗寒性强的贝达作为砧木。但贝达在这些地区由于盐碱胁迫往往表现出严重的缺铁黄化现象, 其替代砧木1103P (Vitis berlandieri×V. rupestris)虽在常用砧木中的耐盐性较强(Galet, 1991), 但在盐渍化严重的地区仍不能满足生产需要。因此, 选育既抗寒又耐盐的葡萄砧木成为我国葡萄产业亟待解决的问题。
因为葡萄根瘤蚜的侵染, 国外很早就开始进行抗性砧木育种, 其中选育的砧木品种SO4和101-1对根瘤蚜免疫且抗寒性较强, 但耐盐性一般(Troncoso et al., 1999)。我国是东亚葡萄种群的发源地之一, 拥有丰富的野生山葡萄(Vitis amurensis)种质资源(贺普超, 1999), 其中选育的山葡萄品种左山一(V. amu- rensis cv. ‘Zuoshan 1’)栽培性状好、抗寒性强(何伟等, 2015), 且其耐盐性在山葡萄种质资源中也较好(秦红艳, 2010)。本实验选择以左山一为母本, 以SO4 (V. berlandieri×V. riparia)和101-1 (V. riparia×V. rupe- stris)分别为父本进行杂交初选出的抗根瘤蚜/抗寒群体中的F1代株系为试材, 研究其抗盐能力, 以期选育出耐盐性较强的砧木新品种。
Walker等(1981)发现葡萄光合速率的显著降低与高盐胁迫密切相关; Troncoso等(1999)以生长量为指标对葡萄砧木进行了耐盐性分类。鉴于耐盐性较为复杂, 单一指标不能全面反映特定品种或基因型的遗传本质; 且选用的耐盐性指标不同, 对植物的耐盐性评价结果也不一致。因此, 需要对耐盐性评价指标进行筛选, 找出与耐盐性评价相关性较大的指标。在作物抗逆性筛选和鉴定方面, 有****提出了总级别法和隶属函数法等(钮福祥等, 1996)。但目前耐盐性评价指标较多, 且各指标间的相关性不同, 因此, 单纯使用总级别法或隶属函数法对作物进行抗逆性评价不够全面(王军等, 2007; 杨淑萍等, 2013)。本研究分析了盐胁迫下葡萄光合和荧光参数的变化, 并通过主成分分析从多个存在相关性的指标中挑选出几个新的综合指标。利用聚类分析对各主成分耐盐性综合评价值进行分类, 所获结果能较好地反映各葡萄株系对盐胁迫的耐受性, 为葡萄种质资源的筛选与鉴定、良种选育和品种改良等提供一定的理论依据。
1 材料与方法1.1 实验材料与设计实验于2015-2016年连续两年在山东农业大学南校区葡萄园避雨棚内进行。实验材料为左山一×SO4杂种F1代的6个葡萄株系(A15、A17、A34、A35、A38和A48); 左山一×101-1杂种F1代的2个株系B24和B26, 以已知砧木中耐盐性较强的1103P品种为标准参照。试材种植于园土:沙:基质=2:1:1 (v/v/v)的花盆中(直径17 cm、高25 cm), 每盆1株, 待长至8-10片完全展开的叶时, 选取长势一致的幼苗进行实验。
设置NaCl处理浓度为60和100 mmol·L-1, 各株系以浇自来水为对照。2015年6-7月, 每隔3天于下午5:00-6:00进行浇灌, 每盆浇1 000 mL, 至溶液从盆底外流。实验发现, 100 mmol·L-1 NaCl处理幼苗6天, B26表现出盐害症状, 处理13天1103P开始出现症状, 20天后其余株系均表现出不同程度的明显的伤害; 而60 mmol·L-1 NaCl处理幼苗20天, 各株系以及1103P并未表现出盐害症状。因此, 选定100 mmol·L-1 NaCl作为正式实验盐处理浓度。
2016年6-7月, 用100 mmol·L-1 NaCl对葡萄幼苗进行处理, 每个株系处理10次重复(10株), 处理20天后进行各项指标的测定。
1.2 测定项目与方法1.2.1 光合作用气体交换参数的测定
使用CIRAS-2便携式光合系统(PP-Systems, 美国), 于上午9:00-11:00测定各葡萄株系梢尖下数第4、5片展开叶的净光合速率(net photosynthetic rate, Pn)、气孔导度(stomatal conductance, Gs)、胞间CO2浓度(intercellular CO2 concentration, Ci)以及蒸腾速率(transpiration rate, Tr)。测定过程中, 光强设定为处理条件下光照强度(1 000 μmol·m-2·s-1), CO2浓度为(360±20) μL·L-1, 叶室温度设定为25°C。
1.2.2 叶绿素荧光参数的测定
叶片暗处理30分钟后, 利用Handy PEA连续激发式荧光仪(Hansatech, 英国)测定快速叶绿素荧光诱导
曲线(OJIP) (李鹏民等, 2005)。OJIP曲线由3 000 μmol·m-2·s-1脉冲光诱导, 记录10 μs-1 s荧光信号。通过JIP-test分析, 得到OJIP曲线及参数最大光化学效率(maximal photochemical efficiency, Fv/Fm)、 PSII受体侧相对电子传递速率(relative electron tr- ansfer rate of PSII receptor side, Ψ0) (Ψ0=ET0/TR0= (1-Vj)、单位面积活性反应中心数目(the number of active reaction centers per unit area, RC/CSm) (RC/ CSm=Fm×φPo×(Vj/Mo)和K点荧光的相对变化值(the relative change of K point fluorescence, WK) (WK= (Ft-F0)/(Fm-F0)。
采用FMS-2型便携脉冲调制式荧光仪(Hansa- tech, 英国)对其它荧光参数进行分析。测定程序如下: 对自然光适应20分钟后的葡萄叶片先打60秒作用光, 打<0.05 μmol·m-2·s-1的测量光测得叶片最小荧光(Initial fluorescence in the light, F0'), 再打12 000 μmol·m-2·s-1饱和脉冲光, 测定光适应下的最大荧光值(maximal fluorescence yield of the light-adapted state, Fm'), 打开作用光待Ft稳定后取此时的Ft为稳态荧光(Steady-state fluorescence, Fs), 然后关闭叶夹, 对叶片测定部位进行30分钟的暗适应, 打12 000 μmol·m-2·s-1饱和脉冲光, 使原初电子受体QA全部处于还原状态, 测定暗适应下的最大荧光(Maximal fluorescence intensity, Fm)。各荧光参数(PSII实际光化学效率(actual photochemical efficiency, ФPSII)、光化学淬灭(photochemical quenching, qP)和非光化学淬灭(non-photochemical quenching, NPQ))的计算公式如下:
ФPSII=(Fm'-Fs)/Fm'; qP=(Fm'-Fs)/(Fm'-F0');
NPQ=Fm/Fm'-1。
1.2.3 其它指标的测定
采用赵世杰等(2002)的方法, 选择每株梢尖下数第4、5片叶测定其叶绿素含量。测定的生长指标包括新梢相对生长量、节间数、地上部鲜重、地下部鲜重、地上部干重、地下部干重、地上部含水量、地下部含水量、全株鲜重、全株干重、根冠比和全株含水量。根冠比和植株含水量的计算公式如下:
根冠比=地下部干重/地上部干重 (1)
植株含水量=(鲜重-干重)/鲜重 (2)
1.3 数据处理与统计分析1.3.1 耐盐系数(salt tolerance coefficient, STC)
STC (%)=(盐胁迫测定值/对照测定值)×100% (3)
1.3.2 各综合指标的隶属函数值
U(Xj)=(Xj-Xmin)/(Xmax-Xmin)×100%, j=1, 2, ……n
(4)
式中, U(Xj)表示第j个综合指标的隶属函数值, Xj表示第j个综合指标, Xmin和Xmax分别表示第j个综合指标的最小值与最大值。
1.3.3 各综合指标的权重(Wj)
Wj=Σnj=1|Pj|, j=1, 2, ……n (5)
式中, Wj表示第j个综合指标在所有综合指标中的重要程度, Pj为各基因型第j个综合指标的贡献率。
1.3.4 各株系的综合耐盐能力大小
D=Σnj=1|μ(Xj)Wj|, j=1, 2, ……n (6)
式中, D为各株系在盐胁迫下的耐盐性综合评价值。
采用Excel 2013软件进行数据统计与整理, 计算各指标的平均值和耐盐系数。使用DPS软件进行方差分析, 利用LSD法进行多重比较差异显著性检验, 使用SPSS 17.0软件进行主成分分析、相关性分析和聚类分析。
2 结果与讨论2.1 盐胁迫株系叶片的表型及盐害分级用100 mmol·L-1 NaCl对试材进行胁迫处理20天后, 观察各葡萄株系的表型及盐害情况。根据盐害轻重分为5个等级(王业遴等, 1990)。0级: 未表现出盐害症状; 1级: 轻度盐害, 有少量的叶片表现叶尖、叶缘或叶脉变黄; 2级: 中度盐害, 有约1/2的叶片出现叶尖和叶缘焦枯症状; 3级: 重度盐害, 大部分叶片出现叶边缘和叶尖焦枯及叶片脱落现象; 4级: 极重度盐害, 枝枯、叶落, 直至死亡。本研究结果表明, A34和A35植株无盐害症状, 盐害级数为0级; A15和A17植株有少部分叶片边缘焦枯, 盐害级数为1级, A38、A48和B24植株大部分叶片萎蔫干枯, 盐害级数为2级; 1103P盐害级数为3级; B26则表现为4级盐害(图1)。
图1
Figure 1

下载原图ZIP
生成PPT
图1
NaCl胁迫对各葡萄株系叶片表型的影响
Figure 1
The phenotype of grape hybrid strains under NaCl stress
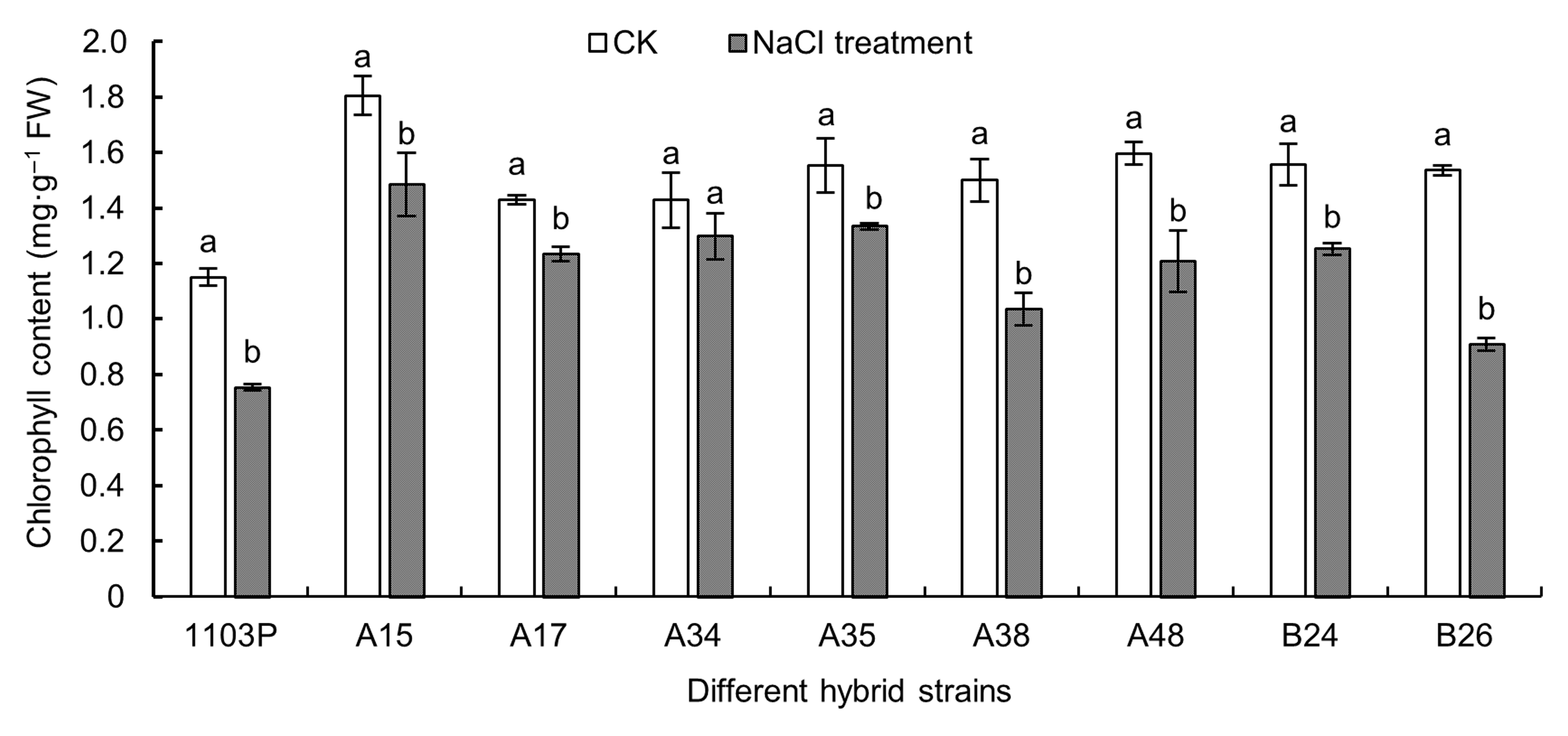
2.2 盐胁迫对各株系叶片叶绿素含量的影响用100 mmol·L-1 NaCl处理各葡萄株系, 其叶片叶绿素含量均表现出不同程度的降低(图2)。A15、A17、A34和A35植株降低幅度较小, 分别比对照降低了17.71%、13.72%、9.06%和14.14%; 而A38、A48和B24植株降低幅度较大, 分别比对照降低了30.95%、24.36%和19.55%; 1103P和B26植株降低幅度最大, 分别比对照降低了34.56%和40.85%。
图2
Figure 2
下载原图ZIP
生成PPT
图2
NaCl胁迫对各葡萄株系叶片叶绿素含量的影响
不同小写字母表示同一株系不同处理间差异显著(P<0.05)。
Figure 2
Effects of NaCl stress on chlorophyll content in different grape hybrid strains
Different lowercase letters indicate significant differences under different treatments of the same strain (P<0.05).
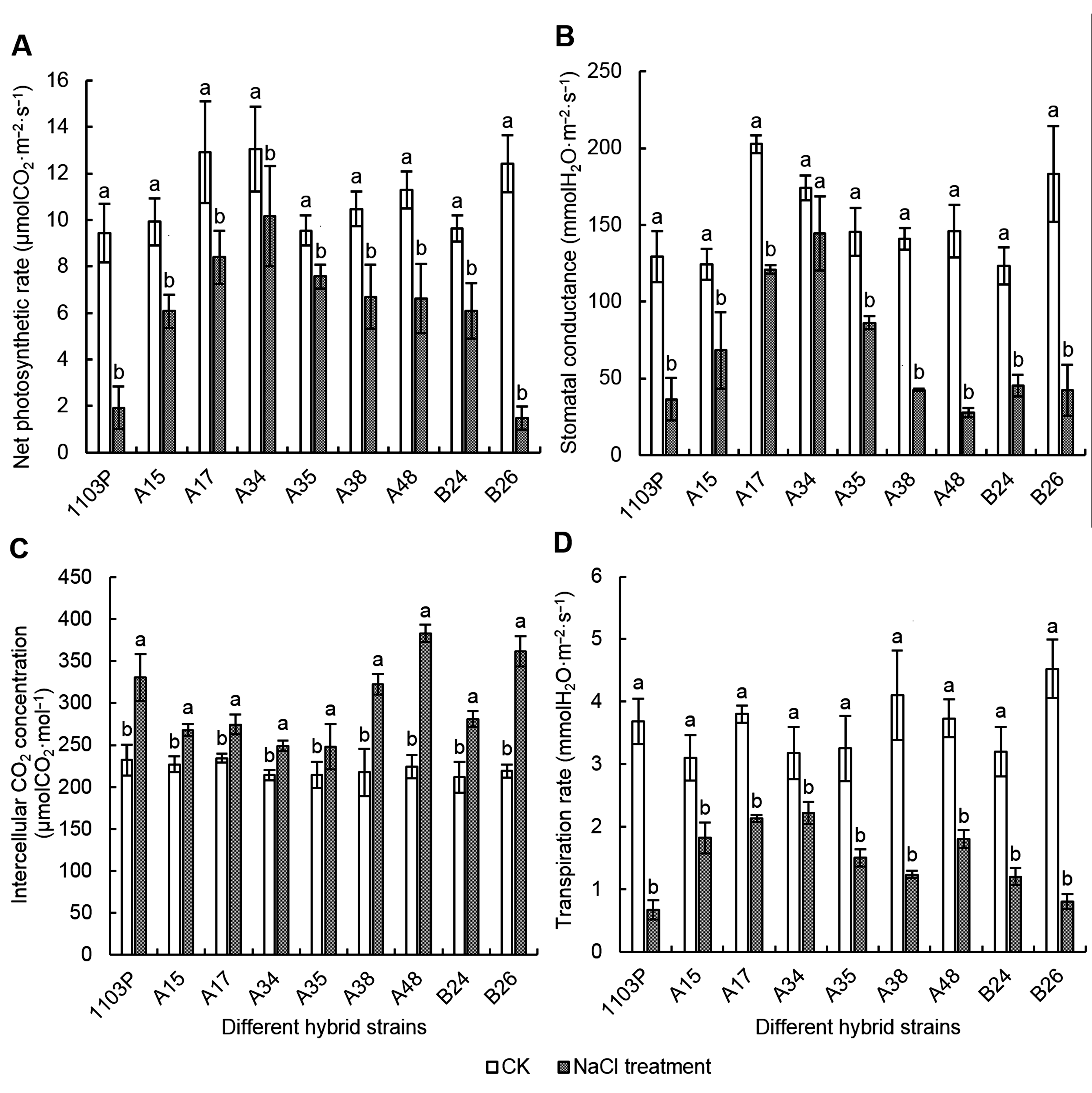
2.3 盐胁迫对各株系叶片光合作用气体交换参数的影响用100 mmol·L-1 NaCl处理各葡萄株系, 其叶片净光合速率、气孔导度和蒸腾速率下降。本研究结果表明, A34以及A35植株的净光合速率降低幅度较小; 而1103P和B26植株的净光合速率降幅最大, 分别比对照降低了79.59%和88.12% (图3A)。A15、A17、A34和A35植株的气孔导度降低较小, 分别比对照降低了45.07%、40.28%、17.07%和40.68% (图3B)。另外, 此4个植株的蒸腾速率分别比对照降低了41.29%、43.86%、30.08%和53.85%; 而1103P和B26植株的蒸腾速率分别比对照降低了81.9%和82.32% (图3D)。
图3
Figure 3
下载原图ZIP
生成PPT
图3
NaCl胁迫对各葡萄株系叶片Pn、Gs、Ci和Tr的影响
Pn: 净光合速率; Gs: 气孔导度; Ci: 胞间CO2浓度; Tr: 蒸腾速率。不同小写字母表示同一株系不同处理间差异显著(P<0.05)。
Figure 3
Effects of NaCl stress on Pn, Gs, Ci and Tr of different grape hybrid strains
Pn: Net photosynthetic rate; Gs: Stomatal conductance; Ci: Intercellular CO2 concentration; Tr: Transpiration rate. Different lowercase letters indicate significant differences under different treatments of the same strain (P<0.05).
用100 mmol·L-1 NaCl处理各葡萄株系, 其叶片胞间CO2浓度增加(图3C)。A15、A17、A34以及A35植株的Ci与对照相比升高的幅度较小, 分别升高了18.08%、17.16%、16.2%和15.71%; 而1103P、A38、A48、B24和B26植株的Ci升高幅度较大, 分别比对照升高了42.21%、48.31%、71.07%、32.7%和65.07%。
2.4 盐胁迫对各株系叶片叶绿素荧光参数的影响最大光化学效率(Fv/Fm)是反映光抑制程度的良好指标(莫伟平等, 2013); 实际光化学效率(ΦPSII)表示在光照条件下PSII反应中心部分关闭后的实际光化学效率, 即植物叶片在光下用于电子传递的能量占所捕获光能的比例。用100 mmol·L-1 NaCl处理各葡萄株系后, 其叶片叶绿素荧光参数发生了显著变化(表1)。各葡萄株系叶片的Fv/Fm和ΦPSII均有不同程度的降低, 但A15、A17、A34和A35植株的Fv/Fm下降幅度较小, 分别比对照降低了17.78%、14.7%、6.08%和8.71%; A17、A34和A35植株的ΦPSII下降幅度较小, 分别比对照降低了29.23%、24.06%和28.25%;1103P以及B26植株的ΦPSII则下降幅度最大, 分别比对照降低了64.88%和68.66%。
表1
Table 1
表1
| Strains | NaCl concentration (mmol·L-1) | Fv/Fm | ΦPSII | Wk | RC/CSm | Ψ0 | qP | NPQ |
|---|---|---|---|---|---|---|---|---|
| 1103p | 0 | 0.811±0.007 a | 0.612±0.053 a | 0.387±0.009 ef | 776.5±34.8 abc | 0.545±0.065 a | 0.856±0.026 ab | 0.703±0.139 g |
| 100 | 0.285±0.150 f | 0.153±0.062 e | 0.536±0.072 b | 111.7±94.21 h | 0.21±0.079 e | 0.322±0.131 f | 4.608±0.81 bc | |
| A15 | 0 | 0.827±0.018 a | 0.61±0.027 a | 0.385±0.025 ef | 793±51.51 ab | 0.539±0.040 a | 0.857±0.012 ab | 1.256±0.04 fg |
| 100 | 0.68±0.024 cde | 0.35±0.071 cd | 0.515±0.045 bc | 621.1±73.95 cd | 0.383±0.062 cd | 0.662±0.051 e | 3.017±0.46 de | |
| A17 | 0 | 0.818±0.023 a | 0.601±0.057 a | 0.402±0.016 ef | 643.2±55.1 bcd | 0.507±0.08 abc | 0.863±0.028 a | 0.908±0.049 g |
| 100 | 0.70±0.016 bcde | 0.425±0.01 cd | 0.475±0.037 cd | 514.9±76.19 de | 0.347±0.059 d | 0.76±0.020 bcd | 1.62±0.146 fg | |
| A34 | 0 | 0.818±0.009 a | 0.609±0.033 a | 0.401±0.016 ef | 668.1±33.3 bcd | 0.52±0.044 ab | 0.875±0.009 a | 1.137±0.07 fg |
| 100 | 0.768±0.023 abc | 0.463±0.03 bc | 0.436±0.014 de | 568.7±19.40 de | 0.491±0.02 abc | 0.712±0.019 de | 2.057±0.403 ef | |
| A35 | 0 | 0.824±0.008 a | 0.543±0.04 ab | 0.397±0.022 ef | 631.3±74.80 cd | 0.517±0.018 ab | 0.82±0.014 abc | 2.235±0.251 ef |
| 100 | 0.75±0.006 abcd | 0.39±0.035 cd | 0.415±0.03 def | 528±25.68 de | 0.395±0.07 bcd | 0.76±0.027 bcd | 2.982±1.11 de | |
| A38 | 0 | 0.813±0.007 a | 0.607±0.023 a | 0.352±0.014 f | 773.3±37.2 abc | 0.563±0.078 a | 0.887±0.018 a | 0.856±0.103 g |
| 100 | 0.629±0.059 e | 0.324±0.065 d | 0.528±0.035 bc | 280.7±130.3 fg | 0.302±0.042 de | 0.682±0.048 de | 5.53±0.287 ab | |
| A48 | 0 | 0.816±0.012 a | 0.607±0.079 a | 0.39±0.022 ef | 748.5±49.6 abc | 0.53±0.042 a | 0.873±0.011 a | 1.076±0.085 g |
| 100 | 0.633±0.077 de | 0.368±0.03 cd | 0.58±0.051 ab | 418.6±51.03 ef | 0.275±0.054 de | 0.675±0.076 de | 3.396±0.391 d | |
| B24 | 0 | 0.806±0.003 ab | 0.576±0.098 a | 0.388±0.011 ef | 901.3±65.99 a | 0.534±0.034 a | 0.861±0.015 a | 1.335±0.26 fg |
| 100 | 0.646±0.030 de | 0.344±0.036 d | 0.548±0.028 ab | 653.5±44.5 bcd | 0.35±0.048 d | 0.715±0.051 de | 3.614±0.07 cd | |
| B26 | 0 | 0.818±0.012 a | 0.57±0.039 ab | 0.369±0.004 f | 875.6±38.21 a | 0.55±0.038 a | 0.851±0.018 ab | 1.27±0.114 fg |
| 100 | 0.256±0.108 f | 0.093±0.079 e | 0.616±0.062 a | 151.3±128.4 gh | 0.188±0.047 e | 0.307±0.307 f | 6.42±1.076 a |
表1
盐胁迫对葡萄叶片叶绿素荧光参数的影响
Table 1
Effects of NaCl stress on chlorophyll fluorescence parameters in leaves of grape
K点荧光的相对变化值(WK)表示PSII供体侧放氧复合体(oxygen evolving complex, OEC)的伤害程度(strasser, 1997), WK值升高表明供体侧受到了伤害; RC/CSm表示单位面积有活性反应中心的比例; Ψ0表示PSII捕获的激子将电子传递到电子传递链中QA以后的其它电子受体的概率, 反映了PSII受体侧电子传递伤害程度(金立桥等, 2015)。用NaCl胁迫处理各葡萄株系后, 其叶片的WK值表现出不同程度的升高, A17、A34和A35植株比对照分别升高了18.14%、8.81%和4.54%; 而1103P、A15、A38、A48、B24和B26植株升高幅度较大, 分别比对照升高了38.47%、33.56%、50.09%、48.58%、41.15%和66.87%。各株系叶片的RC/CSm与Ψ0则表现出不同程度的降低, A15、A17、A34和A35植株的RC/CSm及Ψ0降低的幅度较小, 此4个植株的RC/CSm分别比对照降低了21.68%、19.94%、14.88%和16.36%; 而1103P和B26植株的RC/CSm降低幅度最大。
qP与NPQ分别表示荧光淬灭中的光化学淬灭和非光化学淬灭。各葡萄株系的qP在NaCl胁迫下发生了不同程度的降低, 1103P和B26植株的qP降低幅度较大, 分别比对照降低了62.36%和63.91%; A15、A17、A34、A35、A38、A48和B24植株的qP降低幅度较小, 分别比对照降低了22.7%、11.92%、18.6%、7.75%、23.11%、22.68%和16.92%。
2.5 盐胁迫对各株系生长指标的影响2.5.1 各单项指标的耐盐系数及相关系数分析
为了更加合理地评价各葡萄株系对NaCl胁迫的敏感性, 根据实验所得数据, 利用公式(3)求出各葡萄株系各单项指标的耐盐系数。从表2可以看出, 在100 mmol·L-1 NaCl胁迫下, 少数株系的个别指标的耐盐系数≥100%。例如, B26植株的根冠比为129%, 说明100 mmol·L-1 NaCl处理对其地上部的抑制作用大于地下部(杨升等, 2015)。A35植株的地上部含水量与地下部含水量分别为101%和100%, A34植株的地上部含水量为100%, 与对照差异不显著, 而其它指标均表现下降。但不同葡萄株系各单项指标的变化幅度不尽相同, 因此, 利用单一性状耐盐系数评价各葡萄株系的耐盐性不够全面。从表3可以看出, 多数指标之间存在显著或极显著相关性, 导致它们所反映的信息发生重叠, 进而影响了耐盐性鉴定结果。为弥补各单项指标评价耐盐性的不足, 需要用其它多元统计的方法进行分析。
表2
Table 2
表2
| Strains | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1103P | 0.16 | 0.26 | 0.57 | 0.52 | 0.69 | 0.62 | 0.86 | 0.90 | 0.56 | 0.67 | 0.92 | 0.87 |
| A15 | 0.18 | 0.38 | 0.75 | 0.67 | 0.85 | 0.77 | 0.93 | 0.95 | 0.71 | 0.82 | 0.90 | 0.94 |
| A17 | 0.18 | 0.26 | 0.86 | 0.63 | 0.85 | 0.69 | 0.97 | 0.95 | 0.78 | 0.80 | 0.80 | 0.97 |
| A34 | 0.31 | 0.39 | 0.82 | 0.66 | 0.81 | 0.78 | 1.00 | 0.93 | 0.77 | 0.80 | 0.80 | 0.98 |
| A35 | 0.27 | 0.35 | 0.89 | 0.61 | 0.88 | 0.60 | 1.01 | 1.00 | 0.77 | 0.77 | 0.68 | 0.99 |
| A38 | 0.29 | 0.36 | 0.62 | 0.54 | 0.69 | 0.70 | 0.94 | 0.89 | 0.59 | 0.69 | 0.87 | 0.92 |
| A48 | 0.06 | 0.12 | 0.71 | 0.63 | 0.76 | 0.60 | 0.96 | 1.03 | 0.69 | 0.71 | 0.88 | 0.98 |
| B24 | 0.15 | 0.21 | 0.63 | 0.49 | 0.74 | 0.63 | 0.93 | 0.89 | 0.60 | 0.71 | 0.73 | 0.91 |
| B26 | 0.12 | 0.14 | 0.46 | 0.59 | 0.71 | 0.64 | 0.72 | 0.96 | 0.51 | 0.68 | 1.29 | 0.85 |
表2
NaCl胁迫下各葡萄株系各单项指标的耐盐系数
Table 2
Salt tolerant coefficient of every single index of each grape strains under NaCl stress
表3
Table 3
表3
| Growth parameters | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X1 | 1.000 | |||||||||||
| X2 | 0.881** | 1.000 | ||||||||||
| X3 | 0.437 | 0.523 | 1.000 | |||||||||
| X4 | 0.111 | 0.278 | 0.592 | 1.000 | ||||||||
| X5 | 0.266 | 0.431 | 0.893** | 0.711* | 1.000 | |||||||
| X6 | 0.516 | 0.678* | 0.271 | 0.522 | 0.284 | 1.000 | ||||||
| X7 | 0.458 | 0.525 | 0.852** | 0.309 | 0.576 | 0.275 | 1.000 | |||||
| X8 | -0.358 | -0.321 | 0.378 | 0.628 | 0.495 | -0.317 | 0.116 | 1.000 | ||||
| X9 | 0.373 | 0.473 | 0.984** | 0.702* | 0.898** | 0.347 | 0.821** | 0.436 | 1.000 | |||
| X10 | 0.376 | 0.588 | 0.833** | 0.770* | 0.909** | 0.644 | 0.605 | 0.245 | 0.874** | 1.000 | ||
| X11 | 0.301 | 0.297 | -0.501 | -0.137 | -0.564 | 0.615 | -0.273 | 0.678* | -0.456 | -0.203 | 1.000 | |
| X12 | 0.324 | 0.363 | 0.926** | 0.608 | 0.763* | 0.196 | 0.890** | 0.519 | 0.935** | 0.720* | -0.483 | 1.000 |
表3
NaCl胁迫下各单项指标的相关系数矩阵
Table 3
Correlation matrix of every single index under NaCl stress
2.5.2 主成分分析
主成分分析可以从原始的指标中重新转换成几个不相关的新指标, 更客观可靠地解释原始指标所包含的信息。利用SPSS17.0软件对各株系植株的12个相互关联的单项指标的耐盐系数进行主成分分析, 结果发现前3个综合评价指标的累计贡献率高达0.911 (分别为0.554、0.252和0.105)(表4), 因此其余指标的贡献率可忽略不计。我们把原来12个单项指标转换为3个相互独立的综合指标(comprehensive index, CI)。各综合指标的对应特征向量为:
表4
Table 4
表4
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 | P | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CI1 | 0.17 | 0.17 | 0.36 | 0.12 | 0.29 | 0 | 0.34 | 0.13 | 0.34 | 0.24 | -0.24 | 0.35 | 0.554 |
| CI2 | 0.46 | 0.47 | 0.04 | -0.02 | -0.03 | 0.44 | 0.14 | -0.42 | 0.03 | 0.15 | 0.42 | -0.03 | 0.252 |
| CI3 | -0.04 | 0.15 | 0.29 | 0.83 | 0.49 | 0.55 | 0.03 | 0.42 | 0.41 | 0.61 | 0.05 | 0.28 | 0.105 |
表4
NaCl胁迫下各综合指标的相关系数及贡献率
Table 4
Correlation of comprehensive indexes and their contribution rates under NaCl stress
第1主成分(CI1=0.17X1+0.17X2+0.36X3+0.12 X4+ 0.29X5+0.34X7+0.13X8+0.34X9+0.24X10-0.24X11+
0.35X12)、第2主成分(CI2=0.46X1-0.47X2+0.04X3- 0.02X4-0.03X5+0.44X6+0.14X7-0.42X8+0.03X9+0.15 X10+0.42X11-0.03X12)和第3主成分(CI3=-0.04X1+ 0.15X2+0.29X3+0.83X4+0.49X5+0.55X6+0.03X7+
0.424X8+0.41X9+0.61X10+0.05X11+0.28X12)。
由上述3个主成分的表达式可知: 在第1主成分表达式中, 第3、5、7、9和12项的系数较大, 它们分别代表地上部鲜重、地上部干重、地上部含水量、全株鲜重和全株含水量; 在第2主成分表达式中, 第1、2、6和11项的系数较大, 它们分别代表新梢生长量、节间数、地下部干重和根冠比; 在第3主成分表达式中, 第4项的系数远大于其它指标的系数, 即可认为是地上部干重的影响。以上结果表明, 生物量积累可作为评价各葡萄株系耐盐性的指标。
2.5.3 各葡萄株系的耐盐性综合评价
2.5.3.1 隶属函数分析
利用公式(4)计算各葡萄株系各综合指标的隶属函数值, 每个株系所有综合指标的隶属函数值不尽相同, 表明各株系的耐盐性不同。各综合指标的隶属函数值与耐盐性呈正相关, 在NaCl胁迫下, A34植株在CI2综合指标上表现的耐盐性最强; 而A48植株在CI2综合指标上表现的耐盐性最弱(表5)。
表5
Table 5
表5
| Strains | CI1 | CI2 | CI3 | U(X1) | U(X2) | U(X3) | Value D | Salt tolerance |
|---|---|---|---|---|---|---|---|---|
| 1103P | 1.39 | 0.67 | 2.63 | 0.20 | 0.46 | 0.00 | 0.252 | Weaker |
| A15 | 1.68 | 0.82 | 3.19 | 0.71 | 0.79 | 0.97 | 0.761 | Strong |
| A17 | 1.76 | 0.69 | 3.15 | 0.85 | 0.50 | 0.89 | 0.758 | Strong |
| A34 | 1.76 | 0.92 | 3.21 | 0.84 | 1.00 | 1.00 | 0.907 | Strong |
| A35 | 1.85 | 0.67 | 3.12 | 1.00 | 0.45 | 0.84 | 0.826 | Strong |
| A38 | 1.48 | 0.88 | 2.76 | 0.36 | 0.92 | 0.23 | 0.499 | Medium |
| A48 | 1.61 | 0.46 | 2.94 | 0.59 | 0.00 | 0.53 | 0.418 | Medium |
| B24 | 1.49 | 0.64 | 2.70 | 0.38 | 0.39 | 0.13 | 0.353 | Medium |
| B26 | 1.27 | 0.56 | 2.67 | 0.00 | 0.22 | 0.07 | 0.066 | Weaker |
| Index weight | 0.608 | 0.277 | 0.115 |
表5
NaCl胁迫下各株系的综合指标值、权重、隶属函数值、D值及综合评价
Table 5
Each strain’s comprehensive index, index weight, membership function value, value D and comprehensive evaluation under NaCl stress
2.5.3.2 权重的确定
根据各综合指标的贡献率大小(0.554、0.252和0.105) (表4), 用公式(5)求出各综合指标的权重(0.608、0.277和0.115) (表5)。
2.5.3.3 综合评价
利用公式(6)计算各葡萄株系的综合耐盐能力大小。根据综合评价值D对各株系的耐盐性进行排序, 并对各
图4
Figure 4
下载原图ZIP
生成PPT
图4
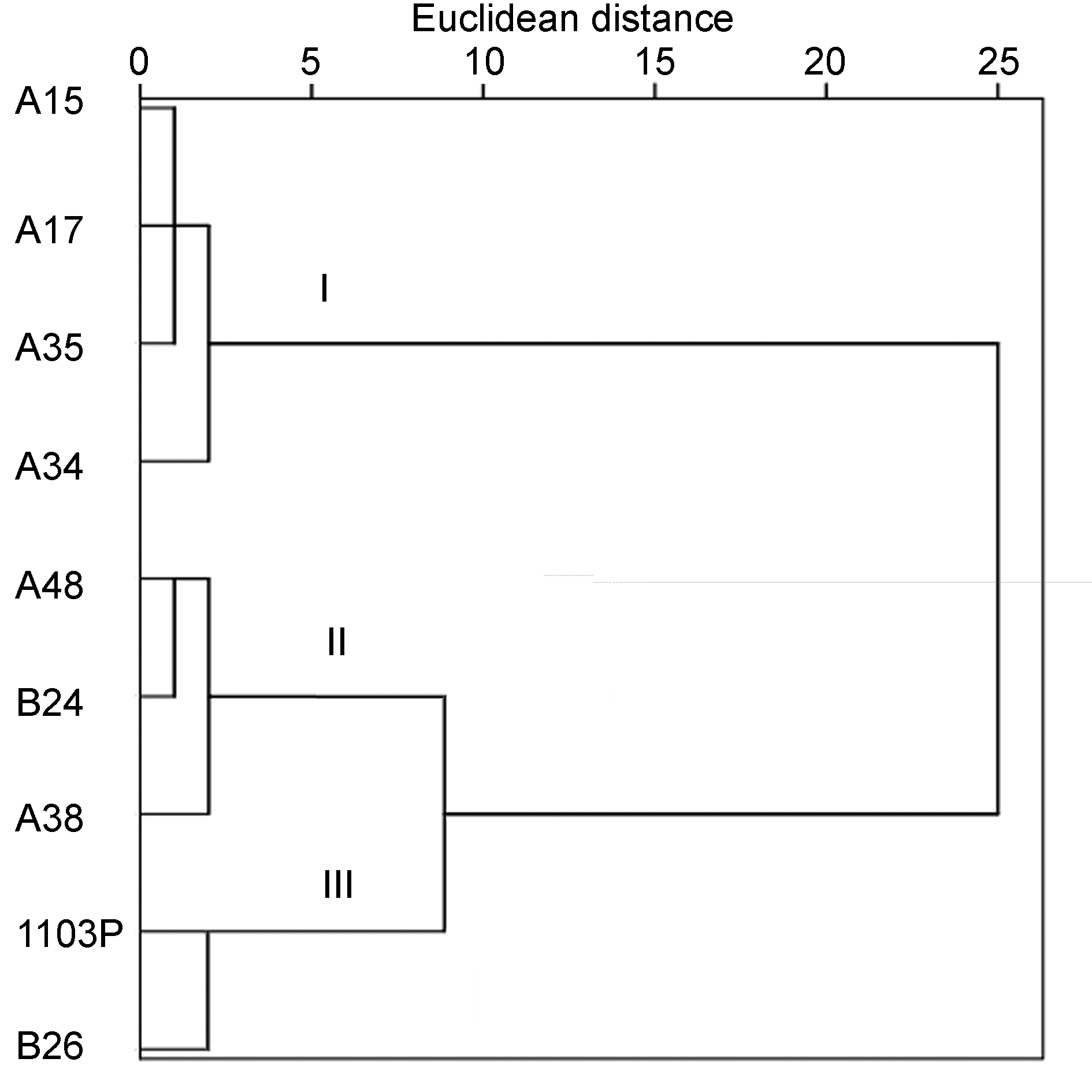
供试材料的耐盐性聚类图
Figure 4
Cluster diagram of salt tolerance of experimental materials
株系的耐盐性强弱进行分类。其中A34植株的D值最大, 说明其耐盐性最强; B26植株的D值最小, 其耐盐性最弱。用SPSS软件对D值采用组间连接法进行聚类分析, 得到1个聚类树形图(图4)。该树形图中的9个葡萄株系分为3类, 其中第1类包括A34、A35、A15和A17植株, 耐盐性较强; 第2类包括A38、A48和B24植株, 耐盐性中等; 第3类包括1103P和B26植株, 耐盐性较弱。
2.6 讨论与结论盐胁迫影响植物的生长和代谢等许多方面, 但对光合作用的影响尤为突出。王邦锡等(1992)研究表明, 叶片叶绿体结构受到破坏是影响植物光合作用正常进行的重要原因。盐胁迫可导致植物叶片叶绿素含量降低, 这主要是由于高盐使得植物叶片中叶绿素与叶绿素蛋白结合变得松弛, 叶绿素酶活性增强, 从而促进叶绿素降解所致(Hoshida et al., 2000)。本研究发现, 在100 mmol·L-1 NaCl胁迫下, 各葡萄株系叶片叶绿素含量以及净光合速率(Pn)均表现出不同程度的降低, 且耐盐性较好株系的下降幅度明显小于盐敏感株系, 这与前人对高粱(Sorghum bicolor) (孙璐等, 2012)和大豆(Glycine max) (薛忠财等, 2011)的研究结果一致。NaCl胁迫下胞间CO2浓度(Ci)均表现出不同程度的升高, 说明叶肉细胞对CO2的消耗能力降低(孙璐等, 2012)。另外, 气孔导度(Gs)也表现出不同程度的下降, 说明此时非气孔限制已成为促使光合速率降低的主要因素(Yang et al., 2006)。A15、A17、A34和A35植株的Gs与对照相比变化幅度较小, 表现出良好的耐盐性。
植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(Foyer and Noctor, 2000; 赵昕等, 2007)。 众多的研究表明, PSII与植物光合作用遭受伤害的最初部位关系密切, 盐胁迫会破坏植物叶片叶绿体光合作用机构, 降低PSII原初光能转化效率和PSII潜在活性(刘家尧等, 1998; Lu and Zhang, 1999)。本研究发现, 各株系叶片PSII最大光化学量子效率(Fv/Fm)和实际光化学效率(ΦPSII)在NaCl胁迫下均降低, 说明葡萄叶片PSII发生了光抑制(Xu et al., 1999)。进一步分析发现, 快速叶绿素荧光参数WK值(供体侧)升高, RC/CSm (反应中心)值和Ψ0 (受体侧)值降低, 表明盐胁迫下葡萄叶片PSII供体侧(Appenroth et al., 2001; 张菂等, 2013)、反应中心(罗海波等, 2010)以及受体侧(Yamasaki et al., 2002)等光合机构均受到伤害。光化学淬灭(qP)表示光合活性的高低, 反映了PSII原初电子受体QA的还原状态; 非光化学淬灭(NPQ)代表叶片的光耗散能力, 即启动热耗散, 以耗散过剩的光能(Gilmore, 1997)。本研究发现, 盐胁迫下qP下降, NPQ升高, 说明盐胁迫会改变葡萄叶片PSII的激发能分配方式, 以热耗散的形式消耗过多激发能来适应逆境环境。A15、A17、A34与A35植株的Fv/Fm、ΦPSII和qP下降幅度明显比其余各植株小, 而NPQ的升高幅度明显大于其它植株, 并且1103P和B26植株的ΦPSII下降幅度最大, 说明A15、A17、A34和A35植株在NaCl胁迫下能够有效保持PSII原初光能转化效率和PSII潜在活性, 同时启动非辐射热能耗散以热的形式将过剩光能耗散掉, 以减轻对PSII供体侧、受体侧及电子传递链的伤害, 从而保护光合机构, 防止净光合速率过度下降。
生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(Samra, 1985; 廖祥儒等, 1996)。生长受到抑制以及生物量降低是盐胁迫对植物最为显著的影响(Zhu, 2002; Munns and Tester, 2008)。植物在逆境胁迫下会以生长缓慢和生物量积累降低等形式来适应不利的环境(Hernández et al., 1995; Chartzoulakis and Klapaki, 2000), 通过调整生物量在各器官的分配来维持植株在逆境条件下的生存(Grotkopp et al., 2002)。本实验发现, 在100 mmol·L-1 NaCl处理下, 各葡萄株系的新梢生长量减小, 节间数变少, 地上、地下、全株干重和鲜重等均受到不同程度的影响; 且大多数葡萄株系的生长指标明显降低, 这与前人的研究结果一致(高建明等, 2012; 赵俊香等, 2015)。李晓芬等(2008)采用多元统计分析方法对10个辣椒(Capsicum annuum)品种的耐盐性进行了分析, 依据具有显著性差异的12个性状指标, 综合评价了辣椒品种的耐盐性; 李丰先等(2013)对35个高粱品种的耐碱性进行了分析, 筛选出耐碱性快速鉴定指标。本实验利用主成分分析、相关性分析、隶属函数分析和聚类分析等多元分析方法对不同株系葡萄幼苗的生长相关指标进行了筛选与综合评价, 并将相互关联的12个单项指标的耐盐系数转换成3个相互独立的综合指标, 评价结果与各株系的盐害分级一致。
目前, 西北部葡萄主产区应用的砧木贝达不耐盐碱, 其在盐碱地种植表现出缺铁黄化现象; 而1103P植株在国外引进砧木中抗盐碱性较强, 在部分地区已开始取代贝达。但本实验发现, 杂交的A系列砧木的耐盐性均优于1103P植株, 具有更好的发展潜力, 尤其是筛选出的耐盐性较强的砧木株系A34、A35、A15和A17, 这为我国盐碱地区选用砧木提供了更多的选择。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | URLA sand culture experiment with Hoagland solution plus NaCl and NaCO was conducted to study the responses of sorghum seedlings to salt-alkaline stress. An assessment method for identifying the salt-alkaline tolerance of sorghum at seedling stage was established, and the salt-alkaline tolerance of 66 sorghum genotypes was evaluated. At the salt concentrations 8.0-12.5 g·L, there was a great difference in the salt-alkaline tolerance between tolerant genotype ‘TS-185’ and susceptive ‘Tx-622B’, suggesting that this range of salt concentrations was an appropriate one to evaluate the saltalkaline tolerance of sorghum at seedling stage. At the salt concentrations 10.0 and 12.5 g·L, there existed significant differences in the relative livability, relative fresh mass, and relative height among the 66 genotypes, indicating a great difference in the salt-alkaline tolerance among these genotypes. The genotype ‘Sanchisan’ was highly tolerant, 16 genotypes such as ‘MN-2735’ were tolerant, 32 genotypes such as ‘EARLY HONEY’ were mild tolerant, 16 genotypes such as ‘Tx-622B’ were susceptive, and genotype ‘MN-4588’ was highly susceptive to salt-alkaline stress. Most of the sorghum genotypes belonging to Sudangrasses possessed a high salt-alkaline tolerance, while the sorghum genotypes belonging to maintainer lines were in adverse. [本文引用: 1] |
| [2] | [本文引用: 1] |
| [3] | URL【目的】探寻适宜葡萄品种及砧木抗寒性研究的评价方法。【方法】以‘北冰红’‘左山一’‘贝达’及‘双红’4个葡萄品种为试材,分别测定了不同低温处理下各品种的相对电导率、丙二醛、游离脯氨酸、可溶性糖及可溶性蛋白含量等抗寒相关指标的变化,以电导法结合隶属函数法综合评价其抗寒性。【结果】随着处理温度的降低,4个品种相对电导率和丙二醛含量逐渐升高,呈“S”型曲线变化,而游离脯氨酸、可溶性糖和可溶性蛋白含量近似呈单峰曲线变化;指标平均隶属度与各理化指标相关性均较高;低温半致死温度几乎与综合评价结果一致,各品种抗寒性为:‘左山一’〉‘双红’〉‘贝达’〉‘北冰红’。【结论】抗寒相关指标平均隶属度变化曲线能够更全面可靠地反映出葡萄品种及砧木抗寒性差异的本质及其可承受的冻害极限温度。 [本文引用: 1] |
| [4] | [本文引用: 1] |
| [5] | URL快速荧光OJIP曲线中的Wk参数被广泛作为放氧复合体(OEC)即PSII供体侧伤害的重要指标。然而Wk这一参数的变化不仅受PSII供体侧伤害的影响,而且还会受到PSII受体侧伤害的影响。很多研究者在使用Wk这个参数时忽略了这个问题。为了阐明PSII供体侧和受体侧伤害程度对Wk影响的规律,我们通过不同温度和强光处理,获得不同PSII供体侧和受体侧伤害的黄瓜叶片,进而分析Wk变化与其供体侧和受体侧伤害程度之间的关系。研究结果表明:用高温黑暗处理黄瓜叶片后,其PSII供体侧受到严重的伤害,此时,Wk随着供体侧伤害程度的增加而显著增加,在这种情况下,Wk的增加直接反映了OEC的伤害程度;但是当用强光和高温共同处理黄瓜叶片后,造成PSII供体侧和受体侧同时伤害时,如果供体侧的伤害程度大于受体侧的伤害程度时,Wk仍然升高,然而,当受体侧的伤害程度大于供体测定伤害程度时,Wk不再显著升高。因此在植物叶片的供体侧和受体共同受到伤害时,要慎重使用Wk这一参数来判断OEC的伤害程度。本文对Wk在不同供体侧和受体侧伤害条件下的变化规律进行了详细的研究,并对如何正确使用Wk来反映逆境对植物叶片OEC的伤害程度进行了探讨,该研究结果可以对正确利用叶绿素快速荧光OJIP分析提供指导。 [本文引用: 1] |
| [6] | |
| [7] | DOI:10.3321/j.issn:1671-3877.2005.06.001URLJIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法。利用该方法可以获得有关光系统II的大量信息。文章介绍了快速叶绿素荧光诱导动力学曲线的定义、数据分析方法及相关参数的意义,并举例说明如何利用该方法分析不同环境条件对光合机构主要是PSII的供体侧、受体侧及PSII反应中心的影响。 [本文引用: 1] |
| [8] | DOI:10.3321/j.issn:0513-353X.2008.03.006URL根据NaCl溶液处理下辣椒萌芽期和幼苗期农艺性状指标,采用多元统计分析方法评价辣椒品种的耐盐性。结果表明:辣椒不同品种间相对发芽势、相对胚根长、相对发芽指数、盐害指数、相对株高等12个性状指标表现出显著性差异;采用因子分析将此12个性状指标用4个主因子表示,保留了原始变量95.35%的信息量;基于品种因子得分值进行聚类分析,10个辣椒品种分为3类:中椒4号、中椒8号、中椒7号、中椒12号为盐敏感品种,中椒5号、中椒11号为中度盐敏感品种,中椒6号、中椒13号、中椒16号、中椒10号为盐不敏感品种。因此,辣椒品种的耐盐性可通过测定萌芽期和幼苗期农艺性状指标,采用多元统计分析方法予以快速鉴定。 |
| [9] | URL0.4-1.0NaCl溶液培养的雷司令葡萄新梢,引起叶片失绿、萎蔫或组织坏死,1.0%NaCl导致叶柄蔫坏死。盐胁迫引起叶细胞膜透性增加,叶片产争光合速率和叶圆片暗呼吸明显降低。盐害症状随NaCl浓度的增加明显加重,并与叶龄有关。 [本文引用: 1] |
| [10] | DOI:10.3969/j.issn.1674-3466.1998.02.007URLEffects of NaCl on chl a fluorescence induction kinetics of leaves from two wheat species with different salt tolerance was investigated. Result show that the growth and chl content of wheat seedling were decreased by 150 mmol/L NaCl and under NaCl stress the potential activity and efficiency of primary conversion of light energy of PS Ⅱ in leaves of two wheat species decreased, light reaction was inhibited by NaCl. [本文引用: 1] |
| [11] | [本文引用: 1] |
| [12] | [本文引用: 1] |
| [13] | URL采用土壤干旱和室内PEG水分胁迫测定相结合的办法,研究有关生理指标与甘薯抗旱性关系。结果表明,水分胁迫下伴随着叶片相对含水量(RWC)的明显下降,丙二醛(MDA)含量显著增加,质膜相对透性(RPP)急剧上升,而过氧化氢酶(CAT)活性迅速下降;抗旱性强的品种叶片RFP和MDA的增加幅度显著低于不抗旱品种,而前者叶片RWC和CAT活性的下降速度显著慢于后者。并具有比后者较强的渗透调节能力,中等抗旱品种介乎两者之间。所选的五种生理指标与抗旱性的相关性次序是:RWCCATRPP可溶性糖蛋白氮,这5种指标的隶属函数加权平均值(D值)与抗旱系数间的相关系数达极显著水平(r=0.956、P0.01),其中 [本文引用: 1] |
| [14] | URL本研究以3个山葡萄品种双丰、左山二和双优一年生扦插苗为试材,以1个砧木品种贝达和1个鲜食葡萄品种金玫瑰一年生扦插苗为材料对照,采用盆栽试验方法,分析了NaCl胁迫下植株生长状况、生理生化指标以及叶绿素荧光参数的响应特点,以及部分葡萄品种Na+、K+含量及其分布和超显微结构的变化特点。并以21份山葡萄种质资源组培苗为试材,采用组培的试验方法,研究了NaCl胁迫对山葡萄组培苗株高、生根情况、侧芽萌发情况以及叶片受害情况的影响,并对山葡萄组培苗种质耐盐性进行了初步评价。结果如下: 1.山葡萄生长状况受害显著。随盐胁迫浓度的增加,各植株生长变化明显,葡萄新梢生长量减小,叶片含水量降低;随胁迫时间的延长植株受害程度逐渐加重。相同盐浓度下,金玫瑰的受害症状最轻,贝达受害症状较为明显,双丰、左山二和双优受害症状最为显著,重度胁迫下死亡。 2.山葡萄膜透性与丙二醛变化幅度较大。各植株细胞膜透性、MDA含量均随盐浓度的增大而升高。不同品种间变化幅度不同,随胁迫程度增加,双丰、左山二和双优变化幅度明显高于金玫瑰和贝达。 3.盐胁迫下超氧化物歧化酶和过氧化物酶活性变化趋势不同。植株SOD活性在盐胁迫期间呈先增后降趋势,金玫瑰、贝达和双优SOD活性比双丰和左山二增加更为迅速,增加幅度也较大。在胁迫前期,植株随胁迫浓度增加POD活性降低,金玫瑰的下降幅度低于贝达,双丰、左山二和双优下降幅度最大。 4.山葡萄游离脯氨酸含量较低。双优、金玫瑰和贝达脯氨酸含量均随盐浓度的升高而升高,在0.5%浓度下达到最大值;但双丰和左山二在0.1%浓度下达到最大值,后随浓度增加又逐渐下降,在0.5%浓度下达到最小值,且明显低于对照。 5.植株不同器官中Na+、K+含量变化趋势不同。植株各器官Na+含量均随NaCl浓度的增大而升高,但茎和根系的升高幅度明显高于叶片。叶片中K+含量随NaCl浓度的增大而升高,茎和根系的、K+含量变化趋势不稳定。 6.不同品种间叶绿素荧光参数变化幅度不同。随胁迫浓度的增加,可变荧光(Fv)、最大荧光(Fm)、可变荧光与最大荧光比(Fv/Fm)、可变荧光与初始荧光比(Fv/F0)、光化学淬灭系数(qP)均降低,而初始荧光(F0)升高,非光化学淬灭系数(NPQ)不同品种间反应不同。 7.山葡萄叶片细胞器受到不同程度损伤。盐胁迫下,叶片叶绿体类囊体片层发生紊乱、空泡化严重,内部大的淀粉粒溶解或消失;线粒体膜溶解,基质着色变浅;细胞核变形,核膜变得模糊不清。 8.山葡萄种质间耐盐性存在差异。21份山葡萄组培苗种质资源耐盐性初步评价结果如下:左山一、84007、九0一、84006和73069属于耐盐山葡萄类型;85013、73231、75086、086919和75023属于较耐盐山葡萄类型;双优、双丰、双红、通化三号、宝图202、087923和087918属于盐较敏感山葡萄类型;75029、73077、长白五号和77023属于盐敏感山葡萄类型。 [本文引用: 1] |
| [15] | DOI:10.3864/j.issn.0578-1752.2012.16.005URL【Objective】 The impacts of salt stress on the photosynthesis and chlorophyll fluorescence parameters of sorghum seedlings were studied for providing a foundation for sorghum cultivation, breeding and artificial regulation of salt stress.【Method】 Salt tolerant cultivar (Liaoza 15) and salt sensitive cultivar (Longza 11) were incubated in the nutrient solution at humidity of 60%, light/dark of 12 h/12 h, illumination of 134 μmol61m-261s-1 and 28℃/25℃ of day/night. NaCl was added into the solution at 3-leaf stage and NaCl concentration levels at 0, 50, 100, 150 and 200 mmol61L-1, respectively. The response of sorghum seedlings to NaCl stress was assessed by measuring the photosynthesis and chlorophyll fluorescence parameters of seedlings. 【Result】 Low NaCl concentration (50 mmol61L-1) increased the chlorophyll content, and high NaCl concentration (100-200 mmol61L-1) reduced the content substantially. Salt stress reduced Pn, Gs, Tr, Fm, Fv/Fo, Fv/Fm, Fv′/Fm′ and qP, and increased Fo and NPQ. Low NaCl concentration (50 mmol61L-1) reduced Ci, and high NaCl concentration (100-200 mmol61L-1) increased it. The adverse impact of salt stress on Liaoza 15 was less than on Longza 11. 【Conclusion】 The small reduction in net photosynthesis rate caused by 50 mmol61L-1 NaCl stress was considered to be a result of non-stomatal restriction; but increased stomatal restriction with increased NaCl concentration resulted in more severe reductions in photosynthesis. Under salt stress, salt tolerant cultivar could protect the photosynthetic organs more effectively than salt sentitive cultivar and thus improve the production of sorghum in salt affected areas. [本文引用: 2] |
| [16] | URL |
| [17] | DOI:10.3321/j.issn:0578-1752.2007.10.004URL: 【Objective】Waterlogging stress is one of the major problems in barley production especially in wetlands. Development of tolerant cultivars is the most cost efficient and environmentally safe way to overcome this problem. The objective of this study was to investigate suitable selection criteria for waterlogging tolerance in barley. 【Method】 165 DH lines derived from a barley cross(Yerong × Franklin) were analysed using principal component analysis, dynamic clustering and subordinate function based on the specimen correlation matrix of 12 traits, including plant height(PH), spike length (SL), the top 1st internode length(TFIL), spike per plant (SPP), grains per spike(GPS), grain weight per plant(GWPP), grains per plant(GPP), straw weight per plant(SWPP), weight of 1000 grains(WTG), number of green leaf (NGL) and Chlorophyll 1-2(Chl 1-2). 【Results】Three principal components and their functional equations, two for spike-grain factor and one for NGL factor, were used to identify lines for waterlogging tolerance, which led to the classification of 165 lines into 3 different levels of tolerance. 30, 93 and 42 lines were classified as waterlogging tolerant, medium tolerant and sensitive, respectively. 【Conclusion】The new selecting indices and comprehensive evaluation method provides a useful tool for further study on QTLs controlling waterlogging tolerance and selecting for waterlogging-tolerance in a breeding program. [本文引用: 1] |
| [18] | URL1985~1986年对5种果 树耐盐力的试验表明:不同树种的盐害指标不同。石榴、无花果在土壤含盐量0.4%时,盐害级别尚仅1~2级;杜梨、葡萄(白香蕉)在土壤含盐量0.2% 时,已出现1~2级盐害;毛桃在土壤含盐量0.1%时,盐害即达1~2级。不同树种不仅存在受害程度的不同,还存在受害敏感性的差异。 [本文引用: 1] |
| [19] | URL以东营野生大豆(Glycine soja Sieb.et Zucc.ZYD 03262)和山东栽培大豆(Glycine max (L.) Merr.山宁11号)为实验材料,通过研究2种大豆植株和离体叶片对不同浓度NaCl(0,100和200mmol/L)处理的响应,探讨2种大豆光合 机构对NaCl胁迫响应的差异和机理。结果表明:NaCl处理完整植株后,2种大豆植株叶片的光合速率(Pn)、PsⅡ最大光化学效率(Fv/Fm)、 PsⅡ实际光化学效率(φPsⅡ)和叶绿素含量都明显降低,而且生长也均受到抑制。但是,NaCl胁迫对栽培大豆各方而的抑制均显著大于野生大豆;野生大 豆叶片中的Na^+含量、Na^+/K^+值都显著低于栽培大豆,而野生大豆根中的Na^+含量却明显高于栽培大豆。当用100和 200mmoL/LNaCl处理2种大豆的离体叶片时,野生大豆的Fv/Fm、φPsⅡ、单位面积有活性反应中心的数目(RC/CS)和光化学性能指数 (川)的下降幅度却显著大于栽培大豆;叶片中的Na^+含量也显著高于栽培大豆。这些结果表明,实验所用的野生大豆的光合机构并不抗盐。但是,在盐胁迫条 件下,野生大豆植株却能够有效地避免过多Na^+进入叶片光合组织,以维持光合机构较高的光合活性,这是野生大豆比栽培大豆更抗盐的原因之一。 [本文引用: 1] |
| [20] | DOI:10.11707/j.1001-7488.20150907MagsciURL<p><strong>[目的]</strong> 研究不同种源沙枣在NaCl胁迫下的生长表现差异,以期为盐碱地沙枣引种提供指导依据,也为筛选优良耐盐碱沙枣种质提供理论基础。<strong>[方法]</strong> 以6个沙枣种源(阿拉尔、昌吉、金昌、银川、盐池和磴口)的实生苗为材料,通过温室盆栽试验,系统调查不同NaCl浓度(0,150和300 mmol·L<sup>-1</sup>)处理60天后,沙枣苗高生长量、地径、叶片参数、生物量积累和根冠比的变化规律,并利用隶属函数法结合权重综合评价各沙枣种源幼苗的耐盐能力。<strong>[结果]</strong> 在150 mmol·L<sup>-1</sup>NaCl胁迫条件下,除阿拉尔种源各生长指标与对照之间无明显差异外,其他5个沙枣种源的苗高生长量、地径、单株叶面积、叶片数和总生物积累量均显著减少,其中,银川种源降幅最大,苗高生长量、单株叶面积和生物积累量降幅分别达到33.32%,24.47%和26.55%。在300 mmol·L<sup>-1</sup>NaCl胁迫条件下,银川种源沙枣幼苗苗高生长量、单株叶面积和生物量积累分别仅为对照的47.53%,41.77%和51.86%,其他沙枣种源各生长指标降幅均小于银川种源。所有沙枣种源根冠比在150 mmol·L<sup>-1</sup>NaCl浓度下与对照之间差异不明显,在300 mmol·L<sup>-1</sup>NaCl浓度下,沙枣幼苗的根冠比均显著增加。利用隶属函数法结合权重综合评价6个种源沙枣幼苗的在2个NaCl浓度下的耐盐能力差异,结果显示,在150 mmol·L<sup>-1</sup>NaCl下,阿拉尔种源耐盐能力最强,其次为磴口,银川种源最差,而在300 mmol·L<sup>-1</sup>NaCl下,金昌种源沙枣幼苗耐盐能力最强,其次为阿拉尔,银川种源最差。综合2个NaCl浓度的综合评价值,6个沙枣种源的耐盐能力排序为:阿拉尔 >金昌 >磴口 >盐池 >昌吉 >银川。<strong>[结论]</strong> 在中、重度盐渍土进行沙枣引种栽培时,适宜选择阿拉尔种源沙枣幼苗,而不适宜选择种植银川种源沙枣幼苗。</p> [本文引用: 1] |
| [21] | URLGermination experiments and hydroponic experiments were carried out to identify the salt tolerance of 30 cotton varieties planted in Xinjiang. The results revealed that, in the 30 cotton varieties, proportion of the cotton varieties with high salt tolerance (at level Ⅰ) at their bud stage was 16.67% only, that with certain salt tolerance (at level Ⅱ) was 43.30%, and that sensitive to salt and with moderate salt tolerance was 40.00%. At seedling stage, there was no any cotton variety with high salt resistance, there were 8 cotton varieties with salt tolerance at level Ⅱ, and the proportion of cotton varieties with salt tolerance at level Ⅲ was 63.30%. At bud stage, there was an extremely significant positive correlation between the relative bud length and the relative root length, but a significant negative correlation between the relative bud length and the saltharmful index. At seedling stage, there was a significant positive correlation between the rootshoot ratio and the saltharmful index, and the salt tolerance at bud stage could be used to indirectly reflect the salt tolerance at cotton seedling stage. [本文引用: 1] |
| [22] | URL本文以苹果砧木平邑甜茶为实验材料,利用快速叶绿素荧光、延迟荧光及820nm光反射同步测 量技术,研究了干旱胁迫对平邑甜茶叶片光合机构的光系统I(PSI)、光系统II(PSII)以及整个光合电子传递链的伤害机制。实验结果表明:干旱胁迫 下平邑甜茶叶片的整个光合电子传递链都受到不同程度的影响。干旱2d,PSII反应中心捕获的光能用于还原QA的能力低于电子从QA向下游传递的能力,电 子从QA向下游传递给QB和PQ库等中间电子传递体的能力小于电子从QB和PQ库向PSI受体侧传递的能力,但干旱5d上述变化则相反,且更加显著。由此 可以得出,严重干旱胁迫下,沿着从PSII到PSI受体侧的光合电子传递链,电子传递的能力越来越低。此外,随着干旱胁迫,PSII和PSI反应中心色素 的降解速率快于天线色素,且PSII的捕光效率和PSI反应中心的含量逐渐降低。快速荧光、延迟荧光及820nm光反射同步测量结果互相印证了上述结论。 我们推测,这可能是平邑甜茶叶片对干旱的一种适应机制。 [本文引用: 1] |
| [23] | DOI:10.13930/j.cnki.cjea.141306URLSoil salinization and alkalinization frequently co-occur in nature. The interactive effects of salt and alkali stress on plants are more complex and serious, greatly limiting crop production. However, the comprehensive effect of salt and alkali stress on crops remains largely unclear. Screening and cultivating salt-tolerant plants are the most efficient way of exploiting saline-alkaline soils. Jerusalem artichoke (SO) and two alkali salts (NaHCO and NaCO) were mixed with mole ratio of 5︰23︰9︰5 to simulate typical saline-alkaline stress. The 16 Jerusalem artichoke accessions were cultivated in plastic pots in a controlled environment chamber. The half-strength Hoagland solution without exogenous saline-alkaline was used as the control. The half-strength Hoagland solution supplemented with 150 mmol·L 1 saline-alkaline mixture was as the treatment. The shoot fresh and dry weight, root fresh and dry weight, SOD activity, and MDA and proline contents were measured after 7 days treatment. The saline-alkaline resistance of different Jerusalem artichoke accessions was evaluated by the fuzzy mathematics method of membership function, principal component analysis (PCA) and clustering analysis (CA). PCA showed that shoot fresh weight, plant dry weight and proline content were the most significant factors, and recommended as the main indexes for identifying saline-alkaline tolerance of Jerusalem artichoke at seedling stage. The degrees of salt-tolerance of 16 Jerusalem artichoke accessions were different. The 16 Jerusalem artichoke accessions were sorted into four groups based on clustering analysis. Jerusalem artichoke accessions of ZH-2 and ZH-3 were strongly tolerant to saline-alkaline stress. Jerusalem artichoke accessions ZH-4, Taiyangdao and Qingyu001 had medium tolerance. Then the remaining 10 Jerusalem artichoke accessions (e.g., ZH-5, ZH-6 and ZH-10) had lower tolerance. Of all 16 Jerusalem artichoke accessions, ZH-9 was the most sensitive to saline-alkaline stress. [本文引用: 1] |
| [24] | [本文引用: 1] |
| [25] | [本文引用: 1] |
| [26] | DOI:10.1016/S0269-7491(01)00091-4PMID:11586773URLChromate (Cr) decreases the growth of Spirodela polyrhiza. The fronds lost their pigments. The O 2 evolution was also decreased. The Cr effect was found to be dose dependent. The toxic effects of Cr have further been studied on the photosynthetic activity of Spirodela polyrhiza by means of the chlorophyll a (Chl a) fluorescence transient O-J-I-P. The Chl a fluorescence transients were recorded in vivo with high time resolution and analyzed according to the JIP-test which can quantify the photosystem II behavior. Cr treated plants show a decrease in yield for primary photochemistry, Po. The performance index of PSII, PI ABS, which is the combination of the indexes of three independent parameters, (1) the total number of active reaction centers per absorption (RC/ABS), (2) yield of primary photochemistry ( Po) and (3) efficiency with which a trapped exciton can move an electron into the electron transport chain ( 0), decreased due to Cr treatment. Chromate sensitivity varies within plant populations. In summary Cr affects several targets of PSII. More specifically, the main targets of Cr, according to the JIP-test, can be listed as a decrease in the number of active reaction centers and damage to the oxygen-evolving complex. [本文引用: 1] |
| [27] | DOI:10.1016/S0304-4238(00)00151-5URLThe salt tolerance of two greenhouse bell-pepper hybrids ( Capsicum annuum L., ‘Sonar’ and ‘Lamuyo’) was studied during germination, seedling growth and vegetative growth in hydroponic culture. Salinity treatments were imposed by irrigating with half-strength Hoagland solution containing 0, 10, 25, 50, 100 and 150 mM/l of NaCl. Salinities up to 50 mM delayed germination but did not reduce the final germination percentage. It was reduced significantly at 100 and 150 mM NaCl in both hybrids. Seedling growth was reduced significantly with salinities higher than 10 mM NaCl. Plant growth parameters such as plant height, total leaf area and dry weight were significantly ( P=0.05) reduced at salinities higher than 25 mM NaCl in both hybrids. Roots had the highest Na + concentration compared to leaves, which increased with increasing salinity, while Cl 61 in leaves was much higher than Na +. Potassium concentration of plant tissues was less affected than Na and Cl by salinity increase. Total fruit yield in both hybrids was significantly reduced at salinities higher than 10 mM NaCl, the reduction being 95% at 150 mM NaCl. Both, fruit number per plant and fruit weight were reduced by the salinity. Our results suggest that during the growth stages studied, the hybrid ‘Lamuyo’ is more sensitive to salinity than the hybrid ‘Sonar’. [本文引用: 1] |
| [28] | DOI:10.1046/j.1469-8137.2000.00667.xURL [本文引用: 1] |
| [29] | [本文引用: 1] |
| [30] | DOI:10.1111/j.1399-3054.1997.tb03449.xURLHigher plants must dissipate absorbed light energy that exceeds the photosynthetic capacity to avoid molecular damage to the pigments and proteins that comprise the photosynthetic apparatus. Described in this minireview is a current view of the biochemical, biophysical and bioenergetic aspects of the primary photoprotective mechanism responsible for dissipating excess excitation energy as heat from photosystem II (PSII). The photoprotective heat dissipation is measured as nonphotochemical quenching (NPQ) of the PSII chlorophyll a (Chl a ) fluorescence. The NPQ mechanism is controlled by the trans-thylakoid membrane pH gradient ( pH) and the special xanthophyll cycle pigments. In the NPQ mechanism, the de-epoxidized endgroup moieties and the trans-thylakoid membrane orientations of antheraxanthin (A) and zeaxanthin ( Z ) strongly affect their interactions with protonated chlorophyll binding proteins (CPs) of the PSII inner antenna. The CP protonation sites and steps are influenced by proton domains sequestered within the proteo-lipid core of the thylakoid membrane. Xanthophyll cycle enrichment around the CPs may explain why changes in the peripheral PSII antenna size do not necessarily affect either the concentration of the xanthophyll cycle pigments on a per PSII unit basis or the NPQ mechanism. Recent time-resolved PSII Chi a fluorescence studies suggest the NPQ mechanism switches PSII units to an increased rate constant of heat dissipation in a series of steps that include xanthophyll de-epoxidation, CP-protonation and binding of the xanthophylls to the protonated CPs; the concerted process can be described with a simple two-step, pH-activation model. The xanthophyll cycle-dependent NPQ mechanism is profoundly influenced by temperatures suboptimal for photosynthesis via their effects on the trans-thylakoid membrane energy coupling system. Further, low temperature effects can be grouped into either short term (minutes to hours) or long term (days to seasonal) series of changes in the content and composition of the PSII pigment-proteins. This minireview concludes by briefly highlighting primary areas of future research interest regarding the NPQ mechanism. [本文引用: 1] |
| [31] | DOI:10.1086/338995PMID:18707424URLAbstract We studied 29 pine (Pinus) species to test the hypothesis that invasive species in disturbed habitats have distinct attributes. Seedling relative growth rate (RGR) and measures of invasiveness were positively associated across species as well as within phylogenetically independent contrasts. High RGR, small seed masses, and short generation times characterize pine species that are successful invaders in disturbed habitats. Discriminant analysis and logistic regression revealed that RGR was the most significant factor among these life-history traits separating invasive and noninvasive species. We also explored the causes of differences in RGR among invasive and noninvasive species. While net assimilation rate, leaf mass ratio, and specific leaf area (SLA) were all found to be contributing positively to RGR, SLA was found to be the main component responsible for differences in RGR between invasive and noninvasive pines. We investigated differences in SLA further by studying leaf anatomy, leaf density, and leaf thickness. We also evaluated relative leaf production rate as an important aspect of SLA. We proposed a hypothetical causal network of all relevant variables. [本文引用: 1] |
| [32] | DOI:10.1016/0168-9452(94)04047-8URLAbstract The possible involvement of activated oxygen species in the mechanism of damage by NaCl strees was studied in chloroplasts from leaves of two cultivars of pea (Pisum sativum L.) with differential sensitivity to NaCl. Intact organelles were purified by centrifugation in density-gradients of Percoll. In chloroplasts from tolerant plants, NaCl stress produced a significant increase of CuZn-SOD II and ascorbate peroxidase activities as well as in ascorbate content, while in those from sensitive plants NaCl produced increases in the H2O2 content and lipid peroxidation and no changes were observed in the enzymatic activities. Chlorophyll content significantly decreased in chloroplasts from sensitive plants and chloroplast integrity was lower in sensitive than in tolerant plants. Electron microscopy showed that the thylakoidal structure of chloroplast was notably disorganized in the NaCl-treated leaves. In purified chloroplasts, an increase in the number and size of plastoglobuli was produced by NaCl in chloroplasts from tolerant plants and to a lesser extent, in chloroplasts from sensitive plants. The relative starch content only decreased in chloroplasts from tolerant plants by NaCl-treatment. Results obtained suggest that in the cellular toxicity of NaCl in pea plants, superoxide- and H2O2-mediated oxidative damage in chloroplasts may play an important role. [本文引用: 1] |
| [33] | DOI:10.1023/A:1006408712416PMID:10949377URLThe potential role of photorespiration in the protection against salt stress was examined with transgenic rice plants. Oryza sativa L. cv. Kinuhikari was transformed with a chloroplastic glutamine synthetase (GS2) gene from rice. Each transgenic rice plant line showed a different accumulation level of GS2. A transgenic plant line, G39-2, which accumulated about 1.5-fold more GS2 than the control plant, had an increased photorespiration capacity. In another line, G241-12, GS2 was almost lost and photorespiration activity could not be detected. Fluorescence quenching analysis revealed that photorespiration could prevent the over-reduction of electron transport systems. When exposed to 150 mM NaCl for 2 weeks, the control rice plants completely lost photosystem II activity, but G39-2 plants retained more than 90% activity after the 2-week treatment, whereas G241-12 plants lost these activities within one week. In the presence of isonicotinic acid hydrazide, an inhibitor of photorespiration, G39-2 showed the same salt tolerance as the control plants. The intracellular contents of NH 4 + and Na + in the stressed plants correlated well with the levels of GS2. Thus, the enhancement of photorespiration conferred resistance to salt in rice plants. Preliminary results suggest chilling tolerance in the transformant. [本文引用: 1] |
| [34] | DOI:10.1093/jexbot/50.336.1199URLModulated chlorophyll fluorescence, rapid fluorescence induction kinetics and the polyphasic fluorescence transients (OJIP) were used to evaluate PSII photochemistry in wheat plants exposed to water stress and/or heat stress (25—45 °C). Water stress showed no effects on the maximal quantum yield of PSII photochemistry (Fv/Fm), the rapid fluorescence induction kinetics, and the polyphasic fluorescence transients in dark-adapted leaves, indicating that water stress had no effects on the primary photochemistry of PSII. However, in light-adapted leaves, water stress reduced the efficiency of excitation energycapture by open PSII reaction centres (F′v/F′m) and the quantum yield of PSII electron transport (01PSII), increased the non-photochemical quenching (qN) and showed no effects on the photochemical quenching (qP). This suggests that water stress modified the PSII photochemistry in the light-adapted leaves and such modifications may be a mechanism to down-regulate the photosynthetic electron transport to match a decreased CO2 assimilation. In addition, water stress also modified the responses of PSII to heat stress. When temperature was above 35 °C, thermostability of PSII was strongly enhanced in water-stressed leaves, which was reflected in a less decrease in Fv/Fm, qP, F′v/F′m, and 01PSII in water-stressed leaves than in well-watered leaves. There were no significant variations in the above fluorescence parameters between moderately and severely water-stressed plants, indicating that the moderate water stress treatment caused the same effects on thermostability of PSII as the severe treatment. It was found that increased thermostability of PSII may be associated with an improvement of resistance of the O2-evolving complex and the reaction centres in water-stressed plants to high temperature. [本文引用: 1] |
| [35] | DOI:10.1146/annurev.arplant.59.032607.092911URL [本文引用: 1] |
| [36] | [本文引用: 1] |
| [37] | DOI:10.1023/A:1005896029778URL [本文引用: 1] |
| [38] | [本文引用: 1] |
| [39] | DOI:10.1071/pp9810359URLRooted cuttings of grapevines (Vitis vinifera L. cv. Sultana; syn. Thompson Seedless) were grown under glasshouse conditions and supplied with dilute nutrient solution containing either 0 or 90 mM of added NaCl. Growth and photosynthetic response to salt treatment and subsequent recovery were followed over 80 days. Shoot growth and photosynthesis were reduced by salt treatment. At relatively low concentrations of leaf chloride (< c. 150 mM, on a tissue water basis), photosynthetic reduction was largely due to increased stomatal resistance. Internal disturbances were involved at higher leaf Cl- concentrations (> c. 150 mM) and included an apparent reduction in photochemical efficiency and a faster rate of photorespiration. Levels of fraction I protein, and specific activity of ribulose-1,5-bisphosphate carboxylase measured in vitro, were not reduced by salt treatment. Vines showed remarkable adaptation to salinity insofar as leaves maintained positive turgor despite leaf Cl- concentrations exceeding 300 mM, implying osmotic adjustment. Cessation of salt treatment led to an immediate decrease in leaf Cl-, a promotion of shoot growth and a progressive recovery in photosynthesis accompanied by a marked but not necessarily concurrent reduction in both stomatal and internal resistances. Leaves tolerated Cl- levels up to 200 mM (under glasshouse conditions) without sustaining permanent reduction in photosynthetic activity. New shoots formed subsequent to stress relief are not a prerequisite for Cl- retranslocation from mature leaves as decapitation at the time of stress relief did not prevent attenuation of leaf Cl- or recovery in photosynthesis. |
| [40] | URLThe age related effect of drought stress on fluorescence parameters and photosynthetic pigments in winter wheat leaves was studied. In all leaves examined, drought induced declines in PSⅡ photochemical efficiency measured as midday values of Fv/Fm were almost completely reversible overnight, such that the predawn Fv/Fm remained largely unchanged during the initial 6~8 days of stress. The violaxanthin (V) deepoxidation into zeaxanthin (Z) increased in response to drought. When the increase of Z reached a plateau level, photoinhibitory damage, as indicated by a net increase in Fo and sustained decline in Fv/Fm, became evident. Compared with older leaves, younger leaves showed a higher capacity for Z synthesis and also showed a lower sensitivity to photodamage caused by a combination of drought and high sunlight stresses. This observation suggests an important role of Z in the protection of photosynthetic apparatus against photoinhibitory damage. We speculate that during drought stress, older leaves senesced irreversibly because they may not be able to increase the Z synthesis to cope with the excessive light stress. [本文引用: 1] |
| [43] | DOI:10.1146/annurev.arplant.53.091401.143329URL [本文引用: 1] |
| [41] | DOI:10.1104/pp.010919PMID:11891263URLWinter wheat (Triticum aestivum L. cv Norin No. 61) was grown at 25°C until the third leaves reached about 10 cm in length and then at 15°C, 25°C, or 35°C until full development of the third leaves (about 1 week at 25°C, but 2-3 weeks at 15°C or 35°C). In the leaves developed at 15°C, 25°C, and 35°C, the optimum temperature for CO2-saturated photosynthesis was 15°C to 20°C, 25°C to 30°C, and 35°C, respectively. The photosystem II (PS II) electron transport, determined either polarographically with isolated thylakoids or by measuring the modulated chlorophyll a fluorescence in leaves, also showed the maximum rate near the temperature at which the leaves had developed. Maximum rates of CO2-saturated photosynthesis and PS II electron transport determined at respective optimum temperatures were the highest in the leaves developed at 25°C and lowest in the leaves developed at 35°C. So were the levels of chlorophyll, photosystem I and PS II, whereas the level of Rubisco decreased with increasing temperature at which the leaves had developed. Kinetic analyses of chlorophyll a fluorescence changes and P700 reduction showed that the temperature dependence of electron transport at the plastoquinone and water-oxidation sites was modulated by the temperature at which the leaves had developed. These results indicate that the major factor that contributes to thermal acclimation of photosynthesis in winter wheat is the plastic response of PS II electron transport to environmental temperature. [本文引用: 1] |
| [42] | DOI:10.1016/j.plantsci.2006.04.010PMID:22980209URLPhotosynthesis and its tolerance to photoinhibition, high temperature and drought stress of flag leaves were investigated in a wheat (Triticum aestivum L.) hybridization line (1-12) and its parents (Jing-411 and Xiaoyan-54). From the beginning of flowering to the 10th day, light-saturated CO2 assimilation rate (Pmax) showed no significant decrease and Pmax of 1-12 was comparable to that of its parents. From the 20th day, Pmax decreased significantly and this decrease was much less in 1-12 than in its parents, whereas no decrease in chlorophyll content was observed in 1-12 and its parents on the 20th day, indicating that photo-oxidative damage occurred in 1-12 and its parents but 1-12 is more resistant to photo-oxidative damage than its parents. To further characterize photo-oxidative damage, tolerance to photoinhibition, high temperature and drought stress was compared in 1-12 and its parents. When exposed to high light (1400μmolm612s611), the maximal efficiency of PSII photochemistry (Fv/Fm) decreased significantly with increasing exposure time and such a decrease was much less in 1-12 than in its parents. When exposed to higher temperatures (30–45°C) for 15min, Fv/Fm started to decrease at 42°C in 1-12 and its parents. The greatest decrease in Fv/Fm was observed in Jing-411. 1-12 and Xiaoyan-54 showed a comparable decrease in Fv/Fm. Similar results were also observed in the actual PSII efficiency (ΦPSII), photochemical quenching (qP) and non-photochemical quenching (qN). During exposure of detached leaves to air under room temperature conditions, relative water content decreased with increasing exposure time. However, such a decrease was greatest in Jing-411 followed by Xiaoyan-54, and 1-12. There were neither changes in Fv/Fm nor qN during water loss in the line and its parents. However, there was a decrease in ΦPSII and qP and greatest decrease was observed in Jing-411 followed by Xiaoyan-54, and 1-12. Also, the decrease in Pmax was greatest in Jing-411, followed by Xiaoyan-54, and 1-12 during water loss. In addition, the activities of ribulose-1,5-bisphosphate carboxylase, phosphoenolpyruvate carboxylase, pyruvate phosphate dikinase, NADP-malate dehydrogenase and NADP-malate enzyme were significantly higher in 1-12 than in its parents. The results in this study suggest that high resistance to photo-oxidative damage of the flag leaves in 1-12 may be the physiological basis for its high yield when grown in north China. Our results also suggest that parents can be selected for improved biochemical and physiological traits and crossed to high yielding agronomically elite materials which can be selected for higher performance in yield. [本文引用: 1] |
高粱种质材料幼苗期耐盐碱性评价
1
2012
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
1999
... 因为葡萄根瘤蚜的侵染, 国外很早就开始进行抗性砧木育种, 其中选育的砧木品种SO4和101-1对根瘤蚜免疫且抗寒性较强, 但耐盐性一般(
葡萄品种及砧木抗寒性评价方法研究
1
2015
... 因为葡萄根瘤蚜的侵染, 国外很早就开始进行抗性砧木育种, 其中选育的砧木品种SO4和101-1对根瘤蚜免疫且抗寒性较强, 但耐盐性一般(
保护地生产条件下的土壤退化问题及其防治对策
1
2004
... 我国葡萄的主产区分布在西北、华北、东北以及沿海等地, 然而受气候变化及不合理耕作等因素的影响, 上述地区的土壤盐渍化程度不断扩大(
高温、强光下黄瓜叶片PSII供体侧和受体侧的伤害程度与快速荧光参数Wk变化的关系
1
2015
... K点荧光的相对变化值(WK)表示PSII供体侧放氧复合体(oxygen evolving complex, OEC)的伤害程度(
高粱品种萌发期耐碱性筛选与综合鉴定
2013
快速叶绿素荧光诱导动力学分析在光合作用研究中的应用
1
2005
... 曲线(OJIP) (
多元统计分析方法在辣椒品种耐盐性评价中的应用
2008
盐胁迫对葡萄离体新梢叶片的伤害作用
1
1996
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
盐胁迫对不同抗盐性小麦叶片荧光诱导动力学的影响
1
1998
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
高温胁迫对‘赤霞珠’葡萄光合作用的影响
1
2010
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
遮阴和环剥对荔枝枝梢生长和光合生理的影响
1
2013
... 最大光化学效率(Fv/Fm)是反映光抑制程度的良好指标(
甘薯品种抗旱性生理指标及其综合评价初探
1
1996
... Walker等(1981)发现葡萄光合速率的显著降低与高盐胁迫密切相关; Troncoso等(1999)以生长量为指标对葡萄砧木进行了耐盐性分类.鉴于耐盐性较为复杂, 单一指标不能全面反映特定品种或基因型的遗传本质; 且选用的耐盐性指标不同, 对植物的耐盐性评价结果也不一致.因此, 需要对耐盐性评价指标进行筛选, 找出与耐盐性评价相关性较大的指标.在作物抗逆性筛选和鉴定方面, 有****提出了总级别法和隶属函数法等(
山葡萄种质资源耐盐性评价研究
1
2010
... 因为葡萄根瘤蚜的侵染, 国外很早就开始进行抗性砧木育种, 其中选育的砧木品种SO4和101-1对根瘤蚜免疫且抗寒性较强, 但耐盐性一般(
盐胁迫对高粱幼苗光合作用和荧光特性的影响
2
2012
... 盐胁迫影响植物的生长和代谢等许多方面, 但对光合作用的影响尤为突出.王邦锡等(1992)研究表明, 叶片叶绿体结构受到破坏是影响植物光合作用正常进行的重要原因.盐胁迫可导致植物叶片叶绿素含量降低, 这主要是由于高盐使得植物叶片中叶绿素与叶绿素蛋白结合变得松弛, 叶绿素酶活性增强, 从而促进叶绿素降解所致(
... 的消耗能力降低(
水分胁迫导致小麦叶片光合作用下降的非气孔因素
1992
大麦耐湿性鉴定指标和评价方法研究
1
2007
... Walker等(1981)发现葡萄光合速率的显著降低与高盐胁迫密切相关; Troncoso等(1999)以生长量为指标对葡萄砧木进行了耐盐性分类.鉴于耐盐性较为复杂, 单一指标不能全面反映特定品种或基因型的遗传本质; 且选用的耐盐性指标不同, 对植物的耐盐性评价结果也不一致.因此, 需要对耐盐性评价指标进行筛选, 找出与耐盐性评价相关性较大的指标.在作物抗逆性筛选和鉴定方面, 有****提出了总级别法和隶属函数法等(
五种果树耐盐力试验初报
1
1990
... 用100 mmol·L-1 NaCl对试材进行胁迫处理20天后, 观察各葡萄株系的表型及盐害情况.根据盐害轻重分为5个等级(
野生大豆和栽培大豆光合机构对NaCl胁迫的不同响应
1
2011
... 盐胁迫影响植物的生长和代谢等许多方面, 但对光合作用的影响尤为突出.王邦锡等(1992)研究表明, 叶片叶绿体结构受到破坏是影响植物光合作用正常进行的重要原因.盐胁迫可导致植物叶片叶绿素含量降低, 这主要是由于高盐使得植物叶片中叶绿素与叶绿素蛋白结合变得松弛, 叶绿素酶活性增强, 从而促进叶绿素降解所致(
NaCl胁迫下不同种源沙枣的生长表现差异
1
2015
... 为了更加合理地评价各葡萄株系对NaCl胁迫的敏感性, 根据实验所得数据, 利用公式(3)求出各葡萄株系各单项指标的耐盐系数.从
新疆主要棉花品种耐盐性筛选与鉴定
1
2013
... Walker等(1981)发现葡萄光合速率的显著降低与高盐胁迫密切相关; Troncoso等(1999)以生长量为指标对葡萄砧木进行了耐盐性分类.鉴于耐盐性较为复杂, 单一指标不能全面反映特定品种或基因型的遗传本质; 且选用的耐盐性指标不同, 对植物的耐盐性评价结果也不一致.因此, 需要对耐盐性评价指标进行筛选, 找出与耐盐性评价相关性较大的指标.在作物抗逆性筛选和鉴定方面, 有****提出了总级别法和隶属函数法等(
利用快速荧光、延迟荧光和820 nm光反射同步测量技术探讨干旱对平邑甜茶叶片光合机构的伤害机制
1
2013
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
16份菊芋种质苗期耐盐碱性筛选与综合鉴定
1
2015
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
2002
... 采用
NaCl胁迫对盐芥和拟南芥光合作用的影响
1
2007
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2001
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2000
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
2000
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
1991
... 我国葡萄的主产区分布在西北、华北、东北以及沿海等地, 然而受气候变化及不合理耕作等因素的影响, 上述地区的土壤盐渍化程度不断扩大(
1
1997
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2002
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
1995
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
2000
... 盐胁迫影响植物的生长和代谢等许多方面, 但对光合作用的影响尤为突出.王邦锡等(1992)研究表明, 叶片叶绿体结构受到破坏是影响植物光合作用正常进行的重要原因.盐胁迫可导致植物叶片叶绿素含量降低, 这主要是由于高盐使得植物叶片中叶绿素与叶绿素蛋白结合变得松弛, 叶绿素酶活性增强, 从而促进叶绿素降解所致(
1
1999
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2008
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
1985
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
1997
... K点荧光的相对变化值(WK)表示PSII供体侧放氧复合体(oxygen evolving complex, OEC)的伤害程度(
1
1999
... 因为葡萄根瘤蚜的侵染, 国外很早就开始进行抗性砧木育种, 其中选育的砧木品种SO4和101-1对根瘤蚜免疫且抗寒性较强, 但耐盐性一般(
1981
1
1999
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2002
... 生长特性是葡萄对盐胁迫的综合反应, 也是评价葡萄耐盐性的最优指标(
1
2002
... 植物对光能的吸收、传递和转化必然会受到光合碳同化速率的影响, 最明显的表现是光化学活性下降(
1
2006
... 盐胁迫影响植物的生长和代谢等许多方面, 但对光合作用的影响尤为突出.王邦锡等(1992)研究表明, 叶片叶绿体结构受到破坏是影响植物光合作用正常进行的重要原因.盐胁迫可导致植物叶片叶绿素含量降低, 这主要是由于高盐使得植物叶片中叶绿素与叶绿素蛋白结合变得松弛, 叶绿素酶活性增强, 从而促进叶绿素降解所致(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}